
Abstract
By the emergence of recombinant DNA technology, many antibody fragments have been developed devoid of undesired properties of natural immunoglobulins. Among them, camelid heavy-chain variable domains (VHHs) and single-chain variable fragments (scFvs) are the most favored ones. While scFv is used widely in various applications, camelid antibodies (VHHs) can serve as an alternative because of their superior chemical and physical properties such as higher solubility, stability, smaller size, and lower production cost. Here, these two counterparts are compared in structure and properties to identify which one is more suitable for each of their various therapeutic, diagnosis, and research applications.
Similar content being viewed by others


Background
Antibodies (Abs) are distinguished binding tools for targeting almost any biomarker specifically. Although the inherent high affinity and specificity of immunoglobulins (Ig) is achieved by series of somatic hyper mutations and affinity maturation processes in the B cells, the recombinant DNA technology facilitates in-vitro production of various antibodies for a diverse set of targets [1]. Up to date, with about 100 FDA-approved antibodies in the market [2], the monoclonal antibody is a 145 billion dollar industry with 11% growth rate [3].
In structure, immunoglobulin (Ig) consists of two separate regions that can be dissociated by proteolytic cleavage with papain and pepsin; namely Fragment antigen-binding (Fab) domain and fragment crystallizable (Fc) region. While Fc region initiates biological processes upon antigen binding, Fab is responsible for antigen recognition, and the binding specificity of the whole Ig molecule is solely dependent on this domain, especially the two variable domains on the top-variable heavy chain (VH) and variable light chain (VL) [4]. This modular structure of immunoglobulin enabled scientists to introduce many structural modifications on the structure of Abs to improve their performance, by means of protein engineering and recombinant DNA technology. Smaller antibody fragments (such as Fab, scFv, diabodies, triabodies, mini bodies, and single-domain antibodies) are among these modified structures, designed to be reliable alternatives for conventional antibodies. Their smaller size, superior properties, and ease of manufacturing while retaining the targeting specificity of the whole Ig molecule make them perfect tools for diagnosis and clinical applications [5, 6].
Among all the engineered and recombinant antibody formats, single-chain variable fragments (scFvs) and camelid heavy-chain variable domains (VHHs) - also known as nanobodies- are the most popular ones. Previously scientists considered single-chain variable fragment (scFv) -composed of VH and VL- as the smallest antibody fragment with the same antigen-binding specificity to the whole Ig molecule. However, the discovery of camelid VHH [7] and shark variable new antigen receptor (VNAR) [8] demonstrated that a single V-like domain can retain the affinity of a whole antibody molecule [9]. Due to the broad and similar applications of scFv and VHH, this article aims to review the differences of these two antibody fragments in structure and function to illustrate whether the superior properties of nanobodies can make them a capable alternative for scFvs or not.
Nanobody and scFv in structure
As stated previously, the variable domains of Fab are responsible for the binding specificity of the whole antibody. Therefore, the smallest unit of Ig with antigen-binding activity is the fragment variable or Fv in which the two variable domains (VH and VL) connect with a disulfide bond. ScFv is an engineered form of Fv that, instead of a disulfide bond, the two variable domains are joined together by a flexible linker (Fig. 1). The length and amino acid composition of this linker play an important role in correct folding of the protein [10], and it is typically 10-25 amino acid long with Glu Lys stretches to increase the solubility and Gly Ser stretches for the flexibility of the final protein [11, 12]. Within each of the two variable domains of the scFv, there are three hyper variable domains or complementary determining regions (CDRs) that are linked together with framework regions (FRs). While the CDRs are responsible for antigen binding, and their structure is complementary to the epitope, the remainder of the variable domains (FRs) acts as a scaffold and has inconsiderable variability compared to CDRs. Interestingly, the contribution of each CDR in antigen binding is different. For instance, the CDR3 in the heavy chain has a critical role by 29% contribution in binding specificity while the involvement of CDR2L is just 4% [13].

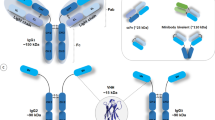
The differences of scFv and nanobody in structure. A conventional IgG structure, scFv structure composed of VH and VL of antibody joined with a flexible linker, the detailed structure of VH in scFv. B camel HCAb structure, nanobody that is derived from camelid VHH, the detailed structure of VHH
The higher contribution of the VH in antigen binding raised a research hypothesis that whether a single heavy chain can retain the parent Ab's binding affinity. In seminal studies, mouse single variable domains were investigated for their functional activity [14]. However, their troublesome properties, such as low affinity, poor solubility, and higher production cost, hampered their broader development. The discovery of heavy chain only antibodies (HCAbs) in camelids [7] and immunoglobulin new antigen receptor (IgNAR) in cartilaginous fish [15] was a new beginning in single domain antibody development. The antigen-binding domain of these specific immunoglobulins (VHH and V-NAR) is a high affinity single V-like domain that has evolved to be devoid of the disadvantages of previous single domain fragments. These superior properties result from major adaptations in sequence and structure [16,17,18,19]. As camelid nanobodies have easier handling, more robust antibody responses [17, 20], and higher yield in recombinant expression [21] than shark V-NARs, VHH fragments are more frequently used and will be the focus of this review.
Similar to VH, camelid VHHs comprise nine beta-strands forming a typical IgV fold; however, VL loss caused notable differences between these two fragments, especially in FR2 and hypervariable loops. In the conventional VH region, the FR2 consist of four highly conserved hydrophobic amino acids (Val37, Gly44, Leu45, and Trp47) that in contribution with Gln39, Gly44, Tyr91, and Trp103 form a conserved hydrophobic interface of ~700 Å2 to facilitate VL joining [22]. With the absence of VL in nanobodies, these four hydrophobic residues are substituted for more hydrophilic amino acids (Phe37, Glu44, Arg45, and Gly47) [23,24,25] to avoid the exposure of such a sizeable hydrophobic region to solvent (Fig. 1). In addition to this substitution, residues adjacent to this interface have rotated their side chains without deforming the Cαbackbone to increase the VHH surface's hydrophilicity. Furthermore, the CDR3 domain of VHH folds over this interface to shield the amino acids formerly covered by the VL partner [26]. These alterations elucidate the augmented solubility of VHHs in comparison to the single VH domain and scFvs [27].
In VHHs, extension in hypervariable loops repays the loss of three VL CDRs and VH–VL combinatorial diversity. Extension of CDR1 and CDR3 provides a 600–800 Å2 antigen-interacting surface as offered by six loops from the VH–VL domain [24, 28]. Furthermore, the elongated CDR1, with mutational hotspots imprinted in the VHH germline, compensates for the VL partner's variability [29] while the somatic mutations in 28 and 30 residues of CDR1 are selected during the affinity maturation process to participate in antigen binding directly [30, 31]. Although the elongated CDR3 can extend into epitopes that are almost inaccessible for specific antibodies, the enlarged loop suggests broader flexibility, impeding antigen-binding entropically [24, 28]. To solve this issue, camellia VHHs evolved with an extra disulfide bond toward either the CDR1, CDR2, or FR2 [24, 28, 32]. All these structural features increase the paratope diversity and allow for a wide variety of geometrical loop structures that deviate fundamentally from the canonical loop structures defined for conventional antibodies and facilitate the orientation of the CDR3 toward the antigens [33, 34].
Nanobody and scFv in properties
As a result of significant structural differences, scFv and Nb display distinct properties in vitro and in vivo. These different characterizations are investigated below in more detail (Table 1).
Size
First and foremost, these two fragments have notable dissimilarity in their size, while scFv is almost twice the Nb size by about 30 kDa weight [35]. This smaller size facilitates VHHs genetic manipulation [36], and the presence of only three antigen-binding loops allows for easy enhancement of their intrinsic tendency to antigen [37]. Due to the renal filtration and degradation, the smaller size of VHHs also results in their short half-life in blood [38]. This feature can be helpful since it results in high tissue permeability but unfavorable because their molecular weight is below glomerular filtration cutoff size (65kDa) and makes problems in some clinical therapies requiring antibody circulation over extended periods.
This limitation has led to the development of half-life extension strategies that combine VHHs with additional molecules. One of the most popular ones is addition of stabilizing groups such as Poly-ethylene glycol (PEG) molecules that slow down blood clearance rate with high tumor or any other target site accumulation. Fusion with long-circulating serum proteins like albumin or albumin's specific binders, effectively increase the VHH half-life in the blood. Fc fusion can also stabilize them in blood while provoking the immune system to the target site. Furthermore, Fc or albumin fusion makes the antibody fragment size larger and implements FcRn-mediated recycling to increase the protein half-life in the blood [39].
Solubility and Stability
As described previously, the substitution of four highly conserved hydrophobic amino acids for more hydrophilic residues in VHH leads to significant differences in properties between Nbs and scFvs. In scFvs, these four residues (V37, G44, L45, and W47) in FR2 form a hydrophobic interface to facilitate VH-VL joining. However, on the downside, this hydrophobic region lowers scFvs’ solubility, resulting in their high tendency for aggregation. The substitution of polar and smaller amino acid residues in this position (F37or Y37, E44, R45, and G47) makes them more hydrophilic and, consequently, more soluble than scFvs. Furthermore, this nonpolar to polar transition leads to the molecular and thermodynamic stability of VHHs in comparison to scFvs. Therefore, Nbs are more resistant to chemical denaturants and protease enzymes [40] and have higher stability under harsh PH or ionic strength [41]. This higher conformational stability also stems from the presence of an extra disulfide bond, which lowers the probability of heat-induced aggregation and limits VHHs flexibility [42,43,44,45,46,47]. Because of higher stability, they show high refolding efficiency, which means raising or lowering the sample temperature does not affect Nb conformation, i.e., it de-binds and binds to the target, respectively, without any aggregation or denaturation [48].
This rigidity in structure is a favorite property in the clinic since non-native protein aggregation is a common downside of antibody treatment, raising the immune response in severe cases [49, 50]. However, although in scFvs the hydrophobic interface between VL and VH dampens their stability, this two domain structures make them more flexible and more advantageous for some applications.
