
Abstract
Background
Leaf morphology and epidermal characters are important for phylogenetic and taxonomic studies of many plants, but there is currently insufficient data to use them to help distinguish species of Gagea, which is a taxonomically difficult genus mainly due to polyploidy and hybridization. Therefore, leaf morphology and epidermal characters of Gagea were studied to assess the characters that can be used to elucidate the taxonomy and systematics of 14 species of Gagea collected in Xinjiang, China. Using light microscopy (LM), six qualitative and three quantitative leaf epidermal anatomical characters were examined for both the adaxial and abaxial surfaces. Hierarchical cluster analysis (HCA) was employed to reveal the similarities based on leaf morphology and epidermal characters of the investigated species.
Results
Basal leaf of these species can be terete or flat, and it is triangle, flat, or circular in transverse section. Anticlinal wall patterns of the leaf epidermal cells were straight and sinuous, and only three species had epidermal hairs. Shape of long cells varies, ranging from quadrangular to irregular. HCA revealed that the 14 species could be divided into two groups. Group A was arranged into three subgroups (A1, A2 and A3), based on the Euclidean distance of 6.96. Subgroup A1 consisted of three species with indumentum; subgroup A2 had four species with sinuous type anticlinal walls; and subgroup A3 comprised of two species with a fistulose basal leaf. Group B included five species with short cells.
Conclusions
Leaf morphology and epidermal characters did not differ significantly among populations of the same species in Gagea, whereas they differ significantly among species. Thus, leaf morphology and epidermal characters provide diagnostic information for differentiating G. nigra and G. filiformis; G. altaica, G. jensii and G. alberti, which are morphologically similar species.
Similar content being viewed by others

Background
Leaves are crucial vegetative organs that mostly are exposed to aerial conditions. They embody the adaptive survival strategies of plants during the long-term evolution of species (Wang et al. 2021). The use of leaf morphological and epidermal characters has long been recognized as an important tool in species taxonomy (Deng et al. 2017; Filartiga et al. 2022; Khan et al. 2022).
The morphological characters of plant leaves serve as a useful instrument for infrageneric classification and species circumscription, and sometimes they also provide significant information regarding the phylogeny and taxonomy of closely related species (Stace 1969; Baranova 1992; Meng et al. 2016; Ullah et al. 2018; Shaheen et al. 2021; Bashir et al. 2020). High intraspecific consistency may be shown by foliar epidermal characters, although diversity is seen at the interspecific and higher taxonomic levels (Wang et al. 2015; Mohtashamian et al. 2017). Therefore, leaf epidermal features, such as epidermal cell shape, stomatal type, size and distribution, and trichome type and distribution, have played an essential role in distinguishing those families and genera where identification is complicated, such as Salicaceae (Ghahremaninejad et al. 2012), Caryophyllaceae (Ullah et al. 2018), Fabaceae (Silva et al. 2018; Shaheen et al. 2021; Zhao et al. 2022), Lamiaceae (Gul et al. 2019) and Poaceae (Khan et al. 2017).
Gagea Salisb. a genus of Liliaceae and tribe Tulipeae (Patterson and Givnish 2022; Peruzzi 2016), is widely distributed from the Mediterranean region throughout Europe and Asia (Zarrei et al. 2007; Peterson et al. 2008, 2019), and plants grow in dry and grasslands, rocky slopes, alpine meadows, open shrublands, and deciduous forest understories (Chen and Turland 2000; Peruzzi 2016; Peterson et al. 2019; Kurbaniyazova et al. 2022). The western Pamir-Alai (97 species) and western Tien-Shan Mountains (65 species) are diversification centers of Gagea (Peterson et al. 2011). Due to the wide variety of Gagea species, the short, ephemeral growth phase, and complex morphological characters, e.g. an overlap in primitive and advanced morphological characters (the formation of subterranean organs and the number of ground leaves) (Levichev 1999; Peterson et al. 2004), it is difficult to distinguish/identify similar species. Previous studies have investigated different aspects of Gagea, including bulb structure (Levichev 1999), morphology and ontogeny (Levichev 2001, 2006; Tison et al. 2013), classical cytotaxonomy (Peruzzi 2003, 2008, 2012; Peruzzi and Aquaro 2005), embryology (Greilhuber et al. 2000; Caparelli et al. 2006), pollen morphology (Kosenko 1999; Zarrei and Zarre 2005; Hu et al. 2021; Sezer and Yildiz 2021; Lin et al. 2023), and molecular systematics (Peterson et al. 2004, 2008, 2016; Peruzzi et al. 2008a, b; Zarrei et al. 2009).
