
Abstract
Plant growth regulators are chemical substances which govern all the factors of development and growth within plants. The application of plant growth regulators to crops modifies hormonal balance and growth leading to increased yield, enhanced crop tolerance against abiotic stress and improved physiological trait of crops. Paclobutrazol (PBZ) [(2RS, 3RS)-1-(4-chlorophenyl)- 4, 4-dimethyl-2-(1H-1, 2, 4-trizol-1-yl)-pentan-3-ol], is one of the members of triazole family having growth regulating property. The growth regulating properties of PBZ are mediated by changes in the levels of important plant hormones including the gibberellins (GAs), abscisic acid (ABA) and cytokinins (CK). PBZ affects the isoprenoid pathway, and alters the levels of plant hormones by inhibiting gibberellin synthesis and increasing cytokinins level and consequent reduction in stem elongation. When gibberellins synthesis is inhibited, more precursors in the terpenoid pathway accumulate and that resulted in the production of abscisic acid. PBZ is more effective when applied to the growing media and application on the growing medium would give longer absorption time and more absorption of active ingredient than foliar spray. The application of PBZ to crops is important in reducing plant height to prevent lodging and in increasing number and weight of fruits per tree, in improving the fruit quality in terms of increases in carbohydrates, TSS, TSS/TA and decreases acidity. It further reduces evapo-transpiration and decreases plant moisture stress by enhancing the relative water content of leaf area and develops resistance in the plants against biotic and abiotic stresses. In addition, it acts as highly active systemic fungicide and used against several economically important fungal diseases. In this review, the current knowledge and possible applications of PBZ, which can be used to improve the growth, yield and quality of crops, have been reviewed and discussed. The role of PBZ to mitigate the harmful effects of environmental stresses in crops is also examined. Moreover, various biochemical and physiological processes leading to improved crop production under the effect of PBZ are discoursed in detail.

Similar content being viewed by others


Background
Plant growth regulators are organic substances produced naturally in higher plants, controlling growth or other physiological functions at a site remote from its place of production and active in minute amounts [1]. They play a role in affecting growth, yield and quality of crops [2, 3]. Besides, they are important in stress protection [4].
Triazole compounds are systemic fungicides having plant growth regulating properties and are called as stress protectants, because of their innate ability to induce abiotic stress tolerance by increasing antioxidant enzymes and molecules in stress-affected plants [5]. The plant growth regulating properties of triazoles are mediated by their ability to alter the balance of important plant hormones including Gibberellic acid (GA), Abscisic acid (ABA) and Cytokinins [6]. They induce a variety of morphological and biochemical responses in plants; inhibited shoot elongation, stimulated root growth, increased cytokinin synthesis and a transient rise in ABA, as well as conferring protection from various environmental stresses [7, 8].
PBZ (Bonzi), a triazole family having growth regulating property, is an extremely active chemical and affects almost all plant species, whether applied as a spray or a soil drench [9]. PBZ inhibits GA biosynthesis by blocking the oxidation of ent-kaurene [10]. It is applied to plants in the floricultural industry to control their size and quality [11]. It is applied to perennials and other pot crops at rates of 1–90 mg L−1 [9]. When applied as a foliar spray, PBZ is absorbed by petioles and stems and is translocated through the xylem to the growing tip. When applied as a soil drench, it is taken up through the roots and then translocated through the xylem to the apical meristems [12]. Soil drenches with PBZ may be more effective than the foliar sprays due to increase activity and less probability of stunting and flowering delay, due to no direct contact with flowers or flower buds [11]. Depending on plant species, PBZ can delay or promote flowering. PBZ half-life in the soil varies between 6 and 12 months depending upon the soil type and environmental conditions [13]. Effectiveness of drenches is reduced if the crop is grown in a bark medium, because the chemical will adsorb to the bark and less will be available in the medium solution for the plant to absorb. Phytotoxicity symptoms are not common when applied to perennials, but care must be taken with those species that are known to be sensitive. Therefore, the aim of this review is to summarize the evidence on the biochemical and physiological responses of PBZ as a growth regulator and as a stress protectant.
Paclobutrazol
Plant growth retardants are compounds which are used to reduce plant growth without changing developmental patterns or being phytotoxic [14]. The largest group of plant growth retardants consists of chemicals antagonistic to gibberellins (GA), the hormone that is responsible for plant growth [7]. Commercially used inhibitors of GA biosynthesis are: (a) onium-type compounds, (b) compounds with a N-heterocycle (triazole-type), (c) structural mimics of 2-oxoglutaric acid, and (d) 16, 17-dihydroGAs [14].
PBZ, a member of triazole plant growth regulator group, is used widely in agriculture [15]. It is a cell elongation and internode extension inhibitor that retards plant growth by inhibition of gibberellins biosynthesis. Gibberellins stimulate cell elongation. When gibberellin production is inhibited, cell division still occurs, but the new cells do not elongate. The result is shoots with the same numbers of leaves and internodes compressed into a shorter length. Reduced growth in the diameter of the trunk and branches has also been observed. Another response of trees to treatment with PBZ is increased production of the hormone abscisic acid and the chlorophyll component phytol, both beneficial to tree growth and health. PBZ may also induce morphological modifications of leaves, such as smaller stomatal pores, thicker leaves, and increased number and size of surface appendages, and increased root density that may provide improved environmental stress tolerance and disease resistance [16]. PBZ also has some fungicidal activity due to its capacity as a triazole to inhibit sterol biosynthesis [16].
Chemistry
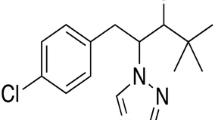
PBZ ([(2R, 3R + 2S, 3S)-1-(4-chloro-phenyl) 4,4-dimethyl-2-(1,2,4-triazol-1-yl)-pentan-3-ol]) has been developed as a plant growth regulator and is registered with trade names such as Bonzi, Clipper, Cultar, and Parsley. It belongs to the triazole compounds that are characterized by a ring structure containing three nitrogen atoms, chlorophenyl and carbon side chains [17]. Structurally, PBZ is a substituted triazole with two asymmetric carbon atoms and is produced as a mixture of 2R, 3R, and 2R, 3R, and 2S, 3S enantiomers [18] (Fig. 1).

The structure of paclobutrazol
Mode of action
Although the precise features of the molecular structure which confer plant growth regulatory activities are not well understood, it appears to be related to the stereochemical arrangement of the substituents on the carbon chain [19]. There are indications that enantiomers having S configuration at the chiral carbon bearing the hydroxyl group are inhibitors of GA biosynthesis. One of the inhibitor of GA biosynthesis, paclobutrazol, is mainly used as growth retardant and stress protectant [20]. This retardation of growth is due to the interference of PBZ with gibberellin biosynthesis by inhibiting the oxidation of ent-kaurene to ent-kauronoic acid through inactivating cytochrome P450-dependent oxygenase [21, 22]. In addition, it tends to be much more effective than various other plant growth regulators at relatively low rate of applications [22].
PBZ is also known to affect the synthesis of the hormone abscisic acid and phytol. Abscisic acid is also synthesized via the terpenoid pathway (Fig. 2). When gibberellins synthesis is blocked, more precursors in the terpenoid pathway are accumulated and shunted to promote the genesis of abscisic acid [23]. It has also been reported to inhibit normal catabolism of ABA [24]. The effect of PBZ on both the synthesis and catabolism processes leads to enhanced concentrations of ABA in leaves. One of the major roles of ABA is to cause closing of stomatal aperture and decreasing loss of water from leaves through transpiration. Improvement of water relations in treated plants takes place because of enhancement in ABA content that decreases stomatal aperture, decreases shoot growth and causing less surface area for transpiration, more roots for uptake of water, and anatomical alterations in leaves that impart barriers to water loss.

Terpenoid pathway for biosynthesis of gibberellins, abscisic acid, phytol, and steroids, and path for degradation of abscisic acid. Steps blocked by paclobutrazol indicated with Geranyl diphosphate synthase (GPS), Farnesyl diphosphate synthase (FPS), Geranyl geranyl diphosphate synthase (GGPS), ent-copalyl-diphosphate synthase (CPS), ent-kaurene synthase (KS), ent-kaurene oxidase (KO), ent-kaurenoic acid oxidase (KAO), Geranyl geranyl reductase (GGRS), Chlorophyll synthase (CHL) and Phytoene synthase (PSY) are the enzymes involved in the terpenoid pathway. ABA 8′-hydroxylase (ABA 8′OH) involved in the enzymatic degradation of ABA into Phaseic acid. KO, KAO and ABA 8′OH are the enzymes inhibited upon PBZ application
Translocation and chemical stability
It was previously believed that triazoles were primarily transported acropetally in the xylem [25]. However, PBZ has been detected in xylem and phloem sap of castor bean [26] and pear [27] indicating that triazoles can be transported acropetally and basipetally. Although the metabolic fate of applied has not been investigated in detail most of them have a high chemical stability [28] and depending on the site of application tend to be metabolized slowly [15]. Early and Martin [29] observed more rapid PBZ metabolism in apple leaves than other plant parts, while Sterrett [30] found little evidence for PBZ metabolism in apple seedlings. PBZ is comparatively more resistant to degradation than BAS 111 [31].
Methods of application
The most common application methods of PBZ are foliar sprays and media drench. PBZ shows good results for both methods [32]; however, drenches act longer and provide uniform control of plant height with lower doses [33]. When PBZ is applied by foliar spray, the compound is poorly soluble in water and consequently little translocated in the phloem. Thus, when applied by spray to the plant canopy, its action is restricted to the wet contact area [34]. On the other hand, the application of PBZ by drench is uniform and increases the product efficiency in lower concentrations compared to foliar spray. Moreover, drench application of PBZ may directly inhibit GA synthesis as roots synthesize large quantities of GA [35]. Similarly, Banon et al. [36] and AlKhassawneh et al. [37] demonstrated that drench applications were more effective, allowing to use lower quantity of PBZ, which is desirable for both ecological and economic reasons. This effectiveness may be directly related to its high persistence in the soil drench [38] and in plant organs [39, 40]. Gent and McAvoy [41] also indicated that PBZ persists in annuals, herbaceous perennials and, especially, woody ornamentals.
PBZ is considered a phloem immobile chemical [7], though some direct [26] and indirect [42] evidence exists that it is partially mobile in phloem. Studies indicate that PBZ and uniconazole-P move in plants acropetally via the xylem, accumulate in leaves, and have very low mobility in phloem [14]. This results in a low level of PBZ residues in seeds and fruits as they are supplied with nutrients via the phloem [25]. However, low phloem mobility of PBZ further reduces the effectiveness of foliar spraying, since PBZ action on plant growth would be restricted to the site of application.
