
Abstract
Background
Restoration practices usually emphasize on the structural part of the biodiversity; also, most studies have focused on plants and very few have been conducted on arthropods and its function after restoration. The Pedregal de San Angel Ecological Reserve (PSAER) is a protected area immersed in Mexico City and it has been drastically affected by different anthropogenic disturbances. The aim of this study was to compare the relative diversity, richness, and abundance of species level identification, but also the composition through an analysis of ordination of taxonomic (species, family, and order level) and functional (trophic guild) traits of arthropods in three sites subjected to ecological restoration within the PSAER. Restored sites were also compared to conserved and disturbed sites, to evaluate whether restoration efforts are effective at the reserve.
Methods
Arthropods were sampled using pan traps during September 2013 in 11 sites (three restored, four conserved and four disturbed) inside the PSAER. All sampled species were taxonomically identified at species of morphospecies (inside a family) and assigned to a trophic guild. Differences in diversity, richness and abundance were evaluated through effective number of species, comparisons of Chao’s1 estimated richness and a non-parametric Kruskal–Wallis test, respectively. Both taxonomic and trophic guild composition were evaluated using a multivariate analysis and a post hoc test.
Results
We found some differences in richness, abundance, and diversity between sites, but not a clear pattern of differentiation between restored to disturbed sites. The NMDS showed differences at species and order level, and with trophic guilds, among site types. Families were not useful to differentiate types of sites. Regarding guilds, predators were more abundant in conserved sites, while phytophagous insects were more abundant in disturbed sites.
Conclusions
Species and order level were useful to identify differences in communities of arthropods in sites with different management. The trophic guild approach provides information about the functional state of the restored sites. Nevertheless, our quick evaluation shows that restoration efforts at PSAER have not been successful in differentiate restored to disturbed sites yet.
Similar content being viewed by others


Background
Ecological restoration arises as an activity to mitigate the negative effects of disturbances on ecosystems, and its main purpose is to initiate or accelerate the process of recovery, and return to a pre-disturbance state [1]. To know whether the developed actions are successful to recover disturbed sites, so they resemble conserved sites, it is critical to monitor changes on the community structure of the taxa inhabiting such ecosystems, which also implies functional structure.
Most of the research on ecological restoration has been conducted in plant communities [2, 3] mainly because the natural succession of vegetation will determine the result of the restoration for other taxa [4]. Nevertheless, according to the Society for Ecological Restoration (SER) [1], the main goal of the ecological restoration is to recover not only the structure of the ecosystem but also its function. For this reason, it is critical to evaluate other taxonomic groups such as arthropods in order to integrate them as indicators of restoration programs, and to evaluate the effect of restoration from different ecosystem perspectives [5, 6].
Regarding to this, arthropods are the most diverse group of organisms on Earth, composing nearly 85% of all described species [7]. They can be classified depending on their guild (herbivores, predators or saprophagous) and the way of life and functions that they provide to the ecosystem (e.g. pollinators, pest controllers, scavengers, preys or biomass recyclers) [8, 9]. The great richness and diversity, variability in size, fast growth, high dispersion rates, sensitivity to environmental changes and short lifespan make arthropods an ideal study system to evaluate the effectiveness of the restoration programs in short periods of time [10,11,12]. Some factors that can determine the arthropod community are: i) changes in the quality, abundance and availability of resources [13,14,15], ii) spatial variation (horizontal and vertical) of plant communities [15], iii) origin (native or exotic) of plant species [16, 17], iv) habitat heterogeneity, v) degree of exposure to natural enemies [18, 19], and vi) disturbances [15, 20].
Although arthropods have been used largely in monitoring of restoration efforts [18], it is difficult to obtain species identification quickly, which commonly lead to the use of morphospecies [19, 21]. The use of coarse taxonomical identifications has been tested to evaluate the response of invertebrate communities to changes in habitats [22, 23], proving that the effectiveness of different levels depends -among other things- on physical and vegetal structures. Thus, an evaluation of the use of different taxonomical levels in a specific area could be useful to design a continuous monitoring in restoration programs.
In cities, overpopulation has induced a massive reduction of natural land uses, which in turn has led to habitat fragmentation, loss of biodiversity and a drastic reduction of the quality and quantity of the ecosystem services they provide [12, 24]. The Pedregal de San Angel Ecological Reserve (PSAER) is a protected area (since 1983) embedded in the south part of Mexico City [25], which has been drastically affected by different anthropogenic disturbances such as garbage accumulation, the reduction of the natural habitat by buildings, unauthorized people presence, accidental fires, extraction of native species and introduction of exotic species (both animals and plants), the extraction of basalt rock, as well as light and noise pollution [26]. For these reasons, several restoration efforts have been carried out to reduce the disturbances and to accelerate the recovery of such sites [27,28,29].
The aim of this study was to evaluate the effectiveness of ecological restoration on three sites within the PSAER. To do so, we compared arthropod richness, abundance, and diversity with conserved and disturbed sites. Also, we compared taxonomical (at different levels) and guild composition between types of sites. Although our sampling method is biased to flying and epigeous insects, this study represents the some of the firsts attempts to use entire arthropod communities to evaluate the restoration progress of three different sites within the PSAER; particularly, we addressed the following questions: 1) are there differences in richness, abundance, diversity and taxonomical composition at species level of the arthropod communities in conserved, restored and disturbed sites?, 2) the use of taxonomic orders or families leads to different results compared to species level? 3) is there a specific affinity of the three main trophic guilds (predators, herbivores and saprophagous) to the management categories used? and 4) are the restoration actions being effective? We expected a higher richness, diversity and abundance in conserved sites than in disturbed ones, according to declining arthropod diversity with plant species loss hypothesis [14], with the restored sites closer to conserved ones [30, 31]. We expected equivalent information provided by different taxonomic levels [22, 23, 32]. Also, we expected a better resolution of the differences between sites using trophic guilds, as trophic structure is a proxy for ecosystem function [33]. With all this information, we expect to be able to determine if restoration efforts in the PSAER have been good enough to differentiate restored to disturbed sites.
