
Abstract
Background
Nest sanitation behavior is one of the most important means to ensure high reproductive efficiency. In avian brood parasitism, nest sanitation behavior may be a pre-adaptation of host birds that allows them to identify the parasitic eggs, so that egg discrimination behavior may have evolved from nest sanitation behavior. However, whether nest sanitation behavior could improve egg rejection in cuckoo hosts was inconclusive.
Methods
In this study, we investigated the relationship between nest sanitation and egg discrimination behavior in a potential cuckoo host, the Brown-breasted Bulbul (Pycnonotus xanthorrhous) with two experimental groups. In the first group, we added a blue, non-mimetic egg to the nest of the host, while in the second group we added a blue, non-mimetic egg and a peanut half-shell.
Results
The results showed that in the first group, the probability of rejecting the non-mimetic eggs was 53.8% (n = 26 nests). In comparison, all of the Brown-breasted Bulbuls in the second group were able to rapidly remove the peanut shells from the nest, but only 52.6% (n = 19 nests) rejected the non-mimetic eggs. The rejection rates of the non-mimetic eggs in both experimental groups were not significantly different.
Conclusions
Our study indicated that nest sanitation behavior of Brown-breasted Bulbuls did not influence their egg recognition and that egg discrimination ability of Brown-breasted Bulbuls was not directly related to nest sanitation behavior.
Similar content being viewed by others


Background
Nest sanitation behavior is the removal of foreign objects from the nest, and is a very common behavior in breeding birds (Alvarez et al. 1976; Moskát et al. 2003; Underwood and Sealy 2006; Guigueno and Sealy 2009, 2017). This behavior can improve the incubation efficiency of the parent birds, reduce physical damage to eggs from foreign objects, lower the chance of egg infection by pathogenic bacteria, and decrease the risk of predation (Guigueno and Sealy 2012). For instance, parent birds usually clear the leaves, gravel, broken eggshells, food debris, nestling feces, and other debris from the nest (Weatherhead 1984; Kemal and Rothstein 1988; Petit et al. 1989; Lang et al. 2002; Evans et al. 2016). Nest sanitation is also prevalent in inter-specific brood parasitism birds, and may be associated with the evolution of egg discrimination in the anti-parasitic strategy of host birds (Rothstein 1975).
Previous work suggested that the evolution of egg rejection in cuckoo hosts is driven by the pressure from cuckoo parasitism; the higher the parasitism pressure, the stronger the egg discrimination ability of the hosts (Davies and Brooke 1989; Moksnes et al. 1991). However, egg rejection by hosts may be an extension of their nest sanitation behavior (Rothstein 1975; Guigueno and Sealy 2009; Yang et al. 2015b). Many studies have shown the relationship between the nest sanitation behavior and egg discrimination ability of cuckoo hosts using various non-ovoid foreign objects (Ortega and Cruz 1988; Moskát et al. 2003; Underwood and Sealy 2006; Yang et al. 2015a, b; Luro and Hauber 2017; Peer 2017; review in Rothstein 1975; Peer and Sealy 2004; Guigueno and Sealy 2017). Moskát et al. (2003) proposed that the less similar the shape of a foreign object to a cuckoo egg, the higher the host rejection rate, so that the ability of the host to recognize a cuckoo egg is an extension of its nest sanitation behavior. In addition, hosts with strong egg discrimination ability tend to be more capable of rejecting non-ovoid objects than are hosts without egg discrimination ability (Yang et al. 2015b), indicating that host egg discrimination ability is associated with nest sanitation behaviour (Underwood and Sealy 2006; Guigueno and Sealy 2009; Yang et al. 2015a). The removal of non-egg-shaped objects is stable across pre-hatching stages, unless precluded by rejection costs (Guigueno and Sealy 2017). Therefore, nest sanitation behaviour may be a preadaptation to egg discrimination behaviour, as hypothesized by Guigueno and Sealy (2012).