Leaf epidermal morphology in Gagea has reported for only 11 species (Elwan 2008; Wang et al. 2013), while anatomical features of the basal leaf has been examined in 22 species (Zarrei et al. 2010). Even at the sectional level, examining the anatomy of the basal leaf and shoot structure in Gagea species, provides key taxonomic insights, with crucial characteristics such as the transverse section contour of the leaf, the number of vascular bundles, and the degree of enrichment in the pith providing profound insights for taxonomic research (Zarrei et al. 2010; Levichev 2013). However, due to the low number of species from China, Egypt or Sweden for which epidermal anatomical studies have been conducted, it has not been possible to make intraspecific and interspecific comparisons and thus to explore their taxonomic significance. Thus, we utilized multiple samples of Gagea from Xinjiang, China, to investigate leaf and their epidermal morphology by light microscopy (LM), and then we compared the leaf epidermal characters among populations and species to assess their taxonomic significance. Regarding certain species with similar macro-morphology, such as G. nigra and G. filiformis from the sect. Minimae, Peterson et al. (2011) clearly distinguished G. nigra from G. filiformis using morphological and molecular evidence. Our objective was to confirm this differentiation through micro-morphology. Similarly, within the sect. Plecostigma, G. altaica, G. jensii, and G. alberti are grouped together, where it was observed that the morphological similarities between G. jensii and G. alberti. Our aim was to determine whether the micro-morphology could be used to distinguish between these species.
Methods
Leaves of the 14 Gagea species were collected from plants growing various sites and habitats in Xinjiang, China, and the number of populations for each species ranged from one to six with at least five populations for widespread species (such as G. bulbifera, G. fedtschenkoana and G. nigra) (Table S1; in total 50 populations of Gagea were sampled). If there were no differences in leaf morphology and epidermal characters between populations of a species, one population was randomly selected as representative of the species. If populations of a species differed, all populations of the species were studied individually. After collection, the species were identified by comparing their morphological characters to those listed in the Flora of China (Chen and Turland 2000); Peterson et al. (2011). The scientific names of species were standardized according to Plants of the World Online (https://powo.science.kew.org).
Basal leaf materials
In each natural population for a species, a mature and healthy basal leaf was collected from 20 individuals. While in the field, half of each fresh basal leaf was preserved in FAA solution (70% ethyl alcohol, 40% formalin and 40% glacial acetic acid), and the other half was taken back to the laboratory to observe basal leaf size, shape, and state and to photograph a transverse section of the middle portion of the basal leaf with a Nikon SMZ-25 (SMZ25, Japan). The herbarium of Xinjiang Agricultural University (XJA) contains the voucher material for the 14 species of Gagea, as listed in Table 1.
Leaf epidermal slides observation using light microscope
For isolating the leaf epidermal layer, we used the method of Wen (1995) with some adjustments. Leaves were removed from the FAA fixative and a 1 cm2 area was taken from it at the middle of the leaf near the midvein. To remove the adaxial and abaxial epidermal layers, the leaf tissue was soaked in a 50% NaClO solution, washed with water and then the two surface layers were torn off and placed on separate slides. Leaf epidermal tissue was stained in a solution of 1% saffron (50% ethyl alcohol) for 2 hours, dehydrated in an ethanol series, and placed in xylene, which made the tissue transparent, and finally sealed with a cover slip using Canadian balsam.
All slides were labeled, including the voucher number. Five slides of the adaxial and abaxial leaf surface were prepared for each specimen collected in the field, and they were examined with a Nikon Eclipse (80i, Japan) microscope by using an objective lens (at 10× magnification). For each slide, qualitative and quantitative data were recorded. For each slide, qualitative and quantitative characters were recorded; a total of 20 characters for each slide. The five qualitative characters observed were: pattern of anticlinal walls (Ant), short cells (S) present or absent, indumentum (Ind), shape of long cells (Shape) and stomatal type, and four quantitative characters were: long cells size (L), stomata size (St), stomatal index (SI) and density of epidermal cell were observed. Terminology for the description of epidermal cells and stomata follows Barthlott et al. (1998) and Carpenter (2005). Long cells were defined as having a width: length ratio > 3, whereas short cells had a width: length ratio < 3. SI was calculated using the following formula: SI = [S/S + E] × 100, where S = number of stomata per unit area; E = number of epidermal cells per unit area.
Data analysis
SPSS program version 26 was used to test for normality and homogeneity of the data for leaf epidermal characters to satisfy the requirements of one-way analysis of variance (ANOVA). The differences among different populations of the same species were analyzed by a one-way ANOVA. Differences among species were determined by the non-parametric Kruskal-Wallis test.
Hierarchical cluster analysis (HCA) was employed to reveal similarities among 14 species of Gagea in leaf morphology and epidermal characters, as well as taxonomic relationships. Quantitative characters are represented by mean ± standard error (Mean ± SE), whereas qualitative characters were given a specific number in the data matrix. Six qualitative (pattern of anticlinal walls, short cells present or absent, indumentum, shape of long cells, transverse section of basal leaf, cauline leaves present or absent) and three quantitative (long cells size, stomata size and stomatal index) leaf morphology and epidermal characters were evaluated in the comparative analysis for their value in distinguishing the studied Gagea species. HCA based on Euclidean distance after normalizing the original data, and the data were clustered using Ward’s method in Origin 2021 software to reflect similarities and differences and achieve data visualization (Ye et al. 2015).
