
Abstract
Background
Central place foragers must acquire resources and return to a central location after foraging bouts. During the egg laying (hereafter laying) period, females are constrained to a nest location, thus they must familiarize themselves with resources available within their incubation ranges after nest site selection. Use of prospecting behaviors by individuals to obtain knowledge and identify profitable (e.g., resource rich) locations on the landscape can impact demographic outcomes. As such, prospecting has been used to evaluate nest site quality both before and during the reproductive period for a variety of species.
Methods
Using GPS data collected from female eastern wild turkeys (Meleagris gallopavo silvestris) across the southeastern United States, we evaluated if prospecting behaviors were occurring during laying and what landcover factors influenced prospecting. Specifically, we quantified areas prospected during the laying period using a cluster analysis and the return frequency (e.g., recess movements) to clustered laying patches (150-m diameter buffer around a clustered laying period location) during the incubation period.
Results
The average proportion of recess movements to prospected locations was 56.9%. Nest fate was positively influenced (μ of posterior distribution with 95% credible 0.19, 0.06–0.37, probability of direction = 99.8%) by the number of patches (90-m diameter buffer around a clustered laying period location) a female visited during incubation recesses. Females selected for areas closer to the nest site, secondary roads, hardwood forest, mixed pine-hardwood forest, water, and shrub/scrub, whereas they avoided pine forest and open-treeless areas.
Conclusions
Our findings suggest that having a diverse suite of clustered laying patches to support incubation recesses is impactful to nest fate. As such, local conditions within prospected locations during incubation may be key to successful reproductive output by wild turkeys. We suggest that prospecting could be important to other phenological periods. Furthermore, future research should evaluate how prospecting for brood-rearing locations may occur before or during the incubation period.
Similar content being viewed by others


Introduction
Central place foragers travel from a central location on foraging excursions and return to that location between foraging bouts [53, 70]. Foraging bouts from centralized locations are known to incur a cost of time, energy, and mortality risk [87]. During incubation, avian species are constrained to nest sites, and their ability to use space is restricted if low-risk loafing and foraging areas are not adequately distributed within their range [39, 43, 80]. Therefore, individuals should familiarize themselves with profitable areas within their incubation ranges that provide reduced risk or energetic benefit [52].
Site prospecting is an exploratory behavior common across taxa which allows animals to determine quality of areas within their ranges that would increase fitness [60, 65, 90]. Prospecting occurs at various time periods (pre-, post-, and during reproduction) during the reproductive season [12, 25, 68]. Among avian species, prospecting behavior has been related to identifying migratory stopover [14, 49] and pre- and post-breeding sites [54, 55, 61, 68]. Gathering information using prospecting behaviors can reduce predation risk, increase foraging efficiency, both contributing to individual fitness and reproductive success [60, 65].
Recursive movements are patterns of returns to previously visited areas which occur when individuals identify resources within a heterogeneous landscape [7, 9, 51, 66]. Recursive movement behaviors benefit fitness by improving forage efficiency [64, 81], increasing predator avoidance [79, 86], or in maintaining territories [36, 41]. For central place foragers, prospecting could be used as the mechanism to identify high-quality foraging areas [56], preceding recursive movement to those profitable areas that were identified on the landscape [7].
The onset of egg laying (hereafter laying) and incubation in avian species is an energetically costly time period during which individuals are spatially constrained [22]. Uniparental incubators are faced with the tradeoff of remaining at the nest site or making recess movements (i.e. directional movements made away from nesting location) to gain resources [74, 83]. Prior to incubation, prospecting by females to familiarize themselves with resource distribution could facilitate efficient travel to and from resources while reducing mortality risk [59, 65, 82]. Therefore, prospecting during the laying period could be important in supporting behavioral strategies used during incubation [59, 65].
Female eastern wild turkeys (Meleagris gallopavo silvestris; hereafter wild turkey) are uniparental ground nesters that maintain ranges, but do not defend territories [32]. During nesting, females are central-place foragers that make foraging bouts from the nest location during incubation [3, 18, 45]. To survive the incubation period, females identify resources that provide foraging opportunities and concealment from predators (Green 1982, [3, 45, 85]. Contemporary research has shown that prior to laying, female wild turkeys do not prospect for potential nest sites [17], but it is plausible that individuals may prospect for resources during the laying period [16]. Furthermore, pre-nesting and laying ranges show little overlap [71], and during laying, females increase daily movements but decrease space use, indicative of a lack of site familiarity [31, 48, 71]. It is plausible that movement behaviors during the laying period may maximize foraging success and reduce predation risk during the incubation period [2, 11]. However, it is unclear if areas identified by females during laying are ultimately selected and visited during incubation when females take incubation recesses [3, 45].
