
Abstract
Background
There is growing evidence that individuals within populations can vary in both habitat use and movement behavior, but it is still not clear how these two relate to each other. The aim of this study was to test if and how individual bats in a Stunira lilium population differ in their movement activity and preferences for landscape features in a correlated manner.
Methods
We collected data on movements of 27 individuals using radio telemetry. We fitted a heterogeneous-space diffusion model to the movement data in order to evaluate signals of movement variation among individuals.
Results
S. lilium individuals generally preferred open habitat with Solanum fruits, regularly switched between forest and open areas, and showed high site fidelity. Movement variation among individuals could be summarized in four movement syndromes: (1) average individuals, (2) forest specialists, (3) explorers which prefer Piper, and (4) open area specialists which prefer Solanum and Cecropia.
Conclusions
Individual preferences for landscape features plus food resource and movement activity were correlated, resulting in different movement syndromes. Individual variation in preferences for landscape elements and food resources highlight the importance of incorporating explicitly the interaction between landscape structure and individual heterogeneity in descriptions of animal movement.
Similar content being viewed by others

Background
Movement is defined as the change in the spatial location of an organism over time, and it has a crucial role in determining the fates of individuals, and consequently the structure and dynamics of populations, communities and ecosystems [1]. Habitat fragmentation and climate change have pervasive impacts on ecosystem maintenance [2]. These anthropogenic processes can influence directly the movement behavior of individuals and species. For example, a survey of a GPS-tracking database of 803 individuals across 57 mammal species found that movement activity in areas with high anthropogenic impact was on average less than half of that in areas with low anthropogenic impact. Reduction in movement may be attributable to an individual-behavioral effect, where individuals alter their movements relative to the anthropogenic impact, or a species occurrence effect, where certain species that exhibit long-range movement simply do not occur in areas of high anthropogenic impact. [3]. To better predict how animals are likely to respond to environmental change, it is therefore important to characterize movement patterns and in particular to infer the underlying mechanisms driving those patterns.
A growing number of studies has shown that individuals differ substantially in resource use [4, 5] and movement patterns [6,7,8,9,10] in many taxa. Interindividual variation in resource use can be related to resource availability and seasonally, thus intraspecific variation level may not be fixed over time. For instance, the scarcity of resources can lead to increased intraspecific competition and increased interindividual variation [11], [12]. Although many studies have demonstrated that individuals vary in movement patterns and habitat use, it has remained less clear what mechanisms explain this variation.
At a proximate level, features that describe movement behavior (or movement syndromes) might potentially be related to morphology (e.g., wing morphology; [13, 14]), have a behavioral basis, or a combination of both. Assuming they have a behavioral basis [15, 16] it is unclear at this point if syndromes are learned (e.g., matrilineal cultural transmission; [17]) or have a genetic basis. Personality traits such as shyness-boldness, exploration-avoidance, activity, sociability and aggressiveness may be heritable, and may have important consequences for several ecological and evolutionary processes at the population (e.g. individual movement, gene flow) and community levels (e.g., individual variation in some sets of correlated personality and morphological traits may be viewed as functional sub-categories in the organization of communities; [18]).
In heterogeneous landscapes, where animals move around to seek shelter and resources needed for their reproduction and survival, landscape structure directly affects the realized movement patterns [19]. For example, in patchy landscapes, individuals typically move fast and in a directed manner when moving among the habitat patches, whereas their movements are slow and tortuous when moving within the patches [20]. In heterogeneous landscapes, individual preferences for food resources and habitat types can interact with landscape features, generating complex movement patterns [21]. Inferring the underlying mechanisms behind such interactions can be challenging, as it requires an appropriate combination of relevant data and analytical tools [22]: spatially explicit data on both animal movements and on the availability of the relevant food resources, as well as analytical tools that capture how variation in resource availability influences individual-specific movement behavior and data on the animal’s state at different times (disease, injuries, hunger-level).
The frugivore species Sturnira lilium (family Phyllostomidae) is one of the most abundant and widespread bats in the Atlantic Forest [23], exhibiting a strong preference for fruits of the genus Solanum, followed by Piper and to a smaller degree Cecropia, Ficus and Vismia [24, 25]. In a previous study on a population of S. lilium inhabiting a fragmented landscape of Brazilian savanna (Cerrado), we found that different individual bats foraged more often in different geographical locations, but whether individuals have different habitat preferences or movement behavior remain to be tested [26].
Natural forest environments such as the Atlantic Forest are increasingly impacted by agricultural, pasture and human expansion (70 % of the Brazilian population live along the Brazilian Atlantic coast; [27]), most of the remaining forest being distributed in small-sized and isolated fragments immersed in anthropogenic matrix [28]. There is evidence for a fragmentation threshold for bat richness, i.e. that the number of bat species strongly depends on the forest cover (%) at the landscape level. Furthermore, studies indicate that resource use correlates with response to fragmentation: frugivorous species are most tolerant to disturbance, being abundantly present also at forest edges, secondary forests and forestry (i.e. commercial Eucalyptus plantations) with dense understory with many chiropterochoric plants, such as Cecropia, Piper and Solanum [29, 30]. This suggests that frugivorous bats can to some extent adapt to environmental changes, at least over the short term. Although S. lilium can also utilize disturbed areas, it needs access to areas of forest, as it needs tall trees for roosting and understory vegetation for foraging [31, 32].