In Tree Sparrows (Passer montanus), it seems that nest sanitation plays a key role in the evolution of the removal of parasitic eggs (Poláček et al. 2013; but see Yang et al. 2016). Yang et al. (2015b) showed that the nest sanitation behavior of Barn Swallows (Hirundo rustica) was a pre-adaptation to identifying the eggs of parasitic cuckoos. They further demonstrated that nest sanitation behavior could significantly improve the ability of parent swallows to reject non-mimetic eggs (Yang et al. 2015a). In contrast to Yang et al. (2015a), the Red-winged Blackbird (Agelaius phoeniceus), a host of the Brown-headed Cowbird (Molothrus ater), was not more likely reject a model cowbird egg, with its maculation but different background colour, if the model egg was simultaneously added to the nest with flagging tape or a pine cone bract scale (Peer 2017). Similarly, whether American Robins (Turdus migratorius) removed a Douglas Fir (Pseudotsuga menziesii) leaf clipping from their nests or not showed no effect on rejection of a model cowbird egg (Luro and Hauber 2017). Therefore, a potential proximate link between nest sanitation and egg ejection in avian brood parasite-host system is still inconclusive (Luro and Hauber 2017).
For research, replicating work is crucial to assess the generality of findings (Nakagawa and Parker 2015). However, the complexity of data collection in ecology, especially in field ecology, often precludes the possibility of going beyond single-population studies (Canestrari et al. 2017). For example, a study by Canestrari et al. (2014) suggested a mechanism based on the malodorous secretion of Great Spotted Cuckoo (Clamator glandarius) chicks to explain why the presence of the parasite in the nests of Carrion Crows (Corvus corone) in northern Spain increased the probability of nest success as compared to non-parasitized nests. However, also in Spain, Soler et al. (2017) found no evidence to support an anti-depredatory function of cuckoo chicks in their studied populations (but see Canestrari et al. 2017). Clearly, more studies addressing whether nest sanitation elicits egg rejection are needed (Guigueno and Sealy 2017).
In this study, we investigated the relationship between nest sanitation and egg rejection in an open-cup nesting potential cuckoo host, the Brown-breasted Bulbul (Pycnonotus xanthorrhous). Although the Brown-breasted Bulbul in China has not been recorded as being parasitized by parasitic cuckoos (Yang et al. 2012; Su et al. 2016), most species of bulbuls were listed as hosts for many cuckoo species (Cuculus spp.) (Davies 2000), and in Africa, Cape Bulbuls (Pycnonotus capensis) and other bulbul species (Pycnonotus spp.) were common hosts of the Jacobin Cuckoo (Clamator jacobinus) (Krüger 2011). Here, we used blue, non-mimetic eggs and peanut half-shells to evaluate whether nest sanitation behavior in Brown-breasted Bulbuls influences their egg discrimination ability following the experimental protocols by Yang et al. (2015a). If nest sanitation behavior is directly associated with egg discrimination ability, then egg discrimination against non-mimetic eggs should be significantly higher in nests with peanut shells than in those without. We predict that nest sanitation behavior would elicit egg rejection in the Brown-breasted Bulbul.