Materials and methods
Site of study
The PSAER is an urban reserve inside Mexico City. Geologically, the area is result of the Xitle volcano eruption, ~ 1600 years ago [34]. It is located to the southwest of Mexico City (19° 20´ 11´´ N, 99° 10´15´´ W; 2,292– 2,365 m.a.s.l). The climate is temperate with a dry winter and a medium annual temperature of 15.6 °C and a pluvial precipitation of 833 mm [35]. The region shows two distinctive seasons: a dry one and a wet one, which promote a xeric scrub community dominated by Pyttocaulom praecox (Cav.) Rob & Brettell (Asteraceae) [36, 37]. The reserve contains three core zones located to the west (ZNP), east (ZNO) and southeast (ZNSO) and 13 buffer zones (from A1 to A13).
Although we provide a brief description of conserved, restored, and disturbed sites below, a detailed characterization is shown in Table 1.
Conserved sites in the PSAER display high spatial heterogeneity caused by the cooling of the lava flow; this geography provide different conditions of soil, moisture, temperature and light exposition [25], as well as caves, promontories, rock fissures and hollows. On the other hand, disturbed sites are characterized by an open vegetation and a relative flatness, caused by the introduction of soil used for gardening. In disturbed areas it is common to find trash, rubble, and gardening wastes.
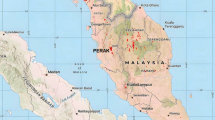
Inside the reserve, three sites have been subjected to ecological restoration: 1) the buffer zone 11 (hereafter called R1), a portion of the buffer zone 8 (R2) and a site located in the southeast core zone (R3). In addition to these three sites subjected to ecological restoration, we randomly selected 4 conserved (C1, C2, C3, C4) and 4 disturbed areas (D1, D2, D3, D4) (Fig. 1) based on our knowledge of the reserve. Characterization of each site is provided in Table 1.

A Location of the Pedregal de San Angel, in Mexico City. B Original extension of the Pedregal and location of the PSAER (2020), C Sampling sites in the PSAER, noted as white polygons
Arthropod sampling
The sampling was performed in September of 2013, corresponding to the wet season where more density of arthropods has been observed in the reserve [39, 40]. A line of eight pan traps with a 10 m distance between them, were placed in all 11 sites (conserved, restored, and disturbed) (lines of 80 m total). Lines were placed 10 m away from the border of the area. Traps consisted in plastic bowls with a diameter of 17.5 cm in which 175 ml of a 5% solution of shampoo with honey scent was poured. To catch a higher variability of arthropods we used four colors [41,42,43]: solid orange, solid yellow, solid white and fluorescent yellow. Thus, we used two bowls of each color in each site. The traps operated for 540 min, from 8:00 to 17:00 h for one day. Here it is important to point out that although there are other methods which are commonly used to monitor insect diversity [44], such as pitfall traps or sweep netting, the use of pitfall traps was logistically impossible in our case because the dept of the soil is very low and most of the land is still basaltic rock. One of the advantages to use pan traps was that all the traps were working simultaneously.
The collected material was conserved in alcohol at 70%. Each specimen was identified at stereomicroscope to the finest taxonomic level possible using dichotomous keys, catalogues, or previous records at species level [45,46,47,48,49,50,51,52,53]. All the specimens were identified at least at the family level, and were assigned a trophic guild (predator, phytophagous or saprophagous) according to the description of the family provided in [34] which corresponds to most of the species in each family. When the taxonomic identification was not possible, we used the morphospecies criteria based on sightly differences of anatomical structures. Neither color nor size was considered to differentiate a morphospecies. Unidentifiable larvae were not considered in our analysis. Although it may be possible that some members of the family have different trophic habits, our lack of a species identification made difficult to use a better guild resolution.
Data analysis
To perform the diversity and richness analysis, we grouped all the replicates (sites) for conserved and for disturbed sites according to their category. The restored sites were considered individually for each analysis to gain a better knowledge of their restoration success.
To test for differences in diversity at species level identifications, we calculated the effective number of recognizable taxonomic units (RTUs) since we have a pool of species and morphospecies. To do so, we used the algorithm to calculate the effective number of species, which represents the number of species with the same abundance in a virtual community that equals the diversity obtained by any traditional index, so that comparisons between communities are more intuitive [54, 55]. In this case, we used the exponential of the values of the centered Confidence Interval (CI) at 95% of Shannon–Weaver entropy index, obtained with a bootstrap of 9999 replicates using PAST4.04 [56]. By doing this, we are not favoring common or rare RTUs [54]. If an overlap in the CI is detected, the evaluated parameters are considered as equal [55].
To compare richness, we calculated the confidence interval at 95% of Chao’s1 index (bias corrected) estimated richness sensu Gotelli and Colwell [57] for site, using centered bootstrapping (9999 replicates) with PAST4.04. This method offers an estimated approach of the existing species (RTUs, in our case), still when they were not sampled. It considers the presence of organisms found in only one site (singletons) and in two sites (doubletons) through the next expression:
where:
S = richness, F1 = number of singletons, and F2 = number of doubletons.
In our case, singletons were considered as RTUs that appears either in just one of the restored sites (independently), or in just one time in the grouped conserved or disturbed sites. The same sites criterion was used for doubletons, as RTUs that appeared twice.
To test for differences in abundance between treatments and because our data did not meet the assumptions of a normal distribution, we perform a non-parametrical Kruskal–Wallis test using the kruskal.test function in the package stats in R 4.0.3 [58]. We used the type of site as a fixed factor. In this case, besides the grouping of conserved and disturbed sites, we also grouped the three restored sites as one group to facilitate comparisons of this data with ANOSIM (see below). Since there were differences between type of sites, we used the pairwise.wilcox.test function (also contained in stats) to detect differences between pairs.