Here, we investigated how the movement patterns of S. lilium individuals inhabiting a heterogeneous landscape respond to landscape features and food resources. We hypothesized that there is correlation between statistics that describe individual habitat preferences and movement activity, so that the variation among individuals can be summarized as low-dimensional behavioral syndromes [33]. To test these hypotheses we acquired mark-recapture data based on radio telemetry, and analyzed the data using the heterogeneous-space diffusion model introduced by [34] and [35].
Materials and methods
Study area
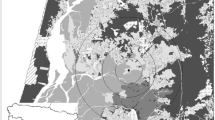
The Vassununga State Park (Parque Estadual de Vassununga - PEV) is a state nature reserve with area of c.a. 2070 ha, divided into six disjoint fragments, all of which are located in the city of Santa Rita do Passa Quatro, state of São Paulo (21º 42’ 37” S, 47º 28’ 41” W). The main vegetation types of the PEV is semi-deciduous forest (Atlantic Forest; 859 ha) and Cerrado (1,213 ha) –Brazilian savanna – surrounded by 2,776 ha of sugar cane and 1,960 ha of natural vegetation composed mainly by different physiognomy of Cerrado and riparian forest (Fig. 1).

Study site: Vassununga State Park and its surroundings, where movements of Sturnira lilium bats were recorded in São Paulo state, Brazil
Bat sampling
Sturnira lilium bats were monitored during the wet season from September to November 2016. Bats were caught using mist nets. For each captured bat, we measured its forearm with a caliper (0.01 mm precision) and its weight with a portable dynamometer (0.1 g precision) – this information contributed in species identification in the field. Captured bats were identified by a combination of taxonomic keys [36, 37]. Twenty-seven adult individuals of S. lilium were selected for radio telemetry. We classified the individuals as adults or juveniles based on the degree of ossification of the epiphyses of the phalanges of the wings [38], and selected only adult individuals to avoid ontogenetic effects [39]. We included both males (N = 23) and females (N = 4) to be able to ask if there is a systematic variation between sex. In order to capture possible variability of habitat preference and food preference within the population, we sampled individuals both inside the forest and in the open areas (Fig. 1).
Radio telemetry
We used the Axabixo equipment developed by Trapa-camera ®. The Axabixo device is a terrestrial radio telemetry system with digital encoding that operates both at the VHF frequency of 173,225 MHz and at the UHF frequency of 433,920 MHz (http://www.trapacamera.com.br/indexbixo.htm). The datalogger (i.e., the receiver) has an omnidirectional antenna, with range about 300 m in open areas, and it identifies the bat individual through a transmitter-specific digital code. We utilized thirty-six dataloggers (Fig. 1) that continuously recorded transmitter signals, storing information on date, time, transmitter identity and signal strength. Radio transmitters were attached to the back of the bats using veterinary glue (Vetbond). The weight of the transmitter was at most 5 % of the weight of the bat individual.
Landscape analysis
We mapped the landscape within a 4.5 km radius buffer with the centroid located in the PEV center. We made this choice based on our previous unpublished study on S. lilium, in which we recorded a maximum flight distance of 6 km. The mapping was made by manual digitalization and visual interpretation using high resolution satellite images (Open Layers Plugin Google Satellite at Quantum Gis 1.8) at the 1:5,000 scale. The following cover classes were mapped: semi-deciduous forest, open natural vegetation, sugar cane, agriculture, forestry, pasture, rural facilities and water. However, for the diffusion model, we reclassified the landscape into classes that we expected to be relevant for modelling bat movements: forest (18 % within the 4.5 km radius buffer; consisting of the PEV), open area (31 %; consisting of regeneration areas and natural open physiognomies such as gramineous-woody savanna, with a predominance of pioneer plants and shrubs) and matrix (51 %; mainly sugarcane, water and highways).
We computed a set of eight landscape attributes for each location of dataloggers: the proportions of forest cover (FC), open area cover (OA) and matrix cover (MA) in a circular buffer with 200 m radius, the spatial heterogeneity (HE) of the buffer calculated by the Shannon’s landscape diversity index; distance of the datalogger to the nearest forest fragment (NN), and estimated numbers of ripe fruits of trees in the genera Piper (PI), Solanum (SO) and Cecropia (CE). These trees provide the main resources consumed by S. lilium [24, 25]. We chose a 200 m radius buffer because it was the maximum signal range of the transmitters in forest areas. The landscape was mapped using Quantum Gis 1.8 (QGIS Development Team) and ArcGis 10.5. The landscape metrics were calculated on the extension V-Late.