Results
The observations of leaf morphology and epidermal characters revealed that while qualitative characters remained consistent across various populations, continuous modifications were observed in the quantitative aspects of leaf morphology and epidermal characters. However, there were differences in qualitative leaf morphology and epidermal characters among species and quantitative characters were intermittently distributed among species.
Basal leaf morphology and epidermal characters was studied for 14 species of Gagea (Tables 2, 3 and 4). Individual plants and a transverse section of a basal leaf are presented in Fig. 1, and selected LM micrographs of the adaxial and abaxial leaf surface are shown in Fig. 2.
Morphology of basal leaf
Leaf morphological of the studied species showed two characters: eight species had cauline leaves (Fig. 1A-C, I, J, L-N) and six species did not (Fig. 1D-H, K). Length and width of basal leaf ranged from 54.12 ± 3.67 to 189.76 ± 15.49 mm and from 0.70 ± 0.04 to 4.65 ± 0.22 mm, respectively (Table 2). Longest basal leaf was found in G. jensii, whereas the shortest one was found in G. bulbifera (Table 2). Widest basal leaf was found in G. granulosa, whereas the narrowest one was found in G. bulbifera (Table 2). The basal leaf of G. altaica (Fig. 1B), G. bulbifera (Fig. 1C), G. divaricata (Fig. 1D), G. filiformis (Fig. 1F), G. kunawurensis (Fig. 1L) and G. tenera (Fig. 1N) was terete, while that of the other species was flat (Table 2). A transverse section of basal leaf was triangular (Fig. 1A1-D1, F1, J1 M1), flat (Fig. 1E1, H1, I1, K1) or circular (Fig. 1G1, L1, N1), or sometimes fistulose (Fig. 1J1).

Individual plant and transverse section of basal leaf of 14 species of Gagea from Xinjiang, China. (A-A1: Gagea alberti; B-B1: G. altaica; C-C1: G. bulbifera; D-D1: G. divaricata; E-E1: G. fedtschenkoana; F-F1: G. filiformis; G-G1: G. fragifera; H-H1: G. granulosa; I-I1: G. jaeschkei; J-J1: G. jensii; K-K1: G. nigra; L-L1: G. kunawurensis; M-M1: G. stepposa; N-N1: G. tenera). Scale bars: 1 mm
Morphology of leaf epidermis
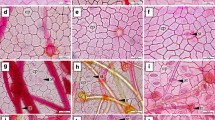
Glandular trichomes were observed on both surfaces of G. stepposa (Fig. 2M, M1), while dendroid-type trichomes were found on the adaxial surface of G. alberti and G. jensii (Fig. 2A, J). Leaf epidermis was comprised of long and short cells, although not all species possessed short cells (Table 4), such as G. filiformis (Fig. 2F, F1), G. fragifera (Fig. 2G, G1), G. kunawurensis (Fig. 2L, L1), G. stepposa (Fig. 2M, M1) and G. tenera (Fig. 2N, N1). Long cells were parallel to the leaf veins and varied from rectangular (Fig. 2A, A1, B, B1, H, H1, J, J1, N, N1), rhomboid (Fig. 2C, C1, D, D1, G, G1, I, I1, M, M1) to irregular (Fig. 2E, E1, F, F1, L, L1). Pattern of anticlinal walls was either straight or sinuous (Table 3). The size of long cells was significantly different between species (H = 87.571, df = 13, p < 0.001). The largest epidermal cells were observed in G. granulosa (Adaxial: 545.82 ± 52.66 × 50.03 ± 1.48 μm, Abaxial: 573.48 ± 17.45 × 37.39 ± 0.94 μm) and smallest ones in G. stepposa (Adaxial: 228.31 ± 7.42 × 24.95 ± 1.28 μm, Abaxial: 254.37 ± 10.33 × 23.08 ± 1.22 μm). Examination of the epidermal cells on both surfaces of Gagea under LM revealed that their morphology was roughly identical (Tables 3 and 4).
Morphology of stomata
Orientation of all stomata on the leaf epidermis of Gagea was roughly consistent (Fig. 2). All species have amphistomatic stomata, i.e. on both surfaces, that are paracytic (Fig. 2). Stomatal size (H = 91.717, df = 13, p < 0.001) and stomatal index (H = 93.654, df = 13, p < 0.001) differed significantly between species. Stomatal index (SI) ranged from 21.05 to 39.17% on adaxial surface in G. filiformis and G. fedtschenkoana respectively, whereas 22.83–39.94% on abaxial surface in G. kunawurensis and G. divaricata respectively (Table 4). Size of stomata ranged from 45.66 ± 1.31 × 28.77 ± 0.92 to 82.16 ± 2.39 × 44.16 ± 0.77 μm on adaxial surface of G. bulbifera and G. tenera, respectively, whereas on abaxial surface size ranged from 43.74 ± 1.38 × 26.80 ± 1.08 μm to 92.82 ± 5.17 × 42.30 ± 1.42 μm in G. bulbifera and G. tenera, respectively (Table 4).