Application rates
A lot has been done to identify the best application rate of PBZ in different places. Factors like age of the trees, extent of vegetative growth and method of application should be considered when determining the rate of PBZ to be applied. The rates also affect the different tree parameters variously. In general, the amount of PBZ required to promote flowering and fruiting in fruit crops is very low [27].
The rate of soil application is a function of tree size and cultivar. The rate is determined by multiplying the diameter of tree canopy in meters by 1–1.5 g of active ingredients of PBZ [43]. They indicated that other factors including soil type, irrigation system, etc. may affect PBZ activity and, thus, may be necessary to improve the effectiveness of the chemical. As to them, overdose may cause undesirable effects such as restricted growth, panicle malformation (too compact), and shoot deformity. They also asserted that to insure uniform flowering and reduce the detrimental side effects, the search for better application methods were investigated and one approach is to apply high volume of low PBZ concentration to improve better coverage.
Optimizing PBZ dose is a prerequisite for any yield improvement programmes. Severe and undesirable loss in seed and oil yield of Camelina was observed when the plants were treated with higher PBZ concentration (125 mg L−1), while PBZ dose between 75 mg L−1 and 100 mg L−1 can effectively improve the economic traits, including higher seed and oil yields in Camelina. Severe retardation of Camelina growth was also reflected in plant height, branch and canopy size when the plants were sprayed with higher PBZ concentration (125 mg L−1) [44]. He also reported that Camelina seed yield increased by 74.23% when compared to the control with the applications of 100 g L−1. Similarly, reduced yields were recorded in peanut [45] and Jatropha [46] associated with higher PBZ concentrations.
Kamran et al. [47] described that soaking of seeds under 300 mg L−1 PBZ increased the average maize grain yield by 61.3% as compared to the control. Patil and Talathi [48] also reported that application of 5 g of PBZ through soil enabled to induce early and regular fruiting with 2.8 times increase in yield in mango var. Alphonso. In addition, PBZ at a rate of 150 mg L−1 in bottle gourd, 100 mg L−1 in bitter gourd, 150 mg L−1 in French bean, 125 mg L−1 in cucumber and 40 mg L−1 in tomato increased the yield and quality of fruits [49].
Response of plants to PBZ
Plant hormone biosynthesis
Gibberellin: Gibberellins (GAs) are a large family of tetracyclic diterpenoid plant growth regulators. Since its original discovery, > 130 GAs have been identified in plants, fungi and bacteria, although only a few GAs have biological activity [50]; many non-bioactive GAs exist in plants, and these act as precursors for the bioactive forms or are de-activated metabolites. Gibberellins (GAs) are plant hormones that are essential for many developmental processes in plants, including seed germination, stem elongation, leaf expansion, trichome development, pollen maturation and the induction of flowering [51]. The major bioactive GAs, which includes GA1, GA3, GA4 and GA7, are derived from a basic diterpenoid carboxylic acid skeleton, and commonly has a C3 hydroxyl group [50].
Triazole compounds are antagonistic to gibberellins and auxins, reducing cell elongation and cell division by inhibiting GA3 biosynthesis [52]. They exhibit varying degrees of both plant growth and fungicidal activity. The intensity of their biological activity is dependent on their isomeric form [17]. The growth retarding property of PBZ is largely attributed to interference with gibberellins biosynthesis. Gibberellins are synthesized from mevalonic acid via the isoprenoid pathway, and the PBZ specifically inhibits the oxidation of ent-kaurene to ent-kaurenoic acid through inactivating cytochrome P-450-dependent oxygenases [18]. Furthermore, PBZ-induced growth inhibition can be reversed by exogenous application of gibberellins [53]. These observations support the hypothesis that growth inhibition due to PBZ is primarily due to reduced gibberellins biosynthesis.
Abscisic acid: The effect of PBZ on ABA is of interest because ABA, like the gibberellins, is synthesized via the isoprenoid pathway, and the two compounds often exhibit opposing physiological activities. The action of PBZ on ABA could be the source of stress protection that has been observed with PBZ [19]. ABA is a natural plant growth regulator that has been implicated in plant acclimation and protection against environmental stress. Exogenous application of ABA has been shown to increase plant resistance to salinity, ozone, heat, chilling and freezing [54].
Mackay et al. [54] demonstrated that PBZ induced stress resistance and it also increased the endogenous concentrations of ABA in snap beans. Hauser et al. [55] also demonstrated that PBZ considerably increased endogenous ABA levels in detached leaves and hydroponically grown seedlings of oilseed rape. ABA accumulated in proportion to PBZ concentration. Mackay et al. [54] also hypothesized that stress protection inferred by PBZ may in part be the result of their effect on endogenous concentrations of ABA. However, both experiments showed that increases in ABA were short lived and eventually decreased to normal or below control levels. Hauser et al. [55] hypothesized that this may be due to stimulated ABA catabolism and/or by an inhibition of its biosynthesis. Therefore, providing a continuous supply, over the growing season, of the PBZ could help to maintain higher levels of endogenous ABA and thereby prolong its stress-protecting effects. In addition, Aly and Latif [56] also reported that PBZ increased the endogenous level of ABA in wheat.
Cytokinin: Cytokinins are synthesized in the roots and translocated acropetally to the shoots where they regulate both plant development and senescence [57]. They are involved in the control of various plant developmental processes such as cell division, apical dominance, stomatal behavior, root formation, leaf senescence, and chloroplast development [58].
Zhu et al. [21] observed an increase in the endogenous cytokinin (Zeatin) level in xylem sap of young apple trees in response to PBZ treatment. PBZ treatment delayed the onset of senescence in grapevine [59] and blueberry [60]. It has been reported that cytokinin or chemicals like thidiazuron with cytokinin-like activity stimulate chlorophyll synthesis and retard senescence [61] and thus, PBZ-induced physiological responses may be associated with increased cytokinin synthesis or prevention of its degradation.
Fletcher et al. [7] also proposed that triazoles stimulate cytokinin synthesis and that enhance chloroplast differentiation, chlorophyll biosynthesis and prevent chlorophyll degradation. An increased level of cytokinins and polyamines over the senescence-promoting hormones ABA and ethylene was reported in plants treated with PBZ. PBZ delayed senescence and extended period of ‘stay-green’ in Camelina sativa [44] by enhancing endogenous levels of cytokinins and that promoted chlorophyll formation and increased activity of certain antioxidant enzymes. A longer ‘staygreen’ character simultaneously increased the period of leaf photosynthesis in PBZ-applied plants by keeping the leaves photosynthetically efficient for a longer time which in turn enhanced the plant productivity of Camelina [44].
Stress protection
Biochemical effects of the triazole include detoxification of active oxygen species, increased contents of antioxidants and chlorophyll (Chl) [22]. More recently, it was found that triazole compounds have been reported to protect plants from various environmental stresses, including chilling, drought, heat, waterlogging, air pollutants, and heavy metals [7, 62]. The triazole-mediated stress protection is often explained in terms of hormonal changes such as an increase in cytokinins, a transient rise in ABA and a decrease in ethylene [19, 54]. Enhanced chilling tolerance in triazole-treated tomato [64] was associated with increased antioxidant enzyme concentrations. In treated tomatoes, apart from the increase in the antioxidants a-tocopherol and ascorbate, free fatty acids were higher and there was a reduction in the loss of membrane phospholipids, as compared to the untreated controls. PBZ prevents the decline in total chlorophyll content in corn plants after exposure to chilling temperatures [64]. PBZ-induced tolerance to low temperature stress has been associated with increased levels of endogenous ABA [7], which has been reported to trigger the genetic processes for hardening. In field studies, winter survival of peas and cereal crops [25] and resistance to frost damage in corn were enhanced by PBZ.
PBZ increases the survival rate of plants under drought conditions through a number of physiological responses. A reduction in the rate of transpiration (due to reduction in leaf area), increased diffusive resistance, alleviating reduction in water potential, increased relative water content, less water use, and increased anti-oxidant activity are some of the reported responses [21, 65]. PBZ also protects plants from high-temperature-induced injuries [64, 65]. Protection against high temperature stress is accompanied by the production of low molecular mass stress proteins [66] and the increase in the activity of antioxidant enzymes [64].
It has also been reported that several environmental factors such as drought, low and high temperature can cause an excess of toxic oxygen-free radicals [67]. Some of the free radical scavenging enzymes are reported to increase in wheat [65] and corn [64] plants after PBZ treatments and their activities are conserved even after exposure to extreme temperature. The triazole compounds enhance the free radical scavenging capacity of treated plants including the levels of carotenoids, ascorbate, superoxide dismutase and ascorbate peroxidase [63]. Berova et al. [68] suggested that the protection caused by PBZ was due to a similar mechanism of enhanced free-radical scavenging systems.
Assimilate partitioning
Assimilate partitioning to the different sinks may be controlled by environmentally regulated, hormonal balances [69]. PBZ treatment increased the root-to-shoot ratio [64], increased partitioning of assimilates to economically important plant parts such as bulbs [70, 71], potato tubers [72, 73], carrot root [8] and rice grain yield [74]. The mechanism of tubers to act as a dominant sink during assimilate partitioning might be associated with PBZ stimulated low GA level in the tuber tissue that increases tuber sink activity [72].
Setia et al. [75] also reported that the application of PBZ resulted in an overall increase in dry weight per plant and better partitioning of assimilates (percent ratio of siliqua dry weight to plant dry matter) in Brassica juncea and Brassica carinata. Similarly, Kumar et al. [45] reported that PBZ treatment enhanced seed yield in Camelina sativa and this enhancement of yield was correlated with improvement in CO2 assimilation physiology, sink activity partitioning of assimilates and rooting.
In addition, Yeshitela et al. [76] reported that the higher PBZ rates suppressed vegetative growth of mango and the assimilate that was to be expended for vegetative growth was diverted to intensifying flowering. This was proved by a higher total non-structural carbohydrate level of the shoots of the treated trees before flowering. Similarly, the reduction in vegetative growth of grape by altering relative sink strengths within the plant had an indirect consequence of allowing a greater partition of the assimilates to reproductive growth, to flower bud formation, fruit formation and fruit growth of treated plants [77].