Our objectives were to (1) determine if female wild turkeys returned to locations prospected during the laying period and how many patches were used when making recess movements during incubation, and to (2) assess the relationship between environmental and movement covariates during incubation recesses to areas they prospected during the laying period. Prospecting behavior in avian species is known to enhance foraging ability and reduce predation risk during reproduction by increasing landscape familiarity (Pärt and Doligez 2011). During incubation, avian species must balance incubating the nest and ensuring their own survival, making the identification of resources crucial for maintaining their survival during this period [22]. Notably, recess movements during the incubation period significantly influence wild turkey nest success [3, 45]. Therefore, we hypothesized that incubating females would return to sites previously visited during the laying period when taking incubation recesses, and such behaviors would positively affect nest fate. Specifically, we predicted that females who did not revisit sites previously visited during laying would have lower nest success. If incubation recesses occurred in locations prospected during laying, it could indicate the presence of high-quality forage or closer proximity to the nest, potentially reducing overall movements [3, 45]. Therefore, we hypothesized sites prospected during laying and selected by incubating females would be closer to landcover that provide foraging opportunity but reduce their distance from the nest location. Specifically, we predicted that females would select for pine and open landcover types that are closer to the nest. Implications from our findings offer a novel perspective on the timing of prospecting behavior and its impact on nest fate, providing contributions to the understanding of spatial–temporal resource selection.
Methods
We used rocket nets to capture wild turkeys from January-March of 2014–2021 (For details on study sites refer to Additional file 1). We aged captured individuals based on presence of barring on the ninth and tenth primary feathers and sexed them by the coloration of the breast feathers [58]. We banded each bird with an aluminum rivet leg band (National Band and Tag Company, Newport, Kentucky,female size = 8, male size = 9) and radio-tagged each individual with a backpack-style GPS-VHF transmitter [30] produced by Biotrack Ltd. (Wareham, Dorset, UK). We programmed transmitters to record 1 GPS location nightly (23:58:58) and hourly GPS locations from 0500 to 2000 (Standard Time and according to the appropriate time zones) for the duration of the study [15]. Each transmitter had a mortality switch that was programmed to activate after > 23 h of no movement. We released turkeys immediately at the capture location after processing. All turkey capture, handling, and marking procedures were approved by the Institutional Animal Care and Use Committee at the University of Georgia (Protocol #A2019 01-025-R2 and #A2020 06-018-R1) and the Louisiana State University Agricultural Center (Protocol #A2014-013, A2015-07, and A2018-13).
We located wild turkeys ≥ 2 times per week using a 3-element handheld Yagi antenna and receiver to monitor survival based on the presence of a mortality signal, general movements of individuals within their ranges, and onset of nesting activity. We remotely downloaded GPS locations from each turkey ≥ 1 time per week. In ArcGIS 10.8 (Environment Systems Research Institute, Redlands, California, USA), we spatially projected GPS locations to identify nest locations by determining when a female’s locations became concentrated, which represented the onset of incubation [3, 18]. When VHF tracking and GPS locations indicated nest termination, we located the nest site to determine if hatching had occurred [17, 88, 89].
We processed and cleaned the raw GPS data by removing fix locations that had dilution of precision values (DOP) > 7 which is considered error in the positional fix due to the geometry of the satellite signal received [29]. To determine dates of nest initiation (i.e. initiation of laying) and onset of incubation initiation, we mapped our spatial–temporal data using ArcGIS 10.8 (Environment Systems Research Institute, Redlands, California, USA). We identified the onset of incubation as the first time an individual remained on the nest overnight [3, 45], and then evaluated hourly locations for the previous 20 days to determine when a female initially visited the nest site (defined as location being < 20 m from the known nest site, [17, 18, 71]. We considered the date of first visit as the date of nest initiation and used it as the beginning of the laying period as wild turkeys rarely visit nest sites before laying the first egg [16, 17]. Incubation recesses are directional movements made away from nesting locations during active incubation, which are thought to allow individuals time to acquire necessary resources while maintaining appropriate egg temperatures [22]. Following Bakner et al. [3], we classified recess movements during incubation as any location > 27.5 m (27.5 m is associated with the 90th percentile error of the transmitter) away from the known nest location and all other locations (< 27.5 m) as incubation and not recessing. We calculated the distance each recess location (any point > 27.5 m away from the nest) was from the nest location (distance to nest) to incorporate into our model to evaluate habitat selection. We performed data processing and analysis in program R (v.4.1.0; [63].
Covariates
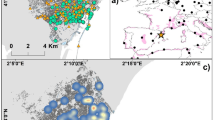
We assigned a unique identification to each female GPS location for the duration of the laying period. To quantify location-specific revisitation for individual females, we combined laying locations that were to the same areas using a cluster analysis in package geosphere in program R [33], v.4.1.0; [63]. Using estimates from Schofield [71] who reported that female wild turkeys moved 300 m/hr during the laying period, we used a 150-m radius buffer of each unique GPS location to perform the cluster analysis (Fig. 1). Specifically, we used a 150-m radius buffer to account for average movement during the laying period, preventing the double-counting of laying period locations and treating multiple locations within the same area as one. The cluster analysis allowed us to associate each female with a set of potential prospecting locations. We then used the clustered laying period locations to quantify how many incubation recesses were made to that area.