Epidermal morphology of adaxial (A-N) and abaxial (A1-N1) surfaces of basal leaf in 14 species of Gagea from Xinjiang, China under light microscope (A-A1: Gagea alberti; B-B1: G. altaica; C-C1: G. bulbifera; D-D1: G. divaricata; E-E1: G. fedtschenkoana; F-F1: G. filiformis; G-G1: G. fragifera; H-H1: G. granulosa; I-I1: G. jaeschkei; J-J1: G. jensii; K-K1: G. nigra; L-L1: G. kunawurensis; M-M1: G. stepposa; N-N1: G. tenera). Scale bars: 100 μm
Hierarchical cluster analysis
A hierarchical clustering dendrogram shows the relationship between the 14 species based on leaf morphological and epidermal characters (Fig. 3). The obtained tree separated the species into two groups based on a Euclidean distance of 8.18. Group A was composed of nine species: G. alberti, G. filiformis, G. fragifera, G. granulosa, G. jensii, G. kunawurensis, G. nigra, G. stepposa and G. tenera. Further, Group A was arranged into three subgroups (A1, A2 and A3), based on a Euclidean distance of 6.96. Whereas group B included G. altaica, G. bulbifera, G. divaricata, G. fedtschenkoana and G. jaeschkei (Fig. 3).

Cluster diagram (Ward’s linkage) of 14 species in Gagea from Xinjiang, China, based on six qualitative and three quantitative morphological and epidermal characters of leaves
Discussion
The present study provides some baseline data about leaf morphology and leaf epidermal anatomy characters of Gagea from Xinjiang, China. Observations of the leaf morphology and epidermal characters of 14 species of Gagea revealed the following characters: (1) except for G. fragifera and G. tenera, the basal leaf of most species was solid; (2) the adaxial and abaxial epidermal cells were mostly quadrilateral but a few are irregular; (3) the leaf epidermal layer consists of long and short cells, but G. filiformis, G. fragifera, G. kunawurensis, G. stepposa and G. tenera do not have short cells; (4) the pattern of anticlinal walls was sinuous or straight; (5) orientations of all stomatal in the leaf epidermis are roughly identical; (6) the basal leaf of G. alberti, G. jensii and G. stepposa is hairy. The indumentum (hairiness) of basal leaf can be divided into two types: glandular trichomes found in G. stepposa and dendroid-type found in all the other species. The leaf epidermis of Gagea shows sinuous or straight patterns of anticlinal walls, rectangular, rhomboid or irregular shape of the long cells, and presence of long and short cells in some species, and these are key characters that can be used for classification and identification of Gagea. These findings emphasize the importance of leaf morphology and epidermal characters in the taxonomic study of Gagea.
Wang et al. (2013) reported differences in leaf epidermal characters, including the microstructure of epidermal cells and stomata for five species of Gagea, each represented by a single population in China. In our comparative study of the five species studied by Wang et al. (2013), we had total of 24 populations for the five species, and we found interspecific differences but no significant differences within and among conspecific populations. Additionally, Elwan (2008) also demonstrated interspecific differences in leaf epidermal characters. These findings reflect the stability of leaf epidermal morphology within populations of the same species. For monocotyledons, the Liliaceae differs considerably from the Asparagaceae and Poaceae in terms of leaf epidermal characters (Aliscioni et al. 2015; Meng et al. 2016; Chao et al. 2022). The types of anticlinal wall of Fritillaria (Wang et al. 2009) and Lilium (Hou et al. 2015) in the Liliaceae are straight or curve, and types of anticlinal walls observed in Gagea are consistent with these genera. However, in addition to rectangular or rhombic shapes of long cells, serrated cells also are present in Fritillaria and Lilium, but are not present in Gagea (Wang et al. 2009; Hou et al. 2015). The stomata of Liliaceae exhibit consistent orientation and shape, providing significant taxonomic value among monocotyledon (Wang et al. 2009; Doğu 2019).
Leaf epidermal characteristics within each section are generally highly consistent. For instance, in sect. Didymobulbos, the basal leaves are typically circular, flattened, or fistulose in transverse section, with 1–2 basal leaves present. Similarly, sect. Plecostigma has 1–2 basal leaves that are circular in transverse section, while sect. Platyspermum has a single basal leaf that is pentagonal in transverse section; sect. Gagea features 1–2 basal leaves that are flattened in transverse section (Zarrei et al. 2010, 2011). Our research corroborates these results, demonstrating that the consistency in leaf epidermal characteristics within each section yields valuable insights for the classification and identification of Gagea.
