
Abstract
The blood plasma of numerous snake species naturally comprises endogenous phospholipase A2 inhibitors, which primarily neutralize toxic phospholipases A2 that may eventually reach their circulation. This inhibitor type is generally known as snake blood phospholipase A2 inhibitors (sbPLIs). Most, if not all sbPLIs are oligomeric glycosylated proteins, although the carbohydrate moiety may not be essential for PLA2 inhibition in every case. The presently known sbPLIs belong to one of three structural classes – namely sbαPLI, sbβPLI or sbγPLI – depending on the presence of characteristic C-type lectin-like domains, leucine-rich repeats or three-finger motifs, respectively. Currently, the most numerous inhibitors described in the literature are sbαPLIs and sbγPLIs, whereas sbβPLIs are rare. When the target PLA2 is a Lys49 homolog or an Asp49 myotoxin, the sbPLI is denominated a myotoxin inhibitor protein (MIP). In this brief overview, the most relevant data on sbPLIs will be presented. Representative examples of sbαPLIs and sbγPLIs from two Old World – Gloydius brevicaudus and Malayopython reticulatus – and two New World – Bothrops alternatus and Crotalus durissus terrificus – snake species will be emphasized.
Similar content being viewed by others


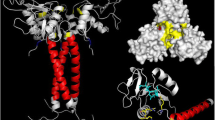
Background
A number of venomous and nonvenomous snake species are naturally resistant to the deleterious actions of snake venom components, in many cases due to the presence of specific antitoxins in their circulating blood [1–10]. These antitoxins were identified as liver-secreted proteins, which prevent any possible damage from toxins that might have reached the snake’s blood stream [11]. Among these inhibitors, phospholipase A2 inhibitors or snake blood phospholipase A2 inhibitors (sbPLIs) play a key role in this type of endogenous resistance.
During the 80’s and 90’s, a number of sbPLIs were purified from different snake species. The first authors to identify various sbPLIs in a single snake species – Gloydius brevicaudus, formerly Agkistrodon blomhoffii siniticus – proposed a classification based on the presence of characteristic domains of known mammalian proteins in their structure and on variations in their PLA2 selectivity [12]. Alpha sbPLIs (sbαPLIs) have a C-type lectin-like domain that is highly similar to the carbohydrate recognition domain of Ca2+-dependent lectins, and preferentially inhibit acidic PLA2s. Beta-type inhibitors (sbβPLIs) exhibit tandem leucine-rich repeats (LRRs), and specifically inhibit basic PLA2s. Gamma inhibitors (sbγPLIs) display a three-finger pattern and are less specific than the aforementioned classes, therefore inhibiting neutral, acidic and basic PLA2s from snake venoms. The structural classification of sbPLIs has been adopted by most authors working on the subject, but the selectivity concept is not absolute [13–16]. In general, α and γ sbPLIs simultaneously occur in several snake species, while sbβPLIs have only been reported in three snake species.
Native sbPLIs are usually homo- or heterooligomers of glycosylated and/or non-glycosylated subunits. Carbohydrates do not seem essential for the inhibition of PLA2 by sbPLIs, since some of them remain functional in the absence of this moiety [16–20]. When the target PLA2s are Lys49 homologues or Asp49 myotoxins, the sbPLIs are specifically called myotoxin inhibitor proteins (MIPs) [13, 14, 16, 21, 22].
The following sections present the most relevant characteristics of the three classes of sbPLIs. Subsequently, examples of sbαPLIs and sbγPLIs from two Old World snake species — Gloydius brevicaudus and Malayopython reticulatus — and two New World ones — Bothrops alternatus and Crotalus durissus terrificus — will be introduced.
Alpha class of sbPLIs (sbαPLIs)
Members of this class of inhibitors are found in solution as homo- or heterooligomers, with molecular masses between 75 kDa and 120 kDa (Table 1).
In addition to the typical C-type lectin-like domain, sbαPLI monomers present two other highly conserved regions in their structure: a hydrophobic core at their carboxy-terminus and an α-helical coiled-coil neck comprising the 13th to 36th amino acid segment in the mature protein [23, 24]. The last amino acid stretch corresponds to the exon 3 reported for the gene of the sbPLI from Protobothrops flavovoridis (formerly Trimeresurus flavoviridis) [25].
Besides the functional sbαPLIs, non-functional homologs were purified from the blood serum of two nonvenomous species, Elaphe quadrivirgata and E. climacophora. Despite displaying not only molecular masses, but also primary and quaternary structures comparable to classical sbαPLIs, these homologs failed to inhibit all tested snake venom PLA2s [26, 27].
The sbαPLI from Asian Gloydius brevicaudus (GbαPLI)
The sbαPLI from G. brevicaudus (formerly Agkistrodon blomhoffii siniticus) is a homotrimer, in which the α-helical coiled-coil neck subunit forms a central pore that constitutes the binding site for the target PLA2s [28–30]. The C-type lectin-like domain was discarded as responsible for PLA2 binding [30].