To evaluate the sites ordination in relation with the abundances and the RTUs composition, we performed a non-metric multi-dimensional scaling (NmMDS) using Bray–Curtis distance since it considers non only presence-absence data, but abundance, trough PAST4.04, using an 11 (sites) × 159 (morphoespecies) matrix. The same analysis was performed for orders using a 11 (sites) × 11 (orders) matrix; for families using an 11 (sites) × 78 (families) matrix and for trophic guilds using an 11 (sites) × 3 (trophic guilds: predators, phytophagous or saprophagous) matrix. In NmMDS, there is a value called stress, which is a measure of goodness of fit. Here, we consider a rule of thumb which states that values under 0.2 are considered adequate, i.e. the lower the value, the better fit. In the latter, we perform a x2 test to identify if trophic guilds were over (or under) represented in each type of site. Also, we performed an analysis of similarities (ANOSIM) to detect differences among categories, both for taxonomical and trophic guilds arrangements. This analysis compares the values of any method used to calculate distances, inter and intra factor [59] through the following expression:
where \({\overline{r} }_{b}\) is the mean of the distance range between factor; \({\overline{r} }_{w}\) is the mean of the distance range within factor; and M = \(\frac{n\left(n-1\right)}{2}\), with n as the sample number. If P < 0.05, the program runs a pairwise test to detect the differences among groups.
Results
We found a total of 857 organisms of 159 RTUs coming from 78 families, 10 orders and three subphyla (Supplementary table 1). Nevertheless, we worked with 11 order-level groups, as we consider Heteroptera and Homoptera as different orders because several heteropterans may be predators, hematophagous, polliniphagous, spermophagous and even folivorous, and homopterans (Sternorrhyncha and Auchenorrhyncha) are only sap feeders. This way, we were able to get a better knowledge of the diverse functional groups in the PSAER. The distribution of the orders among the sites is shown in Table 2.
Restored sites were different to conserved or disturbed sites in terms of diversity, which showed an overlapped CI, going from 31 to 45 effective number of RTUs in conserved sites, and from 36 to 48 in disturbed sites (Fig. 2). Site R1 was different to the other restored sites with a lower CI of two to three effective numbers of RTUs. R2 and R3 showed an overlapped CI, going from 17 to 28 and from 12 to 24, respectively (P < 0.05). In terms of richness, R1 showed less estimated RTUs with 7 to 30. All other conserved, disturbed, and restored sites showed at least 99 to 199 estimated RTUs, and there were no significant differences between them (Fig. 3).

95% Confidence Intervals of arthropod diversity in conserved, disturbed, and restored sites in the PSAER, based on 857 arthropods in total. Conserved and disturbed sites were grouped respectively to facilitate the interpretation. Captures in all conserved and disturbed sites were pooled and treated as just one site, in each case. Dots represent the effective number of RTUs obtained through the Shannon–Weaver entropy index calculated from our real data, which is right sided of the bar. Lower and upper bar represents minimum, and maximum obtained through a bootstrap resampling with 9999 replicates. Different letters represent different groups due the lack of an overlapped area

95% Confidence Intervals of arthropod richness obtained through Chao1 species estimation in conserved, disturbed, and restored sites in the PSAER, based on 857 arthropods in total. Conserved and disturbed sites were grouped to facilitate the interpretation. Lower and upper bar represents minimum, and maximum obtained through a bootstrap resampling with 9999 replicates. The spots represent real richness obtained in our sampling, which is right sided of the bar. Different letters represent different groups due the lack of an overlapped area
The Kruskall-Wallis test showed differences in abundance (χ2 = 11.97, g.l. = 2, P = 0.002) between types of sites. The pairwise test indicated differences between conserved and disturbed sites (P = 0.018) and between restored and disturbed sites (P = 0.003). No differences were found between restored and conserved sites (P = 0.605) (Fig. 4).

Pairwise comparison of arthropod abundance in conserved, restored, and restored sites in the PSAER, based on 857 arthropods in total. Conserved, restored, and disturbed sites were grouped respectively to facilitate the interpretation with ANOSIM differences, which are based in Bray–Curtis distances. The points represent the real mean abundance obtained in our sampling, which is to the right of the bar. Maximum and minimum were calculated through standard error. Different letters represent different groups
In the NmMDS for abundance and composition of RTUs the stress value was 0.1925, and the analysis aggregated two distinctive groups conformed by conserved and disturbed sites. The restored sites resulted scattered around the disturbed ones (Fig. 5).

Non-metric Multidimensional Scaling showing the ordination of the 11 study sites in the PSAER, based on RTUs. Dots represent conserved sites; triangles represent disturbed sites and squares represent restored sites. The lines are the convex hulls of the polygons formed in each case
The NmMDS based on orders (stress = 0.1248) and trophic guilds (stress = 0.1386) showed a better fit. In these cases, conserved and disturbed sites were grouped separately, and restored sites were scattered around conserved and disturbed sites (Figs. 6 and 7), respectively.

Non-metric Multidimensional Scaling showing the ordination of the 11 study sites in the PSAER, based on taxonomic orders. Dots represent conserved sites; triangles represent disturbed sites and squares represent restored sites. The lines are the convex hulls of the polygons formed in each case

Non-metric Multidimensional Scaling showing the ordination of the 11 study sites in the PSAER, based on trophic guilds. Dots represent conserved sites; triangles represent disturbed sites and squares represent restored sites. The lines are the convex hulls of the polygons formed in each case
The NMMDS based on families had a higher stress that accepted (stress = 0.2112), so their ordination plot is not presented here.
The χ2 test showed statistical differences among trophic guilds in each type of site (χ2 = 108.88, g.l. = 4, P = 0.0001). Based on adjusted residuals, we observed an overrepresentation of predators in conserved sites (standardized residual = d = 3.193), but an underrepresentation in restored sites (d = -3.452). Moreover, we found an overrepresentation of phytophagous in disturbed sites (d = 7.015) and an underrepresentation in restored sites (d = -6.722). Finally, there was an overrepresentation of saprophagous in restored sites (d = 9.701), and an underrepresentation in conserved and disturbed sites (d = -2.0349 and -7.427, respectively).