Fruit availability
For estimating the number of fruits, we also used a 200 m radius buffer around each datalogger. Inside each buffer, we first estimated the number of fruits per branch as the average count over 10 individual trees. The trees were selected randomly from the point where the datalogger was located. Second, we defined four perpendicular transects and counted the number of branches of each plant species along each transect. Finally, we multiplied the number of branches by the average number of fruits per branch.
Fitting the heterogeneous-space diffusion model
We fitted to the bat detection data the heterogeneous-space diffusion model introduced by [34] and [40], and extended to account for variation among species by [35]. In this study, we consider a single species, but utilize the structure of the Joint Species Movement Modeling (JSMM) framework [35] to model variation among individuals. The diffusion model (which is technically a partial differential equation, see [34]) accounts for habitat-specific variation in movement and mortality rates, and for habitat selection at edges between habitat types. Prior to the analyses, we triangulated the landscape (as needed to fit the diffusion model to data; [34]) classified into the three habitat types: forests (\( h=1 \)), matrix (\( h=2 \)), and open areas (\( h=3 \)) – this step was done using the software Mapper [40]. The movement model contains seven parameters to be estimated for each individual \( i \): (a) three habitat-specific movement rates (the diffusion parameters \( {D}_i^h \) for each habitat type \( h \)); (b) two habitat preference parameters \( {k}_i^h \) (relative preferences to open areas and to matrix, normalized to 1 for forests); (c) mortality parameter \( {m}_i \) (assumed to be the same for all habitat types) and (d) detection probability \( {q}_i \) (assumed to be the same for all habitat types). We note that in the present study the mortality parameters are more likely to represent the death of the battery of the tracking equipment rather than the death of the individual bat.
The detection probability \( {q}_i \) measures the probability of observing the individual by a datalogger conditional on the individual being within the detection area [34]. We defined for each datalogger a detection area based on a field experiment where we examined from which locations the datalogger was able to detect the tracking equipment. We then doubled the sizes of these areas to account for small-scale bat movements that may bring the individual to the detection area during one-time step when the individual is nearby it. We note that the detection probabilities (\( {q}_i\Big) \) are likely to be similar for each individual, even if the devices may somewhat vary in their technical quality. We chose to include variation among individuals in detection probability partially because it allowed us to utilize the modelling framework of [40].
The \( {n}_p=7 \) individual-specific movement parameters can be described by the vector
We computed the likelihood \(p\left({\varvec{y}}_{i}|{\varvec{\Theta }}_{i\bullet }\right)\) for observing the movement data \({\varvec{y}}_{i}\) using the finite-element scheme to solve the time-evolution of the probability density for the individuals’ location, as described by [34] and implemented for the case of variation among species (here, \({n}_{i}\) individuals) by the JSMM [35]. The data are in continuous-time, but the implementation of the diffusion model requires one to define discrete time intervals at which the detections are attempted, which we set to 12 h intervals.
The JSMM estimates species- and community-level movement parameters as a function of species traits and their phylogenetic relationships. Here, we model the movement activity of bats dependent on individuals’ weight and sex. We combine the individual-specific parameters into the \({n}_{i}\times {n}_{p}\) matrix \(\varvec{\Theta }\), and then model the \({n}_{i}{n}_{p}\times 1\) vector \(\varvec{\uptheta }=\text{v}\text{e}\text{c}\left(\varvec{\Theta }\right)\) using a multivariate normal distribution
Here the \({n}_{p}\times {n}_{p}\) variance-covariance matrix \(\varvec{\Sigma }\) models the individual-specific deviations from the expectations based on the traits, \({{\mathbf{I}}_{n}}_{i}\) is the \({n}_{i}\times {n}_{i}\) identity matrix, and \(\otimes\) is the Kronecker (outer) product. The vector \(\mathbf{m}=\text{v}\text{e}\text{c}\left(\mathbf{M}\right)\) is the vectorized version of the \({n}_{i}\times {n}_{p}\) matrix \(\mathbf{M}\), with the expected movement parameters based on individuals’ sex, and weight. We model the matrix elements \({m}_{ip}\) for each individual \(i\) and parameter \(p\) as
where \( a \) is the index for \( {n}_a=3 \) traits (sex and weight, as well as intercept modelling the overall mean), and \( {t}_{ia} \) is the trait \( a \) for bat \( i \). The parameter \( {\zeta}_{ap} \) measures the effect of traits (sex and weight) on parameter \( p \). We log-transformed the weight values, and converted sex categories into a binary variable (1 for males and 0 for females).
We ran a Monte Carlo Markov Chain (MCMC) algorithm to estimate the parameters, first for 50,000 iterations, during which we adapted the proposal distributions of \(\varvec{\Theta }\) to achieve optimal mixing, and then sampled the posteriors by running the MCMC algorithm further by 350,000 iterations. We replicated the MCMC sampling for 3 independent chains, and assessed the convergence of the MCMC algorithm using the Gelman-Rubin convergence statistic [41] with the package CODA [42].