Production process
Both scFvs and Nbs are generated from immune or naive libraries that will be screened to discover high-affinity fragments against our desired target. Despite significant similarities in library screening, the mechanism of library construction is less challenging for Nbs. VHH libraries are created from blood serum of either naive or immunized camelids. After mRNA isolation, cDNA is reverse transcribed by reverse transcription-polymerase chain reaction (RT-PCR). This pool of VHH sequences is applied to construct a library screened by versatile display technologies such as phage display to discover a specific VHH for any potential antigen [51]. Although the inaccessibility of a camel, dromedary, llama, or alpaca to immunize is a bottleneck, HCAbs generating transgenic mice, or commercial naïve or synthetic Nb libraries can circumvent this issue.
For scFvs, libraries are typically created from either naive or immunized murine or human. Similar to VHH, VH, and VL genes obtain from RT-PCR [52,53,54,55], but in contrast to Nbs, an extra step is required to connect VH and VL cDNAs through SOE-PCR. Pairing these fragments is a challenging step in the library construction of scFvs due to the low efficiency of this technique [56, 57]. Since the mispaired VH and VL developed by this method may not detect and bind to the target antigen [58].
After isolation of a specific scFv or nanobody, attaining a high expression yield is challenging, especially for scFvs. In scFvs, as discussed earlier, the intrinsic hydrophobic interaction between VH and VL domains leads to their higher tendency to aggregate. Therefore it is complicated to express them in various expression systems appropriately. Many methods should be employed to improve these fragments’ stability, such as loop grafting, altering specific positions in their structure, and random mutagenesis [59]. Nevertheless, the superior properties of VHHs, such as their high hydrophilicity lead to a less demanding production cycle with fewer steps [60].
Bacterial expression of scFvs can occur in the either cytoplasmic or periplasmic environment. Each of these systems has its own limitations for the proper formation of scFv. On the one hand, while chaperones and disulfide isomerases in periplasm are preferable for appropriate folding of scFvs [10], they lead to a lower production yield [61]. Furthermore, unpaired cysteine residues of scFvs can form covalent bonds with the periplasmic protein's cysteine, which results in aggregation [62, 63]. Adding a signal peptide such as Pel B should also be considered to guide scFv into the periplasm [61]. On the other hand, the reducing environment of the cytoplasm hampers disulfide bond formation, which leads to exposure of hydrophobic VH-VL patches to the solvent and production of insoluble aggregate forms of scFvs called inclusion bodies. These aggregated products have to be re-folded in one additional time-consuming, costly, and ineffective step with the need of denaturing agents like urea [64].
Another approach to lower scFv aggregation promoted by inter-domain hydrophobic interaction is the humanization of scFvs by replacing hydrophobic amino acids with hydrophilic residues to prevent accumulation. Although protein solubility has improved by this method, these replacements can also have minor effects on the antigen-binding affinity of the final scFv [65]. The single entity nature of VHH makes their production much easier besides scFvs. In VHH cytoplasmic expression, there are no such hydrophobic- interaction-related issues [66], which lower production costs [10].
scFvs are not efficiently expressed in the yeast expression system as well. Due to the higher hydrophobicity of scFvs, they cannot be produced with proper folding in Saccharomyces cerevisiae's Endoplasmic Reticulum (ER) [67]. Some extra refolding steps, such as co- or over-expression of chaperons, should be applied to overcome these limitations [68]. In contrast, S. cerevisiae is the best expression system for VHH [69], as organelles like ER or Golgi ensure proper disulfide bonds and glycosylation [70]. Because of these difficulties, E.coli remains the best host for scFv expression [10, 64, 68].
Immunogenicity
One of the significant drawbacks of scFvs is their rodent origin, as the Hybridoma technique is only well developed for mice and rats and not for humans [71]. Murine VL and VH exhibit only 53% and 51% sequence identity, respectively, with corresponding regions in humans [72], while nanobodies display high sequence similarity with human VH (VH3 gene family) with ~75–90% identity correlated with their Low immunogenicity in clinical applications [73, 74]. As a result, the humanization process is more straightforward in VHHs. Even after the humanization of murine-derived scFv, the variable regions of scFv can elicit an anti-idiotypic response because eliminating critical residues in this region may affect antigen binding [75,76,77]. Also, scFvs’ engineering for reducing human anti-mouse antibody (HAMA) responses [78] will inactivate [79] injected scFvs and lessen their clinical effectiveness [80, 81], and allergic reactions will arise in repeated administration [78, 79]. Furthermore, humanization reduces the binding affinity of these fragments [10, 65], and CDR grafting may represent new immunogenic epitopes [75,76,77, 82]. In general, the humanization of murine-derived scFvs can overcome these immunogenicity problems to some extent but not entirely [59].
Affinity
Although scFvs and Nbs provide similar affinity, they show a distinct preference for epitopes. Nbs have better access for grooves and clefts on the surface of antigens like ion channels [83], viral glycoproteins [84], or immune synapses [85], but scFvs prefer flat linear epitopes. These differences result from the longer CDR3 loop in Nb, allowing for a highly convex shape to access concave epitopes. Nbs also show a good affinity for flat epitopes suggesting that these fragments can form various interface complexes. Furthermore, nonspecific background binding is lower for nanobodies in comparison to scFvs [37].
Nanobody and scFv in application
In recent years, by the rapid progress in antibody fragments engineering, the smaller size of scFv and VHH makes them suited for a broad range of applications ranging from therapy and diagnosis to research and exploration. Here the various utilizations of these prominent antibody fragments are discussed in detail to demonstrate which counterpart is more appropriate for each application.
Therapeutic applications of Nb and scFv in various formats
Development in targeted medicine has expanded treatment options, particularly in cancer therapy [45]. ScFv and VHH as high-affinity antibody fragments play an important role in targeted medicine [86] and are utilized in various formats (Fig. 2) as therapeutic options for several conditions.

Different Formats of Nanobody and scFv in therapeutic applications
Unconjugated neutralizing agent
ScFv and VHH can be utilized as neutralizing agents through direct binding and inactivation of foreign particles such as toxins and viruses [87] and cancer antigens or disease mediating cytokines and growth factors. These antibody fragments are administered naked in monovalent or multivalent format to block the function of their target or fused to an Fc region to increase serum half-life and trigger the immune system.
Nb is mostly a preferable candidate for this concern because of five main reasons. a) Their smaller size, resulting in perfect diffusion in tissues and extending neutralizing performance beyond the vessels. b) Higher flexibility and binding tendency enabling the bivalent and trivalent conjugation of Nanobodies and improving neutralization [88]. c) Their lower immunogenicity and more straightforward humanization process [89]. d) High stability that makes VHH production easier. e) Structural properties of VHHs that provide the power to target inaccessible epitopes [90].
Fatal toxins are a neglected but vital health problem all around the world. Plasma anti-venom serum (PAS) therapy is passive immunotherapy against toxins, with a rapid, effective response of direct antibody injection. Toxin neutralizers need to have rapid diffusion and clearance to provide enough power to identify and neutralize toxins with high tissue penetration [89] and then uptaken by kidneys in a shortened period. Therefore, antibody fragments such as nanobody and scFv are suitable for toxin inactivation, and scFvs were successfully applied in vitro. In a study done by Miethe and his colleagues, a neutralizing scFv-Fc inhibited the endopeptidase activity of botulinum neurotoxin [91]. In another paper, scFv was utilized to neutralize scorpion toxin Cn2 [88]. However, these fragments were not effective in vivo because of their high serum half-life [92]. In contrast, Nbs, with their smaller size and stronger binding affinity, have shown promising results in toxin inactivation in vivo [89]. Anti-scorpions toxin nanobody was able to protect mice more quickly than PAS therapy from Androctonus australis hector (Aah) scorpions [88]. In another study, despite the nanobody mixture failed to stop the venom lethality, the results proved the efficacy and usefulness of VHH developed against Bothrops atrox snake venom [93], which highlights the potential of VHH as anti-toxin agents.
Antibody fragments can neutralize viruses as well. In the last decades, one of the most successful approaches against viral agents was neutralizing monoclonal antibodies, which can suppress viral load partly through humoral immunity [94]. Nanobodies derived from Camelid and murine scFv are considered candidates for neutralizing viral agents because of their smaller size, higher solubility, and stability. Virus-neutralizing nanobodies have been developed against several animal and human virus families such as HIV-1 [95], human respiratory syncytial virus (hRSV) [96], and H5N1 Influenza [97]. The glycoprotein envelope of the virus canyons is filled with VHH, therefore the virus cannot conjugate with a co-receptor on the cell surface [98]. Bivalent and trivalent fragments are more efficient in identifying similar or different epitopes on the envelope glycoproteins, e.g., trivalent nanobodies against HIV [95] and hRSV [96] extended the neutralization breadth compared to the monovalent format. ScFvs are also utilized as viral neutralizing agents. Recombinant scFv has been utilized for targeting many viruses such as HIV [99], influenza [100], Porcine epidemic diarrhea virus (PEDV) [101], and HPV [102]. However, nanobody is more creditable because its smaller size provides the power to match and fill virus canyons more efficiently.
During the current SARS-Co-2 pandemic, anti-viral agents are one of the considerable therapeutic candidates. The receptor-binding domain (RBD) of the spike protein of the virus and the human angiotensin-converting enzyme 2 (ACE2) receptor on the cell surface are the key components in the viral entry that various forms of antibody fragments can target. Nanobodies [103] and scFvs are both appropriate candidates for RBD blocking, but Nbs are more common. The theranostic potential of camelid nanobodies in covid-19 infections is reviewed recently [104, 105]. Although a handful of scientific groups are working on finding the best nanobody set for SARS-cov2 neutralization [106,107,108,109], attempts are still enduring to discover a Recombinant scFv for this mission [110, 111]. Nevertheless, the superior properties of nanobodies for viral neutralization, such as lower production cost, the potential for aerosol delivery because of their high stability, and ease of multimerization put them forward in this race.
In terms of cancer immunotherapy, antibody fragments can inhibit tumor evasion by binding to the tumor itself or blocking the vital component for tumor growth and invasion. As growth factors play a critical role in angiogenesis, especially in the tumor microenvironment, blocking them or their cognate receptor can enhance tumor regression. Both scFv and VHH are utilized to bind and neutralize Vascular endothelial growth factor receptor 2 (VEGFR-2) [112, 113]. It is also demonstrated that the epidermal growth factor receptor (EGFR) blocked by a monovalent and bivalent nanobody called CONAN-1 could be a potential anticancer therapy [114].