The correct configuration of the central pore in GbαPLI is controlled by the primary structures of the α-helical coiled-coil neck in the formation of subunits. Chimeric constructions of GbαPLI and the non-functional sbαPLI homolog from E. quadrivirgata allowed the mapping of important amino acids for PLA2 inhibition in the 13–36 segment, which are expected to be located in the helical neck of the GbαPLI trimer based on the three-dimensional structural model constructed by homology modeling [29, 30]. The trimerization occurs only among subunits having the same α-helical motif in the regions 13–36 and the oligomer is structurally stabilized by intermolecular electrostatic interactions. Two charged residues, E23 and K28, have been found specifically responsible for these essential interactions between the forming subunits in the trimer. The contribution of each subunit to the total inhibitory activity of trimeric GbαPLI has also been investigated. In the trimer, the inhibitory action is driven by one subunit with the highest affinity and is not affected by the number of subunits of this type [29].
GbαPLI displays lower affinities (about 2000-fold less) for neutral or basic PLA2s from the homologous venom compared to acidic PLA2s. In the absence of carbohydrates, the inhibition of acidic and neutral PLA2s has been reported to remain unchanged, while the inhibition of basic PLA2s is affected [19]. The possibility of different inhibition mechanisms, depending on the ionic character of the target PLA2, has been attributed to GbαPLI and other sbαPLIs, but further studies are required to clarify this issue.
The sbαPLI from Latin American Bothrops alternatus (BaltMIP)
This inhibitor was purified from the blood serum of Bothrops alternatus snakes by affinity chromatography using bothropstoxin I – a basic Lys49 PLA2 from the homologous venom – as the immobilized ligand. The monomer of BaltMIP is composed of a single polypeptide chain with apparent molecular mass of 24 kDa. The native molecule is able to inhibit myotoxicity and cytotoxicity caused by both Lys49 and Asp49 PLA2s, possibly by different mechanisms depending on the type of enzyme to be inhibited [15]. Amino acid residues possibly involved in the inhibition by BaltMIP of acidic PLA2s from homologous venom have been recently discussed in comparison to published data for PLA2-sbαPLIs complexes from Asian snake species [23].
The characteristic α-helical coiled-coil neck, the carbohydrate recognition domain and the hydrophobic core of sbαPLIs are well conserved in the BaltMIP monomer, according to the theoretical structural model (available in the Model Archive database under DOI: 105452/ma-a2iil). In the trimeric BaltMIP (available in the Model Archive database under DOI: 105452/ma-a4btt), three monomers fit well in a spherical arrangement [15].
Recombinant BaltMIP, displaying the same apparent molecular mass (24 kDa) as the native inhibitor monomer, has been produced in Pichia pastoris. The expressed protein was heavily glycosylated and formed oligomers of about 77 kDa, a profile fully compatible with a trimeric arrangement. Nevertheless, the functionality of the recombinant protein was reduced in comparison with the native molecule [31].
Beta class of sbPLIs (sbβPLIs)
Beta-type inhibitors are acidic, leucine-rich glycoproteins of 150–160 kDa. The leucines are assembled as leucine-rich repeats (LRRs) in tandem. This particular arrangement creates horseshoe-shaped molecules, similarly to those observed in Toll-like receptors in general [12, 32]. The first sbβPLI described in the literature was purified from G. brevicaudus as a homotrimer (Table 2). The inhibitor is specific for basic PLA2s from homologous venom and forms a stable PLA2-sbβPLI complex at a 1:1 molar ratio [12, 33].
Subsequently, similar sbβPLIs were purified from two non-venomous Colubridae snakes: E. quadrivirgata and E. climacophora [26, 34] (Table 2). Besides nine LRRs of 24 amino acids each, all three known sbβPLIs display a proline-rich amino-terminal region and ten cysteines, eight of which are probably involved in disulfide bonds. The fully conserved LRR1 segment might be responsible for the specific binding of sbβPLIs to basic PLA2s [26].
Gamma class of sbPLIs (sbγPLIs)
Currently, the gamma class of phospholipase A2 inhibitors comprises the greatest number of endogenous sbPLIs (Table 3).
SbγPLIs are acidic glycoproteins characterized by two structural units of highly conserved repeats of half cysteines, known as three-finger motifs, such as those found in proteins belonging to the Ly-6 family, the urokinase-type plasminogen activator, and α-neurotoxins [35, 36]. A subclassification into classes 1 and 2 was subsequently proposed for sbγPLIs, based on predicted structural homologies to urokinase-type plasminogen activator receptor (u-PAR) or to Ly-6. The inhibitors with the highest homology to the u-PAR were located in class 1, whereas those more similar to Ly-6 were assigned to class 2 [37].
Another important characteristic of most sbγPLIs is a highly conserved proline-rich region [38]. Proline residues are commonly found in the flanking segments of protein–protein interaction sites. Known as proline brackets, they may play a structural role by protecting the integrity and conformation of the interaction sites in functional proteins [39].