Mineral uptake
By influencing shoot and root morphology, PBZ alters mineral uptake. Rieger [78] working in hydroponics on ‘Nemaguard’ peach rootstocks, found that PBZ treatment induced decreases in N, P, K, Fe and Mo, whereas levels of Ca, Mg, B and Mn were increased by PBZ. This author stated that the magnitude of changes in foliar nutrition was proportional to the degree of growth suppression. In the case of Fuji apple trees, Huang et al. [79] found that the differences in the total dry matter accumulated per kg of leaves were negligible. On the other hand, Wang et al. [80] observed that the PBZ treatments increase the content of N, P, K, Ca, Mg, B, and Zn in leaves of pear tree. Rieger and Scalabrelli [81] demonstrated in peach tree that the foliar concentrations of N, P, K, and Fe decrease slightly, while increase those of Ca, Mg, B and Mn. Recently, Yeshitela et al. [76] also reported that PBZ increased mango leaf Mg, Cu, Zn, and Fe content without affecting the concentration of N, P, K, and Ca. In addition, this author indicated that the higher concentration of PBZ (8.25 g a.i./tree) resulted in a decreased Cu concentration, while the increase in PBZ concentration (2.75-8.25 g a.i./tree) did not show an increment of the concentration of Zn.
Plant growth, yield and quality
Germination and seedling development
Common problems found when treating seeds with growth regulators are reduction or absence of germination and delay in seedling emergence. Apple seeds (Pyrus malus Mill.) imbibed in 7 mg L−1 PBZ solution had 35% inhibition of germination and a germination delay by 2 days [82]. Similarly, Almond (Prunus dulcis L.) seeds soaked in 4000 or 8000 mg L−1 PBZ solutions during 15 min failed to germinate [83].
Germination percentage of tomato seeds that were soaked in 500 or 1000 mg L−1 PBZ for 6, 16, or 24 h was lower than that of water-soaked seeds [84]. They further found that seedling height suppression at 36 days after sowing was > 30% for seeds that had been soaked for 16 h in 500 or 1000 mg L−1 PBZ compared to those soaked in water. Pill and Gunter [85] also found that exposing Cosmos bipinnatus seeds to 1000 mg L−1 PBZ during soaking or priming reduced seedling height and also reduced seedling emergence with the responses being greater with longer exposure during priming than during soaking. Similarly, Pasian and Bennett [84] noted that 500 or 1000 mg L−1 PBZ reduced and delayed germination of tomato, geranium, and marigold seeds. These could be due to PBZ that adheres to the seed coat of treated seeds and then diffuses into the growth medium where it can be taken up by the seedling roots [86]. However, PBZ may penetrate the seed coat and exert a direct toxic effect on the embryo. The seed coats of tomato have a semipermeable layer [87] that may prevent the PBZ from entering the endosperm and embryo thereby lessening negative effects of PBZ on germination.
In addition, Kar and Gupta [88] described that treating sunflower (Helianthus annus L.) and safflower (Carthamus tinctorius L.) seeds with PBZ diminished the rate of germination and reduced seedling growth. Similarly, treating seeds with PBZ (at 250 mg per 1 kg seeds) retarded elongation of primary leaves in wheat (Triticum durum L.), barley (Hordeum vulgare L.), oat (Avena sativa L.), and rye (Secale cereale L.) [89]. The first true leaves of seedlings from triazole-treated seeds had a disturbed morphology. In these seedlings, root elongation was less severely retarded than shoot growth; roots were thicker and had higher water content [89].
For celery seeds, which require light for germination, the action of PBZ on GA biosynthesis was also light dependent. PBZ had a low effect on seed germination in the light, which might be due to increase in seed sensitivity to GA and, respectively, lower GA requirement for germination [90]. Other data also suggested that germination of PBZ-treated seeds is dependent on seed GA levels influenced by light intensity [91].
Addition of GA or other chemicals (ethephon) to seeds may overcome the influence of growth regulators on seed germination. Only 5% of petunia (Petunia hybrida L.) seeds germinated after imbibing them for 14 days in an agar medium containing 5 mg L−1 PBZ. The addition of 10 μM GA solution to the PBZ-treated substrate improved petunia germination up to 65% [92]. Soaking in PBZ solutions inhibited germination of amaranth (Amaranthus sp.) seeds, while further soaking seeds in gibberellin, ethephon, or 1-aminocyclopropane-1-carboxylic acid reversed the inhibitory effect of PBZ [93].
Shoot growth
Triazole treatments normally decreased the shoot length and increased thickness of the young plant stem, as well as the accelerated root formation is a significant advantage of the paclobutrazol treatment in Lycopersicon esculentum [94]. Triazole treatments have more pronounced effect of reducing height in wheat plants and appeared greener [6].
The most striking growth response observed in different species treated with PBZ is shoot growth reduction [95]. This response could be attributed primarily due to decreased internode length. PBZ was also found to effectively inhibit plant height, leaf expansion and alter the stem in Syzygium campanulatum [96]. Similarly, plant height was significantly reduced by PBZ application in canola [97], Vigna radiata [98], Epidendrum radicans [40], mango [99], wheat [68], Dianthus caryophyllus [36], Sesamum indicum [100], and Ocimum sanctum [101].
The PBZ effectively suppresses growth in a wide range of plant species, where treated plants tend to be smaller and more compact in appearance and have darker green leaves [102,103,104,105]. Terri and Millie [106] and Sebastian et al. [107] also reported that PBZ-treated plants tend to be dark green, shorter and more compact in appearance. Similarly, treating Chrysanthemum plants with PBZ as a soil drench resulted in thicker leaves, reduced stem diameter, and roots with an increased diameter [108]. Modification of shoot growth with the aid of PBZ may be helpful in maximizing return per unit land by allowing increased plant populations of the compact plants per unit land area.
Leaf growth
Leaf area: Triazole treatments significantly reduced the leaf area and the reduction might be due to the reduction in leaf size [109]. Gomathinayagam et al. [110] also reported that triazole treatments reduced the leaf area in Manihot esculenta.
PBZ induces various morphological modifications depending on plant species, growth stage, rate and method of application [76, 107]. Vijayalakshim and Srinivasan [111] found that application of PBZ in mango found to be significantly superior in increasing the leaf area compared to other treatments like potassium nitrate, urea and ethrel recording an average area of 94.89 cm2, whereas the control was only 63.65 cm2. According to these authors, the increase in leaf area has overcome the limitation of depletion for reserve food materials. As the reserve food materials were then plenty, the breaking up of alternate bearing cycle in the cultivars chosen has been achieved. However, this was found to be contradictory to the finding of Fernandez et al. [112] who reported a decrease in leaf area with PBZ in Phillyrea angustifolia. Similarly, Paclobutrazol treatment also reduced the leaf area in Solanum tuberosum [72], Ocimum sanctum [101], Hordeum vulgare [113], Catharanthus roseus [114] and zinnia plants [115]. Although PBZ decreased the surface area of the plants, it improved the durability of leaves; therefore, the decrease in the surface area of leaves was compensated by the lack of leaf falling and by the leaf durability [72].
Chlorophyll synthesis Several studies demonstrated increase in chlorophyll content in triazole-treated plants [7, 94]. The greening effect caused by plant treatments with growth regulators can be explained by an increase in chlorophyll content and/or more densely packed chloroplasts per unit leaf area due to a reduction in leaf area [116]. A similar explanation is suggested for the increased chlorophyll a and b contents in potato leaves [72].
Dewi et al. [117] reported that black rice plants treated with either 25 or 50 ppm PBZ have greener leaves compared to control and the leaves also experienced late senescence. This could be due to an increase in the activity of oxidative enzymes that prevented cell maturation. Similarly, studies on Jatropha [46], tef [118] and Camelina [119] showed that chlorophyll was higher on plants treated with PBZ compared to control. The increased chlorophyll content treated with PBZ might be from minimized damage caused by reactive oxygen and changes in the levels of carotenoids, ascorbate and the ascorbate peroxidase.
The report of Nivedithadevi et al. [120] showed that plants treated with PBZ synthesized more cytokinin, which in turn enhanced chloroplast differentiation and chlorophyll biosynthesis, and prevented chlorophyll degradation. Berova and Zlatev [94] also reported that the increase in chlorophyll content may be ascribed to higher cytokinin content that is known to stimulate chlorophyll biosynthesis and/or reduced chlorophyll catabolism. Furthermore, PBZ appears to have delayed the onset of senescence, represented by the rate of chlorophyll degradation in attached mung bean leaves, which was probably due to the enhanced endogenous level of cytokinins through their secondary effect on plants [7]. In several plant species, PBZ-treated leaves were retained longer and the onset of senescence considerably delayed [59, 60]. The senescence delaying activity may be related to the influence of PBZ on the endogenous cytokinin content [7].
Rate of photosynthesis and transpiration rate Contradictory reports have been published regarding the effects of PBZ on crop photosynthetic efficiency; however, indirectly by reducing leaf area it may reduce photosynthetic surface area and thereby reduce the whole-plant photosynthesis [25]. DeJong and Doyle [121] noted no apparent effect of PBZ on photosynthetic rate of well exposed nectarine leaves during most of the growing season. Davis et al. [25] reported that photosynthetic rate decreased as a result of inhibition of leaf expansion. Leaf is an important plant part as it contains mesophyll cells that are specialized as photosynthetic tissues. Gaussoin et al. [122] also reported that plants treated with plant growth retardants often had a moderate restraining effect on carbon dioxide exchange rate, thus possibly reducing the photosynthetic rate.
Ahmad Nazarudin et al. [123] described that photosynthetic rate and transpiration rate of Syzygium myrtifolium were reduced after treatment with PBZ. This might be due to the reduction in photosynthetic rate that would also imply reduction in the transpiration rate as both processes are associated with the opening and closing of stomata [124]. The decline in transpiration rate would then reduce the percentage of water released through stomata. Olsen and Andersen [125] reported that reduction in transpiration rate would protect the plant against abiotic stress due to water restriction or drought period. In addition, PBZ enhanced stress tolerance of plants by increasing the xylem pressure potential of the treated plants and thereby enhancing plant moisture status during drought period. According to Abod and Jeng [126], the reduction in photosynthetic rate and transpiration rate was influenced by the stomatal activity and leaf area. However, Ahmad Nazarudin et al. [123] reported that stomatal activity was not the main factor in the reduction of photosynthetic and transpiration rate in Syzygium myrtifolium. The possible reason could be the reduction of leaf area, which further contributed to the reduction of the total leaf surface that absorbs the sunlight.
On the contrary, PBZ increased the rate of net leaf photosynthesis [72]. This could be attributed to the higher chlorophyll content and earlier tuberization in response to the PBZ treatment. The increased net photosynthesis in response to PBZ has also been reported in Setaria italic [127] and horse chestnut [128]. Similarly, Soumya [129] reported that foliar and drench applications of PBZ in chickpea were found to maintain higher rates of photosynthesis under water deficit condition and faster recovery after water stress termination. The increase in intercellular CO2 concentration and alteration in stomatal conductance was assessed the reasons for higher photosynthesis in PBZ-treated Amorphophallus campanulatus [20, 130] and potted red fire spike [131].