The ANOSIM showed that the Bray–Curtis distance between groups was statistically significant for RTUs (R = 0.443, P = 0.006), order (R = 0.33, P = 0.014), and trophic guilds (R = 0.2667, P = 0.034), but no for families (R = 0.2517, P = 0.053) (see Table 3).
Discussion
Since our data lack a species identification in most of the cases, we were unable to obtain a finest trophic guild assignment to our RTUs (using nectarivores or folivores instead of phytophagous, for example), which could provide a better understanding of the dynamic of the PSAER.
Although, several limitations were evident in our methodology, such as the sampling method used, the time of collection and the bias of the pan traps for flying insects, we are aware of them. Given those limitations, we found a differentiation in taxonomical composition in terms of RTU’s and orders between types of sites, these apparently absence of differences in diversity and richness are given by different organisms. This way, despite the caveats in our design, we consider that our results are useful to compare on a preliminary way the community of arthropods in the PASER. This is the first approach to examine whether broader classifications may be useful in future monitoring efforts, and to set a baseline of the state of the restoration at the time of the sampling.
Our results showed differences in abundance among conserved and disturbed sites, whereas restored sites shared values with conserved sites. The restored site R1 presented lower richness compared to all other sites. Also, differences in diversity as effective number of RTUs between restored against conserved and disturbed sites, were observed.
It has been reported that arthropods abundance can increase in habitats dominated by invasive plant species [60], due to the availability of resources for generalist species [61]. Although this tendency varies depending on the species or habitats involved (for example, [62]), this could be the case for the PSAER, as the higher abundance was found in disturbed sites, which are dominated by invasive species like the grass Cenchrus clandestinus [38]. Thus, as the restored sites can not be differentiated from disturbed sites in terms of taxonomic nor guild composition (see below), the same species beneficiated of the presence of exotic plant species in disturbed sites may be present in restored sites, displaying similar abundances.
Site R1 showed the lowest richness of all the studied sites. Since its restoration process began at the same time than R2, and prior to the process in R3, it is possible that the lower richness found in R1 is associated with a low habitat heterogeneity (as in [38]) or low availability of food resources [14] as the site is dominated by the shrub Buddleja cordata with 90% of coverture ([62], Table 1), which, although is a native species, offers limited resources compared to a more diverse site. This may also explain the low effective number of RTUs in this site, as we observed a high dominance of a single species of chironomid, which may be feeding on the nectar or pollen of this particular plant species [63].
Conserved sites showed a similar amount of arthropod diversity in comparison with the disturbed sites, although the latter showed a higher dominance of exotic plants. This could mean that arthropods are not responding exclusively to the plant community. As has been showed in other studies, there may be influence of other characteristics such as the area of the patch and the type of substrate [64], the presence of keystones structures [65], the plant coverage [9] or the intake of soil P or NO3− or even the communication between patches. Moreover, it has been shown that sites with different management may sustain different arthropods communities [66], which may increase the overall beta diversity [67]. If that is the case, taxonomical or guild composition could be considered more important than richness or diversity in the PSAER to evaluate the effect of the restoration practices because they reflect the functionality of the ecosystem rather than only the structure-.
Studies using arthropods as monitoring tools of restoration efforts have been conducted in specific taxonomic groups, such as beetles [68], orthopterans [69], ants [70] or lepidopterans [71, 72]; however a wider taxonomical scope may show a broader pattern where the effect of restoration over different sites and/or different strategies could be identified [2, 66, 67, 73], this is our case.
Although the use of species or morphospecies has been considered the most useful approach to evaluate differences in habitats management in some studies [32, 74, 75], the use of high taxonomic classifications has proved to be a cost effective way to obtain similar results [22, 32]. Nevertheless, as data about the utility of different taxonomic levels have not be consistent among distinct studies [22, 76], it is necessary to test the taxonomic sufficiency in order to optimize resources [77].
In this regard, in our study three out of four NmMDS (RTUs, order and trophic guild compositions) showed spatial differences between conserved and disturbed sites, with restored sites scattered distinctly in each plot. In the RTUs plot, restored sites appeared closer to the disturbed sites, while in the order and trophic guilds plot, restored sites were dispersed indistinctly between the disturbed and conserved sites. However, the stress of the order analysis showed that this was the better criterion to summarize the variation in the PSAER.
Thus, our data suggest that if we are interested just in taxonomical similitude of sites in the PSAER, it is possible to obtain reliable information about the effectiveness of restoration using order classification, which is consistent with other studies [22, 23]. Nevertheless, at least a family classification is recommended in order to assign a coarse trophic guild and to evaluate the restoration of functional structure [1].
Although the ANOSIM shows differences in conserved against disturbed sites using RTUs, orders and trophic guilds, in the two last cases restored sites were considered equal to disturbed but also to conserved ones. This implies that with a coarse view or regarding functional structure, restored sites cannot be differentiated of conserved sites in the PSAER. This suggest that besides the absence of clear differences in richness, total abundance and diversity between types of sites, there are differences in the relative abundance and taxonomical or trophic composition, as has been reported in other studies dealing with disturbed or under restoration sites [60, 66].
In general, the presence of predators is considered a signal of a healthy ecosystem, as it suggests the presence of suitable preys [78], which is consistent with their presence in conserved sites, and a desirable result in restored sites. Moreover, predators can facilitate the recovery of plant biomass in restoration projects [79]. On the other hand, the high dominance of phytophagous in disturbed sites could be explained by the “resource concentration hypothesis”, which states that specialist herbivores should be found in high concentrations in habitats dominated by their host plants [80], which is probably the case for C. clandestinus in the PSAER (Table 1).
