
Abstract
Plant cellulose synthase is one of the important glycosyltransferases, which catalyzes the synthesis of the paracrystalline of H-bonded-β-(1,4)-glucose chains. This study isolated the cellulose synthase 3 (Ces3) sequence from sugarcane (Saccharum officinarum L.) leaves. SoCes3 (GenBank accession No. MG324347) has a full-length cDNA sequence of 3625 bp. It contains an open reading frame (3225 bp), encoding 1074 amino acids with a molecular weight of about 120.89 kDa and isoelectric point of 6.26. SoCes3 protein showed high activity with other plant cellulose synthases. The recombinant protein contains plant cellulose synthase (Ces) protein conservative domains. In subcellular localization experiments, the fusion protein of SoCes3 with green fluorescent protein (GFP) was specifically localized in the cell membrane. The gene expression of SoCes3 was found in the leaf, leaf sheath, and internodes of the sugarcane stem. The highest expression level was found in the internode, especially with the highest expression level in the 5th internode and lowest in the leaves, and the gene expression level of SoCes3 was upregulated by PP333 and not in gibberellic acid-treated plants. It was conducted in tobacco plants to understand the biotechnological potential of SoCes3. The contents of cellulose and lignin were increased in SoCes3-overexpressing tobacco. Transcriptomic analysis showed that the transgenic tobacco induced different genes associated with different biological regulatory processes. Differentially expressed genes (DEGs) mediated plant hormone signal transduction, starch and sucrose metabolism signaling pathways were widely induced and mostly upregulated. The transcription levels in SoCes3-overexpressing transgenic lines were higher than wild-type.
Graphical Abstract

Similar content being viewed by others


Introduction
Sugarcane (Saccharum spp. Hybrid) is a major C4 energy crop. It accounts for over 90% of China total sugar production with optimum biomass and fiber. Developing high yield, increased biomass, and stress resistance are the major goals of sugarcane breeding. Fiber is one of the main components in sugarcane stems with higher content (11.5–12.5%) [1, 2]. Cellulose synthase is the most important for sugarcane growth and cell development. Cellulose is the world’s most abundant biopolymer and a major component of plant cell walls [3]. Cellulose consists of a β-1,4-linked glucose chain synthesized by cellulose synthase (CesA, EC2.1.4.12) [4]. The members of the CesA genes family in plants encode a glycosyltransferase that plays a key role in the process of cellulose synthesis [4,5,6,7,8].
Plant cellulose synthase A (CesA) belongs to a multigene family with complex expression patterns. The CesA gene was first cloned from the bacteria that produced extracellular cellulose [9, 10]. Due to the high cellulose content in cotton fiber cell walls, it is no surprise that plant homologs were identified from the cotton fibers undergoing active secondary cellulose synthesis walls (GhCesA1/GhCesA2) [11]. The CesA genes have also been isolated from other plant species, such as Arabidopsis [12, 13], Oryza sativa [14], Zea mays [15], Populus [16,17,18,19], Boehmeria nivea (Linn.) Gaud. [20]. These orthologous CesA proteins possess similar functions and exhibit high sequence similarity. The CesA gene family in monocot crop plants, such as Zea mays, Hordeum vulgare, and Oryza sativa were screened by the analysis of cDNA, expressed sequence tags, and genome sequencing [21,22,23,24].
In higher plants, the family of CesA genes showed interesting similarities and differences from the genes in the bacteria that predicted proteins in plants contain a plant-specific conserved and a hypervariable (HVR-2) domain that separates the domains containing these conserved residues [4]. The CesA proteins are inferred to be the components of cellulose synthase complexes (CSCs) associated in the synthesis of the primary cell wall [3, 23, 25]. Several lines of evidence implicate the plant CesA genes in the process of cellulose synthesis. Genetic and biochemical responses shown that the three unique CesA isoforms are required for CSCs function that separate CSCs associated with cellulose biosynthesis of primary and secondary cell walls [15, 25,26,27,28,29]. In Arabidopsis thaliana, CesA1, CesA3, and CesA6 are required for PCW cellulose biosynthesis, while CesA4, CesA7, and CesA8 are required during the development of secondary cell wall. The remaining CesA (CesA2, CesA5, CesA9, and CesA10) are linked in tissue-specific processes and partially redundant with CesA6 [13, 25, 27,28,29].
CesA genes are poorly identified and annotated in sugarcane plants. We identified a unigene highly homologous to the Zea Mays Ces3 sequence isolated from the sugarcane full-length cDNA library in response to limited water supply (i.e., drought stress). This study utilized an innovative approach to isolate and characterize a full-length cellulose synthase gene (SoCesA3) from sugarcane plants (Saccharum spp.). The functions of the SoCesA3 were inferred based on their phylogenetic relationships and differential expression patterns. This study not only lays a foundation for understanding the biosynthesis of cellulose and further functional analysis of SoCesA3 in sugarcane, but also sheds light on future improvements in the production and properties of cellulosic biomass in sugarcane plants. In addition, a pair of stably expressed housekeeping genes were screened out and validated in sugarcane plants for the quantitative real-time reverse transcription PCR (qRT-PCR) experiment.
Materials and methods
Plant material and growth conditions
The main sugarcane cultivar (Taitang 22 from Guangxi Province, China) was used for gene cloning. Two genotypes (Guitang 46 and Guitang 49) were planted in the greenhouse of Sugarcane Research Institute, Guangxi Academy of Agricultural Sciences, Nanning, Guangxi, China for gene expression analysis. Tobacco (Nicotiana tabacum L.cv. Petit Havana) plants were used in subcellular localization of SoCes3 and gene overexpression.
The first leaf was collected from the Taitang22 cultivar for gene isolation at the pre-elongation stage, which contained 9–10 leaves [30]. Two genotypes (GT 46 and GT 49) at different leaves, such as 1, 3, and 5th leaf position (from top to bottom), leaf sheaths and leaf stalks were collected for gene expression analysis during elongation stage of plant. Gibberellic acid (GA, 100 mg/L) and paclobutrazol (PP333, 100 mg/L) were sprayed on GT 49 variety at seedling stage with contained 3–5 leaves (45 days). The plant leaf samples were collected at specific time intervals, i.e., 15, 30, 40, 50, and 60 days after treatment. During 60 days, different leaves, leaf sheaths and stalks (1, 3 and 5th leaf position from top to bottom) were collected. These plant materials were immediately frozen and stored in liquid nitrogen for RNA isolation.