Currently, classification studies of most species are conducted from a molecular perspective, which can achieve classification results at the genetic level. However, compared to external macroscopic morphological characters, there are limitations in practical species identification (Shneyer et al. 2003; Hou et al. 2015). Morphologically, the Gagea species are discriminated using information on bulbs, leaves, inflorescences, tepals, stigmas and seeds traits (Chen and Turland 2000). The level of variation in anatomical characters is the highest among the basal leaf (Zarrei et al. 2010). In contrast, classification based on leaf morphology and epidermal characters is a relatively intuitive and practical method. Species with similar external morphology can be inferred from leaf morphological and epidermal characters, which have been reported in Ornithogalum (Peruzzi et al. 2007), Fritillaria (Wang et al. 2009), Maianthemum (Meng et al. 2016), Impatiens (Rahman et al. 2017), Aspidistra (Vislobokov et al. 2021) and Zingiber (Zhao et al. 2022). An extensive field investigation and examination of specimens from Xinjiang found that G. nigra and G. filiformis; G. jensii, G. altaica and G. alberti have similar morphological characters, such as bulbs, inflorescences, tepals, stigmas and seeds. Thus, these species are closely related natural taxa and support their placement in the same section (Tison et al. 2013; Peterson et al. 2016). The absence of short cells in G. filiformis (Fig. 2F, F1) can be used to distinguish this species from G. nigra (Fig. 2K, K1). The leaf morphology is consistent with Peterson et al.’s (2016) classification of G. nigra. The sinuous pattern of anticlinal walls of G. altaica (Fig. 2B, B1) also can be used to separate this species from G. jensii (Fig. 2J, J1) and G. alberti (Fig. 2A, A1). Remaining species also can be distinguished based on their pollen morphology, except for G. jensii (Lin et al. 2023). Morphologically, G. jensii and G. alberti exhibit slight differences; G. jensii has a denser inflorescence and larger lateral flowers, as stated by Peterson et al. (2011). Nonetheless, it is difficult to distinguish two species based solely on pollen grains (Lin et al. 2023) and leaf epidermis, and employment of cytological or molecular evidence to identify G. jensii, is necessary. Classification research based on leaf and epidermal characters can not only reflect common characters of Gagea but also interspecific variability. Therefore, leaf morphology and epidermal characters are considered highly valuable for the systematic classification of Gagea.
Conclusion
This study is the first to combine the leaf structure of Gagea with detailed epidermal characters. While not all variable traits hold systematic or taxonomic significance, a considerable number of them play a crucial role in species differentiation, such as the shape of leaf epidermal long cells. The division of species with similar external morphology is further supported by phylogenetic and palynological evidence. The leaf and its epidermal characters provide evidence for species classification, with significant differences in epidermal characters that can help resolve delimitation of species with similar appearances in Gagea. Consequently, these characters hold systematic significance and enhance our understanding of Gagea.
Data Availability
All data generated or analyzed during this study are included in this published article and its additional files.
References
Aliscioni SS, Ospina JC, Gomiz NE (2016) Morphology and leaf anatomy of Setaria s.l. (Poaceae: Panicoideae: Paniceae) and its taxonomic significance. Plant Syst Evol 302:1–13. https://doi.org/10.1007/s00606-015-1251-9
Baranova M (1992) Principles of comparative stomatographic studies of flowering plants. Bot Rev 58:49–99. https://doi.org/10.1007/BF02858543
Barthlott W, Neinhuis C, Cutler D, Ditsch F, Meusel I, Theisen I, Wilhelmi H (1998) Classification and terminology of plant epicuticular waxes. Bot J Linnean Soc 126:237–260. https://doi.org/10.1111/j.1095-8339.1998.tb02529.x
Bashir K, Sohail A, Ali U, Ullah A, Haq ZU, Gul B, Ullah I, Asghar M (2020) Foliar micromorphology and its role in identification of the Apocynaceae taxa. Microsc Res Tech 83(7):1–29. https://doi.org/10.1002/jemt.23466
Caparelli KF, Peruzzi L, Cesca G (2006) A comparative analysis of embryo-sac development in three closely-related Gagea species (Liliaceae), with some considerations on their reproductive strategies. Plant Biosyst 140:115–122. https://doi.org/10.1080/11263500600756090