Methods
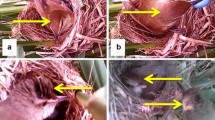
The study was conducted during the breeding season of the Brown-breasted Bulbul (April–August) in 2014–2016 in Kuankuoshui National Nature Reserve (28°10′N, 107°10′E) in Guizhou, Southwest China. Bulbul nests were searched covering all habitats in the study area, and breeding parameters, such as the nesting and hatching dates, were recorded. We estimated the hatching stage following Hays and LeCroy (1971). No cuckoo parasitism was recorded in Brown-breasted Bulbuls although they are abundant in our study area and their nests were easy to find (Yang et al. 2012; Su et al. 2016). We followed the experimental protocols by Yang et al. (2015a) and performed the experiment during the early incubation stage. The blue, non-mimetic eggs were made from baked soft clay, with a mean weight of 4.47 ± 0.16 g, length of 21.42 ± 0.46 mm, and width of 15.99 ± 0.42 mm (n = 15). The mean weight, length, and width of the peanut shells was 1.76 ± 0.45 g, 36.57 ± 4.15 mm, and 15.29 ± 1.38 mm, respectively (n = 15). The mean weight, length, and width of the Brown-breasted Bulbul eggs was 2.92 ± 0.29 g, 21.88 ± 1.02 mm, and 16.35 ± 0.54 mm, respectively (n = 74) (Fig. 1). In the first experimental group, we directly added a non-mimetic blue egg to nests of Brown-breasted Bulbuls to evaluate the ability of the host to discriminate against the non-mimetic egg within 6 days (Davies and Brooke 1989; Moksnes et al. 1991; Yang et al. 2015a). In the second group, we added a peanut half-shell to nests of Brown-breasted Bulbuls in addition to one non-mimetic blue egg. We used Fisher’s exact test to compare the probability of rejecting non-mimetic eggs between different experimental groups. Data analysis was performed using IBM SPSS 19.0 software (IBM Inc., USA).

An example of the experimental group with a half peanut shell addition
Results
In the first experimental group, 26 nests of Brown-breasted Bulbuls were tested for discrimination ability against a non-mimetic egg. In 14 out of 26 nests, the non-mimetic blue eggs were rejected (53.8%) (Table 1). In the second experimental group, 19 nests were tested for discrimination ability against a non-mimetic egg together with a peanut shell. All the peanut shells were removed within the same day that they were added (19 out of 19 nests, 100%), while the non-mimetic eggs were rejected in 10 out of 19 nests (52.6%) (Table 1). Brown-breasted Bulbuls used grasp ejection to reject peanut shells and/or model eggs with no rejection cost. All egg ejections occurred after the peanut shells were removed and all parent birds rejected the non-mimetic eggs within 3 days. There was no significant difference in the probability of rejecting non-mimetic eggs between the two experimental groups (χ2 = 0.007, df = 1, p = 0.936). In the experimental group with both non-mimetic eggs and peanut shells, the rejection rate of non-mimetic eggs was significantly lower than that of peanut shells (χ2 = 11.79, df = 1, p =0.0006), indicating that Brown-breasted Bulbuls did not treat the non-mimetic eggs as foreign objects.
Discussion
Our results showed that although Brown-breasted Bulbuls have strong nest sanitation abilities, their nest sanitation behavior did not directly affect their recognition of non-mimetic eggs. The egg rejection rates were similar for the nests with and without the addition of peanut shells. In addition, the Brown-breasted Bulbuls did not treat the non-mimetic eggs as foreign objects, such as peanut shells. This was shown in the nests with both peanut shells and non-mimetic eggs, in which all of the birds removed the peanut shells, but only 52.6% rejected the non-mimetic eggs. Therefore, this indicated the influence of the shape of the object on egg discrimination ability, similar to the results of previous studies (Ortega and Cruz 1988; Guigueno and Sealy 2009, 2012).
Nest sanitation behavior is considered a pre-adaptation of host bird egg discrimination ability, as the two behaviors share a common aspect of the removal of objects from the nest, although some nest sanitation behavior is accomplished by burying or swallowing (Guigueno and Sealy 2012). However, the two behaviors differ with respect to the shape and color of the foreign object and the similarity to the host egg. There is also no clear link between nest sanitation in cowbird hosts (Rothstein 1975; Peer and Sealy 2004). In addition, some hosts do not practice nest sanitation, but can reject some eggs (Peer 2017). Underwood and Sealy (2006) suggested that the ability of American Robins and Gray Catbirds (Dumetella carolinensis) to recognize non-egg objects was significantly higher than that of other species without egg discrimination ability.