GPS locations of a female eastern wild turkey depicting how we determined the CLP covariate and number of incubation recesses. A Laying and incubation movements used for the analysis with a star showing the nest location. B Clustered laying period GPS locations created from the cluster analysis (150 m radius buffer; hereafter, CLP). C The CLP with a 45 m radius buffer determined from how far a female travels during incubation. D Any incubation point that fell within a CLP contributed to the proportion of recesses made to a laying period location. Any CLP that contained an incubation location was considered a CLP that was used
Following Bracis et al. [8], we calculated the revisit rate to evaluate whether incubation recess movements by females were to locations previously visited during the laying period, we used package recurse in R (v.4.1.0,[63]. We first assigned a unique identification to each of the clustered laying patch (hereafter, CLP. We used estimates of daily distance traveled while on an incubation recess from Bakner et al. [3], 90 m) to set an appropriate circular buffer size around each CLP. We then used the function getRecursionsAtLocation in package recurse in R [8],v.4.1.0; [63] to calculate how many incubation recesses locations fell within a 90 m diameter circular buffer of a CLP. Specifically, the function getRecursionsAtLocation allowed us to evaluate how many times an incubation recess movement was to a CLP.
First we quantified the proportion of recess movements that went to a CLP. We calculated the proportion of recess movements made to CLP by using the number of recesses to any CLP and dividing by the total number of recesses during the incubation period. We then counted the total number of CLP used during the incubation period. Specifically, to calculate the number of CLP used during incubation, we counted the number of CLP that were visited at least once during incubation recess.
Female wild turkeys are constrained to the nest location during incubation [3, 45]. Thus, we calculated the distance from the nest location to the recess movement locations to see if this distance influenced habitat selection. Understanding patterns of resource selection relative to the presence of recursive movements offers a mechanism to link resource availability and female behavioral decisions [2]. So, we evaluated resource selection using a set of landcover covariates relevant to female wild turkey reproductive ecology [3, 13, 45]. We obtained year-specific, 30-m resolution spatial data on landcover from the Cropland Data Layer (Cropscape provided by the National Agricultural Statistics Service (National Agricultural Statistics Service 2015. We recoded and combined landcover in program R (v.4.1.0 [63], to create 6 unique landcover types (water, pine forest, hardwood forest, mixed pine-hardwood forest, open treeless areas, and shrub/scrub, [88, 89]). We calculated the nearest distance from each turkey use and available points to each landcover type, using the Euclidean distance tool in ArcMap 10.8 (Esri, Redlands, CA, USA). We used landcover distance metrics for subsequent analysis instead of a classification or categorical approach [19].
Nest fate model
We constructed a Bayesian logistic regression model to test our hypothesis regarding the relative importance of incubating females revisiting sites previously visited during laying on nest fate. Specifically, we included the covariates proportion of recess movements to CLP and number of CLP visited to predict nest fate. We chose the proportion of recess movements to CLP as a predictor because it reflects the proportion of movements back to prospected locations. Additionally, the choice of incorporating the number of CLP as a predictor was due to the potential benefits associated with having a greater variety of places to recess. We treated the probability of nest fate (success or failure) as a Bernoulli distribution. Our model included a unique identification number for each female turkey as a random effect to account for inter-individual variation. To improve model fit and allow for direct comparison of effect sizes of each predictor variable, we normalized all fixed effects included in the models using the scale function in R. We fitted models using package brms in program R [10]. We computed 4 MCMC chains for 8000 iterations, discarding the first 1000 iterations as a burn-in [28]. We calculated 95% credible intervals that provided a metric of uncertainty. We then computed the probability of direction which provided the probability each covariate either positively or negatively influenced nest fate. All estimated parameters had R-hat values < 1.1, indicating that all chains converged [26].
Resource selection model
We calculated 95% home ranges during the incubation period by fitting dynamic Brownian bridge movement models (dBBMMs) to the time-specific location data [15] using package move [42] in program R. We used an error estimate of 20 m, a moving window size of 7 locations, and a margin setting of 3 locations [11, 15]. These home ranges were estimated to evaluate resource selection within an individual’s incubation range.
We used resource selection functions (RSFs) to examine relationships between 6 landcover types and distances traveled from nests to wild turkey incubation recess movements to CLP within individual incubation ranges (3rd-order selection) following design III approach suggested by Manly et al. [46]. We compared use (incubation recess movements to CLP) points within individual incubation ranges to 500 available points sampled within each range [6]. We tested for collinearity between each of our covariates and excluded covariates using Pearson’s correlation with a r > 0.60 [24]. We found no correlation among covariates in our model. We used a generalized linear mixed model to include a random intercept for each individual turkey, with a binomial response distribution and logit link to the used-available data for turkeys [38, 46]. We used the lme4 R package [4] with a binary (0 = available, 1 = used) response variable to model resource selection. To improve fit, we rescaled all fixed effects by subtracting their mean and dividing by 2 standard deviations prior to modeling [27]. Due to the exploratory nature of our study, we chose to not use a model selection methodology but, instead, used a global model using the covariates selected for their known importance in wild turkey ecology.