Cytokine and chemokine inhibition is a potential therapeutic option for cancer and autoimmune disease [115]. Camelid VHHs and scFvs have been utilized for blocking several inflammatory and immunomodulatory cytokines as a treatment for rheumatoid arthritis [116] and chronic inflammation [117, 118], autoimmune inflammatory diseases [119] as well as cancer [86, 120], and stroke [121]. Cytokine- nanobody complex performs a sufficient neutralizing capacity when its half-life increases using albumin binding. It may act as a superior option in reduction of neuroinflammatory response in brain ischemia by high penetration, even crossing the brain-blood barrier [121]. Moreover, they can inhibit the enzymatic activity of their target protein, such as carbonic anhydrase (CAIX), by blocking their active site [122]. However, the efficiency of antibody fragment mediated immunotherapy can be further augmented by their conjugation with other therapeutic payloads discussed in the next section.
Conjugated to therapeutic payloads
Antibody fragments can be chemically or genetically conjugated to effector domains to deliver therapeutic payloads to a specific target, thereby reducing nonspecific toxicity to normal cells. Furthermore, this conjugation makes them larger, which results in increased circulation time in the blood [123].
One of the familiar domains that are genetically fused to these antibody fragments is toxins. Upon binding to the target cell, the complex is taken in through endocytosis, and finally, the toxin kills the desired cell- usually tumor or virally infected tissue [124]. Both scFv and nanobody have been utilized as an immunotoxin by conjugation with bacterial toxins like Enterobacter cloacae β-Lactamase [125] and Pseudomonas Exotoxin A [126, 127], or less immunogenic human origin cytotoxic elements such as TRAIL [128,129,130] and granzyme B [131].
Immunocytokines are another group of antibody fragment conjugates that can induce immune cell proliferation and anti-tumor activity in cancer immune therapy. Many studies demonstrated the effective function of these conjugates in specific anti-tumor activity [132,133,134,135]. The most popular cytokines delivered by antibody fragments are IL-2, IL-12, and TNF, whose systemic administration may lead to serious side effects [136].
Although scFv and nanobody can both serve as targeting agents for immunotoxins and immune cytokines, since tissue penetration is critical, especially in cancer, VHH is a better option for conjugation with larger fragments. However, the larger size of scFvs makes it suitable for conjugation with a smaller domain such as siRNA [137, 138]. Nevertheless, the superior physicochemical properties, easier humanization and better antigen recognition properties, and higher stability make camelid VHH preferable even in siRNA delivery [139].
Conjugated to immune cell engaging antibody fragments
Due to the critical role of the immune system in cancer, dual-specific antibody fragments can link between tumor and immune system components. Bite or bispecific T cell engagers are the most familiar form of these bispecific conjugates. By binding to CD3 with one of their recognition domains, they can activate T cell-mediated targeted tumor lysis. Bites are historically constructed from scFvs and scFv based the FDA now approves CD3 × CD19 BiTE to treat B-cell acute lymphoblastic leukemia [140]. However, camelid VHH can perform in each binding domain with a smaller size and higher modularity. Nevertheless, in recent studies, these two counterparts are mixed. For example, Harwood et al. created a novel form of T cell engagers named ATTACK by linking three anti-EGFR nanobodies with an anti-CD3 scFv as an ideal format for developing the next generation of T cell-redirecting bispecific antibodies [141].
Other immune system components such as NK cells γδTcells and APC can be redirected to the tumor site similarly by targeting their cognate marker. CD16 for engaging NK cells [142,143,144], T cell receptor (TCR) of Vγ9Vδ2 cells [145] and CD11b for attracting γδTcells, MHC-II or other specific surface proteins for targeting APCs [146, 147].
Targeting moiety of nanocarriers
Because of the binding specificity of these targeting agents, nanobodies and scFvs are broadly used in several drug delivery platforms to deliver their cargo to its specific location. High penetration, stability, pH-temperature resistance, and low aggregation are factors that shed light on VHH importance as a mediating delivery agent [148]. These delivery systems can overcome limitations of drug conjugate systems, such as poor chemical and enzymatic stability, solubility, rapid blood clearance, and adverse side effects to normal tissues. These nanocarriers can also release their cargo in more extended periods to decrease the frequency of drug administration [149]. Both VHH and scFv have been employed as a targeting moiety of nanocarriers such as liposomes [150,151,152], micelles [153, 154], albumin-based nanoparticles (NANAPs) [155, 156], and polymer-based NPS [157, 158]. But studies demonstrate the superiority of Nb in this regard [151]. In a study conducted by Oliveira et al., anti-EGFR-nanobody liposomes downregulate EGFR expression while its scFv based counterpart was unable to do so. The reason behind this effect is probably the sensitive structure of scFv that loses its proper folding in the acidic condition of lysosome after liposome internalization [151]. Furthermore, as these nanocarriers’ primary target is surface receptors that mediate cellular internalization [148], Nb, with its smaller size, is preferable to act as an antagonist for these receptors.
Targeting moiety of viruses
Viruses are eminent gene therapy agents applied for the treatment of various diseases and as a vaccine to induce an immune response against any desired protein. However, these highly efficient gene delivery systems cannot perform specifically by only infecting their target cells [159]. The binding specificity of antibody fragments is helpful to circumvent this issue. Many lentiviral and adenoviral vectors have been modified by either Nb or scFv [159,160,161,162,163,164], but the smaller size and high stability make it a better option in this regard. The superiority of Nb as a binding domain of adenovirus is perfectly investigated by Poulin et al. the result of this study demonstrate that the single domain antibody construct was efficiently incorporated into the Ad capsid and enhanced virus infection of cells expressing the targeted receptor while the scFv construct incorporated into the capsid at a very low level, insufficient to retarget virus infection [164].
Intrabody
Although most studies investigate the potential of antibody fragments in targeting extracellular antigens, most signaling pathways occur intracellularly [165]. Therefore targeting these intracellular factors may be effective in the treatment of various diseases, including cancer [166] and neurodegenerative conditions [167] such as Huntington's disease [168]. Intrabodies have been developed to express and operate within the cell and bind to an intracellular protein specifically. Because the reducing environment of cytoplasm is the main playground of intrabodies, stability under harsh conditions is the factor that makes Nbs outshine all the other antibody fragments as well as scFvs. However, the therapeutic potential of antibodies is hindered by the lack of an effective delivery system [165]. Viral delivery of intrabody coding sequence to the target cell is the most eminent method [167]. However, bacterial systems such as the type III secretion system (T3SS) of E. coli have been utilized to translocate the translated form of intrabody into their eukaryotic target.
Antigen binding domain in synthetic Receptors
Today with the impressive advancement in synthetic biology, a handful of synthetic receptors have been designed to sense and respond to extracellular signals in a programmable way [169, 170]. CAR (Chimeric antigen receptor) is the most popular form of these receptors that recently entered the clinic [171]. Similar to native receptors, the extracellular domain of these proteins is responsible for sensing extracellular factors. To improve the system's modularity and enable recognizing any desired factor, the extracellular part of these receptors is mainly composed of an antibody-derived fragment, such as scFv or nanobody. Although scFv is the most widely used form of the extracellular domain [169, 172], the superior properties of nanobody and its smaller size make it a preferable option in this position [172, 173]. In several studies, camelid VHHs have proven their efficiency as the antigen-binding moiety of CAR T cells [172,173,174,175,176,177,178,179]. Furthermore, the utilization of a bispecific nanobody-based construct in the receptor's extracellular domain broadens CAR T cell therapy specificity by recognizing two distinct antigens simultaneously [180].
Camellia VHH can serve as antigen binding domain in other less popular receptors as well. For instance, nanobodies have been utilized in the recently designed SynNotch receptor [181], GEMA [182], and C-STAR [183]. These new studies suggest that maybe in the future, nanobodies can take the position of scFvs in the extracellular domain of synthetic receptors due to their favorable properties such as high stability, smaller size, and low immunogenicity.
Diagnostic applications of Nb and scFv
Although antibody fragments are well known for their therapeutic application, the binding specificity of these targeting agents suit them for several diagnosis applications. Not only in medical diagnosis but also environmental and food analysis applications. Their broad utilization in diagnosis is summarized in the following paragraphs.
Molecular imaging
Antibody fragments can serve as specific molecular probes for the detection of various disease-related antigens. Especially in cancer treatment, molecular imaging is critical for the early detection of tumor progression or monitoring the success or failure of the therapy in cancer patients. mAbs were the first high-affinity probes in molecular imaging, but their weak tumor penetration and high serum half-life lead to low contrast images with a low tumor to background ratio [184]. Smaller antibody fragments could circumvent this issue; however, Nb, as the smallest antibody-derived targeting domain, is superior in this regard. Compared with scFv and Fab, Nbs were shown to be promising probes for molecular imaging due to high tumor uptake, rapid blood clearance, low liver uptake, and high stability [185]. However, the rapid renal clearance of Nbs result in high signals in the kidneys and bladder, therefore they are not favorable for imaging at their nearby sites [37].
To date, several imaging techniques have been developed and applied for clinical application, and each of the antibody fragments is labeled with different agents to be visualized in that specific imaging technique [186,187,188]. Positron emission tomography (PET), single-photon emission computed tomography (SPECT) are the most popular imaging technique with many radionucleotides labeled nanobodies in clinical trials [189].
Immunoassays
إecause of the antigen-binding activity of immunoglobulins, the various format of the antibody-based immune assay, such as enzyme-linked immunosorbent assay (ELISA) and lateral flow immune assays (LFA) is designed for detection of various factors in medical diagnosis and environmental and food analysis. The mAb is the widely used targeting agent in various immunoassays, but high stability, lower production cost, and the ability to recognize the epitopes out of reach for larger fragments make Nbs a favorable immurement [184]. Nobody-based immune assays have shown promising results in detecting T. solium [190] and Trypanosoma [191], with better outcomes than their whole Ab counterparts.
To date, several formats of nanobody and scFv based immunoreagents have been utilized for detecting microorganisms, natural proteins, and chemicals in various specimens [192,193,194,195,196,197,198,199,200,201]. These detection systems are also packages as a point of care detection device known as a biosensor to enable portability, easier handling, and decrease production costs [202].
Research applications of Nb and scFv
The application of antibody fragments is not limited to theranostics. These high-affinity binders are a proficient tool in research applications to understand the structure and function of proteins. Therefore, in this part, we mainly mention the use of Nbs and scFvs in fundamental research.