Deriving ecological inference from the diffusion model
To assess model fit, we simulated movement tracks, using individual specific parameter values sampled from the posterior distribution. To make real and simulated data as comparable as possible, we released each virtual bat in the location where a real bat was first observed, and we assumed the same spatial distribution of dataloggers as was used in the actual field study. We compared 1000 replicates of simulated data to observed data with sixteen statistics (S1-S16) that we considered relevant indicators of movement behavior. The statistics S1-S8 are the mean values of the eight landscape indices describe above (FC, OA, MA, HE, NN, PI, SO and CE), averaged over the locations where the individual was observed. The remaining statistics are the fraction of observations in forest (S9) and open habitats (S10), the proportion of consecutive observations in which the bat changed from one habitat type to another habitat type (S11), the mean distance between consecutive observations (S12), the distance between the first and the last observation (S13), the number of distinct receivers in which the bat was observed (S14), the proportion of observations in the receiver from which there were most observations (S15), and the proportion of time-steps from first to last observation in which the bat was detected (S16, Table 1). To validate the structural assumptions of the diffusion model, we averaged the statistics S1-S16 over the individuals and compared the statistics for real data to posterior mean and 95 % credible intervals [CI] derived from the simulations.
We assessed the presence of characteristic movement syndromes by examining if the variation among individuals showed correlated patterns in their movement characteristics. As movement characteristics, we used the posterior means of the individual-specific parameters of the movement model (diffusion rates and habitat preferences) and the residual statistics S1-S16, i.e. the difference between observation and the posterior mean prediction by the diffusion model. The reason for including the residual rather than the raw statistics was that they measure deviations of the movement characteristics S1-S16 from the null expectation based on the estimated movement parameters at the population scale. To make statistics with different units comparable, we normalized them to zero mean and unit variance over the individuals. We performed a Principal Component Analysis (PCA) to summarize the information contained in the continuous multivariate movement data considering the 23 residual statistics as the variables, and a k-means clustering to split the individuals into a set of 4 clusters. The optimal number of clusters (4) was determined by the Average Silhouette Method, which determines how well each object lies within its cluster [43]. We performed the PCA analysis using the prcomp function and the k-means analysis in R [44]. We used the factoextra R package to help in the interpretation and visualization of PCA and clustering analysis.
Results
We detected 7607 records (mean ± standard deviation = 271.7 ± 411.9, n = 27) and the mean number of days between first and last record per individual was 6 (+-6, n = 27). At the species level, habitat preference order of the bats was greater for open areas than for forest or matrix, bats preferred locations with high availability of Solanum fruits and bats changed between forest and open area habitats frequently. At the individual level, we detected four classes of individuals: (1) average individuals, (2) forest specialists, (3) explorers which prefer Piper, and (4) open area specialists which prefer Solanum and Cecropia (Fig. 2).

A comparison of population-level movement statistics between real and simulated data. The black lines are the statistics mean values for the simulated data, and the red dots are the statistic values for the real data. The shadowed areas show the distribution of the summary of 1000 simulations
Parameter estimates
The posterior means of the diffusion model parameters for the studied population are shown in Table 2 (see all the parameter estimates and the convergence diagnostics in Supporting Table S5). The model indicates that the habitat preference order of the bats was greater for open areas than for forest or matrix, and that movement rates were not related to habitat preferences so that the movement had similar diffusion rates in all habitats. The mean lifetime of bats (or batteries), which can be computed as the reciprocal of the mortality rate, was ca. 13 days. The estimated detection probability was very close to one, indicating that bats that were close to the vicinity of a datalogger were very likely to be detected by the equipment.
Structural model validation
The simulated data and the real data matched well in terms of the statistics S1-S6 and S8-S10 describing the distribution of the bats with respect to habitat types (Fig. 3). This is to be expected, as the diffusion model accounted explicitly for the habitat composition of the landscape. Deviations between simulated and observed data were minor also for statistics that were not directly fitted in the diffusion model. For example, the simulated data slightly underestimated the amount of Solanum fruits in the locations where the bats visited (S7), meaning that the bats preferred locations with high availability of fruits more than predicted by the diffusion model that does not explicitly account for fruit availability. As a second example, bats changed between habitat types more frequently than proposed by the simulated data (S11), indicating that they alternate between forest and open area habitats more often than predicted by the diffusion model that is based on a random walk assumption. As a third example, simulated data somewhat underestimated the proportion of times the bats were detected (S16), suggesting that the delineated detection area was too small for modelling the detection process of bats at this time scale.

Individual-specific parameters estimates. The dots show the posterior mean for each male (blue) and female bat (red). The lines show the expected posterior means based on bat weight and sex – females (red) and males (blue). The posterior probability of weight having a positive effect on the movement parameter is denoted by pw. The posterior probability of males having a larger movement parameter than females is denoted by ps. The proportion of variation of each movement parameter explained by weight and sex are denoted by r²w and r²s, respectively. We show for each movement parameter the amount of variation not explained by bat weight and sex (r²i)
Individual variation in movement behavior
The sex and the weight of the bats explained less than 10 % of variation in posterior movement parameters, except for the mortality rate (Fig. 4). Despite the overall little amount of variation explained by bats’ traits, we found evidence that larger bats have larger diffusion rates in open areas, and that male bats have a higher mortality rate (probably related to battery life). For all other movement parameters, weight and sex explained only little of the variation among individuals (Fig. 4).