Chapin and Starfield [81] proposed the “novel ecosystem” concept, which refers to a site that presents a new combination of species and relative abundances that do not were present previously, as a result of human activities, which could be the case of R1 in our study, given its tendency to diverge in both plant (Table 1) [38] and arthropod communities. Nevertheless, the SER has proposed a list of nine points that must be accomplished in an ecosystem subjected to restoration actions so they can be effectively restored [1], including the “characteristic assemblage of the species that occur in the reference ecosystem and that provide appropriate community structure”. Under this approach, neither of the sites of the PSAER could be considered successfully restored, although some responses of the arthropods to the restoration efforts were observed. Although, since R3 has the more similar trophic guild composition to conserved sites and the same values of effective number of species and estimated richness than R2, which are also higher than in R1, it seems like passive restoration is the best management in terms of cost-benefits regarding arthropods communities in the PSAER.
In restoration ecology, is it typically assumed that if the original plant community is present, the fauna will be present as well [6, 64, 82]. Nevertheless, it is possible that other factors drive the presence of arthropods in a plant-restored site, such as the physical structure, the chemical properties of the soil, the particular life history traits of each species or the elapsed time after the intervention [2, 67]. In that case, it is possibly that for a successful recovery of the original arthropod community, it would be needed to target specific actions for them, as has been proposed particularly for pollinators [5]. Although a review on the theme was published in 1991 [83], an update on the topic feels necessary to achieve integrated restorations.
Conclusions
In this study we found that restored sites showed lower values of diversity measured as effective number of species compared to conserved or disturbed sites, with one site (R1) exhibiting the lowest effective number of RTUs. Although richness values for two restored sites (R2 and R3) were similar to those in conserved and disturbed sites, the restored site R1 showed a significant decrease in richness. Moreover, abundance in restored sites of the PSAER showed shared values with conserved and disturbed sites. According to results obtained with NmMDS and ANOSIM analysis, information provided by orders, trophic guilds and RTUs are not similar. For order and trophic guilds composition, restored sites are either similar to conserved or disturbed sites, while for RTUs composition, restored sites are similar to disturbed sites only. With de data analyzed in this work, restoration actions should be enhanced to achieve the objectives of the ecological restoration. Also we propose that more studies should be done in order to cover epigeal and soil insects rather than only flying ones.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- PSAER:
-
Pedregal de San Angel Ecological Reserve
- SER:
-
Society of Ecological Restoration
- RTUs:
-
Recognizable Taxonomic Units
- NmMDS:
-
Non-metric Multi-Dimensional Scaling
References
Society for Ecological Restoration International Sicience & Policy Working Group. The SER International primer on ecological restoration [Internet]. Tucson; 2004. Available from: www.ser.org
Longcore T. Terrestrial arthropods as indicators of ecological restoration success in coastal sage scrub (California, U.S.A.). Restor Ecol [Internet]. 2003;11(4):397–409. https://doi.org/10.1046/j.1526-100X.2003.rec0221.x.
McAlpine C, Catterall CP, Mac Nally R, Lindenmayer D, Reid JL, Holl KD, et al. Integrating plant- and animal- based perspectives for more effective restoration of biodiversity. Front Ecol Environ. 2016;14(1):37–45.
Majer J. Rehabilitation of disturbed land: long-term prospects for the recolonisation of fauna. Proc Ecol Soc Aust. 1990;16:509–19.
Menz MHM, Phillips RD, Winfree R, Kremen C, Aizen MA, Johnson SD, et al. Reconnecting plants and pollinators: challenges in the restoration of pollination mutualisms. Trends Plant Sci [Internet]. 2011;16(1):4–12 (http://linkinghub.elsevier.com/retrieve/pii/S1360138510001962 cited 2017 Mar 21).
Shuey JA. Habitat re-creation (ecological restoration) as a strategy for conserving insect communities in highly fragmented landscapes. Insects. 2013;4(4):761–80.
Giribet G, Edgecombe GD. Reevaluating the arthropod tree of life. Annu Rev Entomol. 2012;57(1):167–86.
Losey JE, Vaughan M. The economic value of ecological services provided by insects. Bioscience [Internet]. 2006;56(4):311. (cited 2017 Mar 9). https://doi.org/10.1641/0006-3568(2006)56[311:TEVOES]2.0.CO;2.
Majer JD, Brennan KEC, Moir ML. Invertebrates and the restoration of a forest ecosystem: 30 years of research following bauxite mining in Western Australia. Restor Ecol [Internet]. 2007;15(4):S104–15. https://doi.org/10.1111/j.1526-100X.2007.00298.x.
Jansen A. Terrestrial Invertebrate Community Structure as an Indicator of the Success of a Tropical Rainforest Restoration Project. Restor Ecol [Internet]. 1997;5(2):115–24. https://doi.org/10.1046/j.1526-100X.1997.09714.x (cited 2017 Aug 31).
Andersen AN, Fisher A, Hoffmann BD, Read JL, Richards R. Use of terrestrial invertebrates for biodiversity monitoring in Australian rangelands, with particular reference to ants. Austral Ecol [Internet]. 2004;29(1):87–92. https://doi.org/10.1111/j.1442-9993.2004.01362.x (cited 2017 Mar 15).
Christie F, Hochuli D. Responses of wasp communities to urbanization: effects on community resilience and species diversity. J Insect Conserv. 2009;13:213–21.
Schowalter T. Ecological strategies of forest insects: the need for a community-level approach to reforestation. New For. 1986;1:57–66.
Haddad NM, Crutsinger GM, Gross K, Haarstad J, Knops JMH, Tilman D. Plant species loss decreases arthropod diversity and shifts trophic structure. Ecol Lett [Internet]. 2009;12(10):1029–39. https://doi.org/10.1111/j.1461-0248.2009.01356.x (cited 2017 Apr 17).