Moskát et al. (2003) examined the relationship between nest sanitation and egg rejection in the Great Reed Warbler (Acrocephalus arundinaceus) and showed that the rejection rates of the non-egg models were significantly higher than those of cuckoo model eggs, either during hatching or in the early stage of incubation. Guigueno and Sealy (2009) also showed that in American Yellow Warblers (Setophaga petechia) the burial probability of non-egg objects was significantly greater than that of the simulated cowbird-egg-shaped models during the pre-hatching and hatching stages. These results strongly indicated that host egg discrimination ability is likely to be an extension of nest sanitation behavior.
Yang et al. (2015a) studied the nest sanitation and egg rejection of Barn Swallows, which are hosts of the Common Cuckoo (Cuculus canorus). In nests with only non-mimetic eggs, the probability of swallows rejecting the model eggs was 27%, whereas in nests with model eggs and peanut shells, all of the individuals were able to remove the peanut shells and the probability of rejecting the model eggs increased to 56%, demonstrating the significant influence of host bird nest sanitation behavior on egg discrimination. Similar to Barn Swallows, Brown-breasted Bulbuls also showed moderate egg discrimination ability against non-mimetic eggs (64.1%, Su et al. 2016; 53.8%, this study), which were similar to American Robins (65%; Luro and Hauber 2017) but stronger than the discrimination ability of Red-winged Blackbirds (0%; Peer 2017). However, our results showed no effect of nest sanitation behavior on egg discrimination, but were consistent with those by Luro and Hauber (2017) and Peer (2017), who found that although the tested hosts showed nest sanitation behavior, the addition of foreign objects to the nest did not influence host egg rejection. Rothstein (1975) also found that Red-winged Blackbirds show a very low level of response to cowbird eggs, and foreign objects removal did not influence their egg rejection.
Previous studies have shown that when a host bird sees an adult cuckoo around its nest, it is stimulated to increase the probability of rejecting the parasitic eggs in its nest (Moksnes et al. 1993; Davies 2000; Hosoi and Rothstein 2000; Guigueno and Sealy 2011). Yang et al. (2015a) further showed that this type of stimulation was also produced by foreign objects, which increased egg rejection in Barn Swallows. Such type of stimulation was effective in Barn Swallow populations, but not in American Robins, Red-winged Blackbirds or Brown-breasted Bulbuls. We suggested the possible reason for this discrepancy could be that the nesting habitat of Barn Swallows is different from the latter three, as Barn Swallows are a communally nesting species and their nests are usually built under the eaves of human habitation (Liang et al. 2013; Yang et al. 2015b). In such a nesting habitat, there are relatively fewer pieces of debris that fall into swallow nests during the incubation period. In addition, since the swallow nests are next to the eaves and are relatively dark, small pieces of debris may be difficult for the birds to see. As a result, swallows do not frequently clean their nests during incubation. Thus, when peanut shells suddenly appeared as apparent foreign objects in their nests, nest sanitation behavior was activated and their egg discrimination abilities were also stimulated. By contrast, in the natural habitat of the Brown-breasted Bulbuls, American Robins and Red-winged Blackbirds, many pieces of debris often fall into their nests during the incubation period. In addition, as their nests are cup-shaped and open, the fallen small-sized debris is easily seen. Thus, they often clean their nests throughout the incubation period, and nest sanitation behavior is a normal part of their behavior during incubation. When the peanut shells were added to their nests, the additional stimulation effect was not sufficient to be detected or to induce them to reject the non-mimetic eggs. Clearly, large-scale and multiple-population studies, and more in-depth explanation of these results, e.g., the costs of nest sanitation versus egg rejection and the role of host age structure in nest sanitation, need further investigation in the future.
Conclusions
The results of this study showed that nest sanitation behavior is common among Brown-breasted Bulbuls, but the removal of foreign objects from the nest does not have a direct impact on their egg discrimination ability. In other words, nest sanitation behavior of the Brown-breasted Bulbul was not associated with egg discrimination ability, and nest sanitation did not elicit egg rejection in the Brown-breasted Bulbul. In addition, our study highlighted the necessity to test nest sanitation versus egg rejection in more host species and in multiple populations.