Full-length gene cloning and sequence bioinformatics analysis of SoCes3 genes
RNA was extracted from sugarcane leaves, sheaths, and stems using the TRIzol component (Invitrogen, USA). Then, first-strand cDNA was synthesized from 1 to 5 µg of total RNA using PrimeScript 1st Strand cDNA Synthesis Kit (TaKaRa, Dalian, China). Based on the unigene sequence highly homologous to the Zea Mays Ces3 sequence, it was isolated from the sugarcane full-length cDNA library in response to limited water supply (drought stress). A primer pair CES-F and CES-R was designed and used to amplify the cDNA of SoCesA3 (Table 1). The PCR reaction was performed in a volume of 50 μl containing 1.0 μl of template, 25 µl of Premix Primes STAR HS (TaKaRa, Dalian, China), 1.0 µl of each 20 µmol/L upstream and downstream primer, 22 µl of double-distilled water (DDW), and using the following program: 1 cycle at 94 ℃ for 4 min, 35 cycles at 98 ℃ for 10 s, 55 ℃ for 15 s, 72 ℃ for 3 min and 72 ℃ for 10 min. After PCR amplification, the products were purified and cloned into the PMD19-T vector (TaKaRa, Dalian, China) and sequenced on both strands in Takara Bio Institute (TaKaRa, Dalian, China). Based on the sequence of partial SoCes3 gene fragments, gene-specific primers for 5’-RACE and 3’-RACE was designed (Table 1). 5’-RACE was performed using SMARTer®RACE 5’/3’ Kit (TaKaRa, Dalian, China), and 3’-RACE using 3’-Full RACE Core Set with Prime Script Rtase Kit (TaKaRa, Dalian, China). 5’- and 3’-end cDNA fragments of SoCes3 gene were obtained.
Analysis of sequence alignment
The obtained DNA sequence and amino acids were analyzed using bioinformatics tools (www.ncbi.nlm.nih.gov, www.expasy.org). The multiple sequence alignment was conducted with Vector NTI 8.0. Phylogenetic analysis of SoCes3 was aligned with DNAMAN 5.2.2 software, and evolutionary studies were constructed with the neighbor-joining (NJ) method using MEGA 6.0 software and the neighbor-joining way with 1000 bootstrap replicates [31, 32].
Subcellular localization of SoCes3
To construct the SoCes3-GFP fusion fragment for the expression in tobacco epidermal cells, PCR products were digested with NcoI and SpeI and then ligated to the pCAMBIA1302 vector. The recombinant plasmids (pCAMBIA1302-SoCes3) were confirmed by double digestion with NcoI and SpeI and sequencing, then transferred to the TOP10 clone strain. The empty plasmid pCAMBIA1302 was used as the control. A positive monoclonal antibody containing recombinant plasmid pCAMBIA-SoCes3 was amplified in YEP liquid culture (containing 50 mg/mL rifampicin and 25 mg/mL kanamycin) and cultivated to OD600 nm = 1.5–2.0 at 28 ℃. After centrifugation, Agrobacterium bacteria were collected. Then, 50 mL of Agrobacterium suspension was added, mixed, injected into the tobacco epidermis, and cultured for 72 h under normal illumination conditions. The tobacco epidermis was observed with a confocal laser scanning microscope (excitation wavelength 488 nm; emission wavelength 625–725 nm).
Quantitative real-time PCR analysis
Specific primers for qRT-PCR are shown in Table 1. qRT-PCR experiments were performed using HiScrip®II 1st Strand cDNA Synthesis Kit (Vazyme Biotech Co., Ltd, Nanjing, China) and Power qRT-PCR PreMix Kit (Generay Biotech Co., Ltd, Shanghai, China). The total volume of the reaction system was 7 µL SYBR Green I Master Mix (2 ×); 0.5 µL Forward-Primer (10 µmol/L); 0.5 µL Reverse-Primer (10 µmol/L); 8 µL cDNA template. Three biological replicates (n = 3) were performed for each sample. In order to draw a melting curve for the separation of the genes, the temperature cycle was as follows: 95 ℃ for 10 min, 40 cycles of 95 ℃ for 10 s, 60 ℃ for 34 s, and 95 ℃ for 15 s according to the manufacturer’s protocol. Finally, melting curve profiles were assessed and analyzed using the ABI Step one software. The control was used as a reference sample and treatment as measuring samples. The method of 2−△△CT was adopted to analyze the relative changes in gene expression from real-time quantitative PCR experiments [33]. Statistical analyses were performed using the software in Excel and ASCsee. The results were processed statistically with DPS v14.10 statistical software [34]. The analyzed data subjected to analysis of variance (ANOVA), and Duncan’s multiple range test were employed to determine significant differences between treatments at P < 0.05 level. Mean values ± SD are calculated thrice with two independent biological replicates.
Construction of plant expression vector of transgenic tobacco
The coding region of SoCes3 was inserted in the expression vector pCAMBIA1302 controlled by CaMV 35S promoter using NcoI and SpeI restriction enzyme cutting sites. This plasmid was used to transform Agrobacterium tumefaciens strain EHA105 by triparental mating. The transgenic tobacco (Nicotiana tabacum L. cv. Petit Havana) was obtained via the Agrobacterium-mediated leaf disc method following the standard transformation protocol [35]. Plants rooted on hygromycin selection (50 mg/L) and positive (T0) generation transformants were self-pollinated, and the seeds obtained from T0 were analyzed for segregation by germinating on half-strength Murashige and Skoog medium (MS medium) supplemented with hygromycin (25 mg/L). One-month-old plants were transferred to the greenhouse in pots containing the manure, sand, and soil (1:1:2 ratio). Genomic DNAs were extracted to identify the transgenic lines. The target gene was detected using these DNA as templates via qRT-PCR using GFP primers (GFP-F, R) and the SoCes3 primers (CES-F, R). Applied primers are shown in Table 1. Total RNA was extracted from plant tissue; extracted RNA was used for single-strand cDNA synthesis. Real-time reverse transcription-polymerase chain reaction (qRT-PCR) was used to screen tobacco plants with exogenous SoCes3 and GFP gene expression.
Seeds from a single tobacco plant positive for the overexpression of SoCes3 gene were sown. When the seedlings reached the 4th-leaf stage, each plant was planted in plastic container (12X15 cm, diameter and height). The culture substrate uses a mixture of peat soil and vermiculite (1:1). Plants were grown at 26/25 ℃ (light/dark), light intensity of 400 μmol m−2 s−1, photoperiod of 12 h, and relative humidity of approximately 75% in an artificial culture chamber. When the plants reached the 4th-leaf stage, qRT-PCR was applied to screen T1 tobacco plants with exogenous SoCes3 and GFP gene expression (designated as S). Non-transgenic ‘Nicotiana tabacum L.cv. Petit Havana’ seeds were used as wild-type controls (designated as WT). Two lines (SoCes3-40 and SoCes3-45) were selected for further experiments at 8th-leaf stage of plant. Tobacco plants were used to measure physiological and histological characteristics. Transcriptome sequencing was carried out for photosynthetically mature leaves of WT, SoCes3-40, and SoCes3-45 seedlings.