Leaf thickness: The higher epicuticular wax deposition on treated leaves may be related to the increase in endogenous ABA levels in response to PBZ treatment [23]. An increase in ABA stimulates the synthesis of lipid transfer proteins in barley that play an important role in the formation of epicuticular waxes, a process that affects the water relation of the leaves [132]. PBZ treatments caused an increase of 10% in total wax load and change the proportion of certain wax constitutes in potted rose cultivars within 11 days of application [133]. The development of a thicker epicuticular wax layer provides better protection against some plant pathogens and minor mechanical damage [134].
Tekalign and Hammes [72] observed that potato plants cv. Zemen treated with PBZ at the dosage of 67.5 mg a.i. per plant increased total leaf thickness from 215 to 267 µm bringing about 24% increase higher than the control. These authors also indicated that the increase in leaf thickness is attributed to an increase in epidermal cell diameter, palisade cell length and spongy mesophyll depth. Sopher et al. [35] also reported that in maize, PBZ-treated leaves showed more epicuticular wax deposition and were thicker and broader owing to enlarged vascular elements, epidermal, mesophyll, and bundle sheath cells.
In peanut (Arachis hypogaea L.), leaves treated with PBZ exhibited well-differentiated palisade and spongy mesophyll layers with longer and longer cells [135]. Similarly, Kishorekumar et al. [136] indicated that Chinese potato leaves (Solenostemon rotundifolius) treated with PBZ showed an increased thickness of the upper and lower epidermis, as well as the length of the palisade and spongy cells. In addition, Jaleel et al. [137] reported that cells number in palisade (mesophyll) tissue per unit area have been reported to enhance to a larger extent in PBZ-treated Catharanthus roseus.
Stem growth
Suppression of plant height by PBZ occurs because the compound blocks three separate steps in the terpenoid pathway for the production of gibberellins (GAs). GA enhances internode elongation of intact stems [124]. Liu and Loy [138] showed that GA promotes cell division by stimulating cells in the G1 phase to enter the S phase and by shortening the duration of S phase. They concluded that increased cell numbers lead to more rapid stem growth. But treating plants with PBZ resulted in stems with the same numbers of leaves and internodes compressed into a shorter length [7, 139]. Similarly, reduction in internode length was indicated in tomato in response to PBZ treatment [140]. He further noticed that the application of PBZ at a rate of 400 ppm resulted in a decreased internode length as compared to 200 ppm PBZ application. PBZ can be effective for obtaining sturdy plant and reducing plant height in several species without decreasing flowering quality [141, 142]. Webster and Quinlan [143] also reported that PBZ has great efficacy in reducing height growth of many temperate fruit species and cultivars. Similar reductions in plant height were reported in potato [72], Scaevola [106] and Dianthus caryophyllus [107], Syzygium myrtifolium [123] and Mangifera indica [76] in response to PBZ treatment.
Ahmad Nazarudin et al. [123] reported that PBZ application showed a stunted growth of Syzygium myrtifolium during the 5-month period after application, while the increased PBZ concentration (1.25–3.75 g L−1) did not further reduced the plant height. Similarly, Tekalign and Hammes [72] indicated that both foliar and soil drench application of PBZ reduced the height of potato, though foliar application of PBZ is more effective than soil drench application. These authors further indicated that the increase PBZ concentration (45–90 mg a.i./plant) resulted in a reduced plant height. In addition, Yeshitela et al. [76] also described that the height of mango trees and length of new shoots shortened due to both foliar and soil drench application of PBZ.
PBZ treatment increased cortex thickness, size of the vascular bundles, and pith diameter and resulted in thicker stems [72]. This modification may be attributed to radial expansion of cells due to reduced endogenous GA activities in response to the treatment. Wenzel et al. [144] reported that GA limits the extent of radial expansion of plant organs. In dicot stems, cell shape alterations are apparently caused by a more longitudinal orientation of cellulose microfibrils being deposited in the cell walls, preventing expansion parallel to the these microfibrils but allowing expansion perpendicular to them [145]. The non-uniform distribution and arrangement of the vascular elements in the potato stems resulted in irregularity in the shape of the stems. Various authors reported different results in various plant species with respect to PBZ-induced stem anatomy modifications. PBZ induced both cell number and length in safflower stem [146]. Burrows et al. [108] reported that PBZ treatment brought about a 50% reduction in chrysanthemum stem diameter because of an enhanced development of secondary xylem and a marked reduction in the number of sclerenchyma bundle caps. In peach shoots, PBZ reduced the proportion of xylem and increased that of phloem and cortex, and increased xylem density [147].
Root growth
Triazole treatments induced the root growth in cucumber, which was associated with increased the endogenous cytokinin levels [148]. PBZ treatment increased the root length and enhanced the lateral roots in tomato plants [94], Vigna unguiculata [149] and Festuca plant [150]. PBZ induced the root growth in both maize and wheat [151], mango [152], avocado [153], Abelmoschus esculentus [154] and Ocimum sanctum [101]. Swietlik and Miller [155] reported that root length was stimulated by PBZ applications at low to moderate concentrations. Higher concentrations, however, may reduce root growth.
PBZ increased root diameter by increasing the width of cortex and by favoring the formation of more secondary xylem vessels. This modification may be attributed to radical expansion of cells due to reduced endogenous GA activities in response to the treatment [72]. These authors further indicated that untreated plants had more, thinner and longer roots compared to the treated plants. Increased root diameter has been correlated with larger cortical parenchyma cells in soybean and maize [156]. Increasing root diameter in chrysanthemum was due to an increase number of rows and diameter of cortical cells [108].
PBZ was observed to increase diameter and length of fibrous roots, enhances lateral root formation, reduced the diameter of xylem vessels; however, phloem sieve tubes had shown an increased diameter in PBZ-treated Catharanthus roseus plants [137]. The increased root length in PBZ-treated plants was found to be associated with larger parenchyma cells and promotes cell expansion radially [7].
Flower enhancing
PBZ is effective not only in flower induction but also in early and off season flower induction in mango [77, 157,158,159,160]. PBZ, a gibberellin inhibitor, reduces vegetative promoter level and thereby increases florigenic promoter/vegetative promoter ratio which stimulates flowering shoots in weakly inductive shoots of fruit crops [76, 161,162,163].
Exogenous application of GA as well as endogenous high levels of gibberellins have proved a major hindrance in the way of flower bud differentiation in a number of temperate as well as tropical fruits [164]. PBZ, owing to its anti-gibberellin activity could induce or intensify flowering by blocking the conversion of kaurene into kaurenoic acid. The latter is a precursor of gibberellins. PBZ can considerably enhance the total phenolic content of terminal buds and alter the phloem to xylem ratio of the stem [165]. Such alterations could be important in restricting vegetative growth and enhancing flowering by altering assimilates partitioning and patterns of nutrient supply for new growth.
The application of PBZ before flower bud differentiation or 3 months earlier than anticipated flowering has been effective in inducing flowering in mango without accompanying reduction in shoot length. However, higher concentration leads to canopy and panicle compaction [166, 167]. The response to PBZ varied with cultivar and crop load. The effectiveness of PBZ in promoting flowering in Citrus sp. depends on the crop load as the heavy fruit load trees scarcely flowered. In medium to low fruit load trees, PBZ significantly increased the percentage of sprouted buds and floral shoots and reduced the number of vegetative shoots [168].
Fruit and tuber yield
Fruit yield: Foliar application of PBZ (200 ppm) was effective in increasing yield and minimizing fruit drop and fruit cracking in ber [169]. The effectiveness of PBZ was dependent on stage of development as the application of PBZ at bud bursting and 2 weeks before anthesis of grape increased the yield significantly [77].
Soil application around the tree trunk (collar drench) was more efficacious than foliar application as it ensures proper uptake in inducing fruiting [170]. On the other hand, Yeshitela [77] reported that application of PBZ both as a soil drench and foliar application was effective in suppressing vegetative growth and enhancing yield in mango.
Souza-Machado et al. [171] reported that significant earliness in harvest maturity was recorded in PBZ-treated tomato plants but no significant total yield differences were recorded between the PBZ and control plants. However, Giovinazzo et al. [172] found significant yield increases of 13% due to the PBZ treatments together with earlier harvest maturity by 6%. Similarly, Berova and Zlatev [94] reported that application of PBZ increased both early fruit yield and index of economic earliness in tomato. In addition, Mohamed et al. [173] also reported the remarkable improvement in fruit yield and water use efficiency of tomato.
There is usually a yield increase in mango associated with PBZ treatments, but Voon et al. [161] emphasized the importance of supplying adequate nutrients, irrigation and generally good tree maintenance to maintain these high yields. In the experiments of Medonca et al. [174] also, PBZ increased the productivity of ‘Tommy Atkins’. Similar increase in productivity of ‘Tommy Atkins’ mango (increased total fruit number and total fruit weight per tree) was indicated in response to PBZ treatment [76]. He also indicated that soil application of PBZ increased total fruit number per tree and total fruit weight per tree as compared to foliar spray and the increased application of PBZ (2.78–8.25 g a.i./tree) also resulted in the higher total fruit number per tree and total fruit weight per tree.
Tuber yield: In the trials of Balamani and Poovaiah [175], PBZ application resulted in increased tuber yield per plant. However, it is not clear whether the reported yield increments were a consequence of an increase in tuber size or number. On the contrary, Bandara and Tanino [176] reported that PBZ nearly doubled the number of tubers per plant without affecting the total fresh weight of the tubers. This discrepancy may probably be explained by the cooler growing conditions in their experiment. Tekalign and Hammes [72] showed that PBZ application resulted in decreased tuber number per plant, which could be linked to the decline in stolon number as a result of a decrease in GA activity that may be associated with stolon initiation, and a strong negative correlation between tuber fresh mass and number signifying that the substantial increase in individual tuber size was responsible for the yield increment. This may be due to the interplay of early tuberization, increased chlorophyll content, enhanced rate of photosynthesis, and retaining photosynthetically active leaves longer in response to the treatment. In addition, these authors described that PBZ application increased tuber fresh mass, dry matter content and specific gravity. They further indicated that low concentration of PBZ increased tuber fresh and dry weight, while the increase in PBZ concentration (67.5–90.0 mg a.i./plant) did not increase the value of these parameters.
Similarly, the application of PBZ reduced the number of potato tubers [73, 102], the number of mini tubers [177] and the number of cassava tuber [178]. On the contrary to the increased fresh and dry weight of potato tuber [102, 179], potato mini tuber [177] and elephant foot yam [180], the fresh weight of cassava tuber was reduced in response to PBZ treatment [178].