Carpenter KJ (2005) Stomatal architecture and evolution in basal angiosperms. Am J Bot 92:1595–1615. https://doi.org/10.3732/ajb.92.10.1595
Chao CT, Tzeng HY, Tseng YH (2022) Leaf epidermal morphology of Asparagaceae of Taiwan and its systematic significance. Microsc Res Tech 85:2162–2180. https://doi.org/10.1002/jemt.24074
Chen XQ, Turland NJ (2000) Flora of China, vol 24. Science Press, Beijing
Deng M, Jiang XL, Song YG, Coombes A, Yang XR, Xiong YS, Li QS (2017) Leaf epidermal features of Quercus Group Ilex (Fagaceae) and their application to species identification. Rev Palaeobot Palynology 237:10–36. https://doi.org/10.1016/j.revpalbo.2016.11.006
Doğu S (2019) Comparative anatomical studies of four Hyacinthella Schur species growing in Turkey. Bangladesh J Bot 48:981–987. https://doi.org/10.3329/bjb.v48i4.49038
Elwan ZA (2008) On the taxonomy of Gagea and Calochortus (Liliaceae): evidences from macromophological aspects and cuticular features of leaf. Res J Agric Biol Sci 4:1–15
Filartiga AL, Klime A, Altman J, Nobis MP, Crivellaro A, Schweingruber F, Doleal J (2022) Comparative anatomy of leaf petioles in temperate trees and shrubs: the role of plant size, environment and phylogeny. Ann Bot 129:567–582. https://doi.org/10.1093/aob/mcac014
Ghahremaninejad F, Khalili Z, Maassoumi AA, Mirzaie-Nodoushan H, Riahi M (2012) Leaf epidermal features of Salix species (Salicaceae) and their systematic significance. Am J Bot 99:769–777. https://doi.org/10.3732/ajb.1100019
Greilhuber J, Ebert I, Lorenz A, Vyskot B (2000) Origin of facultative heterochromatin in the endosperm of Gagea lutea (Liliaceae). Protoplasma 212:217–226. https://doi.org/10.1007/BF01282922
Gul S, Ahmad M, Zafar M, Bahadur S, Ayaz A (2019) Taxonomic significance of foliar epidermal morphology in Lamiaceae from Pakistan. Microsc Res Tech 82:1507–1528. https://doi.org/10.1002/jemt.23316
Hou J, Xiao FF, Zhang YL, Liu LX, Yan YX (2015) Structural characters of leaf epidermal and their systematic significance in the 20 wild Lilium species of China. Acta Bot Boreal-Occident Sin 35:1560–1573. https://doi.org/10.7606/j.issn.1000-4025.2015.08.1560
Hu ZM, Zhao CH, Zhao YY, Liu JX (2021) Pollen morphology of Liliaceae and its systematic significance. Palynology 45(3):531–568. https://doi.org/10.1080/01916122.2021.1882601
Khan R, Ahmad M, Zafar M, Ullah A (2017) Scanning electron and light microscopy of foliar epidermal characters: a tool for plant taxonomists in the identification of grasses. Microsc Res Tech 80:1–18. https://doi.org/10.1002/jemt.22909
Khan A, Shen F, Yang L, Xing W, Clothier B (2022) Limited acclimation in leaf morphology and anatomy to experimental drought in temperate forest species. Biology 11:1186. https://doi.org/10.3390/biology11081186
Kosenko VN (1999) Contributions to the pollen morphology and taxonomy of the Liliaceae. Grana 38(1):20–30. https://doi.org/10.1080/001731300750044672
Kurbaniyazova GT, Levichev IG, Kadirov UK (2022) The species of the Genus Gagea Salisb. Is distribution in the Flora of the Urgut Region (Uzbekistan). Am J Plant Sci 13(9):1183–1195. https://doi.org/10.4236/ajps.2022.139080
Leviche IG (2001) New species of the genus Gagea Salisb (Liliaceae) from western regions of Asia (in Russian). Turczaninowia 4:5–35
Levichev IG (1999) Zur Morphologie in Der Gattung Gagea Salisb. (Liliaceae). I. die unterirdischen organe. Flora 194:379–392. https://doi.org/10.1016/S0367-2530(17)30929-5
Levichev IG (2006) A review of the Gagea (Liliaceae) species in the flora of Caucasus. Botanicheskii Zhurnal 91:917–951
Levichev IG (2013) Structural features of shoots in Lloydia, Gagea, Kharkevichia (Liliaceae) as evolutionary variability of the modules of mesome nature in monocotyledons. Botanicheskii Zhurnal 98:409–452
Lin MS, Qiu J, Xie KQ, Tan DY (2023) Palynological features and taxonomic significance for 16 species of Gagea (Liliaceae) from Xinjiang, China. PhytoKeys 225:53–68. https://doi.org/10.3897/phytokeys.225.101518
Meng Y, Wang JJ, Nie EL (2016) Comparative morphology of leaf epidermal in 34 species of Maianthemum (Asparagaceae, Polygonateae) and their systematic significance. Phytotaxa 275:81–96. https://doi.org/10.11646/phytotaxa.275.2.1
Mohtashamian M, Attar F, Kavousi K, Masoudi-Nejad A (2017) Micromorphological studies of leaf epidermal features in populations of maples (Acer L.) from Iran. Phytotaxa 299(1):36–54. https://doi.org/10.11646/phytotaxa.299.1.2
Patterson TB, Givnish TJ (2022) Phylogeny, concerted convergence, and phylogenetic niche Conservatism in the core Liliales: insights from rbcL and ndhF sequence data. Evolution 56:233–252. https://doi.org/10.1111/j.0014-3820.2002.tb01334.x