Measurements of physiological and histological characteristics
When the tobacco plants grow, the top 3rd and 5th leaf and leaf veins were sampled from WT, SoCes3-40, and SoCes3-45 at the eight-leaf stage. Cellulose was isolated and solubilized according to Updegraff [36]. Cellulose content was reduced from Glc concentration measured by the anthrone method as described by Scott and Melvin [37] and Turner and Somerville [38]. The lignin content was analyzed by detection kit from Soraibao Biotechnology Co., Ltd. Stems, leaves, and leaf veins from SoCes3-40 and SoCes3-45 plants together with the same-aged wild-type plants were sectioned and stained with fluorescence single-label staining of paraffin sections.
Transcriptome sequencing
High-quality RNA was extracted with RNAprep Pure kit (Tiangen, Beijing, China) according to the manufacturer’s recommendation. Sequencing libraries were analyzed using the MGIEasy RNA Library Prep Kit, following the manufacturer’s protocol. The sample library was sequenced on the Illumina high-throughput sequencing platform. Use Trimmomatic software to collect sequencing data from raw data and remove low-quality sequencing fragments. Finally, differentially expressed genes (DEGs) were screened using the R software package based on the criteria of log2 (fold change) > 1 or < 1 and P < 0.05. Using the GO (http://geneontology.org/) and KEGG database (https://www.genome.jp/kegg/), annotate differentially expressed genes (DEGs), and the annotated DEGs are used to analyze function and signaling pathways further.
Data analysis
Excel and DPSv14.10 analyzed all the data in this study. All column diagrams were drawn using DPSv14.10. Duncan's multiple comparison of three replications was represented. Significant differences between wild-type and transgenic tobacco were used (*P ≤ 0.05) and (**P ≤ 0.01).
Results
Isolation, characterization, and bioinformatics analysis of SoCes3
A full-length of SoCes3 cDNA sequence of 3625 bp was isolated from the sugarcane plant leaves. The entire open reading frame (ORF) was 3225 bp in length, contained most 5’-end excluding 80 bp and the whole 3’-end with poly(A), named as SoCes3 (GenBank accession No. MG324347). Ten other Ces protein sequences with high homology to sugarcane were selected for phylogenetic analysis using DNAMANv5.2.2 software. The un-rooted phylogenetic tree of the SoCes3 gene and different Ces3 sequences was constructed with MEGA 6.0 software. The results revealed that the SoCes3 protein had the highest (99.00%) similarity with other Ces3 proteins of Miscanthus x giganteus (cellulose synthase catalytic subunit 3) and Zea mays (ZmCes3), other Ces3 proteins in 95–99% of Saccharum hybrid cultivar and Phyllostachys edulis, and other CesA proteins in 92–99% of Sorghum bicolor, Setaria italica and Oryza brachyantha (Fig. 1). The SoCes3 sequence presented high level with the Gramineae plants, such as Miscanthus x giganteus Ces3, Saccharum hybrid cultivar Ces, Sorghum bicolor CesA, Zea mays Ces, etc. After that, most of CesA genes have been observed to occur in the form of gene families in the genome of other higher plants.

The phylogenic tree induced amino acid sequence between sugarcane SoCes3 and other Ces3 proteins
Subcellular localization analysis of SoCes3
According to PSORTb version 3.0.2, SoCes3 has 92.6% possibility of locating in the cell membrane. To determine whether SoCes3 found in the cell membrane, the vector containing SoCes3-GFP fusion genes was constructed. The fusion protein expression vector pCAMBIA1302 was constructed by fusing the C-terminus of SoCes3 protein to the green fluorescent protein (GFP), driven by the 35S promoter of Cauliflower Mosaic Virus (CaMV). The fused protein was expressed transiently in Nicotiana benthamiana while the 35S:GFP functioned as a control. As shown in Fig. 2A, the GFP protein was distributed throughout the cell membrane and nucleus with the pCAMBIA1302 vector. However, green fluorescence was specifically detected in the cell membrane transiently with the pCAMBIA-SoCes3 (Fig. 2B), demonstrating that the product of SoCes3 is located in the cell membrane.

Subcellular localization of pCAMBIA1302-SoCes3. A Histochemical analysis of transactivation activity of pCAMBIA1302 plasmids in N. benthamiana leaves. excitation: 488 nm, emission: 625 nm. (a) GFP images, (b) dark-field images and (c) overlapping GFP fluorescence and dark-field images. B Histochemical analysis of transactivation activity of pCAMBIA1302-SoCes3 plasmids in leaves of N. benthamiana. excitation: 488 nm, emission: 625 nm. (d) GFP images, (e) dark-field images and (f) overlap of GFP fluorescence and dark-field images
Expression patterns of SoCes3 gene in different plant parts of sugarcane
Quantitative reverse transcriptase (qRT)-PCR expression analysis of SoCes3 gene was performed from different plant parts, such as leaves, sheaths, and stalks of GT46 and GT49 sugarcane varieties and specific treatment intervals on GT42 variety with different treatments at early growth stage. The expression of SoCes3 showed significant differences in various organs in sugarcane varieties, i.e., GT46 and GT49 (Fig. 3A). SoCes3 was expressed in leaf, sheath, and internode of both sugarcane varieties. The highest expression level was found in the internode of stem, especially with the highest expression level in the fifth internode and lowest in the leaf. Compared with GA treatment, PP333 treatment can enhance the SoCes3 gene transcription level of first leaf of sugarcane, reaching the highest level on the 40th day after treatment. After 60 days of GA treatment, transcription level of SoCes3 gene was found lower as compared to control and PP333 treatments (Fig. 3B). The lowest transcription level of SoCes3 were found in the leaf sheath (1, 3 and 5th leaf position) of control, GA and PP333 treatment conditions (Fig. 3C).