Valencia-Cuevas L, Tovar-Sánchez E. Oak canopy arthropod communities: which factors shape its structure? Rev Chil Hist Nat. 2015;88:1–22.
Cripps M, Schwarzländer M, Mckenney J, Hinz H, Price W. Biogeographical comparison of the arthropod herbivore communities associated with Lepidium draba in its native, expanded and introduced ranges. J Biogeogr. 2006;33:2107–19.
Hill S, Kotanen P. Phylogenetically structured damage to Asteraceae: susceptibility of native and exotic species to foliar herbivores. Biol Invasions. 2010;12:3333–42.
Hale R, Mac Nally R, Blumstein DT, Swearer SE. Evaluating where and how habitat restoration is undertaken for animals. Restor Ecol. 2019;27(4):775–81.
Krell FT. Parataxonomy vs. taxonomy in biodiversity studies - Pitfalls and applicability of “morphospecies” sorting. Biodivers Conserv. 2004;13(4):795–812.
White PS, Pickett TA. Natural disturbance and patch dynamics: an introduction. In: White PS, Pickett ST, editors. The ecology of natural disturbance and patch dynamics. Orlando: Academic Press; 1985. p. 3–13.
Cotes B, Ruano F, García PA, Pascual F, Campos M. Coccinellid morphospecies as an alternative method for differentiating management regimes in olive orchards. Ecol Indic. 2009;9(3):548–55.
Biaggini M, Consorti R, Dapporto L, Dellacasa M, Paggetti E, Corti C. The taxonomic level order as a possible tool for rapid assessment of arthropod diversity in agricultural landscapes. Agric Ecosyst Environ. 2007;122(2):183–91.
Driessen MM, Kirkpatrick JB. Higher taxa can be effective surrogates for species-level data in detecting changes in invertebrate assemblage structure due to disturbance: a case study using a broad range of orders. Austral Entomol. 2017;58(2):361–9.
Dearborn D, Kark S. Motivations for conserving urban biodiversity. Conserv Biol. 2010;24:432–40.
Peralta-Higuera A, Prado-Molina J. Los límites y la cartografía. In: Lot A, Cano-Santana Z, eds. Biodiversidad del ecosistema del Pedregal de San Ángel. México: Universidad Nacional Autónoma de México; 2009. p. 27–42.
Lot A, Pérez-Escobedo M, Gil-Alarcón G, Rodríguez-Palacios S, Camarena P. La Reserva Ecológica del Pedregal de San Ángel: Atlas de riesgos. Distrito Federal: UNAM, ICyT; 2012. p. 52.
Antonio-Garcés J. Restauración ecológica de la Zona de Amortiguamiento 8 de la Reserva Ecológica del Pedregal de San Ángel, D.F. (México). México: Universidad Nacional autónoma de México; 2008. [cited 2021 Sept 19]. Available from: http://132.248.9.195/ptd2008/junio/0629183/Index.html.
Antonio-Garcés J, Peña M, Cano-Santana Z, Villeda M, Orozco-Segovia A. Cambios en la estructura de la vegetación derivados de acciones de restauración ecológica en las zonas de amortiguamiento Biológicas y Vivero Alto. In: Lot A, Cano-Santana Z, editors. Biodiversidad del Ecosistema del Pedregal de San Ángel. México: Universidad Nacional Autónoma de México; 2009. p. 465–81.
Saucedo-Morquecho E. A. Regeneración de la comunidad vegetal y de dos poblaciones de artrópodos durante un proceso de restauración ecológica en el área de amortiguamiento 8 de la Reserva Ecológica del Pedregal de San Ángel, D.F. México: Universidad Nacional Autónoma de México; 2011. [cited 2021 Sept 19]. Available from: http://132.248.9.195/ptd2012/marzo/0678421/Index.html.
Gómez-Anaya JA, Novelo-Gutiérrez R. A case of successful restoration of a tropical wetland evaluated through its Odonata ( Insecta ) larval assemblage. Rev Biol Trop. 2015;63(December):1043–58.
Cole RJ, Holl KD, Zahawi RA, Wickey P, Townsend AR. Leaf litter arthropod responses to tropical forest restoration. Ecol Evol. 2016;6(15):5158–68.
Schipper AM, Lotterman K, Geertsma M, Leuven RSEW, Hendriks AJ. Using datasets of different taxonomic detail to assess the influence of floodplain characteristics on terrestrial arthropod assemblages. Biodivers Conserv. 2010;19(7):2087–110.
Fraser LH, Harrower WL, Garris HW, Davidson S, Hebert PDN, Howie R, et al. A call for applying trophic structure in ecological restoration. Restor Ecol. 2015;23(5):503–7.
Siebe C. Age and archaeological implications of Xitle volcano, southwestern Basin of Mexico-City. J Volcanol Geotherm Res [Internet]. 2000;104(1–4):45–64 (http://linkinghub.elsevier.com/retrieve/pii/S0377027300001992 cited 2017 Aug 31).
Rzedowski J. Vegetación del Pedregal de San Ángel (D.F., México). An Esc Nac Ciencias Biológicas. 1954;8:59–429.
Rojo A, Rodríguez J. La flora del Pedregal de San Ángel. México: Instituto Nacional de Ecología; 2002. p. 95.
Castillo Argüero S, Montes Cartas G, Romero Romero MA, Martínez Orea Y, Guadarrama Chávez P, Sánchez Gallén I, et al. Boletín de la Sociedad Botánica de México. Boletín la Soc Botánica México. 2004;74:51–75.
Estañol-Tecuatl F, Cano-Santana Z. Recovery of basalt substrate for xeric scrub restoration in a lava field in Mexico City. Ecol Restor. 2017;35(1):41–51.
Ríos-Casanova L, Cano-Santana Z, Godínez-Alvarez H. Patterns of rrthropod diversity in contrasting habitats of el Pedregal de San Angel, a Preserve in Mexico City. Southwest Entomol. 2010;35(2):165–75.