Peruzzi L (2003) Contribution to cytotaxonomical knowledge of Gagea Salisb. (Liliaceae). Sect. Foliatae A. Terrac. And synthesis of karyological data. Caryologia 56:115–128. https://doi.org/10.1080/00087114.2003.10589314
Peruzzi L (2008) Contribution to the cytotaxonomical knowledge of the genus Gagea Salisb. (Liliaceae). III. New karyological data from the central Mediterranean area. Caryologia 61:92–106. https://doi.org/10.1080/00087114.2008.10589615
Peruzzi L (2012) Chromosome diversity and evolution in Gagea (Liliaceae). Bocconea 24:147–158
Peruzzi L (2016) A new infrafamilial taxonomic setting for Liliaceae, with a key to genera and tribes. Plant Biosyst 150:1341–1347. https://doi.org/10.1080/11263504.2015.1115435
Peruzzi L, Aquaro G (2005) Contribution to the cytotaxonomical knowledge of Gagea Salisb. (Liliaceae). II. Further karyological studies on Italian populations. Candollea 60:237–253
Peruzzi L, Caparelli KF, Cesca G (2007) Contribution to the systematic knowledge of the genus Ornithogalum L. (Hyacinthaceae): morpho-anatomical variability of the leaves among different taxa. Bocconea 21:257–265
Peruzzi L, Peterson A, Tison JM, Peterson J (2008a) Phylogenetic relationships of Gagea Salisb. (Liliaceae) in Italy, inferred from molecular and morphological data matrices. Plant Syst Evol 276:219–234. https://doi.org/10.1007/s00606-008-0081-4
Peruzzi L, Tison JM, Peterson A, Peterson J (2008b) On the phylogenetic position and taxonomic value of Gagea trinervia (Viv.) Greuter and Gagea sect. Anthericoides A. Terracc. (Liliaceae). Taxon 57:1201–1214. https://doi.org/10.1002/tax.574013
Peterson A, John H, Koch E, Peterson J (2004) A molecular phylogeny of the genus Gagea (Liliaceae) in Germany inferred from non-coding chloroplast and nuclear DNA sequences. Plant Syst Evol 245:145–162. https://doi.org/10.1007/s00606-003-0114-y
Peterson A, Levichev IG, Peterson J (2008) Systematics of Gagea and Lloydia (Liliaceae) and infrageneric classification of Gagea based on molecular and morphological data. Mol Phylogenet Evol 46(2):446–465. https://doi.org/10.1016/j.ympev.2007.11.016
Peterson A, Levichev IG, Peterson J, Harpke D, Schnittler M (2011) New insights into the phylogeny and taxonomy of Chinese species of Gagea (Liliaceae)-speciation through hybridization. Org Divers Evol 11:387–407. https://doi.org/10.1007/s13127-011-0059-x
Peterson A, Harpke D, Levichev IG, Beisenova S, Schnittler M, Peterson J (2016) Morphological and molecular investigations of Gagea (Liliaceae) in southeastern Kazakhstan with special reference to putative altitudinal hybrid zones. Plant Syst Evol 302:985–1007. https://doi.org/10.1007/s00606-016-1313-7
Peterson A, Harpke D, Peterson J, Harpke A, Peruzzi L (2019) A pre-miocene Irano‐Turanian cradle: origin and diversification of the species‐rich monocot genus Gagea (Liliaceae). Ecol Evol 9:5870–5890. https://doi.org/10.1002/ece3.5170
Rahman F, Ahmad M, Zafar M, Mumtaz AS, Saha SA (2017) Taxonomic implications of foliar epidermis in Impatiens (Balsaminaceae): Investigating 12 Pakistani taxa as an example. Plant Biosystems-An International Journal Dealing with all Aspects of Plant Biology 151:642–648. https://doi.org/10.1080/11263504.2016.1193068
Sezer O, Yildiz AC (2021) Pollen morphology studies on some Gagea Salisb. (Liliaceae) species from Turkey. Pak J Bot 53(3):1073–1077. https://doi.org/10.30848/PJB2021-3(21)
Shaheen S, Sharifi-Rad J, Ali M, Shamim Z, Rasool B, Ashfaq M, Mukhtar H, Harun N, Hanif U, Siddique R (2021) Light and scanning electron microscopy-based foliar morpho-anatomical comparison of selected family Rosaceae members distributed in District Lahore, Punjab, Pakistan. Microsc Res Tech 85:1597–1610. https://doi.org/10.1002/jemt.24021
Shneyer VS, Kutyavina NG, Pimenov MG (2003) Systematic relationships within and between Peucedanum and Angelica (Umbelliferae-Peucedaneae) inferred from immunological studies of seed proteins. Plant Syst Evol 236:175–194. https://doi.org/10.1007/s00606-002-0239-4
Silva NF, de Oliveira Arruda RDC, Alves FM, Sartori ALB (2018) Leaflet anatomy of the Dipterygeae clade (Faboideae: Fabaceae): evolutionary implications and systematics. Bot J Linnean Soc 187:99–117. https://doi.org/10.1093/botlinnean/boy009
Stace CA (1969) The significance of the leaf epidermal in the taxonomy of the Combretaceae II. The genus Combretum Subgenus Combretum in Africa. Bot J Linnean Soc 62:131–168. https://doi.org/10.1111/j.1095-8339.1969.tb01962.x