Results of a k-mean analysis aimed at detecting behavioral syndromes of Sturnira lilium. Numbers represent individuals. Each color represents the syndrome that each individual belongs, determined by k-means analysis. The largest axis of variation PC1 (25 % of statistics variance) separates mainly habitat use, increasing PC1 meaning increasing use of forests, and PC2 (17.3 %) separates mainly long-term movement activity, increasing PC2 meaning exploratory (as compared to stationary) behavior
We identified four main movement syndromes according to a cluster analysis using the correlation among the statistics and a PCA analysis (Fig. 2; Table 3, Additional file S1 and S4. However, it is important to note that the variation between individuals can be gradual rather than very well-defined clusters. We used a k-mean analysis to divided bats into four classes: (1) average individuals, (2) forest specialists, (3) explorers which prefer Piper, and (4) open area specialists which prefer Solanum and Cecropia (Table 3; Fig. 2). Figures with the movement of each individual can be seen in Additional file S2, a synthesis of the data obtained from each individual is provided in Additional file S3.
Discussion
Our results indicate that individuals in a population of a common frugivorous bat species vary in how their movement depends on landscape elements. Importantly, we show that variation in different aspects of movement behavior can be summarized in terms of behavioral movement syndromes. Kerches-Rogeri et al. (2020) demonstrated that different individual bats foraged more often in different geographical locations regardless of habitat type. In our present study, we go one step further and demonstrate that individuals have different habitat preferences and movement characteristics.
The fact that we found a weak effect of sex on movement can be related to the small sample size of females. However, we consider that this deserves future studies because the shape of the wings may be different between the sexes because females carry extra weight during pregnancy and wing shape must keep aerodynamics of flight [45], but we do not know whether movement and habitat selection are related to differences in wing morphology.
Regarding weight, larger bats tended to have high movement rate in open areas. A similar trend of body size effect in space use can be found in interspecific studies at other taxa [3, 46, 47], but there are only indications of this trend among bats (e.g. [48, 49]). Our results thus suggest that also intraspecific variation in body size is relevant for understanding how bats move within a landscape.
Some studies indicate that S. lilium is preferentially associated with secondary forest (e.g. [30], [50]), whereas other studies indicate an association with more mature forests [51]. Our results may help to elucidate these descriptions, by showing that a single population may contain individuals with preferences for different habitat types. Because the degree of inter-individual variation may vary among populations [4], the presence of syndromes may be population-specific rather than universal, since each population faces different environments and conditions, e.g. resource availability and predation pressure [52]. It is possible that some populations are behaviorally monomorphic (either specializing in open or forest habitats) and others (e.g., the population studied here) are polymorphic, which would help to explain these apparent conflicting findings.
Intraspecific variation can affect important processes to the survival of the species. The niche overlap among individuals can increase or decrease competition, since variation between individuals of the same species can decrease competition between conspecifics, but increase competition with other species [11]. In another scenario, the variation between individuals can also be a relief for inter-specific competition, since even if the overlap between the niche of other species increases, this impact can be reduced, because only a subgroup of individuals in each species are affected.
Intraspecific variation can also be important for ecosystem processes performed by the species. Because S. lilium is an important seed disperser and a “fragment connector” for many plant species in the Neotropics [53, 54], the presence of movement syndromes may have implications for this ecological service. The importance of variation within a population of dispersers may be related to Jensen`s inequality mechanism, in which when an ecological interaction depends nonlinearly on a specific characteristic, the variation around the mean of that characteristic can change the average strength the interaction [11, 55]. If a population has different subsets of individuals with respect to the partition of landscape use, it may mean that the real connectivity of the landscape by the bats may be different from the connectivity provided by a population formed by average individuals.
Individuals with different syndromes are likely to interact with different plant species and disperse seeds to different areas with different efficiencies, suggesting that they might vary in their quality as dispersers [55, 56]. Individuals using forest areas, for instance, are likely to be effective dispersers of Piper seeds, whereas individuals using open areas are likely to be effective dispersers of Solanum seeds. Understanding how inter-individual variation in movement patterns in S. lilium affects seed dispersal and the dynamics of the plants they disperse should be addressed in future research.
From a conservation biology point of view, the presence of movement syndromes suggests that individuals are likely to respond differently to habitat change, which may be good news, since the adaptation capacity of these bats to different natural and anthropogenic habitat may guarantee their maintenance within human-modified landscapes. Individuals specialized in open habitats might benefit from forest fragmentation, whereas those specialized in forest habitats should be negatively impacted. By the same token, restoration programs aiming at recovering only dense forest habitats may be advantageous for only a subset of the population. Besides, if individual specialization is correlated with the number of coexisting species (as predicted by the niche variation hypothesis), then protecting a highly variable population of a species may require protecting a habitat of little diversity, while reserves designed to include high interspecific biodiversity can minimize intraspecific diversity (52). Therefore, conservation efforts may benefit from considering how changes in habitat may impact individuals differently in a population.