Protein visualization studies
Due to inefficient folding and chain assembly of scFvs, Nbs are better candidates to be stably expressed and then trace proteins in living cells [48, 203].i.e., reducing intracellular environment hampers proper folding and formation of disulfide bonds [204] in scFvs, results in their poor function and stability [205, 206], .therefore their use as intrabodies is restricted, and just a few of them have been used in fundamental biology researches. Nevertheless, Nbs do not show this limitation and can fold accurately inside the cells, which favors their use as a research tool for versatile applications over scFvs. , Furthermore adding a tag to label and visualize cytoplasmic proteins by vhhs is less challenging than scFvs as they show higher stability. Because of the single domain nature of Nbs, adding a tag does not influence their binding activity [207], while in scFvs, it is harder to add a tag without affecting the binding affinity
The first method for tracing any desired protein in developmental biology was fusing it with a fluorescent protein-like GFP. However, the intracellular expression of a protein and a fluorescent protein may influence the correct formation and function of the protein. To overcome such challenges, chromobodies were developed by fusing nanobodies with a fluorescent tag like RFP. Although target-specific chrome bodies can circumvent many challenges in this regard, the production of a specific nanobody for each target is labor-intensive. Hence targeting the fusion domains by chrome bodies enables the tracing of various proteins by a single nanobody. To date, many tracer nanobodies against small peptide fusions like SunTag and PTM have been developed; however, anti-GFP VHHs, termed (GFP binding proteins) GBPs, are extensively employed against GFP fusion proteins [208].
Another method of protein visualization used in fluorescence microscopy is imaging in a sandwich format utilizing a fluorescently labeled secondary antibody to detect the target bound Nb [209, 210]. However, in super-resolution microscopy, since it is more efficient to have a less distance between the target molecule and fluorescent label [211, 212], coupling organic dyes to GBPs can directly visualize any GFP labeled structure [213, 214]. In addition, chromobodies expressed in living cells can trace endogenous targets, therefore they are also employed in super-resolution imaging techniques [215]. In general, compared to scFvs, the smaller size of VHHs allows for a more accurate determination of the target's location in microcopy [216].
Protein function studies
Understanding the interactions of a protein to its surrounding environment is a critical step in studying the function of the desired protein. Antibody fragments as high-affinity binders can serve as a professional tool in studying protein-protein or even RNA-protein interactions. A study done by Sheetz et al. evaluated the function of human scFv for understanding the biogenesis of a subset of oncogenic microRNAs by using an anti-NCL scFv, they demonstrated that NCL is a critical protein in cancer biogenesis as it reacts with oncogenic microRNA. These antibody fragments can also serve as primary antibodies in ELISA [217, 218].
Another approach for study protein-protein interaction is GBP-based fluorescent-three-hybrid. In this approach, GBP is first coupled with an intracellular anchoring protein resulting in its fixation in the predetermined subcellular compartment. Then two target proteins which each one is fused with a fluorescent tag, interact with the central GBP binding protein. Therefore a GFP–RFP colocalization signal is produced at the location of GBP, which can be monitored to improve our understanding of protein-protein interaction [219].
Another method to understand the function of a protein is inactivation by degradation or interfering in the function, therefore the role of knockout protein in the pathway can be revealed in its absence. Unlike RNAi, Nanobodies, by their smaller size, can bind to the effector domain of desired proteins to defunctionalize that particular domain and enable the fundraising of its role in the process. In contrast, by RNA interference, the function of the whole protein would be disrupted, and the role of each domain will remain unclear [220].
Nbs can knock out their target protein by inducing protein degradation [221,222,223,224]. For instance, GBP nanobodies were combined with the F-box domain (which is a component of the SKP1–CUL1–F-box (SCF) ubiquitin E3 ligases complex) and so initiated polyubiquitination, which results in degradation of intracellular GFP-fused protein through proteasome [225]. Another approach is adding a PEST motif to Nb so that the target antigen will undergo a proteasomal degradation pathway [226].
Another step to studying a protein function is understanding its location of function. GBP nanobodies combined with monomeric RFP can localize GFP fusion proteins as they bind to GFP, and the resulting GFP–chromobody shows the location of GFP fusion proteins in nuclei, cytoplasm, or even membrane [227]. Nb is also a valuable tool to impose a new location for a protein of interest inside a cell. Nb fusion to localization domains redirect the POI to a new cellular compartment leading to its relocalization. In this manner, the role of location on protein function during animal development can be understood. If POI is tagged with a fluorescent molecule, this relocalization can also be monitored by fluorescent microscopy [219]. For instance, Nb coupled to KDEL peptide keeps its antigen inside the endoplasmic reticulum and coupling Nb with lamin molecule locate the antigen to the nuclear membrane [228]. Adding a tag to Nb can also immobilize the antigen is side the cell membrane and prevent its diffusion outside the cell [215, 229, 230]. By preventing the spread of secreted proteins, their effects on neighbor cells can be identified in developmental biology [231].
Nanobodies can also act as a scaffold that binds to two or more different targets in a non-overlapping manner. These target proteins can be DNA binding domains, and transcription factors as a ‘transcription device dependent on GFP’ (T-DDOG) [232] controls gene expression. This tool utilizes two GBPs, one of these nanobodies is equipped with a DNA-binding domain, and the other joined with the activation domain of the viral protein VP16. In GFP expressing living cells, these two Nbs come together in the presence of the GFP, leading to the desired gene expression.
Protein structure studies
Protein crystallography is one of the standard methods for understanding the structure of any desired protein. However, because proteins are highly dynamic, they are bound to crystallization chaperons for reduced conformational heterogeneity. Antibody fragments are superior among various binding partners since they can be engineered to target almost any desired protein [233]. In scFvs, the hydrophobic interface between VL and VH dampens their stability, and this two domain-structure make them more flexible than single-domain fragments like VHHs. This feature hampers their utility as a crystallization chaperon to understand an unknown and challenging protein [37].
In contrast, Nbs lower intrinsic flexibility show effective crystal formation, and it is easier to identify the conformation and mechanism of epitope binding by them rather than scFvs. VHHs can reveal protein conformations in each folding step, so transient intermediates can be determined even of highly dynamic protein [234] without inducing any out-of-native structure. To date, the structure of many proteins has been determined by VHHs, including high-value G protein-coupled receptor [235] and amyloid proteins in different pathological conditions [236, 237].
Affinity purification and Immunoprecipitation
Due to Nbs superior properties, including small size, monovalent mode, and easy directional immobilization to solid substrates, they can purify an increased amount of biomolecules in chromatography [24, 28, 238]. De Genest et al. developed VHHs against Glu–Pro–Glu–Ala (EPEA) in highly efficient affinity chromatography for any EPEA-tagged protein purification [239]. Nbs show lower nonspecific background binding than larger antibody formats like scFvs; besides, their high refolding efficiency and denaturation resistance allows for repeated column regeneration with only milder elution buffer, which later matters for sensitive targets [240]. Nanobodies can precipitate serum immunoglobins like IgG, so with their help, we can purify different types of antibodies from the blood. For example, Klooster et al. could purify human protein HSA and IgG from blood with the help of Nbs [241]. Furthermore, they are also perfect candidates for antibody-based slide, bead arrays [238], and chromatin immunoprecipitation with DNA microarray (chIP-on-chip) to discover new transcription factor-binding sites [242].
Conclusion and future perspective
Antibodies as well-known targeting moieties are utilized for a handful of applications. The emergence of recombinant DNA technology enabled the design and construction of various antibody fragments, including Camelid nanobody and scFv. These two Ab fragments are the most widespread ones with a broad range of applications. Here we compared these fragments in structure and properties to investigate which of these antigen-binding domains is preferable for each application. In General, the higher stability, solubility, and lower production cost of Nb make it favorable in almost all applications, while its smaller size acts as a double-edged sword.
In applications that favor rapid clearance of targeting agents, such as molecular imaging or anti-venom therapy, the smaller size of nanobody is advantageous. While in other therapeutic utilizations that require persistence in the body, including neutralizing foreign invaders or targeting cancer, half-life extension strategies should be employed to increase the size of the targeting agent. On the downside, this size increment is against tissue permeability which is a critical property in cancer treatment. Therefore balancing the tradeoff between serum half-life and tissue penetration is necessary to achieve an optimized size for cancer antibody-based therapeutics.
Another issue that makes choosing between these two counterparts controversial is their preferred antigenic site. In other words, because of the structural difference between scFv and nanobodies, they prefer different epitopes while binding to the same antigen. Nbs are high-affinity binders for grooves and clefts, while scFvs prefer flat linear epitopes. Therefore, the antigen structure is an essential factor to decide which one is a better candidate for targeting the desired protein.
In spite of the superior properties of Nbs, scFvs are still dominant in the clinic with about ten FDA approved products and more than 80 ongoing clinical trials (Table 2). However, in less than 30 years since the discovery of HCAb, the superior properties of this single domain targeting agent make it a pioneer in the field of recombinant Ab engineering. In 2019 the first Nanobody entered the clinic and there are more than 27 Nb based drug candidate in clinical trials, waiting for FDA approval (Table 3). Nevertheless, because of the long history behind scFvs and other fragments derived from conventional Ab, nanobodies still have a long way to become a dominant player in recombinant antibody market.
Availability of data and materials
Not applicable.
Abbreviations
- Ab:
-
Antibody
- Nb:
-
Nanobody
- scFv:
-
Single-chain variable fragment
- Ig:
-
Immunoglobulin
- Fab:
-
Fragment antigen-binding
- Fc:
-
Fragment crystallizable
- VH:
-
Variable heavy chain
- VL:
-
Variable light chain
- V-NAR:
-
Shark variable new antigen receptor
- CDR:
-
Complementary determining regions
- FR:
-
Framework region
- HCAb:
-
Heavy chain only antibody
- IgNAR:
-
Immunoglobulin new antigen receptor
- FcRn:
-
Neonatal Fc receptor
References
Liu JKH. The history of monoclonal antibody development - Progress, remaining challenges and future innovations. Ann Med Surg. 2014;3:113–6.
Kaplon H, Muralidharan M, Schneider Z, Reichert JM. Antibodies to watch in 2020. MAbs. 2020;12(1) [cited 2021 Sep 4] Available from: /pmc/articles/PMC6973335/.
Antibodies Market Size, Share Trends, Growth Analysis | 2021 to 2026 [Internet]. [cited 2021 Sep 4]. Available from: https://www.marketdataforecast.com/market-reports/antibodies-market
Madison S. Introduction to critical ethnography. Theory Method. 2005;911(1):1–16.