References
Alvarez F, de Reyna LA, Segura M. Experimental brood parasitism of the magpie (Pica pica). Anim Behav. 1976;24:907–16.
Canestrari D, Bolopo D, Turlings TCJ, Röder G, Marcos JM, Baglione V. From parasitism to mutualism: unexpected interactions between a cuckoo and its host. Science. 2014;343:1350–2.
Canestrari D, Bolopo D, Turlings TCJ, Röder G, Marcos JM, Baglione V, Formal comment to Soler et al. Great spotted cuckoo nestlings have no antipredatory effect on magpie or carrion crow host nests in southern Spain. PLoS ONE. 2017;12:e0184446.
Davies NB, de Brooke ML. An experimental study of co-evolution between the cuckoo, Cuculus canorus, and its hosts. I. Host egg discrimination. J Anim Ecol. 1989;58:207–24.
Davies NB. Cuckoos, cowbirds and other cheats. London: T & AD Poyser; 2000.
Evans JK, Griffith SC, Klasing KC, Buchanan KL. Impact of nest sanitation on the immune system of parents and nestlings in a passerine bird. J Exp Biol. 2016;219:1985–93.
Guigueno MF, Sealy SG. Nest sanitation plays a role in egg burial by yellow warblers. Ethology. 2009;115:247–56.
Guigueno MF, Sealy SG. Aggression towards egg-removing cowbird elicits clutch abandonment in parasitized yellow warblers, Dendroica petechia. Anim Behav. 2011;81:211–8.
Guigueno MF, Sealy SG. Nest sanitation in passerine birds: implications for egg rejection in hosts of brood parasites. J Ornithol. 2012;153:35–52.
Guigueno MF, Sealy SG. Implications of nest sanitation in the evolution of egg rejection. In: Soler M, editor. Avian brood parasitism: behaviour, ecology, evolution and coevolution. Cham: Springer International Publishing AG; 2017. p. 385–99.
Hays H, LeCroy M. Field criteria for determining incubation stage in eggs of the common tern. Wilson Bull. 1971;83:425–9.
Hosoi SA, Rothstein SI. Nest desertion and cowbird parasitism: evidence for evolved responses and evolutionary lag. Anim Behav. 2000;59:823–40.
Kemal RE, Rothstein SI. Mechanisms of avian egg recognition: adaptive responses to eggs with broken shells. Anim Behav. 1988;36:175–83.
Krüger O. Brood parasitism selects for no defence in a cuckoo host. Proc R Soc Lond B. 2011;278:2777–83.
Lang JD, Straight CA, Gowaty PA. Observations of fecal sac disposal by eastern bluebirds. Condor. 2002;104:205–7.
Liang W, Yang C, Wang L, Møller AP. Avoiding parasitism by breeding indoors: cuckoo parasitism of hirundines and rejection of eggs. Behav Ecol Sociobiol. 2013;67:913–8.
Luro A, Hauber ME. A test of the nest sanitation hypothesis for the evolution of foreign egg rejection in an avian brood parasite rejecter host species. Sci Nat. 2017;104:14.
Moksnes A, Røskaft E, Bičík V, Honza M, Øien IJ. Cuckoo Cuculus canorus parasitism on Acrocephalus warblers in Southern Moravia in The Czech Republic. J Ornithol. 1993;134:425–34.
Moksnes A, Røskraft E, Bráa AT, Korsnes L, Lampe HM, Pedersen HC. Behavioural responses of potential hosts towards artificial cuckoo eggs and dummies. Behaviour. 1991;116:64–89.
Moskát C, Szekely T, Kisbenedek T, Karcza Z, Bártol I. The importance of nest cleaning in egg rejection behaviour of great reed warblers Acrocephalus arundinaceus. J Avian Biol. 2003;34:16–9.