It is important to note that our study describes movement for a short period of time (with an average battery life of 13 days), so we do not know whether bats consistently fall into a single behavioral syndrome or they switch back and forth, just as we do not know whether this is a consistent individual-level trait or is it subject to unmeasured temporal factors. One of the causes of individual specialization is intraspecific competition, which is related to the availability of resources, therefore, the same population may exhibit strong or weak niche overlap between individuals according to the availability of resources. Thus, it would be plausible to assume that the organization of bat individuals in groups is subject to seasonal variation related to the natural seasonal availability of resources.
Conclusions
Our results indicate that individuals may vary consistently in their movement patterns, which can be considered movement syndromes. Individual preferences for elements of the landscape highlight the importance of incorporating explicitly the landscape to descriptions of animal movement. An important step in future research is to understand more mechanistically the eco-evolutionary processes driving such variation, and to account for movement syndromes in conservation efforts, e.g. related to the consequences of habitat fragmentation on seed dispersal.
Availability of data and material
Data will be available on MoveBank (https://www.movebank.org).
Abbreviations
- GPS:
-
Global positioning system
- PEV:
-
Parque Estadual de Vassununga
- FC:
-
forest cover
- OA:
-
open area cover
- MA:
-
matrix cover
- HE:
-
spatial heterogeneity
- NN:
-
distance of the datalogger to the nearest forest fragment
- PI:
-
estimated numbers of ripe fruits of trees in the genus Piper
- SO:
-
estimated numbers of ripe fruits of trees in the genus Solanum
- CE:
-
estimated numbers of ripe fruits of trees in the genus Cecropia
- MCMC:
-
Monte Carlo Markov Chain
- CI:
-
credibility interval
References
Nathan R, Getz WM, Revilla E, Holyoak M, Kadmon R, Saltz D, et al. A movement ecology paradigm for unifying organismal movement research. Proc Natl Acad Sci. 2008;105:19052–9.
Turner IM. Species loss in fragments of tropical rain forest: a review of the evidence. J Appl Ecol. 1996;33:200–9.
Tucker MA, Böhning-Gaese K, Fagan WF, Fryxell JM, Van Moorter B, Alberts SC, et al. Moving in the Anthropocene: Global reductions in terrestrial mammalian movements. Science. 2018;359:466–9.
Bolnick DI, Svanbäck R, Fordyce J, Yang LH, Davis JM, Hulsey CD, et al. The ecology of individuals: incidence and implications of individual specialization. Am Nat. 2003;161:1–28.
Araújo MS, Bolnick DI, Layman CA. The ecological causes of individual specialisation. Ecol Lett. 2011;14:948–58.
Austin D, Bowen WD, McMillan JI. Intraspecific variation in movement patterns: modeling individual behaviour in a large marine predator. Oikos. 2004;105:15–30.
Ciuti S, Muhly TB, Paton DG, McDevitt AD, Musiani M, Boyce MS. Human selection of elk behavioural traits in a landscape of fear. Proc R Soc B Biol Sci. 2012;279:4407–16.
Farwell M, Fuzzen MLM, Bernier NJ, McLaughlin RL. Individual differences in foraging behavior and cortisol levels in recently emerged brook charr (Salvelinus fontinalis). Behav Ecol Sociobiol. 2014;68:781–90.
Bonnot N, Verheyden H, Blanchard P, Cote J, Debeffe L, Cargnelutti B, et al. Interindividual variability in habitat use: Evidence for a risk management syndrome in roe deer? Behav Ecol. 2015;26:105–14.
Spiegel O, Leu ST, Sih A, Godfrey SS, Bull CM. When the going gets tough: behavioural type-dependent space use in the sleepy lizard changes as the season dries. Proc R Soc B Biol Sci. 2015;282:20151768.
Bolnick DI, Amarasekare P, Araújo MS, Bürger R, Levine JM, Novak M, Rudolf VHW, Schreiber SJ, Urban MC, Vasseur DA. Why intraspecific trait variation matters in community ecology. 2011. 26(4): 183–192.
Herrera LG, Korine C, Fleming TH, Arad Z. Dietary Implications of Intrapopulation Variation in Nitrogen Isotope Composition of an Old World Fruit Bat. J Mammal. 2008;89(5):1184–90.
Kalcounis MC, Brigham RM. lntraspecific variation in wing loading affects habitat use by little brown bats (Myotis lucifugus). Can J Zool. 1995;73:89–95.
Adams RA. Morphogenesis in bat wings: Linking development, evolution and ecology. Cells Tissues Organs. 2007;187:13–23.
Fortin D, Beyer HL, Boyce MS, Smith DW, Duchesne T, Mao JS. Wolves influence elk movements: Behavior shapes a trophic cascade in Yellowstone National Park. Ecology. 2005;86:1320–30.
Patterson TA, Thomas L, Wilcox C, Ovaskainen O, Matthiopoulos J. State-space models of individual animal movement. Trends Ecol Evol. 2008;23:87–94.