Results
We monitored 692 nesting attempts by 485 (427 adults and 55 juveniles, 3 unknown) female wild turkeys during 2014‒2021. We removed 107 nesting attempts that were incubated < 3 days since we were unable to isolate incubation behaviors from nests of such short duration. We used 585 nesting attempts (initial attempts = 407, renesting attempts = 178) by 435 females to quantify whether females were revisiting CLP. We identified 31,145 recess movements during incubation, of which 56.9% (SD = 22.2, median = 58.7) were made to CLP (Fig. 2). Mean number of CLP used during laying that were visited during incubation recesses was 5 (SD = 1.9, range = 0–17 patches). The random effect of individual had a variance of 1.44 ± 0.63 within our nest fate model. The proportion of recess movements to CLP had no effect on nest fate (μ of posterior distribution with 95% credible − 0.0, − 0.01 to 0.01, probability of direction = 61.6%). However, as the number of CLP used during incubation recesses increased there was a positive impact on nest fate (μ of posterior distribution with 95% credible 0.19, 0.06–0.37, probability of direction = 99.8%), where the probability of nest success increased by 2.8% for every additional CLP visited (Fig. 3).

Proportion of incubation recess movements made to CLP for 585 nesting attempts made by 435 female eastern wild turkeys (Meleagris gallopavo silvestris) across the southeastern United States during 2014–2021. Violin plots are the distribution of the data with corresponding boxplot inside. The solid line identifies the median and the dot corresponds with the average proportion

Probability of nest success as a function of the number of CLP visited during incubation recesses for 585 nesting attempts made by 435 female eastern wild turkeys (Meleagris gallopavo silvestris) across the southeastern United States during 2014–2021. Gray shading represents the 95% credible intervals
For our RSF, we used 16,278 GPS locations from recess locations that were to 2831 CLP and created 585 individual home ranges. The random effect of individual had a variance of 1.84 (SE ± 1.36) in our RSF. Female wild turkeys selected for areas closer to hardwoods (β = − 0.27, SE ± 0.031), water (β = − 0.48, SE ± 0.037), mixed pine-hardwoods (β = − 0.10, SE ± 0.025), secondary roads (β = − 0.39, SE ± 0.051), shrub/scrub (β = − 0.16, SE ± 0.583), and areas closer to the nest (β = − 2.04, SE = 0.020; Fig. 4). Female wild turkeys avoided areas closer to open treeless areas (β = 0.27, SE ± 0.030) and pine (β = 0.06, SE ± 0.022; Fig. 4).

Coefficient plot depicting resource selection of eastern wild turkeys (Meleagris gallopavo silvestris) during incubation recesses to CLP across the southeastern United States during 2014–2021. The whiskers depict 95% confidence intervals around regression coefficient estimates
Discussion
Prospecting behavior before the onset of incubation has been found to occur in a variety of avian species [65]. Presumably, species rely on prospecting to determine areas capable of conferring greater nest success [23] and profitable patches ensuring availability of resources [56]. Using prospecting movements, our results indicate that ~ 57% of incubation recess movements were to patches visited during laying. Our findings support contemporary research demonstrating that wild turkeys increase daily movements during laying, indicative of a lack of site familiarity [71]. Similar behaviors have been described in waterfowl (Anas sp.) that visit future brood-rearing ponds prior to hatching [12], ruff (Philomachus pugnax) and black grouse (Tetrao tetrix) where females visit leks prior to the breeding season [5] and is presumed to occur in sage grouse (Centrocercus urophasianus) prior to incubation [25].
We observed that nest fate was not influenced by the number of times a female returned to CLP but was affected by how many different CLP she visited while on incubation recesses. Observational work by Williams and Austin [84] reported the unpredictability of timing and movement patterns by female wild turkeys during incubation. When individuals are faced with patchy resource distributions, they become constrained by the spatial distribution of resources [72]. Where resources are sparse, prey may have to endure periods of overlap with predators which makes prey more predictable, providing cues into their nesting behavior [69, 72]. Alternatively, when prey are surrounded by multiple safe sites where predators are less efficient, predators may avoid these locations [73]. Having multiple profitable foraging patches allows prey to be more unpredictable in their movements which favors the prey’s behaviors during nesting instead of the predator [75], thus, reducing cues to nest site locations [37]. Moreover, the familiarity of sites due to prospecting behavior could play a crucial role in reducing predation risks. Site familiarity may lead to a decrease in the duration and number of incubation recess bouts, a factor known to influence predation in many ground-nesting bird species [21, 44, 45, 76]. Our findings suggest that female wild turkeys not confined to repeatedly using the same patches within their incubation ranges had increased nest success. Collectively, these findings highlight the importance of site familiarity in shaping nesting behaviors and ultimately influencing nest success in the context of predator–prey dynamics. Overall, we found that female wild turkeys that were not confined to repeatedly using the same patches within their incubation ranges had increased nest success.
During nesting, avian species should surround themselves with adequate resources to survive incubation while reducing predation risk [22, 74]. Wild turkeys are habitat generalists [62], so we were not surprised that females used a variety of landcover types during incubation recess movements to sites previously visited during laying. Presumably, females were simply going to places that offered conditions capable of supporting survival. Our results indicated that pine and open-treeless areas were avoided by wild turkeys. Open-treeless areas and pine forest on our sites were typically open pastures dominated by forages planted for livestock, sod-forming grasses, or industrial pine forest. Similar types of open areas and pine forest fail to offer high quality foraging habitats for incubating females relative to other early successional vegetation communities [1, 47]. Likewise, during incubation females often try to avoid other females, hence reducing predation risk [32, 67, 71]. Therefore, remaining in forested areas could provide concealment to reduce intraspecific interactions and predation risk. Alternatively, environmental thermal regimes can shape avian behavior [35], and in warmer environments, Galliformes have been found to adjust habitat use to select for areas with cooler temperatures [34, 77]. Specific to wild turkeys, Nelson et al. [50] found that broods on our study sites avoided pine forests and selected cooler locations as the day progressed. Therefore, avoidance of open-treeless areas and pine forest may be due to thermal regulatory constraints.