Khan FH. Antibodies and Their Applications. Animal Biotechnology. 2013:473–90. Available from:. https://doi.org/10.1016/B978-0-12-416002-6.00025-0.
Holliger P, Hudson PJ. Engineered antibody fragments and the rise of single domains. Nat Biotechnol. 2005;23(9):1126–36.
Hamers-Casterman C, Atarhouch T, Muyldermans S, Robinson G, Hammers C, Songa EB, et al. Naturally occurring antibodies devoid of light chains. Nature. 1993;363(6428):446–8 [cited 2020 Sep 12] Available from: https://www.nature.com/articles/363446a0.
Streltsov VA, Carmichael JA, Nuttall SD. Structure of a shark IgNAR antibody variable domain and modeling of an early-developmental isotype. Protein Sci. 2005;14(11):2901–9 [cited 2020 Sep 12]. Available from: /pmc/articles/PMC2253229/?report=abstract.
Gonzalez-Sapienza G, Rossotti MA, Tabares-da RS. Single-domain antibodies as versatile affinity reagents for analytical and diagnostic applications. Front Immunol. 2017;8:977 [cited 2020 Aug 31]. Available from: www.frontiersin.org.
Ahmad ZA, Yeap SK, Ali AM, Ho WY, Alitheen NBM, Hamid M. ScFv antibody: Principles and clinical application. Clin Dev Immunol. 2012;2012 [cited 2020 Aug 31]. Available from: https://pubmed.ncbi.nlm.nih.gov/22474489/.
Alfthan K, Takkinen K, Sizmann D, Söderlund H, Teeri TT. Properties of a single-chain antibody containing different linker peptides. Protein Eng Des Sel. 1995;8(7):725–731. [cited 2020 Sep 20]. Available from: https://academic.oup.com/peds/article/8/7/725/1465664
Whitlow M, Bell BA, Feng SL, Filpula D, Hardman KD, Hubert SL, et al. An improved linker for single-chain fv with reduced aggregation and enhanced proteolytic stability. Protein Eng Des Sel. 1993;6(8):989–95 [cited 2020 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/8309948/.
Wilson IA, Stanfield RL. Antibody-antigen interactions: new structures and new conformational changes. Curr Opin Struct Biol. 1994;4(6):857–67.
Ward ES, Güssow D, Griffiths AD, Jones PT, Winter G. Binding activities of a repertoire of single immunoglobulin variable domains secreted from Escherichia coli. Nature. 1989;341(6242):544–6. Available from. https://doi.org/10.1038/341544a0.
Greenberg AS, Avila D, Hughes M, Hughes A, McKinney EC, Flajnik MF. A new antigen receptor gene family that undergoes rearrangement and extensive somatic diversification in sharks. Nature. 1995;374(6518):168–73. Available from. https://doi.org/10.1038/374168a0.
Streltsov V, Nuttall S. Do sharks have a new antibody lineage? Acta Crystallogr Sect A Found Crystallogr. 2005;61(a1):c91 [cited 2020 Dec 19] Available from: //scripts.iucr.org/cgi-bin/paper?a32285.
Dooley H, Flajnik MF. Shark immunity bites back: Affinity maturation and memory response in the nurse shark, Ginglymostoma cirratum. Eur J Immunol. 2005;35(3):936–45 [cited 2020 Dec 19]. Available from: https://pubmed.ncbi.nlm.nih.gov/15688348/.
De Genst E, Handelberg F, Van Meirhaeghe A, Vynck S, Loris R, Wyns L, et al. Chemical basis for the affinity maturation of a camel single domain antibody. J Biol Chem. 2004;279(51):53593–601 [cited 2020 Dec 19]. Available from: https://pubmed.ncbi.nlm.nih.gov/15383540/.
De Genst E, Silence K, Ghahroudi MA, Decanniere K, Loris R, Kinne J, et al. Strong in vivo maturation compensates for structurally restricted H3 loops in antibody repertoires. J Biol Chem. 2005;280(14):14114–21 [cited 2020 Dec 19]. Available from: https://pubmed.ncbi.nlm.nih.gov/15659390/.
Crouch K, Smith LE, Williams R, Cao W, Lee M, Jensen A, et al. Humoral immune response of the small-spotted catshark, Scyliorhinus canicula. Fish Shellfish Immunol. 2013;34(5):1158–69 [cited 2020 Dec 19]. Available from: https://pubmed.ncbi.nlm.nih.gov/23439398/.
Zielonka S, Weber N, Becker S, Doerner A, Christmann A, Christmann C, et al. Shark Attack: High affinity binding proteins derived from shark vNAR domains by stepwise in vitro affinity maturation. J Biotechnol. 2014;191:236–45 [cited 2020 Dec 19]. Available from: https://pubmed.ncbi.nlm.nih.gov/24862193/.
Padlan EA. Anatomy of the antibody molecule. Mol Immunol. 1994;31:169–217 [cited 2021 Jan 10]. Available from: https://pubmed.ncbi.nlm.nih.gov/8114766/.
Muyldermans S, Atarhouch T, Saldanha J, Barbosa JARG, Hamers R. Sequence and structure of vh domain from naturally occurring camel heavy chain immunoglobulins lacking light chains. Protein Eng Des Sel. 1994;7(9):1129–35 [cited 2021 Jan 10]. Available from: https://pubmed.ncbi.nlm.nih.gov/7831284/.
Muyldermans S. Nanobodies: Natural single-domain antibodies. Annu Rev Biochem. 2013;82:775–97 [cited 2021 Jan 10]. Available from: https://pubmed.ncbi.nlm.nih.gov/23495938/.
Vu KB, Ghahroudi MA, Wyns L, Muyldermans S. Comparison of llama V(H) sequences from conventional and heavy chain antibodies. Mol Immunol. 1997;34(16–17):1121–31.
Muyldermans S, Cambillau C, Wyns L. Recognition of antigens by single-domain antibody fragments: The superfluous luxury of paired domains. Trends Biochem Sci. 2001;26:230–5.
Davies J, Riechmann L. “Camelising” human antibody fragments: NMR studies on VH domains. FEBS Lett. 1994;339(3):285–90 [cited 2021 Jan 10] Available from: https://pubmed.ncbi.nlm.nih.gov/8112468/.
Siontorou CG. Nanobodies as novel agents for disease diagnosis and therapy. International Journal of Nanomedicine. 2013;8:4215–27 Available from: /pmc/articles/PMC3818023/?report=abstract.
Nguyen VK, Hamers R, Wyns L, Muyldermans S. Camel heavy-chain antibodies: Diverse germline V(H) H and specific mechanisms enlarge the antigen-binding repertoire. EMBO J. 2000;19(5):921–30 [cited 2021 Jan 10]. Available from: /pmc/articles/PMC305632/?report=abstract.
Decanniere K, Desmyter A, Lauwereys M, Ghahroudi MA, Muyldermans S, Wyns L. A single-domain antibody fragment in complex with RNase A: Non-canonical loop structures and nanomolar affinity using two CDR loops. Structure. 1999;7(4):361–70 [cited 2021 Jan 10]. Available from: https://pubmed.ncbi.nlm.nih.gov/10196124/.
Desmyter A, Transue TR, Ghahroudi MA, Thi MHD, Poortmans F, Hamers R, et al. Crystal structure of a camel single-domain V(H) antibody fragment in complex with lysozyme. Nat Struct Biol. 1996;3(9):803–11 [cited 2021 Jan 10]. Available from: https://www.nature.com/articles/nsb0996-803.
Govaert J, Pellis M, Deschacht N, Vincke C, Conrath K, Muyldermans S, et al. Dual beneficial effect of interloop disulfide bond for single domain antibody fragments. J Biol Chem. 2012;287(3):1970–9 [cited 2021 Jan 10]. Available from: https://pubmed.ncbi.nlm.nih.gov/22128183/.
Flajnik MF, Deschacht N, Muyldermans S. A Case Of Convergence: Why Did a Simple Alternative to Canonical Antibodies Arise in Sharks and Camels? PLoS Biol. 2011;9(8):e1001120. [cited 2021 Jan 10]. Available from. https://doi.org/10.1371/journal.pbio.1001120.
Decanniere K, Muyldermans S, Wyns L. Canonical antigen-binding loop structures in immunoglobulins: More structures, more canonical classes? J Mol Biol. 2000;300(1):83–91 [cited 2021 Jan 10]. Available from: https://pubmed.ncbi.nlm.nih.gov/10864500/.
Bannas P, Hambach J, Koch-Nolte F, Johnson M. Nanobodies and nanobody-based human heavy chain antibodies as antitumor therapeutics. Front Immunol. 2017;8(NOV):1603 [cited 2020 Aug 10]. Available from: www.frontiersin.org.
Hassani M, Hajari Taheri F, Sharifzadeh Z, Arashkia A, Hadjati J, van Weerden WM, et al. Construction of a chimeric antigen receptor bearing a nanobody against prostate a specific membrane antigen in prostate cancer. J Cell Biochem. 2019;120(6):10787–95. Available from. https://doi.org/10.1002/jcb.28370.
Hassanzadeh-Ghassabeh G, Devoogdt N, De Pauw P, Vincke C, Muyldermans S. Nanobodies and their potential applications. Nanomedicine. 2013;8:1013–26.
Kim TY, Park JH, Shim HE, Choi DS, Lee D-E, Song J-J, et al. Prolonged half-life of small-sized therapeutic protein using serum albumin-specific protein binder. J Control Release. 2019;315:31–9.
Kontermann RE. Strategies to extend plasma half-lives of recombinant antibodies. BioDrugs. 2009;23:93–109 [cited 2021 May 19]. Available from: https://pubmed.ncbi.nlm.nih.gov/19489651/.
Hussack G, Mackenzie CR, Tanha J. Characterization of single-domain antibodies with an engineered disulfide bond. Methods Mol Biol. 2012;911:417–29.
Van Audenhove I, Gettemans J, Hussack G, Mackenzie CR, Tanha J. Nanobodies as Versatile Tools to Understand, Diagnose, Visualize and Treat Cancer. EBioMedicine. 2016;8:40–8. [cited 2020 May 13]. Available from. https://doi.org/10.1016/j.ebiom.2016.04.028.