SbγPLIs may be assembled as hetero- or homomeric molecules and a subclassification was proposed based on the monomer composition [40]. The sbγPLIs from elapids (Naja naja kaouthia, Notechis ater, Notechis scutatus and Oxyuranus scutellatus), colubrid (Elaphe quadrivirgata), Old World viperid (Gloydius brevicaudus) and hydrophiid (Laticauda semifasciata) were placed in subclass I (heteromeric). All these inhibitors are composed of two different subunits with distinct primary structures (called α and β, or A and B) typically under a 2:1 ratio for A and B, respectively.
Subclass II is comprised of homomeric sbγPLIs from New World viperid Bothrops asper, Cerrophidion godmani, and C. d. terrificus, as well as Malayopyton reticulatus (Pythonidae) and P. flavovirids (Viperidae) from the Old World. However, the identification of a secondary subunit, similar to the subunit B of heteromeric inhibitors, in the sbγPLI-IIs from C. d. terrificus, P. elegans P. flavoviridis and several Australian elapid species challenged the homomeric composition of those inhibitors [17, 41–43]. However, a single subunit remained in sbγPLI-IIs from M. reticulatus, C. godmani, and B. jararacussu. The last two were originally purified by affinity chromatography using the target PLA2s as an immobilized ligand, whereas the purified inhibitors were confirmed as being composed of single subunits A, as expected for sbγPLI-IIs. All three sbγPLI-IIs were fully functional as homomers [21, 22]. The actual contribution of the secondary subunits B to the full functionality of the sbγPLI-IIs, whenever applicable, remains to be clarified. It has been speculated that the subunit B might play a structural rather than a functional role in the sbγPLIs from Australian elapid species [42]. On the other hand, an ancestral role has been suggested for the subunit B compared to subunit A, in the sbγPLI from the Asian P. flavoviridis [43]. In any case, both subunits, A and B, may be present as a heterogeneous mixture of more and less conserved isoforms, therefore generating subtle structural changes depending on the combination of isoforms, and increasing the PLA2-binding repertoire of sbγPLIs [38, 41].
The sbγPLI from Asian Malayopython reticulatus
This inhibitor was denominated phospholipase inhibitor from python (PIP). The native protein is a glycosylated oligomer formed by six identical subunits of 23 kDa each. After full deglycosylation, the molecular mass of the subunits decreases to 20 kDa.
Native PIPs occur as hexamers of apparent molecular mass of 140 kDa. The monomer precursor in snake liver tissue has a 19-residue signal sequence and an open reading frame of 603 bp encoding for a 182-residue protein. PIPs neutralize both lethal and PLA2 activities of daboiatoxin – the major toxin of Daboia russelli siamensis snake venom – by forming a toxin-inhibitor complex at 1:1 molar ratio. A recombinant PIP homologue produced in Escherichia coli was shown to neutralize not only daboiatoxin PLA2 activity in vitro, but other toxic PLA2s belonging to groups I (from Elapidae snake venoms), II (from Viperidae snake venoms) and III (from bee venom) at inhibitor-enzyme molar ratios between 0.1 and 5.0. In addition, this PIP homolog inhibited the edematogenicity of bee venom PLA2 and daboiatoxin up to 92.1 and 78.2%, respectively [20].
The functional site of PIP was predicted based on the hypothesis of proline brackets, and the data were employed to design PIP-derived bioactive peptides [39]. In general, the inhibition of PLA2s by these peptides has been explained by the blockage of the hydrophobic channel of secreted PLA2, as presented by other known inhibitors of this enzyme type [44].
Among a number of linear and cyclic PIP-derived peptides tested, PGLPPLSLQNG decapeptide (called P-PB.III) was able to inhibit groups I, II and III of PLA2s, including PLA2 from human synovial fluid of arthritis patients belonging to subgroup IIA [45]. The heptadecapeptide LGRVDIHVWDGVYIRGR (named PNT.II) was found to selectively inhibit human secreted IIA-PLA2. It also reduces neurotoxin-induced high levels of secreted PLA2 in rat hippocampal homogenates and modulates joint destruction in a mouse model of human rheumatoid arthritis [44, 46, 47]. An analog of PNT.II, known as PIP18, has been more recently devised [48]. Besides potent neutralization effects against Crotalus adamanteus snake venom, PIP18 has shown high bactericidal action against a number of pathogens, in a dose-dependent manner, with a remarkable activity against Staphylococcus aureus. Topical application of PIP18 has also modulated in vivo wound repair in a mouse model of S. aureus infection [49].
A structurally-related PIP homolog was later isolated from another pythonid species, Python sebae. Despite displaying poor PLA2 inhibition activity, the primary structure is highly similar to that of PIP. Two subunits (A and B) were characterized in this novel molecule. Nevertheless, both of them display the same amino-terminal sequence and show no similarity with the previously described B subunits from typical heteromeric sbγPLI-Is [50]. As to the complete primary structure, subunits A and B in the PIP homolog differ in eight of 182 amino acids, which suggests that they are actually isoforms of subunit A. Henceforth, the homomeric character of PIP appears preserved in PIP homolog.