Our results emphasize the complexity of how prior behavioral processes can affect future events, such as nesting behavior. While researchers often concentrate on characterizing nest sites, there is a tendency to neglect the critical aspect of movement decisions [22]. This neglects results in characteristics that frequently fail to describe the spatial scale at which nesting occurs [22, 78] and are not clearly linked to nest success [20, 40]. Our research highlights the biologically relevant spatial and temporal scale at which prospecting decisions influence nest fate. We also provide a different approach to understanding resource selection during avian reproduction. Specifically, we suggest that focus on identifying resource selection and activities during the laying period could be relevant and appropriate for other ground-nesting species. Furthermore, prospecting behavior has been thought to occur during recess bouts to identify brood-rearing habitat [25]. Future research should evaluate how prospecting for brood-rearing habitat may occur prior to or during the incubation period.
Conclusions
Central place foragers are faced with understanding their surroundings to maximize foraging ability while reducing predation risk. Prospecting behavior allows individuals to determine suitability of areas within their ranges that would increase fitness. We found that wild turkeys frequently (~ 57%) return to locations previously visited during the laying period. Furthermore, we found that this behavior was common among wild turkeys, but nest fate was influenced by numbers of prospected locations within the incubation range. Our findings suggest that having more patches could reduce cues to predators and positively influence nest fate. Alternatively, increased site familiarity through prospecting could reduce incubation recess bouts and duration which positively influence nest fate.
Availability of data and materials
The dataset supporting the conclusions of this article is archived in the Dryad repository.
Abbreviations
- CLP:
-
Clustered laying patch
- GPS:
-
Global positioning system
- RSFs:
-
Resources selection function
- VHF:
-
Very high frequency
References
Backs SE, Bledsoe LW. Invertebrate abundance and vegetative structure in forest openings. Natl Wild Turkey Sympos. 2011;10:51–63.
Bakner NW, Cohen BS, Collier BA, Chamberlain MJ. Recursive movements of eastern wild turkey broods in the southeastern United States. Wildl Soc Bull. 2022;46:e1274.
Bakner NW, Schofield LR, Cedotal C, Chamberlain MJ, Collier BA. Incubation recess behaviors influence nest survival of wild turkeys. Ecol Evol. 2019;9:14053–65.
Bates, D., M. Maechler, B. Bolker, and S. Walker. 2014. lme4: Linear mixed-effects models using eigen and S4. R package version 1.1–7. Available from: http://CRAN.R-project.org/package=lme4.
Beehler BM, Foster MS. Hotshots, hotspots, and female preference in the organization of lek mating systems. Am Nat. 1988;131:203–19.
Benson JF. Improving rigour and efficiency of use-availability habitat selection analyses with systematic estimation of availability. Methods Ecol Evol. 2013;4:244–51.
Berger-Tal O, Bar-David S. Recursive movement patterns: review and synthesis across species. Ecosphere. 2015;6:1–12.
Bracis C, Bildstein KL, Mueller T. Revisitation analysis uncovers spatio-temporal patterns in animal movement data. Ecography. 2018;41:1801–11.
Bracis C, Mueller T. Memory, not just perception, plays an important role in terrestrial mammalian migration. Proc R Soc B: Biol Sci. 2017;284:20170449.
Bürkner PC. brms: An R package for Bayesian multilevel models using stan. J Stat Softw. 2017;80:1–28.
Byrne ME, Guthrie JD, Hardin J, Collier BA, Chamberlain MJ. Evaluating wild turkey movement ecology: an example using first-passage time analysis. Wildl Soc Bull. 2014;38:407–13.
Casazza ML, McDuie F, Lorenz AA, Keiter D, Yee J, Overton CT, Peterson SH, Feldheim CL, Ackerman JT. Good prospects: high-resolution telemetry data suggests novel brood site selection behaviour in waterfowl. Anim Behav. 2020;164:163–72.
Chamberlain MJ, Cohen BS, Bakner NW, Collier BA. Behavior and movement of wild turkey broods. J Wildl Manag. 2020;84:1139–52.
Chernetsov N. Habitat selection by nocturnal passerine migrants en route: mechanisms and results. J Ornithol. 2006;147:185–91.
Cohen BS, Prebyl TJ, Collier BA, Chamberlain MJ. Home range estimator method and GPS sampling schedule affect habitat selection inferences for wild turkeys. Wildl Soc Bull. 2018;42:150–9.
Collier BA, Fyffe N, Smallwood A, Oleson B, Bakner NW, Heffelfinger JR, Chamberlain MJ. Reproductive ecology of Gould’s wild turkeys (Meleagris gallopavo mexicana) in Arizona. Wilson J Ornithol. 2019;131:667–79.