Tison JM, Peterson A, Harpke D, Peruzzi L (2013) Reticulate evolution of the critical Mediterranean Gagea sect. Didymobulbos (Liliaceae) and its taxonomic implications. Plant Syst Evol 299(2):413–438. https://doi.org/10.1007/s00606-012-0731-4
Ullah F, Zafar M, Amhad M, Sultana S, Ullah A, Shah SN, Butt MA, Mir S (2018) Taxonomic implications of foliar epidermal characteristics in subfamily Alsinoideae (Caryophyllaceae)-ScienceDirect. Flora 242:31–44. https://doi.org/10.1016/j.flora
Vislobokov NA, Fu LF, Wei YG, Nuraliev MS (2021) Leaf epidermal micromorphology in Aspidistra (Asparagaceae): diversity and taxonomic significance. PhytoKeys 185:65–86. https://doi.org/10.3897/phytokeys.185.72259
Wang Q, Zhou SD, Deng XY, Zheng Q, He XJ (2009) Comparative morphology of the leaf epidermal in Fritillaria (Liliaceae) from China. Bot J Linnean Soc 160:93–109. https://doi.org/10.1111/j.1095-8339.2009.00855.x
Wang H, Zhang WH, Lu YM, Yang F, Wang YX (2013) Leaf epidermal micro-morphology structures of 5 Gagea species from Xinjiang. J Anhui Agri Sci 41:4264–4267
Wang YH, Lu L, Fritsch PW, Wang H, Wang YH, Li DZ (2015) Leaf epidermal character variation and evolution in Gaultherieae (Ericaceae). Bot J Linnean Soc 178:686–710. https://doi.org/10.1111/boj.12296
Wang ZY, Chen XP, Cheng Y, Wang MT, Zhong QLi, Li M, Cheng DL (2021) Leaf and fine root economics spectrum across 49 woody plant species in Wuyi Mountains. Chin J Plant Ecol 45:242–252. https://doi.org/10.17521/cjpe.2020.0280
Wen J (1995) A rapid mathod of isolating and clearing organs on foliar epidermal. Chin Bull Bot 12:77
Ye WP, Min J, Ren K, Yang F, Li W, Chen P, Fang AP (2015) Origin 9.1 scientific graphing and data analysis. China Machine Press, Beijing
Zarre S, Zarrei M (2005) Pollen morphology of the genus Gagea (Liliaceae) in Iran. Flora 200:96–108. https://doi.org/10.1016/j.flora.2004.04.001
Zarrei M, Zarre S, Wilkin P, Rix M (2007) Systematic revision of the genus Gagea Salisb. (Liliaceae) in Iran. Bot J Linnean Soc 154:559–588. https://doi.org/10.1111/j.1095-8339.2007.00678.x
Zarrei M, Wilkin P, Fay MF, Ingrouille MJ, Zarre S, Chase MW (2009) Molecular systematics of Gagea and Lloydia (Liliaceae; Liliales): implications of analyses of nuclear ribosomal and plastid sequences for infrageneric classification. Ann Bot 104:125–142. https://doi.org/10.1093/aob/mcp103
Zarrei M, Wilkin P, Ingrouille MJ, Zarrei S, Chase MV (2010) The systematic importance of anatomical data in Gagea (Liliaceae) from the Flora Iranica area. Bot J Linnean Soc 164:155–177. https://doi.org/10.1111/j.1095-8339.2010.01081.x
Zarrei M, Wilkin P, Ingrouille MJ, Chase MW (2011) A revised infrageneric classification for Gagea (Tulipeae; Liliaceae): insights from DNA sequence and morphological data. Phytotaxa 15:44–56. https://doi.org/10.1186/1471-2229-11-26
Zhao H, Xiao MH, Zhong Y, Wang YQ (2022) Leaf epidermal micromorphology of Zingiber (Zingiberaceae) from China and its systematic significance. PhytoKeys 190:131–146. https://doi.org/10.3897/phytokeys.190.77526
Acknowledgements
We thank Dr Carol C. Baskin (University of Kentucky, Lexington, Kentucky, USA) for correcting the English and we are grateful to Dr. Xiaojun Shi and Dr. Kaiqing Xie (Xinjiang Agricultural University, Ürümqi, China) for their assistance in collecting samples from Gagea populations. We also thank undergraduate student Pulu Wang for his assistance with the experiments on leaf epidermal preparations.
Funding
This research was supported by the Natural Science Foundation of Xinjiang Uygur Autonomous Region of China (2022D01A71), the Innovative Team Foundation of Biology of Xinjiang Agricultural University, China (ITB202103), the Grant of Innovation Environment Construction of the Xinjiang Uygur Autonomous Region, China (PT2315) and the Third Xinjiang Scientific Expedition Program (2022xjkk1505).
Author information
Authors and Affiliations
Contributions
DYT and JQ conceived and designed the study; JQ and MSL performed experiments; MSL analyzed the data; MSL, JQ and DYT interpreted the results of the statistical analyses. All authors wrote the paper. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary Material: Table S1
. The list of plant materials examined and vouchers of 50 populations belonging to 14 species of Gagea from Xinjiang, China
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Qiu, J., Lin, M. & Tan, D. Taxonomic implications of leaf morphology and epidermal anatomy for 14 species of Gagea (Liliaceae) from Xinjiang, China. Bot Stud 64, 33 (2023). https://doi.org/10.1186/s40529-023-00405-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40529-023-00405-9