Average expression pattern values of SoCes3 gene in sugarcane (n = 3). A Impact of gene expression levels in leaf sheath and stem internode of GT46 and 49 varieties, B gene expression levels on 1st leaf position of sugarcane during GA and PP333 treatments of GT49 variety on early growth stage at specific time interval and C gene expression levels in different plant organs during GA and PP333 treatments of GT49 variety at 60 days of growth stage
Development and screening assay of transgenic tobacco plants
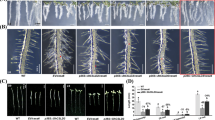
Identified eight hygromycin-resistant transgenic tobacco lines integrated with sugarcane gene (Fig. 4A, B), and their mRNA levels of SoCes3 by qRT-PCR (Fig. 4C). qRT-PCR was used to analyze the transcript expression of SoCes3 in transgenic plants with a single copy of transgenic. The constitutively expressed NtActin was used as the internal standard in expression analysis. Compared with the wild-type tobacco, these transgenic lines showed the increasing expression pattern of SoCes3 gene at different levels. When tobacco plants reached the 4th-leaf stage, SoCes3-33 showed a relatively lower expression level of the SoCes3 gene, the same as WT, while SoCes3-2, 3, 40, 45, 47, 55, 58 showed the highest expression level. For GFP, SoCes3-2, 3, 47, 55, 58 showed a relatively low expression level of SoCes3 gene as same as WT. In contrast, SoCes3-40 showed the highest expression level (Fig. 4C). Based on the transcripts of SoCes3, two lines (SoCes3-40 and SoCes3-45) were selected for the further experiments at 8th-leaf growth stage from top to bottom (Fig. 4A). Third and fifth leaf of plant, SoCes3-40 showed a relatively high expression level of SoCes3 gene than those of SoCes3-45 and WT. Third leaf of SoCes3-45 showed a relatively higher expression level. On the third leaf and vein, and fifth leaf vein of plants, SoCes3-45 showed a relatively higher expression level than SoCes3-40 and WT plants.

Growth and development of transgenic tobacco plants. A Morphological development of transgenic with wild-type tobacco plant (WT) under normal conditions. B The expected fragment of SoCes3 was observed from all transgenic lines except WT. C The expression levels of different transgenic tobacco lines were detected via qRT-PCR. D The expression levels of other leaves and lines 40, 45, and WT leaf veins were detected via qRT-PCR
SoCes3-overexpression of tobacco increases cellulose and lignin contents
The cellulose and lignin contents were found higher in transgenic tobacco leaves and leaf veins. As shown in Fig. 5A, B, the cellulose content increased in the transgenic plant SoCes3-40 of third and fifth leaf, and third leaf, but not in the transgenic plant SoCes3-45. The cellulose content was found higher in 5th leaf vein of WT as compared to SoCes3-40 and SoCes3-45. Lignin content increased in the transgenic plants SoCes3-40 and SoCes3-45 as compared to WT plants (Fig. 5B).

Measurements of cellulose contents (A) and lignin contents (B). Error bars indicate standard error of three biological replicates (n = 3)
Histological observation of SoCes3-overexpression
Figure 6 shows that the histological differences between SoCes3 transgenic tobacco plants and WT plants. It can be seen from the transverse stem sections stained with fluorescence single-label staining of paraffin sections in transgenic tobacco plants. SoCes3 transgenic tobacco has stronger fluorescence signals than the WT and mainly primary cell wall tissues. Cell wall thickening suggests that the cellulose content in transgenic materials can upregulate (Fig. 6A). The expression pattern of SoCes3 in heterologous plant tobacco is similar to that of Arabidopsis AtCESA3 [25, 27, 39], and related to the biosynthesis of primary cell wall.

Immunofluorescence test using GFP antibodies of transgenic tobacco in 3rd leaf vein. A SoCes3 tobacco, B WT tobacco. Red fluorescent signals indicate SoCes3 foci, cell wall, and nucleus indicated by DAPI staining. Bars = 50 μm
Regulation of SoCes3 gene overexpression in tobacco plants
Here, the WT and SoCes3 transgenic lines SoCes3-40 and SoCes3-45 were compared for further analyses. Performed a comparative transcriptomic analysis between WT (K1) and SoCes3 transgenic lines, i.e., SoCes3-40 (A1) and SoCes3-45 (A5) 3rd leaves using high-throughput RNA sequencing (RNA-seq). After background correction and normalization, a total of 2993 and 4481 differentially expressed genes (DEGs) were identified from the leaves of SoCes3-overexpressing transgenic tobacco plants, respectively (Fig. 7A, B).

Numbers of DEGs in various comparisons. A The bar graph presents the number of up- and down-regulated DEGs. B KEGG pathway enrichment analysis. K1: wild type; A1: SoCes3-overexpressing transgenic tobacco lines SoCes3-40; A5: SoCes3-overexpressing transgenic tobacco lines SoCes3-45
GO annotation is mainly divided into three categories, such as biological processes, molecular functions, and cellular composition, and the results of A1 vs K1 and A5 vs K1 showed that the 22 and 17 most significantly enriched subcategories among the three major categories revealed that the cell wall biogenesis and metabolic, and cellulose biosynthetic processes (Fig. 8). Cell wall biogenesis, xyloglucan metabolic process, xyloglucan: xyloglucosyl transferase activity was the most significant biological process of differential gene enrichment. However, all the upregulated genes were selected to elucidate the cell wall biogenesis mechanism (Fig. 9).

Assessment of cellular biosynthetic-related genes induced by SoCes3 gene. Comparison of GO annotation of DEGs between A1-vs-K1 and A5-vs-K1 biological processes (A, B). K1: wild type; A1: SoCes3-overexpressing transgenic tobacco lines SoCes3-40; A5: SoCes3-overexpressing transgenic tobacco lines SoCes3-45

Characterization of upregulated DEGs in transgenic tobacco. A The heatmap of DEGs between A1-vs-K1 and A5-vs-K1. B Top 20 GO functional annotations in biological process corresponding to A. C The heatmap of DEGs between A1-vs-K1 and A5-vs-K1 related to cellular biosynthesis. D Top 20 GO functional annotations in biological process corresponding to C. A, C Heatmap of selected DEGs related to cellular biosynthesis based on TPM or log2FC values. In A, C, the TPM values of DEGs were standardized by log2 (TPM + 1). The dark color, the higher gene expression level. In B, D, the gene ratio represents the percentage of selected genes, and the circle size represents gene numbers. The bigger circle means more gene numbers. The color of circle represents the p-value. The dark color smaller the p-value with higher significant difference. The darker color, the larger gene fold change. K1: wild type; A1: SoCes3-overexpressing transgenic tobacco lines SoCes3-40; A5: SoCes3-overexpressing transgenic tobacco lines SoCes3-45
Discussion
Cellulose is a main component of plant biomass and represents 42–46% (w/w) of sugarcane cell wall components, is responsible for the rigidity of the cell wall, providing the backbone to determine cell structure and maintaining the resistance of turgor pressure. The predominant polysaccharide component in culm cell walls is cellulose [39, 40]. Cellulose accounts for 28–30% of the above-ground dry matter in typical forage grasses [41], 42–45% in wood [42], and in sugarcane and energy cane cultivars [43]. It is composed of UDP-α-D-glucose through the catalysis by cellulose synthase, and degraded by cellulase. Cellulose biosynthesis in various species varies on specific members of the CesA family [44]. As the most abundant reservoir of C in nature, cellulose and other polymers can be prominent competing sinks for C in sugarcane [45]. Cellulose synthesis is catalyzed by enzyme complexes of cellulose synthase (CesA), arranged in rosette formations at the bases of growing cellulose fibrils. The CesA reactions are coordinated with those of SuSy that operate in the degradative direction in supplying UDP-Glu substrates at cellulose synthesis sites [46].