Conley MD, Yeldell NA, Chamberlain MJ, Collier BA. Do movement behaviors identify reproductive habitat sampling for wild turkeys? Ecol Evol. 2016;6:7103–12.
Conley M, Oetgen J, Barrow J, Chamberlain M, Skow K, Collier B. Habitat selection, incubation, and incubation recess ranges of nesting female Rio Grande wild turkeys in Texas. Natl Wild Turkey Sympos. 2015;11:117–26.
Conner LM, Smith MD, Burger LW. A comparison of distance-based and classification-based analyses of habitat use. Ecology. 2003;84:526–31.
Crawford JC, Porter WF, Chamberlain MJ, Collier BA. Wild turkey nest success in pine-dominated forests of the southeastern United States. J Wildl Manag. 2021;85:498–507.
Criscuolo F, Gauthier-Clerc M, Gabrielsen GW, Maho YL. Recess behaviour of the incubating common eider Somateria mollissima. Polar Biol. 2000;23:571–4.
Deeming DC, Reynolds SJ. Nests, eggs, and incubation: new ideas about avian reproduction. Oxford: Oxford University Press; 2015.
Doligez B, Danchin E, Clobert J. Public information and breeding habitat selection in a wild bird population. Science. 2002;297:1168–70.
Dormann CF, Elith J, Bacher S, Buchmann C, Carl G, Carré G, Marquéz JRG, Gruber B, Lafourcade B, Leitão PJ, Münkemüller T, McClean C, Osborne PE, Reineking B, Schröder B, Skidmore AK, Zurell D, Lautenbach S. Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography. 2013;36:27–46.
Dudko JE, Coates PS, Delehanty DJ. Movements of female sage grouse Centrocercus urophasianus during incubation recess. Ibis. 2019;161:222–9.
Gelman A. Parameterization and Bayesian modeling. J Am Stat Assoc. 2004;99:537–45.
Gelman A. Scaling regression inputs by dividing by two standard deviations. Stat Med. 2008;27:2865–73.
Gelman A, Rubin DB. Inference from iterative simulation using multiple sequences. Stat Sci. 1992;7:457–511.
Gupte PR, Beardsworth CE, Spiegel O, Lourie E, Toledo S, Nathan R, Bijleveld AI. A guide to pre-processing high-throughput animal tracking data. J Anim Ecol. 2022;91:287–307.
Guthrie JD, Byrne ME, Hardin JB, Kochanny CO, Skow KL, Snelgrove RT, Butler MJ, Peterson MJ, Chamberlain MJ, Collier BA. Evaluation of a global positioning system backpack transmitter for wild Turkey research. J Wildl Manag. 2011;75:539–47.
Heathcote RJP, Whiteside MA, Beardsworth CE, Van Horik JO, Laker PR, Toledo S, Orchan Y, Nathan R, Madden JR. Spatial memory predicts home range size and predation risk in pheasants. Nat Ecol and Evolution. 2023;7:461–71.
Healy WM. Behavior. In: Dickson JG, editor. The wild turkey: biology and management. Mechanicsburg: Stackpole Books; 1992. p. 46–65.
Hijmans RJ, Williams E, Vennes C, Hijmans MRJ. Package ‘geosphere.’ Spherical Trigonom. 2017;1:1–45.
Hovick T, Elmore R, Allred B, Fuhlendorf S, Dahlgren D. Landscapes as a moderator of thermal extremes: a case study from an imperiled grouse. Ecosphere. 2014;5:1–12.
Huey RB. Physiological consequences of habitat selection. Am Nat. 1991;137:S91–115.
Hughes M, Hyman J. Should I stay or should I go now: late establishment and low site fidelity as alternative territorial behaviors. Ethology. 2011;117:979–91.
Ibáñez-Álamo JD, Magrath RD, Oteyza JC, Chalfoun AD, Haff TM, Schmidt KA, Thomson RL, Martin TE. Nest predation research: recent findings and future perspectives. J Ornithol. 2015;156:247–62.
Johnson CJ, Nielsen SE, Merrill EH, McDonald TL, Boyce MS. Resource selection functions based on use-availability data: Theoretical motivation and evaluation methods. J Wildl Manag. 2006;70:347–57.
Kacelnik A (1984) Central place foraging in starlings (Sturnus vulgaris). I. Patch residence time. J Anim Ecol 283–299.
Keever AC, Collier BA, Chamberlain MJ, Cohen BS. Early nest initiation and vegetation density enhance nest survival in wild turkeys. Ornithology. 2023;140:ukac050.
Kokko H, López-Sepulcre A, Morrell LJ. From hawks and doves to self-consistent games of territorial behavior. Am Nat. 2006;167:901–12.
Kranstauber B, Kays R, LaPoint SD, Wikelski M, Safi K. A dynamic Brownian bridge movement model to estimate utilization distributions for heterogeneous animal movement. J Anim Ecol. 2012;81:738–46.
Lalla KM, Fraser KC, Frei B, Fischer JD, Siegrist J, Ray JD, Cohn-Haft M, Elliott KH. Central-place foraging poses variable constraints year-round in a neotropical migrant. Mov Ecol. 2022;10:1–12.
Lecomte N, Gauthier G, Giroux J. A link between water availability and nesting success mediated by predator-prey interactions in the Arctic. Ecology. 2009;90:465–75.