The sbγPLI from Latin American Crotalus durissus terrificus
The cDNA of C. d. terrificus sbγPLI – called Crotalus neutralizing factor (CNF) – encodes a 19-residue signal peptide characteristic of secreted proteins, followed by 181 amino acids in the mature protein, including sixteen cysteines. CNF is a glycosylated alpha1-globulin with a single N-linked carbohydrate site at Asn157 [51–54]. The carbohydrate moiety, however, is not essential for PLA2 inhibition, since CNF remains functional after enzymatic deglycosylation [17].
Native CNF is a globular-shaped, predominantly tetrameric molecule with an average molecular mass of 100 kDa in solution. It innately occurs as a mixture of non-glycosylated and glycosylated monomers of 22 kDa and 25 kDa, respectively [55]. The oligomerization of CNF is independent of the presence of carbohydrates, since it occurs equally with native or enzymatically deglycosylated monomers. Tyrosine residues at the interface of the monomers composing CNF may contribute to the oligomerization process, according to a theoretical structural model constructed for the inhibitor (available with DOI:10.5452/ma-avb44 at ModelArchive database). The U monomer of the crystallographic structure of urokinase plasminogen activator from Homo sapiens (PDB ID: 2FD6) was used as the template ab initio [17].
Besides inhibiting lethal and PLA2 actions of C. d. terrificus venom, CNF is also able to inhibit the lethal activity of heterologous viperid venoms, such as those from Bothrops alternatus, B. atrox, B. jararaca. B. jararacussu, B. moojeni, B. neuwiedi and Lachesis muta, but not that of the elapid Micrurus frontalis [51]. In relation to PLA2 inhibition of heterologous venoms, CNF is capable of fully inhibiting the PLA2 activity of crude venom and of a semi-purified fraction of L. muta, which comprise PLA2s of different ionic character. It is important to note that the crude venom of L. muta is about eight times more active than C. d. terrificus venom, whereas the aforementioned fraction displays even higher activity – almost 24 times higher than that of C. d. terrificus venom [56]. Soon after, investigations of a protein highly similar to CNF purified from C. d. terrificus serum, named crotoxin inhibitor from Crotalus serum (CICS) by Perales and co-workers, showed complex formation with monomeric and multimeric Viperidae β-neurotoxins [53]. The enzymatic activity of pancreatic and non-pancreatic PLA2 from mammals, bee venom and Elapidae venoms remained unaffected [57].
The natural target of CNF in homologous venom is crotoxin, a heterodimeric β-neurotoxin formed by an enzymatically inactive subunit (crotoxin A or CA) and a PLA2 counterpart (crotoxin B or CB). CA and CB are non-covalently bonded in the crotoxin complex (CA/CB) [58]. CNF is able to displace CA in the native crotoxin in vitro to form a non-toxic CNF/CB complex, most likely at a 1:1 molar ratio [52]. In the presence of CNF, the newly formed CNF/CB complex no longer interacts with the target acceptor of crotoxin on rat brain synaptosomes to deliver CB to cause its toxic effect [55]. The formation of a new nontoxic complex by displacement of CA from the native crotoxin was confirmed by studies on the interaction of CICS and heterodimeric β-neurotoxins (Mojave toxin, CbICbII from Pseudocerastes fieldi venom, and crotoxin itself) [57].
The interaction in CNF/CB complex may be reminiscent of the crotoxin-receptor interaction at the presynaptic site. Competitive binding experiments were performed in vitro on rat brain synaptosomes, in an attempt to clarify the role of CNF as a CB receptor. Although the model encompasses a unique mechanism with three molecules – the receptor on synaptosomal membrane, CNF, and CA – competing for one ligand (CB), comparable IC50 values of around 100 nM were found for both CNF and CA [55].
It has been suggested that amino-terminus, beta-wing and carboxyl-terminus regions of CB participate in the formation of CA/CB and CNF/CB complexes [59]. The counter segments in CA and CNF remain to be clarified. A screening of highly conserved regions in CNF and putative sbγPLIs from Latin American pit vipers, using the multiple EM for motif elicitation (MEME) software for multiple alignment [60] indicated the decapentapeptide QPFPGLPLSRPNGYY as the best consensus motif possibly involved in the PLA2 interaction. Compared to the aforementioned decapeptide P-PBIII from PIP, the motif displays an amino-acid deletion and two amino-acid substitutions in its internal decapeptide [61].
Conclusion
A brief review on past and recent achievements on sbPLIs is presented herein. Although they have been studied by different groups over the years, many gaps remain to be filled, especially concerning their action mechanism and scope. In the near future, a better understanding of sbPLIs may guide practical applications of these fascinating molecules in biotechnology and therapeutics on PLA2-related disorders.
Abbreviations
- BaltMIP:
-
Myotoxin inhibitor protein from Bothrops alternatus
- CA:
-
Crotoxin A
- CB:
-
Crotoxin B
- CICS:
-
Crotoxin inhibitor from Crotalus serum
- CNF:
-
Crotalus neutralizing factor
- GbPLI:
-
Phospholipase A2 inhibitor from Gloydius brevicaudus
- LRR:
-
Leucine-rich repeats
- MEME:
-
Multiple EM for motif elicitation
- MIP:
-
Myotoxin inhibitor protein
- PIP:
-
Phospholipase A2 inhibitor from Python
- PLA2 :
-
Phospholipase A2
- sbPLI:
-
Snake blood phospholipase inhibitor
- u-PAR:
-
urokinase-type plasminogen activator receptor
References
Abalos JW. The ophiophagus habits of Pseudoboa cloelia. Toxicon. 1963;1:90.