Fruit and tuber quality
Fruit quality: Fruit quality of mango and lemon (TSS and acid content) increased with PBZ application [160, 181]. Similarly, Vijayalakshmi and Srinivasan [111] and Yeshitela et al. [76] reported that applying PBZ in mango had the greatest effect in increasing total sugar, reducing sugar, TSS, sugar: acid ratio and a decrease in titratable acidity. On the other hand, PBZ had shown no improvement in fruit quality of grapes [77], strawberries [182] and peach [183].
Foliar application of 2500–3000 ppm PBZ three weeks after full bloom increased TSS and reduced acidity in peach [184] and cherries [185]. Yeshitela et al. [76] also reported that the application of 8.25 g a.i./tree PBZ resulted in increased TSS. Even though, PBZ increased the quality of fruits, it was ascertained that the accumulation of PBZ residues on the surface or inside mango fruit (especially due applications of higher rates) is unfriendly to human health [42].
Tuber quality: The PBZ application to cassava plant significantly increased starch content was reported by Yang and Cao [186] and Medina et al. [178]. Similarly, in potato, the application of PBZ decreased the assimilate partitioning to stem, leaves, root and stolon but increased the partitioning of dry mass production to tubers [72]. An increase in specific gravity and dry matter content of the tubers in response to PBZ may be attributed to reduced GA activity in the tuber tissue that in turn increased sink strength to attract more assimilates and enhance starch synthesis [72]. Under favorable conditions for tuberization (GA content below threshold level due to PBZ), the activities of enzymes involved in potato tuber starch biosynthesis such as ADPG-pyrophosphorylase, starch phosphorylase and starch synthase increased [187].
PBZ also help to increase the production of antioxidant such as carotenoid content in plants to fight against oxidative stress [7, 188, 189]. Similar results were reported on tuber crop species that utilize low concentrations of triazole compound derivatives as treatments with concentrations ranging from 10 to 30 ppm to increase carotenoid content in the tuber of white yam [190].
Conclusion
PBZ, a triazole, is an extremely active chemical and affects almost all plant species, whether applied as a spray or a soil drench. It is more effective when applied to the growing media and application on the growing medium would give longer absorption time and more absorption of active ingredient than foliar spray. It inhibits GA biosynthesis by blocking the oxidation of ent-kaurene. PBZ has been used to provide plant protection against numerous abiotic stresses such as chilling, water deficit stress, flooding and salinity. PBZ depressed the vegetative growth components, but GA induced vegetative growth components through total shoot length and total bud number increases. PBZ induced the increase in tuber yield, specific gravity and dry matter yield, fruit number and yield, TSS, TSS/TA, reducing sugar and total sugar, and a decrease in TA. This review has compiled and discussed the nature of PBZ, the role of PBZ as a protection against numerous abiotic stresses such as chilling, water deficit stress and heat stress, the effects of PBZ on the vegetative growth, yield and quality of crops. This review will be useful for the professionals and researchers working on plant growth regulators to improve crop production through the use of PBZ.
Availability of data and materials
Not applicable.
References
Kumari S, Bakshi P, Sharma A, Wali VK, Jasrotia A, Kour S. Use of plant growth regulators for improving fruit production in sub tropical crops. Int J Curr Microbiol App Sci. 2018;7(03):659–68.
Yan W, Zhang XX, Yuan A. Effects of two plant growth regulators on the growth and recovery of alfalfa seedlings exposed to aluminum stress. J Shanghai Jiaotong Univ. 2011;29:75–82.
Vineeth TV, Kumar P, Yadav S, Pal M. Optimization of bio-regulators dose based on photosynthetic and yield performance of chickpea (Cicer arietinum L.) genotypes. Indian J Plant Physiol. 2015;20:177–81.
Vineeth TV, Kumar P, Krishna GK. Bioregulators protected photosynthetic machinery by inducing expression of photorespiratory genes under water stress in chickpea. Photosynthetica. 2016;54:234–42.
Jaleel CA, Gopi R, Manivannan P, Panneerselvam R. Responses of antioxidant defense system of Catharanthus roseus (L.) to paclobutrazol treatment under salinity. Acta Physiol Plant. 2007;29:205–9.
Hajihashemi S, Kiarostami K, Saboora A, Enteshari S. Exogenously applied paclobutrazol modulates growth in salt-stressed wheat plants. Plant Growth Regul. 2007;53:117–28.
Fletcher R, Gilley A, Sankhla N, Davis T. Triazoles as plant growth regulators and stress protectants. Hort Rev. 2000;24:55–137.
Gopi R, Jaleel C, Sairam R, Lakshmanan GMA, Gomathinayagam M, Panneerselvam R. Differential effects of hexaconazole and paclobutrazol on biomass, electrolyte leakage, lipid peroxidation and antioxidant potential of Daucus carota L. Colloids Surf B. 2007;60:180–6.
Barrett JE. Mechanisms of action. In: Gaston ML, editor. Tips on regulating growth of floriculture crops. Batavia: Ball Publishing; 2001. p. 32–47.
Sponsel VM. The biosynthesis and metabolism of gibberellins in higher plants. In: Davis PJ, editor. Plant hormones: physiology, biochemistry, and molecular biology. 2nd ed. Dordrecht: Kluwer Academic Pub; 1995.
Million JB, Barrett JE, Nell TA, Clark DG. Paclobutrazol distribution following application to two media as determined by bioassay. Hort Sci. 1999;34:1099–102.
Syngenta. Product Details: Bonzi.20 Mar. 2003. http://www.syngentaprofessionalproducts.com/labels/Index.asp?nav=PrdLst&F=PrdDsp.
Norremark I, Andersen A. Effect of paclobutrazol on seed propagated Pelargonium x hortorum L.H. Bailey. Gartenbauwissenschaft 1990; 55:1-8.
Rademacher E. Growth retardants: effects on gibberellin biosynthesis and other metabolic pathway. Annu Rev Plant Physiol Mol Biol. 2000;51:501–31.
Davis TD, Curry EA. Chemical regulation of vegetative growth. Crit. Rev. Pl. Sci. 1991;10:151–88.
Chaney WR. Growth retardants: A promising tool for managing urban trees. Purdue Extension document FNR-252-W; 2005.
Fletcher RA, Hofstra G, Gao J. Comparative fungi toxic and plant growth regulating properties of triazole derivatives. Plant Cell Physiol. 1986;27:367–71.
Hedden P, Graebe JE. Inhibition of gibberellins biosynthesis by paclobutrazol in cell-free homogenates of Cucurbita maxima endosperm and Malus pumila embryos. J Plant Growth Regul. 1985;4:111–2.
Fletcher RA, Hofstra G. Triazoles as potential plant protectant. In: Berg D, Plempel M, editors. Sterol synthesis inhibitors in plant protection. Ellis Horwood Ltd: Cambridge; 1988. p. 31–331.
Gopi R, Jaleel C. Photosynthetic alterations in Amorphophallus campanulatus with triazoles drenching. Glob J Mol Sci. 2009;4:15–8.
Zhu L, Van De Peppel A, Li X. Changes of leaf water potential and endogenous cytokinins in young apple trees treated with or without paclobutrazol under drought conditions. Sci Hort. 2004;99:133–41.
Rady M, Gaballah S. Improving barley yield grown under water stress conditions. Research Journal of Recent Sciences. 2012;1:1–6.
Rademacher W. Bioregulation of crop plants with inhibitors of gibberellins biosynthesis. Proc Plant Growth Regul Soc Am. 1997;24:27–31.
Marshall JG, Rutledge RG, Blumwald E, Dumbroff EB. Reduction in turgid water volume in Jack pine, white spruce and black spruce in response to drought and paclobutrazol. Tree Physiol. 2000;20:701–7.
Davis TD, Steffens GL, Sankhla N. Triazole plant growth regulators. Hort. Rev. 1988;10:63–105.
Witchard M. Paclobutrazol is phloem mobile in castor oil plants (Ricinus communis L.). J Plant Growth Regul. 1997;16:215–7.
Browning G, Kuden A, Blake P. Paclobutrazol promotion of axillary flower initiation in pear cv. Doyenne du Comice. J Hort Sci. 1992;67:121–8.
Jung J, Rentzea C, Rademacher W. Plant growth regulation with triazoles of the dioxanyl type. J Plant Growth Regul. 1986;4:181–8.
Early JD, Martin GC. Translocation and breakdown of 14C-labeled paclobutrazol in ‘Nemaguard’ peach seedling. HortSci. 1988;23(1):197–9.
Sterret JP. XE-1019: plant response, translocation, and metabolism. J Plant Growth Regul. 1988;7:19–26.
Reed AN, Curry EA, Williams MW. Translocation of triazole growth retardants in plant tissue. J Am Soc Hort Sci. 1989;114:893–8.
Rademacher W. Plant growth regulators: backgrounds and uses in plant production. J Plant Growth Regul. 2015;34(4):845–72.
Franca CFM, Costa LC, Ribeiro WS, Mendes TDC, Santos MNS, Finger FL. Evaluation of paclobutrazol application method on quality characteristics of ornamental pepper. Ornam Hort. 2017;23(3):821–9.
Ribeiro DM, Muller C, Bedin J, Rocha GB, Barros RS. Effects of autoclaving on the physiological action of paclobutrazol. Agric Sci. 2011;2:191–7.
Sopher CR, Krol M, Huner NPA, Moore AE, Fletcher RS. Chloroplastic changes associated with paclobutrazol-induced stress protection in maize seedling. Can J Bot. 1999;77(2):279–90.
Banon S, Gonzalez A, Cano EA, Franco JA, Fernandez JA. Growth, development and colour response of potted Dianthus caryophyllus cv. Mondrian to paclobutrazol treatment. Sci Hort. 2002;94:371–7.
Alkhassawneh NM, Karam NS, Shibli RA. Growth and flowering of black iris (Iris nigricans Dinsm.) following treatment with plant growth regulators. Sci Hort. 2006;107:187–93.
Sharma D, Awasthi MD. Uptake of soil paclobutrazol in mango (Mangifera indica L.) and its persistence in fruit and soil. Chemosphere. 2005;60:164–9.
Latimer JG, Scoggins HL, Banko TJ. Persistence of plant growth regulator effects on perennial plants in the nursery. Acta Hort. 2003;624:229–32.
Pateli P, Papafotiou M, Chronopoulos J. Comparative effects of four plant growth retardants on growth of Epidendrum radicans. J Hortic Sci Biotechnol. 2004;79:303–7.