Estes JA, Riedman ML, Staedler MM, Tinker MT, Lyon BE. Individual Variation in Prey Selection by Sea Otters: Patterns, Causes and Implications. J Anim Ecol. 2003;72:144–55.
Réale D, Reader SM, Sol D, McDougall PT, Dingemanse NJ. Integrating animal temperament within ecology and evolution. Biol Rev. 2007;82:291–318.
Van Moorter B, Rolandsen CM, Basille M, Gaillard JM. Movement is the glue connecting home ranges and habitat selection. J Anim Ecol. 2016;85:21–31.
Morales JM, Haydon DT, Frair J, Holsinger KE, Fryxell JM. Extracting More Out of Relocation Data: Building Movement Models As Mixtures of Random Walks. Ecology. 2004;85:2436–45.
Leclerc M, Vander Wal E, Zedrosser A, Swenson JE, Kindberg J, Pelletier F. Quantifying consistent individual differences in habitat selection. Oecologia. 180. Berlin Heidelberg: Springer; 2016. pp. 697–705.
Spiegel O, Leu ST, Bull CM, Sih A. What’s your move? Movement as a link between personality and spatial dynamics in animal populations. Ecol Lett. 2017;20:3–18.
Muylaert R, de L, Stevens, Esbérard RD, Mello CEL, Garbino MAR, Varzinczak GST. LH, et al. ATLANTIC BATS: a data set of bat communities from the Atlantic Forests of South America. Ecology. 2017;98:3227–7.
Mello MAR, Kalko EKV, Silva WR. Diet and Abundance of the Bat Sturnira lilium (Chiroptera) in a Brazilian Montane Atlantic Forest. J Mammal. 2008;89:485–92.
Andrade TY, Thies W, Rogeri PK, Kalko EKV, Mello MAR. Hierarchical fruit selection by Neotropical leaf-nosed bats (Chiroptera: Phyllostomidae). J Mammal. 2013;94:1094–101.
Kerches-Rogeri P, Niebuhr BB, Muylaert RL, Mello MAR. Individual specialization in the use of space by frugivorous bats. J Anim Ecol. 2020. Accepted Author Manuscript. doi:https://doi.org/10.1111/1365-2656.13339.
Tabarelli M, Aguiar AV, Ribeiro MC, Metzgeer JP, Peres CA. Prospects for biodiversity conservation in the Atlantic Forest: Lessons from aging human-modified landscapes. Biol Cons. 2010;143(10):2328–40. https://doi.org/10.1016/j.biocon.2010.02.005.
Ribeiro MC, Metzger JP, Martensen AC, Ponzoni FJ, Hirota MM. The Brazilian Atlantic Forest: How much is left, and how is the remaining forest distributed? Implications for conservation. Biol Conserv. 142: Elsevier Ltd; 2009. pp. 1141–53.
Muylaert RL, Stevens RD, Ribeiro MC. Threshold effect of habitat loss on bat richness in cerrado- forest landscapes. Ecol Appl. 2016;26:1854–67.
Castro-Luna AA, Sosa VJ, Castillo-Campos G. Bat diversity and abundance associated with the degree of secondary succession in a tropical forest mosaic in south-eastern Mexico. Anim Conserv. 2007;10:219–28.
Evelyn MJ, Stiles DA. Roosting Requirements of Two Frugivorous Bats (Sturnira lilium and Arbiteus intermedius) in Fragmented Neotropical Forest A. Biotropica. 2003;35:405–18.
Mello MAR, Kalko EKV, Silva WR. Movements of the bat Sturnira lilium and its role as a seed disperser of Solanaceae in the Brazilian Atlantic forest. J Trop Ecol. 2008;24:225–8.
Sih A, Bell AM, Johnson JC, Ziemba RE. Behavioral Syndromes: An Integrative Overview. The Quarterly Review of Biology. 2004. 79(3) http://www.jstor.org/stable/10.1086/422893.
Ovaskainen O. Habitat-specific movement parameters estimated using mark-recapture data and a diffusion model. Ecology. 2004;85:242–57.
Ovaskainen O, Ramos DL, Slade EM, Merckx T, Tikhonov G, Pennanen J, et al. Joint species movement modeling: how do traits influence movements? Ecology. 100: John Wiley & Sons, Ltd; 2019. p. e02622.
Vizotto LD, Taddei VA. Chave para determinação de quiropteros brasileiros. São José do Rio Claro: Universidade Estadual Paulista; 1973.
Gardner AL. Mammals of South America: Volume I. Marsupials, xenarthras, shrews, and bats. Chicago.: University of Chicago Press; 2008.
Kunz TH, Fenton M. Bat ecology. Chicago.: University of Chicago Press; 2003.
Powers LV, Kandarian SC, Kunz TH. Ontogeny of flight in the little brown bat, Myotis lucifugus: behavior, morphology, and muscle histochemistry. J Comp Physiol A. 1991;168:675–85.
Ovaskainen O, Rekola H, Meyke E, Arjas E. Bayesian Methods for Analyzing Movements in Heterogeneous Landscapes from Mark- Recapture Data. Ecology. 2008;89:542–54.