Lohr AK, Martin JA, Wann GT, Cohen BS, Collier BA, Chamberlain MJ. Behavioral strategies during incubation influence nest and female survival of wild turkeys. Ecol Evol. 2020;10:11752–65.
Manly BFJ, McDonald LL, Thomas DL, McDonald TL, Erickson WP. Resource selection by animals: statistical design and analysis for field studies. 2nd ed. Dordrecht: Kluwer Academic; 2002.
Martin JA, Palmer WE, Juhan SM Jr, Carroll JP. Wild turkey habitat use in frequently-burned pine savanna. For Ecol Manage. 2012;285:179–86.
Moscicki DJ, White JH, Hardin JB, Chamberlain MJ, Collier BA. Phenology-specific space use by Rio Grande wild turkeys. J Wildl Manag. 2023;87:e22331.
Moore FR, Aborn DA. Mechanisms of en route habitat selection: How do migrants make habitat decisions during stopover? Studies in Avian Biology. 2000;20:34–42.
Nelson SD, Keever AC, Wightman PH, Bakner NW, Argabright CM, Byrne ME, Collier BA, Chamberlain MJ, Cohen BS. Fine-scale resource selection and behavioral tradeoffs of eastern wild turkey broods. J Wildl Manag. 2022;86:e22222.
Ohashi K, Thomson JD. Efficient harvesting of renewing resources. Behav Ecol. 2005;16:592–605.
Olsson O, Brown JS, Helf KL. A guide to central place effects in foraging. Theor Popul Biol. 2008;74:22–33.
Orians, G. H. and N. E. Pearson. 1979. On the theory of central place foraging. Analysis of Ecological Systems 157–177.
Oro D, Bécares J, Bartumeus F, Arcos JM. High frequency of prospecting for informed dispersal and colonisation in a social species at large spatial scale. Oecologia. 2021;197:395–409.
Ottosson U, Bäckman J, Smith HG, Dickinson J. Nest-attenders in the pied flycatcher (Ficedula hypoleuca) during nestling rearing: a possible case of prospective resource exploration. Auk. 2001;118:1069–72.
Pärt T, Doligez B. Gathering public information for habitat selection: Prospecting birds cue on parental activity. Proc R Soc London Series B: Biol Sci. 2003;270:1809–13.
Pärt T, Arlt D, Doligez B, Low M, Qvarnström A. Prospectors combine social and environmental information to improve habitat selection and breeding success in the subsequent year. J Anim Ecol. 2011;80:1227–35.
Pelham PH, Dickson JG. Physical characteristics. In: Dickson JG, editor. The wild turkey: biology and management. Mechanicsburg: Stackpole Books; 1992. p. 32–45.
Piper WH. Making habitat selection more “familiar”: a review. Behav Ecol Sociobiol. 2011;65:1329–51.
Ponchon A, Gremillet D, Doligez B, Chambert T, Tveraa T, González-Solís J, Boulinier T. Tracking prospecting movements involved in breeding habitat selection: insights, pitfalls and perspectives. Methods Ecol Evol. 2013;4:143–50.
Ponchon A, Iliszko L, Grémillet D, Tveraa T, Boulinier T. Intense prospecting movements of failed breeders nesting in an unsuccessful breeding subcolony. Anim Behav. 2017;124:183–91.
Porter WF. Habitat requirements. In: Dickson JG, editor. The wild turkey: biology and management. Mechanicsburg: Stackpole Books; 1992. p. 202–13.
R Core Team. R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing, 2022. Retrieved from http://www.R-project.org.
Ranc N, Moorcroft PR, Ossi F, Cagnacci F. Experimental evidence of memory-based foraging decisions in a large wild mammal. Proc Natl Acad Sci. 2021;118:e2014856118.
Reed JM, Boulinier T, Danchin E, Oring LW. Informed dispersal: prospecting by birds for breeding sites. Curr Ornithol. 1999;15:189–259.
Riotte-Lambert L, Benhamou S, Chamaillé-Jammes S. Periodicity analysis of movement recursions. J Theor Biol. 2013;317:238–43.
Schaap JN, Silvy NJ, Peterson MJ, Aguirre R, Perotto-Baldivieso HL. Spatial distribution of female Rio Grande wild turkeys during the reproductive season. Natl Wild Turkey Sympos. 2005;9:231–5.
Schjørring S, Gregersen J, Bregnballe T. Prospecting enhances breeding success of first-time breeders in the great cormorant, Phalacrocorax carbo sinensis. Anim Behav. 1999;57:647–54.
Schmitz OJ, Miller JRB, Trainor AM, Abrahms B. Toward a community ecology of landscapes: predicting multiple predator–prey interactions across geographic space. Ecology. 2017;98:2281–92.
Schoener TW. Generality of the size-distance relation in models of optimal feeding. Am Nat. 1979;114:902–14.
Schofield LR. Evaluation of reproductive phenology and ecology of wild turkey (Meleagris gallopavo) across the southeastern United States. Thesis. Baton Rouge: Louisiana State University; 2019.