Razo-González M. Estructura de las comunidades de artrópodos edáficos asociados a Pittocaulon (~Senecio) praecox (Asteraceae) en la Reserva Ecológica del Pedregal de San Ángel, D.F. México: Universidad Nacional Autónoma de México; 2013. [cited 2021 Sept 20]. Available from: http://132.248.9.195/ptd2013/enero/0687634/Index.html.
Leong JM, Thorp RW. Colour-coded sampling: The pan trap colour preferences of oligolectic and nonoligolectic bees associated with a vernal pool plant. Ecol Entomol. 1999;24(3):329–35.
Saunders ME, Luck GW. Pan trap catches of pollinator insects vary with habitat. Aust J Entomol [Internet]. 2013;52(2):106–13. https://doi.org/10.1111/aen.12008 (cited 2018 Nov 4).
Popic TJ, Davila YC, Wardle GM. Evaluation of common methods for sampling invertebrate pollinator assemblages: net sampling out-perform pan traps. PLoS One. 2013;8(6):e66665.
Yi Z, Jinchao F, Dayuan X, Weiguo S, Axmacher JC. A Comparison of terrestrial arthropod sampling methods. J Resour Ecol. 2012;3(2):174–82.
Herring J, Ashlock P. A key to the nymphs of the families of Hemiptera (Heteroptera) of America North of Mexico. Florida Entomol. 1971;54(3):207–2012.
Triplehorn C, Johnson N. Borror and DeLong’s Introduction to the study of insects. 7th ed. San Francisco: Thomson Learnings; 2005. p. 864.
Wilcken CF, Brasil DE, Orlato C, Ferreira PJ, Firmito DC. Ocorrência do psilídeo-de-concha (Glycapsis brimblecombei) (Hemiptera: Psyllidae) em florestas de eucalipto no Brasil. 2003.
Smith DR. A synopsis of the sawflies (Hymenoptera, Symphyta) of America south of the United States: Pergidae. Rev Bras Entomol. 1990;34:7–200.
Castellanos-Vargas I, García-García PL, Cano-Santana Z. Diversidad ortopterológica de la Reserva Territorial Sureste de la Ciudad Universitaria (UNAM). Acta Zoológica Mex. 2015;31(1):97–108.
Bridwell JC. Bruchidae infesting seeds of Compositae, with descriptions of new genera and species (Coleoptera). Proc Entomol Soc Washingt. 1931;31:37–42.
Romero Nápoles J, Romero RM. New species of Dahlibruchus Bridwell, 1931 (Coleoptera: Bruchidae) from an archaeological site in Texcoco, Mexico, with some comments about history of the site and bionomics of the insect. Acta Zoológica Mex. 2011;27(2):377–91.
Schmidt C, Leistikow A. Catalogue of genera of the terrestrial Isopoda (Crustacea: Isopoda: Onoscidea). Steenstrupia. 2004;28:191–226.
Wilson J, Messinger Carril O. A guide to North America’s bees. In: The bees in your backyard. New Jersey: Princeton University Press; 2016. p. 288.
Jost L. Entropy and diversity. Oikos. 2006;113:363–75.
Moreno CE, Barragán F, Pineda E, Pavón NP. Reanálisis de la diversidad alfa: Alternativas para interpretar y comparar información sobre comunidades ecológicas. Rev Mex Biodivers. 2011;82(4):1249–61.
Hammer Ø, Harper D, Ryan P. Past: paleontological statistics software package for education and data analysis. Palaeontol Electron [Internet]. 2001;4(1):9.
Gotelli NJ, Colwell RK. Estimating species richness. In: Magurran AE, McGill BJ, editors. Biological diversity: Frontiers in measurement and assessment. Oxford: Oxford University Press; 2010. p. 39–54.
Team RC. R: A languaje and environment for statistical computing. Vienna: R. Foundation for Statistical Computing; 2020.
Clarke KR. Non-parametric multivariate analyses of changes in community structure. Austral Ecol [Internet]. 1993;18(1):117–43. https://doi.org/10.1111/j.1442-9993.1993.tb00438.x (cited 2017 Sep 5).
Jesse WAM, Molleman J, Franken O, Lammers M, Berg MP, Behm JE, et al. Disentangling the effects of plant species invasion and urban development on arthropod community composition. Glob Chang Biol. 2020;26(6):3294–306.
Harvey JA, Fortuna TM. Chemical and structural effects of invasive plants on herbivore–parasitoid/predator interactions in native communities. Entomol Exp Appl [Internet]. 2012;144(1):14–26. https://doi.org/10.1111/j.1570-7458.2012.01252.x (cited 2021 Aug 9).
Andersen EM, Cambrelin MN, Steidl RJ. Responses of grassland arthropods to an invasion by nonnative grasses. Biol Invasions [Internet]. 2019;21(2):405–16. https://doi.org/10.1007/s10530-018-1831-z.
Armitage P. Behaviour and ecology of adults. In: Armitage P, Cranston P, Pinder LC, editors. The Chironomidae: Biology and ecology of non-biting midges. London: Springer-Science+Business Media, B.V; 1995. p. 194–229.
Ksiazek-Mikenas K, Herrmann J, Menke SB, Köhler M. If you build it, will they come? Plant and arthropod diversity on urban green roofs over time. Urban Nat [Internet]. 2018;(Special Issue No. 1: Green Roofs and Urban Biodiversity):52–72. Available from: https://www.eaglehill.us/URNAspecial/pdfs-URNA-sp1/14.U127b.Ksiazek-Mikenas 21.pdf
Tews J, Brose U, Grimm V, Tielbörger K, Wichmann MC, Schwager M, et al. Animal species diversity driven by habitat heterogeneity/diversity: the importance of keystone structures. J Biogeogr [Internet]. 2004;31(1):79–92. https://doi.org/10.1046/j.0305-0270.2003.00994.x.