Bonnett DE, Guttman SI. Inhibition of moccasin (Agkistrodon piscivorus) venom proteolytic activity by the serum of the Florida king snake (Lampropeltis getulus floridana). Toxicon. 1971;9(4):417–25.
Clark WC, Voris HK. Venom neutralization by rattlesnake serum albumin. Science. 1969;164(3886):1402–4.
Deoras PJ, Mhasalkar VB. Antivenin activity of some snake sera. Toxicon. 1963;1(2):89–90.
Fontana F. La venin de la Vipere n’est point un poison pour son espece. In: Fontana F. La Traité sur le vénin de la vipere sur les poisons Americains, sur le laurier-cerise et sur quelques autres poisons vegetaux. Tome Premier. Florence, Italy; 1781. p.22-7
Omori-Satoh T, Sadahiro S, Ohsaka A, Murata R. Purification and characterization of an antihemorrhagic factor in the serum of Trimeresurus flavoviridis, a crotalid. Biochim Biophys Acta. 1972;285(2):414–26.
Kihara H. Studies on the phospholipase A2 in Trimeresurus flavoviridis venom. III. Purification and some properties of phospholipase A2 inhibitor in Habu serum. J Biochem. 1976;80(2):341–9.
Kogaki H, Inoue S, Ikeda K, Samejima Y, Omori-Satoh T, Hamagushi K. Isolation and fundamental properties of a phospholipase A2 inhibitor from the blood plasma of Trimeresurus flavoviridis. J Biochem. 1989;106(6):966–71.
Ovadia M, Kochva E, Moav B. The neutralization mechanism of Vipera palestinae neurotoxin by a purified factor from homologous serum. Biochim Biophys Acta. 1977;491(2):370–86.
Rosenfeld S, Glass S. The inhibiting effect of snake bloods upon the hemorrhagic action of viper venoms on mice. Am J Med Sci. 1940;199(4):482–6.
Kochva E. Evolution and secretion of venom and its antidotes in snakes. Period Biol. 1978;80 Suppl 1:11–23.
Ohkura N, Okuhara H, Inoue S, Ikeda K, Hayashi K. Purification and characterization of three distinct types of phospholipase A2 inhibitors from the blood plasma of the Chinese mamushi, Agkistrodon blomhoffii siniticus. Biochem J. 1997;325(Pt 2):527–31.
Quirós S, Alape-Girón A, Angulo Y, Lomonte B. Isolation, characterization and molecular cloning of AnMIP, a new alpha-type phospholipase A2 myotoxin inhibitor from the plasma of the snake Atropoides nummifer (Viperidae: Crotalinae). Comp Biochem Physiol B Biochem Mol Biol. 2007;146(1):60–8.
Oliveira CZ, Menaldo DL, Marcussi S, Santos-Filho NA, Silveira LB, Boldrini-França J, et al. An alpha-type phospholipase A2 inhibitor from Bothrops jararacussu snake plasma: structural and functional characterization. Biochimie. 2008;90(10):1506–14.
Santos-Filho NA, Fernandes CA, Menaldo DL, Magro AJ, Fortes-Dias CL, Estevão-Costa MI, et al. Molecular cloning and biochemical characterization of a myotoxin inhibitor from Bothrops alternatus snake plasma. Biochimie. 2011;93(3):583–92.
Soares AM, Marcussi S, Stabeli RG, Franca SC, Giglio JR, Ward RJ, et al. Structural and functional analysis of BmjMIP, a phospholipase A2 myotoxin inhibitor protein from Bothrops moojeni snake plasma. Biochem Biophys Res Commun. 2003;302(2):193–200.
Fortes-Dias CL, Ortolani PL, Fernandes CA, Lobo KR, Amaral de Melo L, Borges MH, et al. Insights on the structure of native CNF, an endogenous phospholipase A2 inhibitor from Crotalus durissus terrificus, the South American rattlesnake. Biochim Biophys Acta. 2014;1844(9):1569–79.
Ohkura N, Kitahara Y, Inoue S, Ikeda K, Hayashi K. Isolation and amino acid sequence of a phospholipase A2 inhibitor from the blood plasma of the sea krait, Laticauda semifasciata. J Biochem. 1999;125(2):375–82.
Okumura K, Inoue S, Ikeda K, Hayashi K. cDNA cloning and bacterial expression of phospholipase A2 inhibitor PLIalpha from the serum of the Chinese mamushi, Agkistrodon blomhoffii siniticus. Biochim Biophys Acta. 1999;1441(1):51–60.
Thwin MM, Gopalakrishnakone P, Kini RM, Armugam A, Jeyaseelan K. Recombinant antitoxic and anti-inflammatory factor from the nonvenomous snake Python reticulatus: phospholipase A2 inhibition and venom neutralizing potential. Biochemistry. 2000;39(31):9604–11.