Gent MPN, McAvoy RJ. Plant growth retardants in ornamental horticulture. In: Basra AS, editor. Plant growth retardants in agriculture and horticulture: Their role and commercial uses. Binghamton: Food Prod Press; 2000. p. 89–146.
Singh DK, Ram S. Level of paclobutrazol residues in shoot and fruit of mango. Indian J Plant Physiol. 2000;5:186–8.
Tongumpai P, Jutamanee K, Subhadrabandhu S. Effect of paclobutrazol on flowering of mango cv. ‘Khiew Sawoey’. Acta Hort. 1991;291:67–79.
Kumar S, Ghatty S, Satyanarayana J, Guha A, Chaitanya BSK, Reddy A. Paclobutrazol treatment as a potential strategy for higher seed and oil yield in field-grown Camelina sativa L. Crantz. BMC Res Notes. 2012;5:1–13.
Senoo S, Isoda A. Effects of paclobutrazol on dry matter distribution and yield in peanut. Plant Prod Sci. 2003;6:90–4.
Ghosh A, Chikara J, Chaudhary DR, Prakash AR, Boricha G, Zala A. Paclobutrazol arrests vegetative growth and unveils unexpressed yield potential of Jatropha curca. J Plant Growth Regul. 2010;29:307–15.
Kamran M, Wennan S, Ahmad I, Xiangping M, Wenwen C, Xudong Z, Tiening L. Application of paclobutrazol affect maize grain yield by regulating root morphological and physiological characteristics under a semi-arid region. Scientific Reports. 2018;8:1–5.
Patil HK, Talathi JM. Economic viability of paclobutrazol application in mango cv. Alponso. South Indian Horticulturae. 1999;47(1–6):145–6.
Rai N, Yadav DS, Yadav RK. Standardization of Paclobutrazol concentrations in vegetable crops. Veg Sci. 2002;29:149–53.
Yamaguchi S. Gibberellin metabolism and its regulation. Annu Rev Plant Biol. 2008;59:225–51.
Achard P, Genschik P. Releasing the brakes of plant growth: how GAs shutdown DELLA proteins. J Exp Bot. 2009;60:1085–92.
Hartmann A, Senning M, Hedden P, Sonnewald U, Sonnewald S. Reactivation of meristem activity and sprout growth in potato tubers require both cytokinin and gibberellin. Plant Physiol. 2011;155(2):776–96.
Lever BG. ‘‘Cultar’’—A technical overview. Acta Hort. 1986;179:459–66.
Mackay C, Hall J, Hofstra G, Fletcher R. Uniconazole-induced changes in abscisic acid, total amino acids and proline in Phaseolus vulgaris. Pesti Biochem Physiol. 1990;37:74–82.
Hauser C, Kwiatkowski J, Rademacher W, Grossmann K. Regulation of endogenous abscisic acid levels and transpiration in oilseed rape by plant growth retardants. J Plant Physiol. 1990;137:201–7.
Aly A, Latif H. Differential effects of paclobutrazol on water stress alleviation through electrolyte leakage, phytohormones, reduced glutathione and lipid peroxidation in some wheat genotypes (Triticum aestivum L.) grown in-vitro. Romanian Biotechnological Letters. 2011;16:6710–21.
Binns AN. Cytokinin accumulation and action: biochemical, genetic, and molecular approaches. Annu Rev Plant Physiol. 1994;45:173–96.
Mok MC. Cytokinins and plant development - an overview. In: Mok DWS, Mok MC, editors. Cytokinins: chemistry and function. Boca Raton: CRC Press; 1994.
Hunter DM, Proctor JTA. Paclobutrazol affects growth and fruit composition of potted grape vines. Hort Sci. 1992;27:319–21.
Basiouny FM, Sass P. Shelf life and quality of rabbit eye blueberry fruit in response to pre-harvest application of CaEDTA, nutrical and paclobutrazol. Acta Hort. 1993;368:893–900.
Visser C, Fletcher RA, Saxena PK. Thidaizuron stimulates expansion and greening in cucumber cotyledons. Physiol Mol Biol Plants. 1992;1:21–6.
Zhang M, Duan L, Tian X, He Z, Li J, Wang B, Li Z. Uniconazole-induced tolerance of soybean to water deficit stress in relation to changes in photosynthesis, hormones and antioxidant system. J Plant Physiol. 2007;164:709–17.
Senaratna T, Mackay C, McKersie B, Fletcher R. Uniconazole-induced chilling tolerance in tomato and its relationship to antioxidant content. J Plant Physiol. 1988;133:56–61.
Pinhero RG, Fletcher RA. Paclobutrazol and ancymidol protects corn seedlings from high and low temperatures stresses. J Plant Growth Regul. 1994;15:47–53.
Kraus TE, Fletcher RA. Paclobutrazol protects wheat seedlings from heat and paraquat injury: is detoxification of active oxygen involved? Plant Cell Physiol. 1994;35:45–52.
Larsen MH, Davis TD, Evans RP. Modulation of protein expression in uniconazole treated soybeans in relation to heat stress. Proc Plant Growth Regul Soc Am. 1988;15:177–82.
Scandalios J. Oxygen stress and superoxide dismutases. Plant Physiol. 1993;101:7–12.
Berova M, Zlatev Z, Stoeva N. Effect of paclobutrazol on wheat seedlings under low temperature stress. Bulg J Plant Physiol. 2002;28(1–2):75–84.
Almekinders CJM, Struik PC. Shoot development and flowering in potato (Solanum tuberosum L.). Potato Res. 1996;39:581–607.
Le Guen-Le Saos F, Hourmant A, Esnault F, Chauvin JE. In vitro bulb development in shallot (Allium cepa L. Aggregatum Group): effects of anti-gibberellins, sucrose and light. Ann Bot. 2002;89:419–25.
De Resende GM, De Souza RJ. Effects of paclobutrazol doses on garlic crop. Pesquisa Agropecuaria Brasileira. 2002;37(5):637–41.
Tekalign T, Hammes PS. Growth and biomass production in potato grown in the hot tropics as influenced by paclobutrazol. Plant Growth Regul. 2005;45:37–46.
Mabvongwe O, Manenji BT, Gwazane M, Chandiposha M. The effect of paclobutrazol application time and variety on growth, yield, and quality of potato (Solanum tuberosum L.). Adv Agric. 2016;2016:1–5.
Pan S, Rasul F, Li W, Tian H, Mo Z, Duan M. Roles of plant growth regulators on yield, grain qualities and antioxidant enzyme activities in super hybrid rice (Oryza sativa L.). Rice. 2013;6:6–9.
Setia RC, Kaur P, Setia N. Influence of paclobutrazol on growth and development of fruit in Brassica juncea (L.) Czern and Coss. J Plant Growth Regul. 1996;20:307–16.
Yeshitela T, Robbertse PJ, Stassen PJC. Effects of various inductive periods and chemicals on flowering and vegetative growth of ‘Tommy Atkins’ and ‘Keitt’ mango (Mangifera indica) cultivars. NZ J Crop Hort Sci. 2004;32:209–15.
Christov C, Tsvetkov I, Kovachev V. Use of paclobutrazol to control vegetative growth and improve fruiting efficiency of grapevines (Vitis vinifera L.). Bulg J Plant Pysiol. 1995;21:64–71.
Rieger M. Paclobutrazol, root growth, hydraulic conductivity, and nutrient uptake of ‘Nemaguard’ peach. HortScience. 1990;25(1):95–8.
Huang WD, Shen T, Han ZH, Liu S. Influence of paclobutrazol on photosynthesis rate and dry matter partitioning in the apple tree. J Pl Nut. 1995;18:901–10.
Wang SY, Byun JK, Steffens GL. Controlling plant growth via the gibberelllin biosynthesis system. II. Biochemical and physiological alterations in apple seedlings. Physiol Plant. 1985;63:169–75.
Rieger M, Scalabrelli G. Paclobutrazol, root growth, hydraulic conductivity, and nutrient uptake of ‘Nemaguard’ peach. HortScience. 1990;25:95–8.
Mage F, Powell L. Inhibition of stratification and germination of apple seeds by paclobutrazol. HortScience. 1990;25:577.
Koukourikou-Petridou MA. Paclobutrazol affects growth of almond fruits and germination of almond seeds. Plant GrowR reg. 1996;20:267–9.
Pasian CC, Bennett MA. Paclobutrazol soaked marigold, geranium, and tomato seeds produce short seedlings. HortScience. 2001;36:721–3.
Pill WG, Gunter JA. Emergence and shoot growth of cosmos and marigold from paclobutrazol-treated seed. J Environ Hort. 2001;19:11–4.
Pasian CC, Bennett MA. Seed coats as plant growth regulator carriers in bedding plant production. Acta Hort. 1999;504:93–7.
Beresniewicz MM, Taylor AG, Goffinet MC, Terhune BT. Characterization and location of a semipermeable layer in seed coats of leek and onion (Liliaceae), tomato and pepper (Solanaceae). Seed Sci Technol. 1995;23:123–34.
Kar C, Gupta K. Effect of triazole-type plant growth regulators on sunflower and safflower seed viability. Can J Bot. 1991;69:1344–8.
Buchenauer H, Kutzner B, Koths T. Effect of various triazole fungicides on the growth of cereal seedlings and tomato plants as well as on gibberellin contents and lipid metabolism in barley seedlings. 1984.
Pressman E, Shaked R. Germination of annual celery Apium graveolens seeds inhibition by paclobutrazol and its reversal by gibberellins and benzyladenine. Physiol Plant. 1988;73:323–6.
Li FL, Chen JC, Zhao YJ. Effects of gibberellin and light on seed germination and seedling growth of Arabidopsis. Acta Phytophysiol. 2000;26:101–4.
Izhaki A, Swain SM, Tseng TS, Borochov A, Olszewski NE, Weiss D. The role of SPY and its TRP domain in the regulation of gibberellin action throughout the life cycle of Petunia hybrida plants. Plant J. 2001;28:181–90.
Kerczynski J, Kerczynska E, Knypl JC. Effects of gibberellic acid, ethephon and 1-aminocyclopropane-1-carboxylic acid on germination of Amaranthus caudatus seeds inhibited by paclobutrazol. J Plant Growth Reg. 1988;7:59–66.
Berova M, Zlatev Z. Physiological response and yield of paclobutrazol treated tomato plants (Lycopersicon esculentum Mill.). Plant Growth Regul. 2000;30:117–23.
Rossini Pinto A, Deleo Rodrigues T, Leite I, Barbosa J. Growth retardants on development and ornamental quality of potted ‘lilliput’ Zinnia elegans Jacq. Scientia Agricola. 2005;62:337–45.