CesA gene families, which belong to the cellulose synthase gene superfamily consisting of more than eight membranes in higher plants [47, 48]. The CesA gene that synthesizes the β-1,4-linked glycan backbone of cellulose and hemicellulose polysaccharides encodes enzymes related to cellulose and hemicellulose polysaccharides [49]. Cellulose in plants are synthesized by multimeric protein complexes, which consist of hexameric, rosette-like structures in the plasma membrane [50, 51]. Cellulose is made of glucose linear polymer obtained from activated sugar donor UDP‐glucose which is available due to sucrose cleavage by sucrose synthase (SuSy) [51,52,53]. The use of glucose by cleaving the stored sucrose in mature internodes may not occur in sugarcane, because other sources, including the depolymerization of the cell wall, improved photosynthate manufacture, or other metabolic pathways are available to provide glucose, so sucrose content is not affected, particularly in internodes of high sucrose sugarcane [54, 55].
All members of the CesA family contain a putative LIM-like Zn-binding domain/RING finger domain in the N-terminal region, which is similar to several putative plant Leucine zipper transcription factors [6, 7, 46]. All CesA gene products contain a D, D, D, QxxRW motif [21]. The amino-terminal domains are two predicted transmembrane domains near positions 270 and 300 in the Arabidopsis CesA proteins. The carboxy-terminal portion of the protein, extending from nearly 850 amino acid positions, contains six additional predicted transmembrane domains [56]. The various members of the plant CesA family range between 985 and 1088 amino acids and can vary in sequence identity from 53 to 98% [56]. The present study identified sugarcane CesA3 of 1074 amino acids in length. The predicted protein contains random coils, α-helices, extended strands, and non-signal peptide structure.
There are more than 1,250 CesA and Csl sequences from 29 different plant species in GenBank. The genes are responsible for three distinct brittle mutations of rice, induced by the insertion of the endogenous retrotransposon Tos17, which correspond to CesA (cellulose synthase catalytic subunit) genes, OsCesA4, OsCesA7, and OsCesA9. Three CesA genes were expressed in seedlings, culms, premature panicles, and roots but not in mature leaves, and the expression profiles were almost identical among the three genes [23]. The Eucalyptus Cellulose Synthase 3 (EgCesA3) gene had its expression pattern evaluated in leaf and xylem tissues among the three most economically important Eucalyptus species, such as E. grandis, E. globulus, and E. urophylla in Brazil [57]. Three of the maize genes, ZmCesA10-12, are associated with secondary wall formation [15].
Among 25 annotated sorghum cellulose synthase and 28 cellulase genes, 5 (20%) and 7 (25%) of them, were upregulated in optimum sugarcane biomass and no down-regulation was monitored [43, 58, 59]. In Arabidopsis thaliana CesA4, 7, and 8 are associated with the deposition of the secondary cell wall [13, 60], while CesA10 and CesA12 are responsible for the same process in Zea mays plants [15], and CesA7 and CesA8 in Oryza sativa. CesA3 is related to the primary cell wall biosynthesis in Zea mays plants [15, 61]. The upregulation of CesA10, 11, 12 in the rind, as compared to vascular parenchyma and vascular bundles of mature internodes of sugarcane [62]. Primary and secondary cell wall biosynthesis is a dynamic process and is active in immature and mature culms of sugarcane [63]. Poplar is a model tree with better genome annotation and contains 18 CesA genes. It is reported that several CesA genes in poplar are involved in cellulose synthesis [64].
Stalk lodging in sugarcane results in significant yield losses. In maize, an increase in cellulose concentration in the wall might permit simultaneous improvements in stalk strength and harvest index [15]. Isolation of the expressed CesA genes from sugarcane and their association with primary or secondary wall formation has allowed testing of their respective roles in cellulose synthesis through mutational genetics or transgenic approaches. This information would be useful in improving stalk strength. Isolation of the expressed CesA genes from maize and their association with primary or secondary wall formation, possible to test their respective roles in cellulose synthesis in different cell types through association genetics, mutational genetics, or a transgenic approach. It would be useful in improving stalk strength in cereals. AtCesA3(IXR1) was highly expressed in stems, flowers, roots, and shoots. AtCesA3(IXR1) appeared to be expressed more in roots than shoots [65]. The expression of 10 Arabidopsis CesA genes in the embryo at three postembryonic stages by semiquantitative reverse transcription (RT)-PCR. All 10 CesA genes were represented at the young plant, stem, and flowering stage, whereas only CesA1, CesA2, CesA3, and CesA9 were significantly expressed in the embryo [66].
Auxin has been shown to promote the expression of a putative cellulose synthase gene (At2g32610) while inhibiting another putative cellulose synthase gene (At2g32530) in Arabidopsis [67]. In response to auxin treatment, the GhCesA4 promoter was inducible in transgenic tobacco plants [68]. Such results were similar to the expression profile of GhCesA4 in the fibers where IAA levels were low in the stage of rapid cell elongation and then accumulated after the fiber entered the secondary thickening phase [69]. The upregulation of multi-CeS complex DEGs in immature internodes is a sign of the abundant requirement of cellulose in actively growing internodes. Some members of the CesA complex were upregulated in the maturing internodes of the mutant clones, suggesting that cellulose synthesis is still turned on, driving the internodes to final maturity [49, 51]. During culm maturation in sugarcane, the cellulose synthesis is regulated through coordinated expression of diverse genes and gene families including those encoding cellulose synthases, cellulose synthase-likes, enzymes for lignin biosynthesis, and a range of other genes identified in clusters from expression profiles of different tissues [63, 70]. CesAs are responsible for cellulose biosynthesis in secondary walls, and in most cases, the genotypes showed increasing expression levels across time points, this likely indicates ample synthesis of secondary cell walls in these young sugarcane culms [61].
In conclusion, transgenic N. tabacum overexpressing the SoCes3 gene was successfully obtained, and transgenic plant analyses are being performed. Although we are unable to define whether the genetic differences in response to aberrant cell walls exist in sugarcane, it is evident that the process of how CesA works in higher plants is very complicated. Various questions regarding the CesA function remain to be answered. However, it is already possible to notice, by empirical view, growth improvement in the transgenic plants compared to the wild type.
Availability of data and materials
All data generated or analyzed during this study are included in this published article.
References
Tew TL, Cobill RM. Genetic Improvement of sugarcane (Saccharum spp.) as an energy crop. In: Vermerris W, editor. Genetic improvement of bioenergy crops. New York: Springer; 2008. p. 273–94. https://doi.org/10.1007/978-0-387-70805-8_9.