Lizano S, Angulo Y, Lomonte B, Fox JW, Lambeau G, Lazdunski M, et al. Two phospholipase A2 inhibitors from the plasma of Cerrophidion (Bothrops) godmani which selectively inhibit two different group-II phospholipase A2 myotoxins from its own venom: isolation, molecular cloning and biological properties. Biochem J. 2000;346(Pt 3):631–9.
Oliveira CZ, Santos-Filho NA, Menaldo DL, Boldrini-Franca J, Giglio JR, Calderon LA, et al. Structural and functional characterization of a γ-type phospholipase A2 inhibitor from Bothrops jararacussu snake plasma. Curr Top Med Chem. 2011;11(20):2509–19.
Estevão-Costa MI, Fernandes CA, Mudadu Mde A, Franco GR, Fontes MR, Fortes-Dias CL. Structural and evolutionary insights into endogenous alpha-phospholipase A2 inhibitors of Latin American pit vipers. Toxicon. 2016;112:35–44.
Zelensky AN, Gready JE. The C-type lectin-like domain superfamily. FEBS J. 2005;272(24):6179–217.
Nobuhisa I, Inamasu S, Nakai M, Mimori T, Ogawa T. Characterization and evolution of a gene encoding a Trimeresurus flavoviridis serum protein that inhibits basic phospholipase A2 isozymes in the snake’s venom. Eur J Biochem. 1997;249(3):838–45.
Shirai R, Toriba M, Hayashi K, Ikeda K, Inoue S. Identification and characterization of phospholipase A2 inhibitors from the serum of the Japanese rat snake, Elaphe climacophora. Toxicon. 2009;53(6):685–92.
Okumura K, Inoue S, Ikeda K, Hayashi K. Identification and characterization of a serum protein homologous to alpha-type phospholipase A2 inhibitor (PLIalpha) from a nonvenomous snake, Elaphe quadrivirgata. IUBMB Life. 2003;55(9):539–45.
Inoue S, Shimada A, Ohkura N, Ikeda K, Samejima Y, Omori-Satoh T, et al. Specificity of two types of phospholipase A2 inhibitors from the plasma of venomous snakes. Biochem Mol Biol Int. 1997;41(3):529–37.
Nishida M, Okamoto M, Ohno A, Okumura K, Hayashi K, Ikeda K, et al. Inhibitory activities of the heterotrimers formed from two α-type phospholipase A2 inhibitory proteins with different enzyme affinities and importance of the intersubunit electrostatic interaction in trimer formation. Biochim Biophys Acta. 2010;1804(11):2121–7.
Okumura K, Ohno A, Nishida M, Hayashi K, Ikeda K, Inoue S. Mapping the region of the alpha-type phospholipase A2 inhibitor responsible for its inhibitory activity. J Biol Chem. 2005;280(45):37651–9.
Santos-Filho NA, Boldrini-França J, Santos-Silva LK, Menaldo DL, Henrique-Silva F, Sousa TS, et al. Heterologous expression and biochemical and functional characterization of a recombinant alpha-type myotoxin inhibitor from Bothrops alternatus snake. Biochimie. 2014;105:119–28.
Kobe B, Deisenhofer J. Proteins with leucine-rich repeats. Curr Opin Struct Biol. 1995;5(3):409–16.
Okumura K, Ohkura N, Inoue S, Ikeda K, Hayashi K. A novel phospholipase A2 inhibitor with leucine-rich repeats from the blood plasma of Agkistrodon blomhoffii siniticus. Sequence homologies with human leucine-rich alpha2-glycoprotein. J Biol Chem. 1998;273(31):19469–75.
Okumura K, Inoue S, Ikeda K, Hayashi K. Identification of beta-type phospholipase A2 inhibitor in a nonvenomous snake, Elaphe quadrivirgata. Arch Biochem Biophys. 2002;408(1):124–30.
Ohkura N, Inoue S, Ikeda K, Hayashi K. The two subunits of a phospholipase A2 inhibitor from the plasma of Thailand cobra having structural similarity to urokinase-type plasminogen activator receptor and Ly-6 related proteins. Biochem Biophys Res Commun. 1994;204(3):1212–8.
Ohno M, Ménez R, Ogawa T, Danse JM, Shimohigashi Y, Fromen C, et al. Molecular evolution of snake toxins: is the functional diversity of snake toxins associated with a mechanism of accelerated evolution? Prog Nucleic Acid Res Mol Biol. 1998;59:307–64.
Hains PG, Nield B, Sekuloski S, Dunn R, Broady K. Sequencing and two-dimensional structure prediction of a phospholipase A2 inhibitor from the serum of the common tiger snake (Notechis scutatus). J Mol Biol. 2001;312(4):875–84.
Dunn RD, Broady KW. Snake inhibitors of phospholipase A2 enzymes. Biochim Biophys Acta. 2001;1533(1):29–37.
Kini RM. Proline brackets and identification of potential functional sites in proteins: toxins to therapeutics. Toxicon. 1998;36(11):1659–70.