Ahmad Nazarudin MR, Mohd Fauzi R, Tsan FY. Effects of paclobutrazol on the growth and anatomy of stems and leaves of Syzygium campanulatum. J Trop Forest Sci. 2007;19(2):86–91.
Hua S, Zhang Y, Yu H, Lin B, Ding H, Zhang D, Ren Y, Fang Z. Paclobutrazol application effects on plant height, seed yield and carbohydrate metabolism in canola. Int J Agric Biol. 2014;16:471–9.
Bekheta MA, Talaal IM. Physiological response of mung bean Vigna radiata plants to some bioregulators. J Appl Bot Food Qual. 2009;83:76–84.
Murti GSR, Upreti KK. Effect of paclobutrazol on leaf water potential, Ethylene production, ACC, ACC-oxidase and polyamines in mango seedlings. J Plant Biol. 2005;32:183–8.
Abraham SS, Jaleel CA, Chang-Xing Z, Somasundaram R, Azooz MM, Manivannan P, Panneerselvam R. Regulation of Growth and Metabolism by Paclobutrazol and ABA in Sesamum indicum L. under drought condition. Glob J of Mol Sci. 2008;3(2):57–66.
Divya Nair V, Jaleel CA, Gopi R, Panneerselvam R. Changes in growth and photosynthetic characteristics of Ocimum sanctum under growth regulator treatments. Front Biol China. 2009;4(2):192–9.
Esmaielpour B, Hokmalipour S, Jalilvand P, Salimi G. The investigation of paclobutrazol effects on growth and yield of two potato (Solanum tuberosum) cultivars under different plant density. J Food Agric Environ. 2011;9(3&4):289–94.
Brito CLL, Matsumoto SN, Santos JL, Goncalves DN, Ribeiro AF. Effect of paclobutrazol in the development of ornamental sunflower. Revista de Ciências Agrárias. 2016;39(1):153–60.
Rahman MN, Shaharuddin HA, Wahab NA, Wahab PEM, Abdullah MO, Parveez GKA. Impact of paclobutrazol on the growth and development of nursery grown clonal oil palm (Elaeis guineensis Jacq.). J Oil Palm Res. 2016;28(4):404–14. https://doi.org/10.21894/jopr.2016.2804.02.
Hamdani JS, Nuraini A, Mubarok S. The use of paclobutrazol and shading net on growth and yield of potato ‘Medians’ tuber of G2 in medium land of Indonesia. J. Agron. 2018;17:62–7.
Terri WS, Millie SW. Growth retardants affect growth and flowering of Scaevola. HortSci. 2000;35(1):36–8.
Sebastian B, Alberto G, Emilio AC, Jose AF, Juan AF. Growth, development and color response of potted Dianthus carophyllus cv. Mondriaan to paclobutrazol treatment. Sci Hort. 2002;1767:1–7.
Burrows GE, Boag TS, Stewart WP. Changes in leaf, stem, and root anatomy of Chrysanthemum cv. Lillian Hoek following paclobutrazol application. J Plant Growth Regul. 1992;11:189–94.
Sheena A, Sheela VL. Effects of the growth retardant triadimefon on the Ex vitro establishment of Gladiolus (Gladiolus grandiflorus L.) cv. Vinks Glory. Plant Tissue Cult Biotech. 2010;20(2):171–8.
Gomathinayagam M, Jaleel CA, Alagu Lakshmanan GM, Panneerselvam R. Changes in carbohydrate metabolism by triazole growth regulators in Cassava Manihot esculenta Crantz; effects on tuber production and quality. Comples Rendus Biologies. 2007;330:644–55.
Vijayalakshmi D, Srinivasan PS. Morpho-physiological changes as influenced by chemicals and growth regulators in alternate bearing mango cv. “Alphonso”. Madaras Agr. J. 1999;86:485–7.
Fernandez JA, Balenzategui L, Banon S, Franco JA. Induction of drought tolerance by paclobutrazol and irrigation deficit in Phillyrea angustifolia during the nursery period. Sci Hort. 2006;107:277–83.
Sunitha S, Perras MR, Falk DE, Zhang R, Pharis RP, Fletcher RA. Relationship between gibberellins, height and stress tolerance on barley seedlings. Plant Growth Regul. 2004;42:125–35.
Jaleel CA, Gopi R, Chang-xing Z, Azooz MM, Panneerselvam R. Plant growth regulators and fungicides alters growth characteristics in Catharanthus roseus; comparative study. Glob J Mol Sci. 2008;3:93–9.
Asgarian H, Nabigol A, Taheri M. Effects of paclobutrazol and cycocel for height control of Zinnia. Int J Agron Plant Prod. 2013;4:3824–7.
Khalil IA, Rahman H. Effect of paclobutrazol on growth, chloroplast pigments and sterol biosynthesis of maize (Zea mays L.). Plant Sci. 1995;105:15–21.
Dewi K, Agustina RZ, Nurmalika F. Effects of blue light and paclobutrazol on seed germination, vegetative growth and yield of black rice (Oryza Sativa L. ‘CempoIreng’). Biotropia. 2016;23(2):85–96.
Tekalign T. Growth, photosynthetic efficiency, rate of transpiration, lodging, and grain yield of Tef (Eragrostis tef (Zucc.)Trotter) as influenced by stage and rate of paclobutrazol application. East Afr J Sci. 2007;1(1):35–44.
Sumit K, Ghatty S, Satyanarayana J, Guha A, Chaitanya BSK, Reddy AR. Paclobutrazol treatment as a potential strategy for higher seed and oil yield in field-grown Camelina sativa L. Crantz. BMC Res Notes. 2012;5:137.
Nivedithadevi D, Somasundaram R, Pannerselvam R. Effect of abscisic acid, paclobutrazol and salicylic acid on the growth and pigment variation in Solanum trilobatum (l). Int J Drug Dev Res. 2015;4:236–46.
DeJong T, Doyle JF. Leaf gas exchange and growth responses of mature ‘fantasia’ nectarine trees to Paclobutrazol. J Am Soc Hortic Sci. 1984;109:878–82.
Gaussoin RE, Branham BE, Flore JA. Carbon dioxide exchange rate and chlorophyll content of turf grasses treated with flurprimidol and mefluidide. J Plant Growth Regul. 1997;16:73–8.
Ahmad Nazarudin MR, Tsan FY, Mohd FR. Morphological and physiological response of Syzygium myrtifolium (Roxb) Walp. to paclobutrazol. Sains Malays. 2012;41(10):1187–92.
Salisbury FB, Ross CW. Plant physiology. 4th ed. California: Wadsworth Publishing Com; 1992.
Olsen WW, Andersen AS. The influence of five growth retardants on growth and postproduction qualities of Osteospermum ecklonis cv. Calypso. Sci Hort. 1995;62:263–70.
Abod SA, Jeng LT. Effects of Paclobutrazol and its method of application on the growth and transpiration of Acacia mangium Seedlings. Pertanika J Trop Agric Sci. 1993;16(2):143–50.
Bisht R, Singariya P, Bohra SP, Mathur N. Triazoles: their effects on net photosynthetic rate, transpiration rate and stomatal resistance in Setaria italica plants grown in vivo. Asian J Exp Sci. 2007;21:271–6.
Percival C, Noviss K. Triazole induced drought tolerance in horse chestnut (Aesculus hippocastanum). Tree Physiol. 2008;28:1685–92.
Soumya PR. Role of paclobutrazol in amelioration of water deficit stress in chickpea (Cicer arietinum L.). New Delhi: M.Sc. thesis, ICAR-Indian Agricultural Research Institute; 2014.
Manivannan P, Jaleel CA, Kishorekumar A, Sankar B, Somasundaram R, Sridharan R. Changes in antioxidant metabolism of Vigna unguiculata (L.) Walp. by propiconazole under water deficit stress. Colloids Surf B. 2007;57:69–74.
Rezazadeh A, Harkess RL, Guihong B. Effects of paclobutrazol and flurprimidol on water stress amelioration in potted red fire spike. Hort Technol. 2016;26:26–9.
Hollenbach B, Schreiber L, Hartungs W, Dietz KJ. Cadmium leads to stimulated expression of lipid transfer protein (ltp) in barley: implications for the involvement of LTP in wax assembly. Planta. 1997;203:9–19.
Jenks MA, Andersen L, Teusink RS, Williams MH. Leaf cuticular waxes of potted rose cultivars as affected by plant development, drought and paclobutrazol treatments. Physiol Plant. 2001;112:62–70.
Kolattukudy PE. Lipid-derived defensive polymers and waxes and their role in plant microbe interaction. In: Stumpf PK, editor. The metabolism, structure and function of plant lipids. New York and London: Plenum Press; 1987. p. 291–314.
Sankar B, Karthishwaran R, Somasundaram BSK. Leaf anatomical changes in peanut plants in relation to drought stress with or without paclobutrazol and ABA. J Phytol. 2016;5:25–9.
Kishorekumar A, Jaleel CA, Manivannan P, Sankar B, Sridharan R, Somasundaram R, Panneerselvam R. Differential effects of hexaconazole and paclobutrazol on the foliage characteristics of Chinese potato (Solenostemon rotundifolius Poir, JK Morton). Acta Biologica Szegediensis. 2006;50:127–9.
Jaleel CA, Manivannan P, Sankar B, Kishorekumar A, Sankari S, Panneerselvam R. Paclobutrazol enhances photosynthesis and ajmalicine production in Catharanthus roseus. Process Biochem. 2007;42:1566–70.
Liu PBW, Loy JB. Action of gibberellic acid on cell proliferation in the sub apical shoot meristem of watermelon seedlings. Am J Bot. 1976;63:700–4.
Taiz L, Zeiger E. Plant Physiology. 4th ed. Sunderland: Sinauer Associates Inc., Publishers; 2006.
Rahman H, Khan M, Khokhar M. Effect of paclobutrazol on growth and yield of tomato. Pak J Agric Res. 1989;10:49–51.
Mansuroglu S, Karaguzel O, Ortacesme V, Sayan MS. Effect of paclobutrazol on flowering, leaf and flower colour of Consolida orientalis. Pak J Bot. 2009;41(5):2323–32.
Currey CJ, Lopez RG. Paclobutrazol pre-plant bulb dips effectively control height of ‘Nellie White’ easter lily. HortTechnology. 2010;20:357–60.
Webster AD, Quinlan JD. Chemical control of tree growth of Plum (Prunus domestica L.). I. Preliminary studies with the growth retardant paclobutrazol. J Hort Sci. 1984;59:367–75.