Gelman A, Rubin DB. Inference from Iterative Simulation Using Multiple Sequences. Stat Sci. 1992;7:457–72.
Plummer M, Best N, Cowles K, Vines K. CODA: convergence diagnosis and output analysis for MCMC. R News Technische Univ. 2006;6:7–11.
Kaufman L, Rousseeuw PJ. Finding Groups in Data: An Introduction to Cluster Analysis. New York: Wiley; 1990.
Core R Team 2020.
de Camargo NiF, de Oliveira HFM. Sexual Dimorphism in Sturnira lilium (Chiroptera, Phyllostomidae): Can Pregnancy and Pup Carrying Be Responsible for Differences in Wing Shape? PLoS One. 2012;7.
Bowman J, Jaeger JAG, Fahrig L. Dispersal distance of mammals is proportional To home range size. Ecology. 2002;83:2049–55.
Harestad AS, Bunnel FL. Home Range and Body Weight–A Reevaluation. Ecology. 1979;60:389–402.
Kalko EKV, Herre EA, Handley COJ. Relation of Fig Fruit Characteristics to Fruit-Eating Bats in the New and Old World Tropics. J Biogeogr. 1996;23:565–76.
Albrecht L, Meyer CFJ, Kalko EKV. Differential mobility in two small phyllostomid bats, Artibeus watsoni and Micronycteris microtis, in a fragmented Neotropical landscape. Acta Theriol (Warsz). 2007;52:141–9.
Pinto N, Keitt TH. Scale-dependent responses to forest cover displayed by frugivore bats. Oikos. 2008;117:1725–31. https://doi.org/10.1111/j.1600-0706.2008.16495.x.
Gorresen PM, Willig MR. Landscape Responses of Bats To Habitat Fragmentation in Atlantic Forest of Paraguay. J Mammal. 2004;85:688–97.
Nilsson J-A, Bronmark C, Hansson L-A, Chapman BB. Individuality in movement: the role of animal personality. In: Hansson L-A, Akesson S, editors. Anim Mov Across Scales. Oxford University Press; 2014. pp. 45–66.
Bello C, Galetti M, Montan D, Pizo MA, Mariguela TC, Culot L, et al. Atlantic frugivory: a plant-frugivore interaction data set for the Atlantic Forest. Ecology. 2017;98:1729–9.
Loayza AP, Loiselle BA. Preliminary Information on the Home Range and Movement Patterns of Sturnira lilium (Phyllostomidae) in a Naturally Fragmented Landscape in Bolivia. Biotropica. 2008;40:630–5. https://doi.org/10.1111/j.1744-7429.2008.00422.x.
Zwolak R. How intraspecific variation in seed-dispersing animals matters for plants. Biol Rev. 2018;93:897–913.
Pollux BJA. Consistent individual differences in seed disperser quality in a seed – eating fish. Oecologia. 183. Berlin Heidelberg: Springer; 2017. pp. 81–91.
Acknowledgements
We thank the logistic support provided by the State Park Vassununga and surroundings. We thank Bernardo Brandão S. Niebuhr for the fruitful discussions and review of the early versions of the manuscript. We thank the reviewers and editor for careful revision. We acknowledge the computational resources provided by the Aalto Science-IT project.
Funding
The research was funded by the Academy of Finland (grants 1273253, 250444 and 284601 to OO) and Research Council of Norway (CoE grant 223257). PKR was funded by grant #2014/24219-4 and #2016/14939-5, São Paulo Research Foundation (FAPESP). PKR and MCR was funded by grant #2013/50421-2, São Paulo Research Foundation (FAPESP). MCR also thanks to CNPq (processes # 312045/2013-1; #312292/2016-3) and PROCAD/CAPES (project # 88881.068425/2014-01) for their financial support.
Author information
Authors and Affiliations
Contributions
PKR, MCR and MSA conceived the ideas and designed methodology; PKR, BOT, RSCA and CFPB collected the data; PKR, JS, DLR and OO analyzed the data; PKR, MSA, DLR and OO led the writing of the manuscript. All authors contributed critically to the drafts and gave final approval for publication.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Approval for capturing the bats was obtained from ICMBio (capture authorization SISBIO 43730-1). Approval for handling the bats was obtained from the Ethics Committee on Animal Use of the Institute of Biosciences - UNESP - Rio Claro Campus / CEUAIB-UNESP-CRC (No 18/2015).
Consent for publication
Not applicable.
Competing interest
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Additional file 1
S1. The rationale behind the interpretation of the syndromes.
Additional file 2
S2. Individuals-movement.
Additional file 3
S3. Datalogger records summary.
Additional file 4
S4. Clusters definition and PCA output.
Additional file 5
S5. Parameter estimates and convergence diagnostics.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Kerches-Rogeri, P., Ramos, D.L., Siren, J. et al. Movement syndromes of a Neotropical frugivorous bat inhabiting heterogeneous landscapes in Brazil. Mov Ecol 9, 35 (2021). https://doi.org/10.1186/s40462-021-00266-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40462-021-00266-6