Sih A. Predator-prey space use as an emergent outcome of a behavioral response race. In: Barbosa P, Castellanos I, editors. Ecology of predator-prey interactions. London: Oxford University Press; 2005. p. 241–55.
Sih A. The behavioral response race between predator and prey. Am Nat. 1984;123:143–50.
Skutch AF. The constancy of incubation. Wilson Bull. 1962;74:115–52.
Smith JA, Donadio E, Pauli JN, Sheriff MJ, Bidder OR, Middleton AD. Habitat complexity mediates the predator–prey space race. Ecology. 2019;100:e02724.
Smith PA, Tulp I, Schekkerman H, Gilchrist HG, Forbes MR. Shorebird incubation behaviour and its influence on the risk of nest predation. Anim Behav. 2012;84:835–42.
Tanner EP, Elmore RD, Fuhlendorf SD, Davis CA, Dahlgren DK, Orange JP. Extreme climatic events constrain space use and survival of a ground-nesting bird. Glob Change Biol. 2017;23:1832–46.
Ulrey EE, Cedotal CA, Chamberlain MJ, Collier BA. Spatial distribution of potential wild turkey nest predators in west-central Louisiana. Wildl Soc Bull. 2022;46:e1285.
Van Beest F, Vander Wal E, Stronen A, Paquet PC, Brook R. Temporal variation in site fidelity: scale-dependent effects of forage abundance and predation risk in a non-migratory large herbivore. Oecologia. 2013;173:409–20.
Van Gils J, Tijsen W. Short-term foraging costs and long-term fueling rates in central-place foraging swans revealed by giving-up exploitation times. Am Nat. 2007;169:609–20.
Van Moorter B, Visscher D, Benhamou S, Börger L, Boyce MS, Gaillard J. Memory keeps you at home: a mechanistic model for home range emergence. Oikos. 2009;118:641–52.
Wakefield ED, Cleasby IR, Bearhop S, Bodey TW, Davies RD, Miller PI, Newton J, Votier SC, Hamer KC. Long-term individual foraging site fidelity—Why some gannets don’t change their spots. Ecology. 2015;96:3058–74.
Williams JB. Energetics of avian incubation. In: Carey C, editor. Avian energetics and nutritional ecology. Boston: Springer; 1996. p. 375–415.
Williams LE Jr, Austin DH. Studies of the wild turkey in Florida. Bulletin of the Florida Game and Fresh Water Fish Commission. Gainesville: University Presses of Florida; 1988.
Williams LE Jr, Austin DH, Peoples TE, Phillips RW. Laying data and nesting behavior of wild turkeys. Proc Southeast Assoc Game Fish Comm. 1971;25:90–106.
Wittmer HU, McLellan BN, Hovey FW. Factors influencing variation in site fidelity of woodland caribou (Rangifer tarandus caribou) in southeastern British Columbia. Can J Zool. 2006;84:537–45.
Ydenberg R, Welham C, Schmid-Hempel R, Schmid-Hempel P, Beauchamp G. Time and energy constraints and the relationships between currencies in foraging theory. Behav Ecol. 1994;5:28–34.
Yeldell NA, Cohen BS, Little AR, Collier BA, Chamberlain MJ. Nest site selection and nest survival of eastern wild turkeys in a pyric landscape. J Wildl Manag. 2017;81:1073–83.
Yeldell NA, Cohen BS, Prebyl TJ, Collier BA, Chamberlain MJ. Prescribed fire influences habitat selection of female eastern wild turkeys. J Wildl Manag. 2017;81:1287–97.
Zicus MC, Hennes SK. Nest prospecting by common goldeneyes. The Condor. 1989;91:807–12.
Zipkin EF, Grant EHC, Fagan WF. Evaluating the predictive abilities of community occupancy models using AUC while accounting for imperfect detection. Ecol Appl. 2012;22:1962–72.
Acknowledgements
We thank A. P. Gerrits, A. K. Lohr, C. M. Argabright, N. A. Yeldell, L. Schofield, C. J. Wakefield, P. H. Wightman, A. M. Byers, S. D. Nelson, S. Watkins, P. Goodman, and J. W. Wood for their efforts collecting field data.
Funding
This material is partially based on work supported by the National Institute of Food and Agriculture and United States Department of Agriculture under McIntire Stennis project (7001494). We appreciate funding and logistical support provided by the Georgia Department of Natural Resources—Wildlife Resources Division, the Louisiana Department of Wildlife and Fisheries, the School of Renewable Natural Resources and Louisiana State University Agricultural Center, the South Carolina Department of Natural Resources, and the Warnell School of Forestry and Natural Resources at the University of Georgia.
Author information
Authors and Affiliations
Contributions
NWB: Data curation; formal analysis; investigation; methodology; writing-original draft. EEU: Data curation; formal analysis; methodology; writing-review and editing. BAC: Conceptualization; funding, methodology; project administration, writing-review and editing. MJC: Conceptualization; funding acquisition; methodology; project administration; supervision; writing-review and editing.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1:
Study site descriptions.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Bakner, N.W., Ulrey, E.E., Collier, B.A. et al. Prospecting during egg laying informs incubation recess movements of eastern wild turkeys. Mov Ecol 12, 4 (2024). https://doi.org/10.1186/s40462-024-00451-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40462-024-00451-3