Nakagawa S, Parker TH. Replicating research in ecology and evolution: feasibility, incentives, and the cost-benefit conundrum. BMC Biol. 2015;13:88.
Ortega CP, Cruz A. Mechanisms of egg acceptance by marsh-dwelling blackbirds. Condor. 1988;90:349–58.
Petit KE, Petit LJ, Petit DR. Fecal sac removal: do the pattern and distance of dispersal affect the chance of nest predation? Condor. 1989;91:479–82.
Peer BD. Nest sanitation does not elicit egg ejection in a brown-headed cowbird host. Anim Cogn. 2017;20:371–4.
Peer BD, Sealy SG. Correlates of egg rejection in hosts of the brown-headed cowbird. Condor. 2004;106:580–99.
Poláček M, Griggio M, Bartíková M, Hoi H. Nest sanitation as the evolutionary background for egg ejection behaviour and the role of motivation for object removal. PLoS ONE. 2013;8:e78771.
Rothstein SI. An experimental and teleonomic investigation of avian brood parasitism. Condor. 1975;77:250–71.
Soler M, de Neve L, Roldán M, Pérez-Contreras T, Soler JJ. Great spotted cuckoo nestlings have no antipredatory effect on magpie or carrion crow host nests in southern Spain. PLoS ONE. 2017;12:e0173080.
Su T, Yang C, Liang G, Ding C, Liang W. Association between nest defense and egg rejection behavior of brown-breasted bulbuls (Pycnonotus xanthorrhous). Auk. 2016;133:231–5.
Underwood TJ, Sealy SG. Influence of shape on egg discrimination in American robins and gray catbirds. Ethology. 2006;112:164–73.
Weatherhead PJ. Fecal sac removal by tree swallows: the cost of cleanliness. Condor. 1984;86:187–91.
Yang C, Chen M, Wang L, Liang W, Møller AP. Nest sanitation elicits egg discrimination in cuckoo hosts. Anim Cogn. 2015a;18:1373–7.
Yang C, Liang W, Antonov A, Cai Y, Stokke BG, Fossøy F, Moksnes A, Røskaft E. Diversity of parasitic cuckoos and their hosts in China. Chin Birds. 2012;3:9–32.
Yang C, Wang L, Liang W, Møller AP. Nest sanitation behavior in hirundines as a pre-adaptation to egg rejection to counter brood parasitism. Anim Cogn. 2015b;18:355–60.
Yang C, Wang L, Liang W, Møller AP. Contrasting egg recognition between European and Asian populations of tree sparrows (Passer montanus). Behav Proc. 2016;125:85–8.
Authors’ contributions
WL and SC conceived and designed the study. TS conducted the experiments in the field. CY performed the data analysis. TS wrote the early draft, WL and SC discussed and revised the manuscript. All authors read and approved the final manuscript.
Acknowledgements
We would like to thank Dr. Ken Cheng and three anonymous reviewers for their helpful comments on an early version of this manuscript. We thank Kuankuoshui National Nature Reserve, Guizhou, China, for permission to undertake this study. We are grateful to Juan Huo and Guoxian Liang for their assistance with fieldwork.
Competing interests
The authors declare that they have no competing interests. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Consent for publication
Not applicable.
Ethical approval
This article does not contain any studies with human participants performed by any of the authors. The experiments comply with the current laws of China. Fieldwork was carried out under permission from Kuankuoshui National Nature Reserve, Guizhou, China. Experimental procedures were in agreement with the Animal Research Ethics Committee of Hainan Provincial Education Centre for Ecology and Environment, Hainan Normal University (permit no. HNECEE-2016-003).
Funding
This work was supported by the National Natural Science Foundation of China (Nos. 31672303 to CY, 31472013 and 31772453 to WL).
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Su, T., Yang, C., Chen, S. et al. Does nest sanitation elicit egg rejection in an open-cup nesting cuckoo host rejecter?. Avian Res 9, 27 (2018). https://doi.org/10.1186/s40657-018-0119-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40657-018-0119-4