Li YR. Modern sugarcane science. Beijing: China Agriculture Press; 2010. p. 60–2.
Taylor NG. Cellulose biosynthesis and deposition in higher plants. New Phytol. 2008;178(2):239–52. https://doi.org/10.1111/j.1469-8137.2008.02385.x.
Delmer DP. (1999) Cellulose biosynthesis: exciting times for a difficult field of study. Ann Rev Plant Physiol Plant Mol Biol. 1999;50(1):245–76. https://doi.org/10.1146/annurev.arplant.50.1.245.
Brown RM Jr, Saxena IM, Kudlicka K. Cellulose biosynthesis in higher plants. Trends Plant Sci. 1996;1(5):149–56. https://doi.org/10.1016/S1360-1385(96)80050-1.
Kawagoe Y, Delmer DP. Pathways and genes involved in cellulose biosynthesis. Genet Eng. 1997;19:63–87. https://doi.org/10.1007/978-1-4615-5925-2_4.
Kawagoe Y, Delmer DP. Recent progress in the field of cellulose synthesis. Trends Glycosci Glycotechnol. 1998;10(54):291–305. https://doi.org/10.4052/tigg.10.291.
Delmer DP, Haigler CH. The regulation of metabolic flux to cellulose, a major sink for carbon in plants. Metab Eng. 2002;4(1):22–8. https://doi.org/10.1006/mben.2001.0206.
Saxena IM, Lin FC, Brown RM. Cloning and sequencing of the cellulose synthase catalytic subunit gene of Acetobacter xylinum. Plant Mol Biol. 1990;15(5):673–83. https://doi.org/10.1007/BF00016118.
Wong HC, Fear AL, Caboon RD, Eichinger GH, Mayer R, Amikam D, Benziman M, Gelfand DH, Measde JH, Emerick AW. Genetic organization of the cellulose synthase operon in Acetobacter xylinum. Proc Nat Acad Sci U S A. 1990;87(20):8130–4. https://doi.org/10.1073/pnas.87.20.8130.
Pear JR, Kawagoe Y, Schreckengost WE, Delmer DP, Stalker DM. Higher plants contain homologs of the bacterial celA genes encoding the catalytic subunit of cellulose synthase. Proc Nat Acad Sci U S A. 1996;93(22):12637–42. https://doi.org/10.1073/pnas.93.22.12637.
Burn JE, Hocart CH, Birch RJ, Cork AC, Williamson RE. Functional analysis of the cellulose synthase genes CesA1, CesA2, and CesA3 in arabidopsis. Plant Physiol. 2002;129(2):797–807. https://doi.org/10.1104/pp.010931.
Taylor NG, Howells RM, Huttly AK, Vickers K, Turner SR. Interactions among three distinct CesA proteins essential for cellulose synthesis. Proc Nat Acad Sci U S A. 2003;100(3):1450–5. https://doi.org/10.1073/pnas.0337628100.
Wang LQ, Guo K, Li Y, Tu YY, Hu HZ, Wang BR, Cui XC, Peng LC. Expression profiling and integrative analysis of the CESA/CSL superfamily in rice. BMC Plant Biol. 2010;10(1):282. https://doi.org/10.1186/1471-2229-10-282.
Appenzeller L, Doblin M, Barreiro R, Wang HY, Niu XM, Kollipara K, Carrigan L, Tomes D, Chapman M, Dhugga KS. Cellulose synthesis in maize: isolation and expression analysis of the cellulose synthase (CesA) gene family. Cellulose. 2004;11(3–4):287–99. https://doi.org/10.1023/B:CELL.0000046417.84715.27.
Sundberg B, Aspeborg H, Nilsson P, Teeri TT, Mellerowicz E, Djerbi S, Blomqvist K. Identification and expression analysis of genes encoding putative cellulose synthases (CesA) in the hybrid aspen, Populus tremula (L. × P. tremuloides (Michx.). Cellulose. 2004;11:301–12. https://doi.org/10.1023/B:CELL.0000046408.84510.06.
Kalluri UC, Joshi CP. Isolation and characterization of a new, full-length cellulose synthase cDNA PtrCesA5 from developing xylem of aspen trees. J Exp Bot. 2003;54(390):2187–8. https://doi.org/10.1093/jxb/erg232.
Djerbi S, Lindskog M, Arvestad L, Sterky F, Teeri TT. The genome sequence of black cottonwood (Populus trichocarpa) reveals 18 conserved cellulose synthase (CesA) genes. Planta. 2005;221(5):739–46. https://doi.org/10.1007/s00425-005-1498-4.
Takata N, Taniguchi T. Expression divergence of cellulose synthase (CesA) genes after a recent whole genome duplication event in Populus. Planta. 2015;241(1):29–42. https://doi.org/10.1007/s00425-014-2217-9.
Tian ZJ, Yi R, Chen JR, Guo QQ, Zhang XW. Cloning and expression of cellulose synthase gene in ramie [Boehme-rianivea (Linn.) Gaud.]. Acta Agron Sin. 2008;34(1):76–83. https://doi.org/10.3724/sp.j.1006.2008.00076.
Richmond TA, Somerville CR. The cellulose synthase superfamily. Plant Physiol. 2000;124(2):495–8. https://doi.org/10.1104/pp.124.2.495.
Holland N, Holland D, Helentjaris T, Dhugga KS, Xoconostle-Cazares B, Delmer DP. A comparative analysis of the plant cellulose synthase (CesA) gene family. Plant Physiol. 2000;123(4):1313–24. https://doi.org/10.1104/pp.123.4.1313.
Tanaka K, Murata K, Yamazaki M, Onosato K, Miyao A, Hirochika H. Three distinct rice cellulose synthase catalytic subunit genes required for cellulose synthesis in the secondary wall. Plant Physiol. 2003;133(1):73–83. https://doi.org/10.1104/pp.103.022442.
Burton RA, Shirley NJ, King BJ, Harvey AJ, Fincher GB. The CesA gene family of barley. Quantitative analysis of transcripts reveals two groups of Co-expressed genes. Plant Physiol. 2004;134(1):224–36. https://doi.org/10.1104/pp.103.032904.
Desprez T, Juraniec M, Crowell EF, Jouy H, Pochylova Z, Parcy F, Höfte H, Gonneau M, Vernhettes S. Organization of cellulose synthase complexes involved in primary cell wall synthesis in Arabidopsis thaliana. Proc Nat Acad Sci U S A. 2007;104(39):15572–7. https://doi.org/10.1073/pnas.0706569104.
Taylor NG, Scheible WR, Cutler S, Somerville CR, Turner SR. The irregular xylem3 locus of Arabidopsis encodes a cellulose synthase required for secondary cell wall synthesis. Plant Cell. 1999;11(5):769–80. https://doi.org/10.1105/tpc.11.5.769.