Fernandes CFC, dos Pereira SS, Luiz MB, Zuliani JP, Furtado GP, Stabeli RG. Camelid single-domain antibodies as an alternative to overcome challenges related to the prevention, detection, and control of neglected tropical diseases. Front Immunol. 2017;8(JUN):1–8.
Jovčevska I, Muyldermans S. The Therapeutic Potential of Nanobodies. BioDrugs. 2020;34(1):11–26.
Kunz P, Ortale A, Mücke N, Zinner K, Hoheisel JD. Nanobody stability engineering by employing the Δtm shift; A comparison with apparent rate constants of heat-induced aggregation. Protein Eng Des Sel. 2019;32(5):241–9.
Lecocq Q, De Vlaeminck Y, Hanssens H, D’Huyvetter M, Raes G, Goyvaerts C, et al. Theranostics in immuno-oncology using nanobody derivatives. Theranostics. 2019;9(25):7772–91.
Kunz P, Zinner K, Mücke N, Bartoschik T, Muyldermans S, Hoheisel JD. The structural basis of nanobody unfolding reversibility and thermoresistance. Sci Rep. 2018;8(1):1–10.
Ewert S, Cambillau C, Conrath K, Plückthun A. Biophysical properties of camelid VHH domains compared to those of human VH3 domains. Biochemistry. 2002;41(11):3628–36.
Dmitriev OY, Lutsenko S, Muyldermans S. Nanobodies as probes for protein dynamics in vitro and in cells. J Biol Chem. 2016;291(8):3767–75.
Rosenberg AS. Effects of protein aggregates: an immunologic perspective. AAPS J. 2006;8(3):E501–7.
Roberts CJ. Therapeutic protein aggregation: mechanisms, design, and control. Trends Biotechnol. 2014;32(7):372–80.
Sabir JSM, Atef A, El-Domyati FM, Edris S, Hajrah N, Alzohairy AM, et al. Construction of naïve camelids VHH repertoire in phage display-based library. C R Biol. 2014;337(4):244–9.
Yip YL, Hawkins NJ, Clark MA, Ward RL. Evaluation of different lymphoid tissue sources for the construction of human immunoglobulin gene libraries. Immunotechnology. 1997;3(3):195–203.
Graus YF, Verschuuren JJ, Degenhardt A, van Breda Vriesman PJ, De Baets MH, Posner JB, et al. Selection of recombinant anti-HuD Fab fragments from a phage display antibody library of a lung cancer patient with paraneoplastic encephalomyelitis. J Neuroimmunol. 1998;82(2):200–9.
Rothe A, Klimka A, Tur MK, Pfitzner T, Huhn M, Sasse S, et al. Construction of phage display libraries from reactive lymph nodes of breast carcinoma patients and selection for specifically binding human single chain Fv on cell lines. Int J Mol Med. 2004;14(4):729–35.
Xu M-Y, Xu X-H, Chen G-Z, Deng X-L, Li J, Yu X-J, et al. Production of a human single-chain variable fragment antibody against esophageal carcinoma. World J Gastroenterol. 2004;10(18):2619–23.
Hilgarth RS, Lanigan TM. Optimization of overlap extension PCR for efficient transgene construction. MethodsX. 2020;7:100759.
Karkhane AA, Yakhchali B, Rastgar Jazii F, Bambai B, Aminzadeh S, Rahimi F. A Nested-Splicing by Overlap Extension PCR Improves Specificity of this Standard Method. Iran J Biotechnol. 2015;13(2):56–9.
Ingram JR, Schmidt FI, Ploegh HL. Exploiting Nanobodies’ Singular Traits. Annu Rev Immunol. 2018;36(1):695–715 [cited 2020 May 18]. Available from: http://www.annualreviews.org/doi/10.1146/annurev-immunol-042617-053327.
Borras L, Gunde T, Tietz J, Bauer U, Hulmann-Cottier V, Grimshaw JPA, et al. Generic approach for the generation of stable humanized single-chain Fv fragments from rabbit monoclonal antibodies. J Biol Chem. 2010;285(12):9054–66.
Bannas P, Hambach J, Koch-Nolte F. Nanobodies and nanobody-based human heavy chain antibodies as antitumor therapeutics. Front Immunol. 2017;8(NOV):1–13.
Bates A, Power CA. David vs. Goliath: The Structure, Function, and Clinical Prospects of Antibody Fragments. Antibodies. 2019;8(2):28.
Albrecht H, Burke PA, Natarajan A, Xiong C-Y, Kalicinsky M, DeNardo GL, et al. Production of soluble ScFvs with C-terminal-free thiol for site-specific conjugation or stable dimeric ScFvs on demand. Bioconjug Chem. 2004;15(1):16–26.
Schmiedl A, Breitling F, Winter CH, Queitsch I, Dübel S. Effects of unpaired cysteines on yield, solubility and activity of different recombinant antibody constructs expressed in E. coli. J Immunol Methods. 2000;242(1–2):101–14.
Spadiut O, Capone S, Krainer F, Glieder A, Herwig C. Microbials for the production of monoclonal antibodies and antibody fragments. Trends Biotechnol. 2014;32(1):54–60.
Khantasup K, Chantima W, Sangma C, Poomputsa K, Dharakul T. Design and Generation of Humanized Single-chain Fv Derived from Mouse Hybridoma for Potential Targeting Application. Monoclon Antib Immunodiagn Immunother. 2015;34(6):404–17.
Ingram JR, Schmidt FI, Ploegh HL. Exploiting Nanobodies ’ Singular Traits; 2018. p. 1–21.
Joosten V, Lokman C, Van Den Hondel CA, Punt PJ. The production of antibody fragments and antibody fusion proteins by yeasts and filamentous fungi. Microb Cell Fact. 2003;2(1):1.
Xu P, Raden D, Doyle FJ, Robinson AS. Analysis of unfolded protein response during single-chain antibody expression in Saccaromyces cerevisiae reveals different roles for BiP and PDI in folding. Metab Eng. 2005;7(4):269–79.
Chee MK, Haase SB. New and Redesigned pRS Plasmid Shuttle Vectors for Genetic Manipulation of Saccharomycescerevisiae. G3 (Bethesda). 2012;2(5):515–26.
van der Vaart JM. In: O’Brien PM, Aitken R, editors. Expression of VHH Antibody Fragments in Saccharomyces cerevisiae BT - Antibody Phage Display: Methods and Protocols. Totowa: Humana Press; 2002. p. 359–66.
Moldenhauer G. Selection Strategies for Monoclonal Antibodies. In: Handbook of Therapeutic Antibodies. Weinheim: John Wiley & Sons, Ltd; 2014. p. 15–42.
Kügler M, Stein C, Schwenkert M, Saul D, Vockentanz L, Huber T, et al. Stabilization and humanization of a single-chain Fv antibody fragment specific for human lymphocyte antigen CD19 by designed point mutations and CDR-grafting onto a human framework. Protein Eng Des Sel. 2009;22(3):135–47.
Vincke C, Loris R, Saerens D, Martinez-Rodriguez S, Muyldermans S, Conrath K. General strategy to humanize a camelid single-domain antibody and identification of a universal humanized nanobody scaffold. J Biol Chem. 2009;284(5):3273–84.
Harmsen MM, De Haard HJ. Properties, production, and applications of camelid single-domain antibody fragments. Appl Microbiol Biotechnol. 2007;77(1):13–22.
Tamura M, Milenic DE, Iwahashi M, Padlan E, Schlom J, Kashmiri SV. Structural correlates of an anticarcinoma antibody: identification of specificity-determining residues (SDRs) and development of a minimally immunogenic antibody variant by retention of SDRs only. J Immunol. 2000 Feb;164(3):1432–41.
Iwahashi M, Milenic DE, Padlan EA, Bei R, Schlom J, Kashmiri SVS. CDR substitutions of a humanized monoclonal antibody (CC49): contributions of individual CDRs to antigen binding and immunogenicity. Mol Immunol. 1999;36(15):1079–91.
Glaser SM, Vásquez M, Payne PW, Schneider WP. Dissection of the combining site in a humanized anti-Tac antibody. J Immunol. 1992;149(8):2607–14.
Berger M, Shankar V, Vafai A. Therapeutic applications of monoclonal antibodies. Am J Med Sci. 2002;324(1):14–30.
Legouffe E, Liautard J, Gaillard JP, Rossi JF, Wijdenes J, Bataille R, et al. Human anti-mouse antibody response to the injection of murine monoclonal antibodies against IL-6. Clin Exp Immunol. 1994;98(2):323–9.
Weiner LM. An overview of monoclonal antibody therapy of cancer. Semin Oncol. 1999;26(4 Suppl 12):41–50.
Funaro A, Horenstein AL, Malavasi F. Monoclonal antibodies in clinical applications. J Biol Regul Homeost Agents. 1996;10(4):72–82.
Stephens S, Emtage S, Vetterlein O, Chaplin L, Bebbington C, Nesbitt A, et al. Comprehensive pharmacokinetics of a humanized antibody and analysis of residual anti-idiotypic responses. Immunology. 1995;85(4):668–74.
Danquah W, Meyer-Schwesinger C, Rissiek B, Pinto C, Serracant-Prat A, Amadi M, et al. Nanobodies that block gating of the P2X7 ion channel ameliorate inflammation. Sci Transl Med. 2016;8(366):366ra162.
Henry KA, MacKenzie CR. Antigen recognition by single-domain antibodies: structural latitudes and constraints. MAbs. 2018;10(6):815–26.
Cartwright ANR, Griggs J, Davis DM. The immune synapse clears and excludes molecules above a size threshold. Nat Commun. 2014;5(1):5479.
Yang EY, Shah K. Nanobodies: Next Generation of Cancer Diagnostics and Therapeutics. Front Oncol. 2020;10:1182.
Blažek D, Celer V. The Production and Application of Single-Chain Antibody Fragments. Folia Microbiol (Praha). 2003;48(5):687–98. Available from. https://doi.org/10.1007/BF02993480.
Hmila I, Saerens D, Abderrazek RB, Vincke C, Abidi N, Benlasfar Z, et al. A bispecific nanobody to provide full protection against lethal scorpion envenoming. FASEB J. 2010;24(9):3479–89.
Ben Abderrazek R, Vincke C, Hmila I, Saerens D, Abidi N, El Ayeb M, et al. Development of Cys38 knock-out and humanized version of NbAahII10 nanobody with improved neutralization of AahII Scorpion toxin. Protein Eng Des Sel. 2011;24(9):727–35.