Wenzel C, Williamson RE, Wasteneys GO. Gibberellins induced changes in growth anisotropy precede gibberellins dependent changes in cortical microtubule orientation in developing epidermal cells of barley leaves. Kinematics and cytological studies on gibberellins-responsive dwarf mutant, M489. Plant Physiol. 2000;124:813–22.
Elsinger W. Regulation of pea internode expansion by ethylene. Ann Rev Plant Physiol. 1983;34:225–40.
Potter TI, Zanewich KP, Rood SB. Gibberellin biosynthesis of safflower. Endogenous gibberellins and response to gibberellic acid. Plant Growth Regul. 1993;12:133–40.
Aguirre R, Blanco A. Pattern of histological differentiation induced by paclobutrazol and GA3 in peach shoots. Acta Hort. 1992;315:7–12.
Feng Z, Guo A, Feng Z. Amelioration of chilling stress by triadimefon in cucumber seedling. Plant Growth Regul. 2003;39:277–83.
Manivannan P, Jaleel CA, Kishorekumar A, Sankar B, Somasundaram R, Panneerselvam R. Protection of Vigna unguiculata (L.) Walp. plants from salt stress by paclobutrazol. Colloids Surf B. 2008;61(2):315–8.
Mahsa S, Ali T, Haniye H, Yahya S. Effect of drought stress and paclobutrazol- treated seeds on physiological response of Festuca arundinacea L. Master and Lolium perenne L. Barrage. J Biol Environ Sci. 2011;5(14):77–85.
Nayyar H, Gupta D. Differential sensitivity of C3 and C4 plants to water deficit stress: association with oxidative stress and antioxidants. Environ. and Exp. Bot. 2006;58(1–3):106–13.
Tahir FM, Ibrahim M, Hamid K. Seasonal variation in nutrient concentration of bearing and non-bearing terminals in mango. Asian J Plant Sci. 2003;2(1):113–5.
Chartzoulakis K, Patakas A, Kofidis G, Bosabalidis A, Nastou A. Water stress affects leaf anatomy, gas exchange, water relations and growth of two avocado cultivars. Scientia Horticult. 2002;95:39–50.
Sankar B, Jaleel CA, Manivannan P, Kishorekumar A, Somasundaram R, Panneeelvam R. Effect of paclobutrazol on water stress amelioration through antioxidants and free radical scavenging enzymes in Arachis hypogaea L. Colloids Surf B. 2007;60:229–35.
Swietlik D, Miller SS. The effects of paclobutrazol on growth and response to water stress of apple seedlings. J Amer Soc For Hort Sc. 1983;108:1076.
Barnes AM, Walser RH, Davis TD. Anatomy of Zea mays and Glycine max seedlings treated with triazole plant growth regulators. Biol Plant. 1989;31:370–5.
Protacio CM, Bugante RD, Quinto J, Molinyawe G, Paelmo G. Regulation of flowering in Carabao mango tree by paclobutrazol. Philipp J Crop Sci. 2000;25:27–33.
Blaikie SJ, Kulkarni VJ, Muller WJ. Effects of morphactin and paclobutrazol flowering treatments on shoot and root phenology in mango cv. Kensington Pride. Sci Hortic. 2004;101:51–68.
Nafees M, Faqeer M, Ahmad S, Alam KM, Jamil M, Naveed A. Paclobutrazol soil drenching suppresses vegetative growth, reduces malformation, and increases production in mango. Int J Fruit Sci. 2010;10:431–40.
Burondkar MM, Rajan S, Upreti KK, Reddy YTN, Singh VK, Sabale SN, Naik MM, Ngade PM, Saxena P. Advancing Alphonso mango harvest season in lateritic rockysoils of Konkan region through manipulation in time of paclobutrazol application. J Appl Hortic. 2013;15:178–82.
Voon CH, Pitakpaivan C, Tan SJ. Mango cropping manipulation with cultar. Acta Hort. 1991;291:219–28.
Iglesias DJ, Cercos M, Olmenero-Flores JM, Naranjo MA, Rios G, Carrera E, Ruiz-Rivero O, Lliso I, Morillon R, Tadeo FR, Talon M. Physiology of citrus fruiting. Braz J Plant Physiol. 2007;19:333–62.
Adil OS, Rahim A, Elamin OM, Bangerth FK. Effects of paclobutrazol (PBZ) on floral induction and associated hormonal and metabolic changes of beinnially bearing mango (Mangifera indica L.) cultivars during off year. ARPN J Agric Biol Sci. 2011;6:55–67.
Tomer E. Inhibition of flowering in mango by Gibberellic acid. Sci Hort. 1984;24:299–303.
Kurian RM, Iyer CPA. Stem anatomical characteristics in relation to tree vigor in mango (Mangifera indica L.). Sci Hort. 1992;50:245–53.
Negi ND, Sharma N. Effect of paclobutrazol application and planting systems on growth and production of peach (Prunus persica). Indian J Agric Sci. 2009;79:1010–2.
Husen S, Kuswanto SA, Basuki N. Induction of flowering and yield of mango hybrids using paclobutrazol. J Agric Food Technol. 2012;2:153–8.
Martinez-Fuentes A, Mesejo C, Munoz-Fambuena N, Reig C, Gonzalez-Mas MC, Iglesias DJ. Fruit load restricts the flowering promotion effect of paclobutrazol in alternate bearing Citrus spp. Sci Hortic. 2013;151:122–7.
Singh Z. Effect of (2RS, 3RS) paclobutrazol on tree vigour, flowering, fruit set and yield in mango. Acta Hort. 2000;525:459–62.
Kulkarni VJ, Hamilthon D, Mc Mahon G. Flowering and fruiting in mangoes in top end with paclobutrazol. Crops, Forestry and Horticulture, Darwin, Northern Territory Govt. 2006. pp 1–3.
Souza-Machado V, Pitblado R, Ali A, May P, Bieche BJ. Paclobutrazol in tomato (Lycopersicon esculentum) for improved tolerance to early transplanting and earlier harvest maturity. Acta Hort. 1999;487:139–43.
Giovinazzo R, Souza-Machado V, Hartz TK. Paclobutrazol responses with processing tomato in France. Acta Hort. 2001;542:355–8.
Mohamed GF, Agamy RA, Rady MM. Ameliorative effects of some antioxidants on water-stressed tomato (Lycopersicon esculentum Mill.) plants. J Appl Sci Res. 2011;7:2470–8.
Medonca V, Araujo Neto SE, Ramos JD, Pio R, Souza PA. Production and quality of Tommy Atkins mango fruits after application of paclobutrazol. Rev Cienc Agr. 2002;38:39–149.
Balamani V, Poovaiah BW. Retardation of shoot growth and promotion of tuber growth of potato plants by paclobutrazol. Am Potato J. 1985;62:363–9.
Bandara PMS, Tanino KK. Paclobutrazol enhanced minituber production in Norland potatoes. J Plant Growth Regul. 1995;14:151–5.
Kianmehr B, Otroshy M, Parsa M, Mohallati MN, Moradi K. Effect of Plant Growth Regulation during in vitro Phase on Potato Minituber Production. Int J Agri Crop Sci. 2012;4(15):1060–7.
Medina R, Burgos A, Difranco V, Mroginski L, Cenoz P. Effects of chlorocholine chloride and paclobutrazol on cassava (Manihot esculenta Crantz cv. Rocha) plant growth and tuberous root quality. Agriscientia. 2012;29:51–8.
Samy MM, El Aal AA, Khalil MM. The impact of some treatments on improving seed potato production in the summer season. Middle East J. 2014;3(4):1065–73.
Gopi R, Sridharan R, Somasundaram R, Lakshmanan GMA, Panneerselvam R. Growth and photosynthetic characteristics as affected by triazoles in Amorphophallus campanulatus Blume. Gen Appl Plant Physiol. 2005;31(3–4):171–80.
Jain SK, Sing R, Misra KK. Effect of paclobutrazol on growth, yield and fruit quality of lemon (Citrus limon). Indian J Agric Sci. 2002;72:488–90.
Lolaei A, Kaviani B, Khorrami Raad M, Rezaei MA, Maghsoudi M. Effect of Paclobutrazol and salinity on vegetative and sexual growth and fruit quality of strawberry (Fragaria × Ananassa Duch. cv. Selva). Ann Biol Res. 2012;3(10):4663–7.
Arzani K, Bahadori F, Piri S. Paclobutrazol reduces vegetative growth and enhances flowering and fruiting of mature ‘J. H. Hale’ and ‘Red Skin’ Peach Trees. Hort Env Biotech. 2009;50:84–93.
Marini RP. Growth and cropping of ‘Red haven’ peach trees following foliar applications of flurprimidol and paclobutrazol. J Amer Soc Hort Sci. 1986;111:849–53.
Webster AD, Quinlan JD, Richardson PJ. The influence of paclobutrazol on growth and cropping of sweet cherry cultivar. The effect of annual soil treatments on the growth and cropping of cultivar ‘early rivers’. J Hortic Sci. 1986;61:471–8.
Yang QS, Cao XH. Effects of CPPU and paclobutrazol on yield increase in cassava. J South Agric. 2011;42:594–8.
Appeldoorn NJG, De Bruijn SM, Koot-Gronsveld EAM, Visser RGF, Vreugdenhil D, Van der plas LHW. Developmental changes of enzymes involved in sucrose to hexose-phosphate conversion during early tuberization of potato. Planta. 1997;202:220–6.
Sivakumar T, Alagu Lakshmanan GM, Murali PV, Panneerselvam R. Alteration of antioxidative metabolism induced by triazole in sweet potato. J Exp Sci. 2010;1(3):10–3.
Tuna AL. Influence of foliarly applied different triazole compounds on growth, nutrition, and antioxidant enzyme activities in tomato (Solanum lycopersicum L.) under salt stress. Aust J Crop Sci. 2014;8(1):71–9.
Jaleel CA, Gopi R, Panneerselvam R. Biochemical alterations in white yam (Dioscorea rotundata Poir.) under triazole fungicides: impacts on tuber quality. Czech J Food Sci. 2008;26:298–307.
Acknowledgements
We acknowledge horticulture department of Debre Berhan university.
Funding
There is no funding granted.
Author information
Authors and Affiliations
Contributions
BDA is the first author of the review article, whereas the co-author has contributed equally for the literature collection, manuscript documentation and its revision. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
It is not applicable.
Consent for publication
The authors confirm that the content of the manuscript has not been published, or submitted for publication elsewhere.
Competing interests
There are no competing interests between authors.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Desta, B., Amare, G. Paclobutrazol as a plant growth regulator. Chem. Biol. Technol. Agric. 8, 1 (2021). https://doi.org/10.1186/s40538-020-00199-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40538-020-00199-z