Persson S, Paredez A, Carroll A, Palsdottir H, Doblin M, Poindexter P, Khitrov N, Auer M, Somerville CR. Genetic evidence for three unique components in primary cell-wall cellulose synthase complexes in Arabidopsis. Proc Nat Acad Sci U S A. 2007;104(39):15566–71. https://doi.org/10.1073/pnas.0706592104.
Atanassov II, Pitman JK, Turner SR. Elucidating the mechanisms of assembly and subunit interaction of the cellulose synthase complex of Arabidopsis secondary cell walls. J Biol Chem. 2009;284(6):3833–41. https://doi.org/10.1074/jbc.M807456200.
Hill JL, Hummudi MB, Tien M. The Arabidopsis cellulose synthase complex: a proposed hexamer of CESA trimers in an equimolar stoichiometry. Plant Cell. 2014;26(12):4834–42. https://doi.org/10.1105/tpc.114.131193.
Chen R, Fan Y, Yan H, Zhou H, Zhou Z, Weng M, Huang X, Lakshmanan P, Li YR, Qiu L, Wu JM. Enhanced activity of genes associated with photosynthesis, phytohormone metabolism and cell wall synthesis is involved in gibberellin-mediated sugarcane internode growth. Front Genetics. 2020;11:1–13. https://doi.org/10.3389/fgene.2020.570094.
Kumar S, Tamura K, Jakobsen IB, Nei M. MEGA2: molecular evolutionary genetics analysis software. Bioinformatics. 2001;17(12):1244–5. https://doi.org/10.1093/bioinformatics/17.12.1244.
Saitou NM, Nei M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol. 1987;24(4):189–204. https://doi.org/10.1093/oxfordjournals.molbev.a040454.
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2-△△Ct method. Methods. 2000;25(4):402–8. https://doi.org/10.1006/meth.2001.
Tang QY, Zhang CX. Data Processing System (DPS) software with experimental design, statistical analysis and data mining developed for use in entomological research. Insect Sci. 2013;20(2):254–60. https://doi.org/10.1111/j.1744-7917.2012.01519.x.
Burow MD, Chlan CA, Sen P, Lisca A, Murai N. High-frequency generation of transgenic tobacco plants after modified leaf disk cocultivation with Agrobacterium tumefaciens. Plant Mol Biol Rep. 1990;8(2):124–39. https://doi.org/10.1007/BF02669766.
Updegraff DM. Semi-micro determination of cellulose in biological materials. Anal Biochem. 1969;32(3):420–4. https://doi.org/10.1016/S0003-2697(69)80009-6.
Scott TA, Melvin EH. Determination of dextran with anthrone. Anal Chem. 1953;25(11):1656–61. https://doi.org/10.1021/ac60083a023.
Turner SR, Somerville CR. Collapsed xylem phenotype of Arabidopsis identifies mutants deficient in cellulose deposition in the secondary cell wall. Plant Cell. 1997;9(5):689–701. https://doi.org/10.1105/tpc.9.5.689.
Lingle SE, Tew TL, Hale A, Cobill R. Cell wall composition of sugarcane and related Saccharum species. In: 30th Symposium on Biotechnology for Fuels and Chemicals, New Orleans, USA, abstract 1–24. 2008. http://sim.confex.com/sim/30th/techprogram/P5488.HTM.
Sainz MB. Commercial cellulosic ethanol: the role of plant-expressed enzymes. In Vitro Cell Dev Biol Plant. 2009;45:314–29. https://doi.org/10.1007/s11627-009-9210-1.
Theander O, Westerlund E. Quantitative analysis of cell wall components. In: Jung HG, Buxton DR, Hatfield RD, Ralph J, editors. Forage cell wall structure and digestibility. Madison: American Society of Agronomy/CSSA/SSSA; 1993. p. 83–104.
Smook GA. Handbook for pulp and paper technologists. Vancouver: Angus Wilde Publications; 1992.
Kim M, Day DF. Composition of sugar cane, energy cane, and sweet sorghum suitable for ethanol production at Louisiana sugar mills. J Ind Microbiol Biotechnol. 2011;38(7):803–7.
Gu Y, Somerville C. Cellulose synthase interacting protein: a new factor in cellulose synthesis. Plant Signal Behav. 2010;5:1571–4. https://doi.org/10.4161/psb.5.12.13621.
Wang J, Nayak S, Koch K, Ming R. Carbon partitioning in sugarcane (Saccharum species). Front Plant Sci. 2013;4:201. https://doi.org/10.3389/fpls.2013.00201.
Arioli T, Peng L, Betzner AS, Burn J, Wittke W, Herth W, Camilleri C, Höfte H, Plazinski J, Birch R, Cork A, Glover J, Redmond J, Williamson RE. Molecular analysis of cellulose biosynthesis in Arabidopsis. Sci. 1998;279(5351):717–20. https://doi.org/10.1126/science.279.5351.717.
Kaur S, Dhugga KS, Gill K, Singh J. Novel structural and functional motifs in cellulose synthase (CesA) genes of bread wheat (Triticum aestivum L.). PLoS ONE. 2016;11(1):e0147046. https://doi.org/10.1371/journal.pone.0147046.
Liu X, Zhang H, Zhang W, Xu W, Li S, Chen X, Chen H. 2022 Genome-wide bioinformatics analysis of cellulose synthase gene family in common bean (Phaseolus vulgaris L.) and the expression in the pod development. BMC Genom Data. 2022;23:9. https://doi.org/10.1186/s12863-022-01026-0.
Daras G, Templalexis D, Avgeri F, Tsitsekian D, Karamanou K, Rigas S. Updating insights into the catalytic domain properties of plant cellulose synthase (CesA) and cellulose synthase-like (Csl) Proteins. Molecules. 2021;26:4335. https://doi.org/10.3390/molecules26144335.
McFarlane HE, Doring A, Persson S. The cell biology of cellulose synthesis. Annu Rev Plant Biol. 2014;65:69–94.
Khan Q, Qin Y, Guo DJ, Chen JY, Zeng XP, Mahmood A, Yang LT, Liang Q, Song XP, Xing YX, Li YR. Sucrose metabolism analysis in a high sucrose sugarcane mutant clone at a mature stage in contrast to low sucrose parental clone through the transcriptomic approach. Chem Biol Technol Agric. 2023;10:39. https://doi.org/10.1186/s40538-023-00408-5.
Coleman HD, Ellis DD, Gilbert M, Mansfield SD. Up-regulation of sucrose synthase and UDP-glucose pyrophosphorylase impacts plant growth and metabolism. Plant Biotechnol J. 2006;4(1):87–101.