Lizano S, Domont G, Perales J. Natural phospholipase A2 myotoxin inhibitor proteins from snakes, mammals and plants. Toxicon. 2003;42(8):963–77.
So S, Murakami T, Ikeda N, Chijiwa T, Oda-Ueda N, Kuraishi T, et al. Identification and evolution of venom phospholipase A2 inhibitors from Protobothrops elegans serum. Biosci Biotechnol Biochem. 2011;75(3):480–8.
Hains PG, Broady KW. Purification and inhibitory profile of phospholipase A2 inhibitors from Australian elapid sera. Biochem J. 2000;346(Pt 1):139–46.
So S, Chijiwa T, Ikeda N, Nobuhisa I, Oda-Ueda N, Hattori S, et al. Identification of the B subtype of gamma-phospholipase A2 inhibitor from Protobothrops flavoviridis serum and molecular evolution of snake serum phospholipase A2 inhibitors. J Mol Evol. 2008;66(3):298–307.
Thwin MM, Samy RP, Satyanarayanajois SD, Gopalakrishnakone P. Venom neutralization by purified bioactive molecules: Synthetic peptide derivatives of the endogenous PLA2 inhibitory protein PIP (a mini-review). Toxicon. 2010;56(7):1275–83.
Thwin MM, Satish RL, Chan ST, Gopalakrishnakone P. Functional site of endogenous phospholipase A2 inhibitor from python serum. Eur J Biochem. 2002;269(2):719–27.
Thwin MM, Ong WY, Fong CW, Sato K, Kodama K, Farooqui AA, et al. Secretory phospholipase A2 activity in the normal and kainate injected rat brain, and inhibition by a peptide derived from python serum. Exp Brain Res. 2003;150(4):427–33.
Thwin MM, Douni E, Aidinis V, Kollias G, Kodama K, Sato K, et al. Effect of phospholipase A2 inhibitory peptide on inflammatory arthritis in a TNF transgenic mouse model: a time-course ultrastructural study. Arthritis Res Ther. 2004;6(3):R282–94.
Samy RP, Thwin MM, Stiles BG, Bow H, Chow VT, Gopalakrishnakone P. Therapeutic potential of peptides with neutralizing ability towards the venom and toxin (CaTx-I) of Crotalus adamanteus. Curr Top Med Chem. 2011;11(20):2540–55.
Samy RP, Thwin MM, Stiles BG, Satyanarayana-Jois S, Chinnathambi A, Zayed ME, et al. Novel phospholipase A2 inhibitors from Python serum are potent peptide antibiotics. Biochimie. 2015;111:30–44.
Donnini S, Finetti F, Francese S, Boscaro F, Dani FR, Maset F, et al. A novel protein from the serum of Python sebae, structurally homologous with type-γ phospholipase A2 inhibitor, displays antitumour activity. Biochem J. 2011;440(2):251–62.
Fortes-Dias CL, Fonseca BC, Kochva E, Diniz CR. Purification and properties of an antivenom factor from the plasma of the South American rattlesnake (Crotalus durissus terrificus). Toxicon. 1991;29(8):997–1008.
Fortes-Dias CL, Lin Y, Ewell J, Diniz CR, Liu TY. A phospholipase A2 inhibitor from the plasma of the South American rattlesnake (Crotalus durissus terrificus). Protein structure, genomic structure, and mechanism of action. J Biol Chem. 1994;269(22):15646–51.
Perales J, Villela C, Domont GB, Choumet V, Saliou B, Moussatché H, et al. Molecular structure and mechanism of action of the crotoxin inhibitor from Crotalus durissus terrificus serum. Eur J Biochem. 1995;227(1–2):19–26.
Fortes-Dias CL, Diniz CR, Kochva E. Neutralization by homologous plasma of Crotalus durissus terrificus (South American rattlesnake) venom and crotoxin. Ciênc Cult. 1990;42(7):501–6.
dos Santos RM, Oliveira LC, Estevão-Costa MI, de Lima ME, Santoro MM, Fortes-Dias CL. Inhibition of crotoxin binding to synaptosomes by a receptor-like protein from Crotalus durissus terrificus (the South American rattlesnake). Biochim Biophys Acta. 2005;1717(1):27–33.
Fortes-Dias CL, Jannotti ML, Franco FJ, Magalhães A, Diniz CR. Studies on the specificity of CNF, a phospholipase A2 inhibitor isolated from the blood plasma of the South American rattlesnake (Crotalus durissus terrificus). I. Interaction with PLA2 from Lachesis muta muta snake venom. Toxicon. 1999;37(12):1747–59.
Faure G, Villela C, Perales J, Bon C. Interaction of the neurotoxic and nontoxic secretory phospholipases A2 with the crotoxin inhibitor from Crotalus serum. Eur J Biochem. 2000;267(15):4799–808.
Habermann E, Breithaupt H. Mini-review. The crotoxin complex--an example of biochemical and pharmacological protein complementation. Toxicon. 1978;16(1):19–30.
Fortes-Dias CL, dos Santos RMM, Magro AJ, Fontes MRM, Chávez-Olórtegui C, Granier C. Identification of continuous interaction sites in PLA2-based protein complexes by peptide arrays. Biochimie. 2009;91(11–12):1482–92.