Teodoro AV, Muñoz A, Tscharntke T, Klein AM, Tylianakis JM. Early succession arthropod community changes on experimental passion fruit plant patches along a land-use gradient in Ecuador. Agric Ecosyst Environ. 2011;140(1–2):14–9.
Burkhalter JC, Moon DC, Rossi AM. Diversity and community similarity of arthropods in response to the restoration of former pine plantations. Southeast Nat. 2013;12(1):121–36.
Blake S, Foster GN, Fisher GEJ, Ligertwood GL. Effects of management practices on the carabid faunas of newly established wildflower meadows in southern Scotland. Ann Zool Fennici. 1996;33:139–47.
Parmenter RR, Macmahon JA, Gilbert CAB. Early Successional Patterns of Arthropod Recolonization on Reclaimed Wyoming Strip Mines: The Grasshoppers (Orthoptera: Acrididae) and Allied Faunas (Orthoptera: Gryllacrididae, Tettigoniidae). Environ Entomol [Internet]. 1991;20(1):135–42. https://doi.org/10.1093/ee/20.1.135 (cited 2017 Mar 21).
Bisevac L, Majer JD. Comparative Study of Ant Communities of Rehabilitated Mineral Sand Mines and Heathland, Western Australia. Restor Ecol [Internet]. 1999;7(2):117–26. https://doi.org/10.1046/j.1526-100X.1999.72002.x (cited 2017 Mar 21).
Hernández AY, Boege K, Al DEL. Lepidopteran herbivory in restored and successional sites in a tropical dry forest. Southwest Nat. 2014;59(1):66–74.
Juan-Baeza I, Martínez-Garza C, Del-Val E. Recovering More than Tree Cover: Herbivores and Herbivory in a Restored Tropical Dry Forest. PLoS One [Internet]. 2015;10(6):e0128583. https://doi.org/10.1371/journal.pone.0128583.
Nakamura BA, Proctor H, Catterall CP, Proctor H, Rain- CPC. Using soil and litter arthropods to assess the state of rainforest restoration. Ecol Manag Restor. 2003;4:20–8.
Andersen AN. Measuring more of biodiversity: Genus richness as a surrogate for species richness in Australian ant faunas. Biol Conserv. 1995;73(1):39–43.
Verdonschot PFM. Data composition and taxonomic resolution in macroinvertebrate stream typology. Hydrobiologia. 2006;566(1):59–74.
Moreno CE, Guevara R, Sánchez-Rojas G, Téllez D, Verdú JR. Community level patterns in diverse systems: A case study of litter fauna in a Mexican pine-oak forest using higher taxa surrogates and re-sampling methods. Acta Oecologica. 2008;33(1):73–84.
Jones FC. Taxonomic sufficiency: The influence of taxonomic resolution on freshwater bioassessments using benthic macroinvertebrates. Environ Rev. 2008;16:45–69.
Estes JA, Terborgh J, Brashares JS, Power ME, Berger J, Bond WJ, et al. Trophic downgrading of planet earth. Science. 2011;333(6040):301–6.
Renzi JJ, He Q, Silliman BR. Harnessing positive species interactions to enhance coastal wetland restoration [Internet]. Vol. 7, Frontiers in Ecology and Evolution. Frontiers Media S.A.; 2019 [cited 2020 Dec 7]. p. 131. Available from: www.frontiersin.org
Root RB. Organization of a Plant-Arthropod Association in Simple and Diverse Habitats: The Fauna of Collards (Brassica Oleracea). Ecol Monogr [Internet]. 1973;43(1):95–124. https://doi.org/10.2307/1942161 (cited 2020 Dec 7).
Chapin FS, Starfield A. Time lags and Novel Ecosystems in response to transient climatic change in artic Alaska. Clim Change [Internet]. 1997;35(4):449–61. https://doi.org/10.1023/A:1005337705025 (cited 2017 Aug 31).
Gratton C, Denno RF. Restoration of arthropod assemblages in a Spartina salt marsh following removal of the invasive plant phragmites australis. Restor Ecol. 2005;13(2):358–72.
Usher MB, Jefferson RG. Creating New and Successional Habitats for Arthropods. In: The Conservation of Insects and their Habitats [Internet]. London: The Royal Entomological Society of London; 1991. p. 263–91. https://doi.org/10.1016/B978-0-12-181370-3.50017-8.
Acknowledgements
R.A. Farfán-Beltrán for the elaboration of Fig. 1, Iván Castellanos-Vargas for its technical support, and Paulina Corona-Tejeda for the data analysis.
Funding
We thanks to the next projects: PAPIIT-UNAM IV200117 (Análisis ecosocial de una reserva urbana para la sustentabilidad en el campus de Ciudad Universitaria) given to Luis Zambrano González, Zenón Cano Santana & Alicia Castillo Álvarez. PAPIIT-UNAM IN222006 (Evaluación del estado de conservación y restauración ecológica de la Reserva del Pedregal de San Ángel) given to Zenón Cano Santana and PAPIME-UNAM PE204809 (Regeneración de áreas verdes de vegetación natural de Ciudad Universitaria y su Reserva Ecológica sujetas a restauración) given to Zenón Cano Santana.
Author information
Authors and Affiliations
Contributions
EF, ZC, JHC conceived and designed the research, EF performed insect collections; EF analyzed the data; ZCS, JHC contributed reagents/materials/analysis tools; EF, ZC, MCHP & JHC wrote and edited the manuscript. The author(s) read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Table S1.
List of species and morphospecies registered in four conserved sites (CS), three restored sites (RS) and four disturbed sites (DS) in the PSAER. The number represents the abundance of each morphospecies.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Farfán-Beltrán, M.E., Chávez-Pesqueira, M., Hernández-Cumplido, J. et al. A quick evaluation of ecological restoration based on arthropod communities and trophic guilds in an urban ecological preserve in Mexico City. Rev. Chil. de Hist. Nat. 95, 4 (2022). https://doi.org/10.1186/s40693-022-00108-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40693-022-00108-8