Safdari Y, Ahmadzadeh V, Khalili M, Jaliani HZ, Zarei V. Use of Single-Chain Antibody Derivatives for Targeted Drug Delivery; 2016. p. 258–70.
Miethe S, Rasetti-escargueil C, Liu Y, Pelat T, Avril A, Frenzel A, et al. Development of neutralizing scFv-Fc against botulinum neurotoxin A light chain from a macaque immune library Development of neutralizing scFv-Fc against botulinum neurotoxin A light chain from a macaque immune library, vol. 0862; 2014.
Quintero-Hernández V, Juárez-González VR, Ortíz-León M, Sánchez R, Possani LD, Becerril B. The change of the scFv into the Fab format improves the stability and in vivo toxin neutralization capacity of recombinant antibodies. Mol Immunol. 2007;44(6):1307–15.
Bailon Calderon H, Yaniro Coronel VO, Cáceres Rey OA, Colque Alave EG, Leiva Duran WJ, Padilla Rojas C, et al. Development of Nanobodies Against Hemorrhagic and Myotoxic Components of Bothrops atrox Snake Venom. Front Immunol. 2020;11(May):1–12.
Burton DR, Hangartner L. Broadly Neutralizing Antibodies to HIV and Their Role in Vaccine Design. Annu Rev Immunol. 2016;34(1):635–59 Available from: http://www.annualreviews.org/doi/10.1146/annurev-immunol-041015-055515.
Weiss RA, Verrips CT. Nanobodies that neutralize HIV. Vaccines. 2019;7(3):1–14.
Palomo C, Mas V, Detalle L, Depla E, Cano O, Vázquez M, et al. Trivalency of a nanobody specific for the human respiratory syncytial virus fusion glycoprotein drastically enhances virus neutralization and impacts escape mutant selection. Antimicrob Agents Chemother. 2016;60(11):6498–509.
De Filette M, Hultberg A, Verrips T, Temperton N, Weiss RA, Vandevelde W, et al. Nanobodies With In Vitro Neutralizing Activity Protect Mice Against H5N1 Influenza Virus Infection. J Infect Dis. 2011;203:1063–72.
Koch K, Kalusche S, Torres JL, Stanfield RL, Danquah W, Khazanehdari K, et al. Selection of nanobodies with broad neutralizing potential against primary HIV-1 strains using soluble subtype C gp140 envelope trimers. Sci Rep. 2017;7(1):1–15. Available from:. https://doi.org/10.1038/s41598-017-08273-7.
Tanaka K, Kuwata T, Alam M, Kaplan G, Takahama S, Valdez KPR, et al. Unique binding modes for the broad neutralizing activity of single-chain variable fragments (scFv) targeting CD4-induced epitopes. Retrovirology. 2017;14(1):1–15.
Lukosaityte D, Sadeyen JR, Shrestha A, Sealy JE, Bhat S, Chang P, et al. Engineered recombinant single chain variable fragment of monoclonal antibody provides protection to chickens infected with H9N2 avian influenza. Vaccines. 2020;8(1):118.
Zhang F, Chen Y, Ke Y, Zhang L, Zhang B, Yang L, et al. Single chain fragment variable (scFv) antibodies targeting the spike protein of porcine epidemic diarrhea virus provide protection against viral infection in piglets. Viruses. 2019;11(1):58.
Culp TD, Spatz CM, Reed CA, Christensen ND. Binding and neutralization efficiencies of monoclonal antibodies , Fab fragments , and scFv specific for L1 epitopes on the capsid of infectious HPV particles. Virology. 2007;361:435–46.
Konwarh R. Nanobodies: Prospects of Expanding the Gamut of Neutralizing Antibodies Against the Novel Coronavirus, SARS-CoV-2. Front Immunol. 2020;11(June):1–6.
Bessalah S, Jebahi S, Mejri N, Salhi I, Khorchani T, Hammadi M. Perspective on therapeutic and diagnostic potential of camel nanobodies for coronavirus disease-19 (COVID-19). 3 Biotech. 2021;11(2):1–14. Available from. https://doi.org/10.1007/s13205-021-02647-5.
Zare H, Aghamollaei H, Hosseindokht M, Heiat M, Razei A, Bakherad H. Nanobodies, the potent agents to detect and treat the Coronavirus infections: A systematic review. Mol Cell Probes. 2021;55(December 2020):101692. Available from. https://doi.org/10.1016/j.mcp.2020.101692.
Koenig P-A, Das H, Liu H, Kümmerer BM, Gohr FN, Jenster L-M, et al. Structure-guided multivalent nanobodies block SARS-CoV-2 infection and suppress mutational escape. Science (80- ). 2021;371(6530):eabe6230. [cited 2021 Mar 30]. Available from. https://doi.org/10.1126/science.abe6230.
Schoof M, Faust B, Saunders RA, Sangwan S, Rezelj V, Hoppe N, et al. An ultrapotent synthetic nanobody neutralizes SARS-CoV-2 by stabilizing inactive Spike. Science (80- ). 2021;370(6523):1473–9 [cited 2021 Mar 30]. Available from: http://science.sciencemag.org/.
Lu Q, Zhang Z, Li H, Zhong K, Zhao Q, Wang Z, et al. Development of multivalent nanobodies blocking SARS-CoV-2 infection by targeting RBD of spike protein. J Nanobiotechnology. 2021;19(1):33 [cited 2021 Mar 30]. Available from: https://jnanobiotechnology.biomedcentral.com/articles/10.1186/s12951-021-00768-w.
Huo J, Bas A, Ruza RR, Duyvesteyn HM, Mikolajek H, Malinauskas T, et al. Neutralizing nanobodies bind SARS-CoV-2 spike RBD and block interaction with ACE2. Nat Struct Mol Biol. 2020;27(9):846–54. [cited 2021 Mar 30]; Available from. https://doi.org/10.1038/s41594-020-0469-6.
Liu X, Gao F, Gou L, Chen Y, Gu Y, Ao L, et al. Neutralizing Antibodies Isolated by a site-directed Screening have Potent Protection on SARS-CoV-2 Infection. bioRxiv. 2020;2020.05.03.074914. [cited 2021 Mar 30]. Available from. https://doi.org/10.1101/2020.05.03.074914.
Yuan AQ, Zhao L, Bai L, Meng Q, Wen Z, Li Y, et al. Isolation of and characterization of neutralizing antibodies to Covid-19 from a large human naïve scFv phage display library. bioRxiv. 2020 [cited 2021 Mar 30]. p. 2020.05.19.104281. Available from. https://doi.org/10.1101/2020.05.19.104281.
Zhang J, Li H, Qi H, Wang M. Phage-derived fully human antibody scFv fragment directed against human vascular endothelial growth factor receptor 2 blocked its interaction with VEGF. Biotechnol Prog. 2012;28(4):981–9.
Behdani M, Zeinali S, Khanahmad H, Karimipour M, Asadzadeh N, Azadmanesh K, et al. Generation and characterization of a functional Nanobody against the vascular endothelial growth factor receptor-2; angiogenesis cell receptor. Mol Immunol. 2012;50(1–2):35–41. Available from:. https://doi.org/10.1016/j.molimm.2011.11.013.
Roovers RC, Vosjan MJWD, Laeremans T, El Khoulati R, De Bruin RCG, Ferguson KM, et al. A biparatopic anti-EGFR nanobody efficiently inhibits solid tumour growth. Int J Cancer. 2011;129(8):2013–24.
Camacho-Villegas T, Mata-Gonzalez T, Paniagua-Solis J, Sanchez E, Licea A. Human TNF cytokine neutralization with a vNAR from Heterodontus francisci shark. MAbs. 2013;5(1):80–5.
Van Roy M, Ververken C, Beirnaert E, Hoefman S, Kolkman J, Vierboom M, et al. The preclinical pharmacology of the high affinity anti-IL-6R Nanobody® ALX-0061 supports its clinical development in rheumatoid arthritis. Arthritis Res Ther. 2015;17(1) Available from:. https://doi.org/10.1186/s13075-015-0651-0.
Krebs B, Griffin H, Winter G, Rose-John S. Recombinant human single chain Fv antibodies recognizing human interleukin-6: Specific targeting of cytokine-secreting cells. J Biol Chem. 1998;273(5):2858–65. Available from:. https://doi.org/10.1074/jbc.273.5.2858.
Mackenzie CR, Cambillau C, Rommelaere H. Neutralization of Human Interleukin 23 by Multivalent Nanobodies Explained by the Structure of Cytokine–Nanobody Complex. Front Immunol. 2017;8(August):1–10.
Spangler JB, Moraga I, Jude KM, Savvides CS, Garcia KC. A strategy for the selection of monovalent antibodies that span protein dimer interfaces. J Biol Chem. 2019;294(38):13876–86. Available from. https://doi.org/10.1074/jbc.RA119.009213.
Lu Z, Kamat K, Johnson BP, Yin CC, Scholler N, Abbott KL. Generation of a Fully Human scFv that binds Tumor-Specific Glycoforms. Sci Rep. 2019;9(1):1–11.
Jank L, Pinto-Espinoza C, Duan Y, Koch-Nolte F, Magnus T, Rissiek B. Current Approaches and Future Perspectives for Nanobodies in Stroke Diagnostic and Therapy. Antibodies. 2019;8(1):5.
Araste F, Ebrahimizadeh W, Rasooli I, Rajabibazl M, Mousavi Gargari SL. A novel VHH nanobody against the active site (the CA domain) of tumor-associated, carbonic anhydrase isoform IX and its usefulness for cancer diagnosis. Biotechnol Lett. 2014;36(1):21–8.
Hussack G, Ryan S, Van Faassen H, Rossotti M, Roger MacKenzie C, Tanha J. Neutralization of clostridium difficile toxin b with vHH-Fc fusions targeting the delivery and CROPs domains. PLoS One. 2018;13(12):1–17.
Akbari B, Farajnia S, Ahdi Khosroshahi S, Safari F, Yousefi M, Dariushnejad H, et al. Immunotoxins in cancer therapy: Review and update. Int Rev Immunol. 2017;36(4):207–19. Available from:. https://doi.org/10.1080/08830185.2017.1284211.
Cortez-Retamozo V, Backmann N, Senter PD, Wernery U, De Baetselier P, Muyldermans S, et al. Efficient Cancer Therapy with a Nanobody-Based Conjugate. Cancer Res. 2004;64(8):2853–7 [cited 2021 Apr 28]. Available from: https://pubmed.ncbi.nlm.nih.gov/15087403/.