Bailey TL, Williams N, Misleh C, Li WW. MEME: discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 2006;34(Web Server Issue):W369–73.
Estevão-Costa MI, Rocha BC, De Alvarenga Mudado M, Redondo R, Franco GR, Fortes-Dias CL. Prospection, structural analysis and phylogenetic relationships of endogenous gamma-phospholipase A2 inhibitors in Brazilian Bothrops snakes (Viperidae, Crotalinae). Toxicon. 2008;52(1):122–9.
Lizano S, Lomonte B, Fox JW, Gutiérrez JM. Biochemical characterization and pharmacological properties of a phospholipase A2 myotoxin inhibitor from the plasma of the snake Bothrops asper. Biochem J. 1997;326(Pt 3):853–9.
Ohkura N, Inoue S, Ikeda K, Hayashi K. Isolation and amino acid sequence of a phospholipase A2 inhibitor from the blood plasma of Agkistrodon blomhoffii siniticus. J Biochem. 1993;113(4):413–9.
Inoue S, Kogaki H, Ikeda K, Samejima Y, Omori-Satoh T. Amino acid sequences of the two subunits of a phospholipase A2 inhibitor from the blood plasma of Trimeresurus flavoviridis. Sequence homologies with pulmonary surfactant apoprotein and animal lectins. J Biol Chem. 1991;266(2):1001–7.
Shimada A, Ohkura N, Hayashi K, Samejima Y, Omori-Satoh T, Inoue S, et al. Subunit structure and inhibition specificity of alpha-type phospholipase A2 inhibitor from Protobothrops flavoviridis. Toxicon. 2008;51(5):787–96.
Nobuhisa I, Chiwata T, Fukumaki Y, Hattori S, Shimohigashi Y, Ohno M. Structural elements of Trimeresurus flavoviridis serum inhibitors for recognition of its venom phospholipase A2 isozymes. FEBS Lett. 1998;429(3):385–9.
Lima RM, Estevão-Costa MI, Junqueira-de-Azevedo IL, Ho PL, Diniz MR, Fortes-Dias CL. Phospholipase A2 inhibitors (βPLIs) are encoded in the venom glands of Lachesis muta (Crotalinae, Viperidae) snakes. Toxicon. 2011;57(1):172–5.
Okumura K, Masui K, Inoue S, Ikeda K, Hayashi K. Purification, characterization and cDNA cloning of a phospholipase A2 inhibitor from the serum of the non-venomous snake Elaphe quadrivirgata. Biochem J. 1999;341(Pt 1):165–71.
Le Z, Li X, Yuan P, Liu P, Huang C. Orthogonal optimization of prokaryotic expression of a natural snake venom phospholipase A2 inhibitor from Sinonatrix annularis. Toxicon. 2015;108:264–71.
Hains PG, Sung KL, Tseng A, Broady KW. Functional characteristics of a phospholipase A2 inhibitor from Notechis ater serum. J Biol Chem. 2000;275:983–91.
Gimenes SN, Ferreira FB, Silveira AC, Rodrigues RS, Yoneyama KA, Izabel Dos Santos J, et al. Isolation and biochemical characterization of a γ-type phospholipase A2 inhibitor from Crotalus durissus collilineatus snake serum. Toxicon. 2014;81:58–66.
Fortes-Dias CL, Barcellos CJ, Estevão-Costa MI. Molecular cloning of a gamma-phospholipase A2 inhibitor from Lachesis muta muta (the bushmaster snake). Toxicon. 2003;41(7):909–17.
Okumura K, Inoue S, Ohkura N, Ikeda K, Hayashi K. cDNA cloning of the two subunits of phospholipase A2 inhibitor PLIgamma from blood plasma of the Chinese mamushi, Agkistrodon blomhoffii siniticus. IUBMB Life. 1999;48(1):99–104.
Acknowledgments
Thanks are due to the Center for the Study of Venoms and Venomous Animals (CEVAP) of UNESP for enabling the publication of this paper (Edital Toxinologia CAPES no. 063/2010, Process no. 230.38.006285/2011-21, AUXPE Toxinologia 1219/2011).
Funding
The present study was funded by CAPES (Edital Toxinologia 063/2010, Process no. 230.38.006280/2011-07, AUXPE 1810/11), FAPEMIG, and CNPq/FAPESP through the National Institute of Science and Technology in Toxins (INCTTox). FAPEMIG granted productivity fellowships to CLFD, PCC and LAM. GLFD is an undergraduate student of Chemical Engineering at the Pontifical University of Minas Gerais (PUC/Minas) granted a scientific initiation fellowship from FAPEMIG (Program PIBIC/FAPEMIG/FUNED).
Authors’ contributions
PCC and LAM equally contributed to this review. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Campos, P.C., de Melo, L.A., Dias, G.L.F. et al. Endogenous phospholipase A2 inhibitors in snakes: a brief overview. J Venom Anim Toxins Incl Trop Dis 22, 37 (2016). https://doi.org/10.1186/s40409-016-0092-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40409-016-0092-5