Coleman HD, Beamish L, Reid A, Park J-Y, Mansfield SD. Altered sucrose metabolism impacts plant biomass production and flower development. Transgenic Res. 2010;19(2):269–83.
Liu J, Osbourn A, Ma P. MYB transcription factors as regulators of phenylpropanoid metabolism in plants. Mol Plant. 2015;8(5):689–708.
McCarthy RL, Zhong R, Ye Z-H. MYB83 is a direct target of SND1 and acts redundantly with MYB46 in the regulation of secondary cell wall biosynthesis in Arabidopsis. Plant Cell Physiol. 2009;50(11):1950–64.
Richmond TA. Higher plant cellulose synthases. Genome Biol. 2000;1(4):1–5. https://doi.org/10.1186/gb-2000-1-4-reviews3001.
Marques WL, Salazar M, Camargo E, Lepikson-Neto J, Gonçalves DC, Do Nascimento LC, Garcia C, Almeida A, Pereira G. Transgenic Arabidopsis thaliana and Nicotiana tabacum overexpressing the Eucalyptus grandis cellulose synthase 3 and its expression pattern in different Eucalyptus species and tissues. BMC Proc. 2011;5(l7):167–9. https://doi.org/10.1186/1753-6561-5-S7-P167.
Wai CM, Zhang J, Jones TC, Nagai C, Ming R. Cell wall metabolism and hexose allocation contribute to biomass accumulation in high yielding extreme segregants of a Saccharum interspecific F2 population. BMC Genomics. 2017;18:773. https://doi.org/10.1186/s12864-017-4158-8.
Kasirajan L, Hoang NV, Furtado A, Botha FC, Henry RJ. Transcriptome analysis highlights key differentially expressed genes involved in cellulose and lignin biosynthesis of sugarcane genotypes varying in fiber content. Sci Rep. 2018;8:11612. https://doi.org/10.1038/s41598-018-30033-4.
Hernandez-Blanco C, Feng DX, Hu J, Sánchez-Vallet A, Deslandes L, Llorente F, et al. Impairment of cellulose synthases required for Arabidopsis secondary cell wall formation enhances disease resistance. Plant Cell. 2007;19:890–903. https://doi.org/10.1105/tpc.106.048058.
Hosaka GK, Correr FH, Sforça DA, Barreto FZ, Almeida Balsalobre TW, Pasha A, Provart NJ, Carneiro MS, Alves Margarido GR. Temporal gene expression in apical culms shows early changes in cell wall biosynthesis genes in sugarcane. Front Plant Sci. 2021. https://doi.org/10.3389/fpls.2021.736797.
Casu RE, Rae AL, Nielsen JM, Perroux JM, Bonnett GD, Manners JM. Tissue-specific transcriptome analysis within the maturing sugarcane stalk reveals spatial regulation in the expression of cellulose synthase and sucrose transporter gene families. Plant Mol Biol. 2015;89:607–28. https://doi.org/10.1007/s11103-015-0388-9.
Casu RE, Jarmey JM, Bonnett GD, Manners JM. Identification of transcripts associated with cell wall metabolism and development in the stem of sugarcane by Affymetrix GeneChip Sugarcane Genome Array expression profiling. Funct Integr Genomics. 2007;7:153–67. https://doi.org/10.1007/s10142-006-0038-z.
Zhan N, Shang X, Wang Z, Xie Y, Liu G, Wu Z. Screening cellulose synthesis related genes of EgrEXP and EgrHEX in Eucalyptus grandis. Gene. 2022;824:146396. https://doi.org/10.1016/j.gene.2022.146396.
Scheible WR, Eshed R, Richmond T, Delmer D, Somerville C. Modifications of cellulose synthase confer resistance to isoxaben and thiazolidinone herbicides in Arabidopsis Ixr1 mutants. Proc Nat Acad Sci U S A. 2001;98(18):10079–84. https://doi.org/10.1073/pnas.191361598.
Beeckman T, Przemeck GKH, Stamatiou G, Lau R, Terryn N, de Rycke R, Inzé D, Berleth T. Genetic complexity of cellulose synthase a gene function in Arabidopsis embryogenesis. Plant Physiol. 2002;130(4):1883–93. https://doi.org/10.1104/pp.102.010603.
Goda H, Sawa S, Asami T, Fujioka S, Shimada Y, Yoshida S. Comprehensive comparison of auxin-regulated and brassinosteroid-regulated genes in Arabidopsis. Plant Physiol. 2004;134(4):1555–73. https://doi.org/10.1104/pp.103.034736.
Wu AM, Hu JS, Liu JY. Functional analysis of a cotton cellulose synthase A4 gene promoter in transgenic tobacco plants. Plant Cell Rep. 2009;28(10):1539–48. https://doi.org/10.1007/s00299-009-0753-8.
Gokani SJ, Thaker VS. Physiological and biochemical changes associated with cotton fiber development. Field Crops Res. 2002;77(2):127–36. https://doi.org/10.1016/S0378-4290(02)00062-X.
Hu R, Xu Y, Yu C, He K, Tang Q, Jia C, He G, Wang X, Kong Y, Zhou G. Transcriptome analysis of genes involved in secondary cell wall biosynthesis in developing internodes of Miscanthus lutarioriparius. Sci Rep. 2017;7(1):1–16. https://doi.org/10.1038/s41598-017-08690-8.
Acknowledgements
The authors would like to thank the Guangxi Academy of Agricultural Sciences, Nanning, Guangxi, China, for providing the necessary facilities for this study.
Funding
This work was supported by the Natural Science Foundation of China (32060468, 31400281), Guangxi Natural Science Foundation (2022GXNSFAA035444, 2023GXNSFAA026482, 2023GXNSFAA02646) and Fundamental Research Fund of Guangxi Academy of Agriculture Sciences (2023YM55, 2021YT118, 2017YM35, 2015JM24, 2016JM05).
Author information
Authors and Affiliations
Contributions
Conceptualization: C.M.H., K.C.W., Y.W.W., and L.X.; writing—original draft: C.M.H., K.C.W, and K.K.V.; resources: H.B.L., Z.N.D., G.Y.S., H.Q.C., X.J.W., and L.P.Y.; writing—review and editing: Y.W.W., and L.X.; supervision: Y.W.W., and L.X.; project administration: K.C.W., Y.W.W., and L.X.; funding acquisition: K.C.W., Y.W.W., and L.X. All authors have read and agreed to the published version of the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Huang, CM., Wu, KC., Verma, K.K. et al. Isolation and expression analysis of cellulose synthase 3 (Ces3) genes from sugarcane (Saccharum officinarum L.). Chem. Biol. Technol. Agric. 11, 71 (2024). https://doi.org/10.1186/s40538-024-00597-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40538-024-00597-7