Moricoli D, Muller WA, Carbonella DC, Balducci MC, Dominici S, Watson R, et al. Blocking monocyte transmigration in in vitro system by a human antibody scFv anti-CD99. Efficient large scale purification from periplasmic inclusion bodies in E. coli expression system. J Immunol Methods. 2014;408:35–45 [cited 2021 Apr 28]. Available from: /pmc/articles/PMC4427036/.
Behdani M, Zeinali S, Karimipour M, Khanahmad H, Schoonooghe S, Aslemarz A, et al. Development of VEGFR2-specific Nanobody Pseudomonas exotoxin A conjugated to provide efficient inhibition of tumor cell growth. N Biotechnol. 2013;30(2):205–9 [cited 2021 Apr 28]. Available from: https://pubmed.ncbi.nlm.nih.gov/23031816/.
Wiersma VR, De Bruyn M, Shi C, Gooden MJM, Wouters MCA, Samplonius DF, et al. C-type lectin-like molecule-1 (CLL1)-targeted TRAIL augments the tumoricidal activity of granulocytes and potentiates therapeutic antibody-dependent cell-mediated cytotoxicity. MAbs. 2015;7(2):321–30 [cited 2021 Apr 28]. Available from: https://pubmed.ncbi.nlm.nih.gov/25760768/.
Hartung F, Krüwel T, Shi X, Pfizenmaier K, Kontermann R, Chames P, et al. A Novel Anti-Kv10.1 Nanobody Fused to Single-Chain TRAIL Enhances Apoptosis Induction in Cancer Cells. Front Pharmacol. 2020;11 [cited 2021 Apr 28]. Available from: https://www.readcube.com/articles/10.3389%2Ffphar.2020.00686.
Van De Water JAJM, Bagci-Onder T, Agarwal AS, Wakimoto H, Roovers RC, Zhu Y, et al. Therapeutic stem cells expressing variants of EGFR-specific nanobodies have antitumor effects. Proc Natl Acad Sci U S A. 2012;109(41):16642–7 [cited 2021 Apr 28]. Available from: https://pubmed.ncbi.nlm.nih.gov/23012408/.
Stahnke B, Thepen T, Stöcker M, Rosinke R, Jost E, Fischer R, et al. Granzyme B-H22(scFv), a human immunotoxin targeting CD64 in acute myeloid leukemia of monocytic subtypes. Mol Cancer Ther. 2008;7(9):2924–32 [cited 2021 Apr 28]. Available from: https://pubmed.ncbi.nlm.nih.gov/18790773/.
Spitaleri G, Berardi R, Pierantoni C, De Pas T, Noberasco C, Libbra C, et al. Phase I/II study of the tumour-targeting human monoclonal antibody-cytokine fusion protein L19-TNF in patients with advanced solid tumours. J Cancer Res Clin Oncol. 2013;139(3):447–55 [cited 2021 Apr 28]. Available from: www.ema.europa.eu/docs/en_GB.
Liu Y, Wang Y, Xing J, Li Y, Liu J, Wang Z. A novel multifunctional anti-CEA-IL15 molecule displays potent antitumor activities. Drug Des Devel Ther. 2018;12:2645–54 [cited 2021 Apr 28]. Available from: /pmc/articles/PMC6120566/.
Dougan M, Ingram JR, Jeong HJ, Mosaheb MM, Bruck PT, Ali L, et al. Targeting cytokine therapy to the pancreatic tumor microenvironment using PD-L1–specific VHHs. Cancer Immunol Res. 2018;6(4):389–401 Available from: https://pubmed.ncbi.nlm.nih.gov/29459478/.
Pasche N, Wulhfard S, Pretto F, Carugati E, Neri D. The antibody-based delivery of interleukin-12 to the tumor neovasculature eradicates murine models of cancer in combination with paclitaxel. Clin Cancer Res. 2012;18(15):4092–103 [cited 2021 Apr 28]. Available from: https://pubmed.ncbi.nlm.nih.gov/22693354/.
Neri D, Sondel PM. Immunocytokines for cancer treatment: Past, present and future. Curr Opin Immunol. 2016;40:96–102. Available from:. https://doi.org/10.1016/j.coi.2016.03.006.
Lu Y, Liu L, Wang Y, Li F, Zhang J, Ye M, et al. SiRNA delivered by EGFR-specific scFv sensitizes EGFR-TKI-resistant human lung cancer cells. Biomaterials. 2016;76:196–207 [cited 2021 Apr 29]. Available from: https://pubmed.ncbi.nlm.nih.gov/26524539/.
Jiang K, Li J, Yin J, Ma Q, Yan B, Zhang X, et al. Targeted delivery of CXCR4-siRNA by scFv for HER2+ breast cancer therapy. Biomaterials. 2015;59:77–87 [cited 2021 Apr 28]. Available from: https://pubmed.ncbi.nlm.nih.gov/25956853/.
Zavoiura O, Brunner B, Casteels P, Zimmermann L, Ozog M, Boutton C, et al. Nanobody-siRNA Conjugates for Targeted Delivery of siRNA to Cancer Cells. Mol Pharm. 2021; [cited 2021 Apr 25]; Available from: https://pubs.acs.org/doi/pdf/10.1021/acs.molpharmaceut.0c01001.
Huehls AM, Coupet TA, Sentman CL. Bispecific T-cell engagers for cancer immunotherapy. Immunol Cell Biol. 2015;93(3):290–6 [cited 2021 Apr 29]. Available from: https://onlinelibrary.wiley.com/doi/10.1038/icb.2014.93.
Harwood SL, Alvarez-Cienfuegos A, Nuñez-Prado N, Compte M, Hernández-Pérez S, Merino N, et al. ATTACK, a novel bispecific T cell-recruiting antibody with trivalent EGFR binding and monovalent CD3 binding for cancer immunotherapy. Oncoimmunology. 2018;7(1) [cited 2021 Apr 29]. Available from: https://pubmed.ncbi.nlm.nih.gov/29296540/.
Schmohl JU, Gleason MK, Dougherty PR, Miller JS, Vallera DA. Heterodimeric Bispecific Single Chain Variable Fragments (scFv) Killer Engagers (BiKEs) Enhance NK-cell Activity Against CD133+ Colorectal Cancer Cells. Target Oncol. 2016;11(3):353–61 [cited 2021 Apr 29]. Available from: /pmc/articles/PMC4873478/.
Schmohl JU, Felices M, Taras E, Miller JS, Vallera DA. Enhanced ADCC and NK Cell Activation of an Anticarcinoma Bispecific Antibody by Genetic Insertion of a Modified IL-15 Cross-linker. YMTHE. 2016;24:1312–22 [cited 2021 Apr 29]. Available from: www.moleculartherapy.org.
Li Y, Zhou C, Li J, Liu J, Lin L, Li L, et al. Single domain based bispecific antibody, Muc1-Bi-1, and its humanized form, Muc1-Bi-2, induce potent cancer cell killing in muc1 positive tumor cells. PLoS One. 2018;13(1) [cited 2021 Apr 29]. Available from: https://pubmed.ncbi.nlm.nih.gov/29357376/.
de Bruin RCG, Veluchamy JP, Lougheed SM, Schneiders FL, Lopez-Lastra S, Lameris R, et al. A bispecific nanobody approach to leverage the potent and widely applicable tumor cytolytic capacity of Vγ9Vδ2-T cells. Oncoimmunology. 2018;7(1) [cited 2021 Apr 29]. Available from: /pmc/articles/PMC5739573/.
Knight JB, Halperin SA, West KA, Lee SF. Expression of a functional single-chain variable-fragment antibody against complement receptor 1 in Streptococcus gordonii. Clin Vaccine Immunol. 2008;15(6):925–31 [cited 2021 Apr 29]. Available from: https://pubmed.ncbi.nlm.nih.gov/18385459/.
Duarte JN, Cragnolini JJ, Swee LK, Bilate AM, Bader J, Ingram JR, et al. Generation of Immunity against Pathogens via Single-Domain Antibody–Antigen Constructs. J Immunol. 2016;197(12):4838–47 [cited 2021 Apr 29]. Available from: http://www.jimmunol.org/content/197/12/4838.
Hu Y, Liu C, Muyldermans S. Nanobody-based delivery systems for diagnosis and targeted tumor therapy. Front Immunol. 2017;8:1442.
Lu Y, Liang K, Li X, Fan Z. Responses of cancer cells with wild-type or tyrosine kinase domain-mutated epidermal growth factor receptor (EGFR) to EGFR-targeted therapy are linked to downregulation of hypoxia-inducible factor-1α. Mol Cancer. 2007;6 [cited 2021 May 1]. Available from: https://pubmed.ncbi.nlm.nih.gov/17931419/.
Yin W, Yu X, Kang X, Zhao Y, Zhao P, Jin H, et al. Remodeling Tumor-Associated Macrophages and Neovascularization Overcomes EGFRT790M-Associated Drug Resistance by PD-L1 Nanobody-Mediated Codelivery. Small. 2018;14(47) [cited 2021 May 1]. Available from: https://pubmed.ncbi.nlm.nih.gov/30307695/.
Oliveira S, Schiffelers RM, van der Veeken J, van der Meel R, Vongpromek R, van Bergenen Henegouwen PMP, et al. Downregulation of EGFR by a novel multivalent nanobody-liposome platform. J Control Release. 2010;145(2):165–75 [cited 2021 May 1]. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0168365910002257.
Katakowski JA, Mukherjee G, Wilner SE, Maier KE, Harrison MT, Di Lorenzo TP, et al. Delivery of siRNAs to dendritic cells using DEC205-targeted lipid nanoparticles to inhibit immune responses. Mol Ther. 2016;24(1):146–55 [cited 2021 May 1]. Available from: https://pubmed.ncbi.nlm.nih.gov/26412590/.
Wang Y, Wang Y, Chen G, Li Y, Xu W, Gong S. Quantum-Dot-Based Theranostic Micelles Conjugated with an Anti-EGFR Nanobody for Triple-Negative Breast Cancer Therapy. ACS Appl Mater Interfaces. 2017;9(36):30297–305 [cited 2021 May 1]. Available from: https://pubmed.ncbi.nlm.nih.gov/28845963/.
Costa SA, Mozhdehi D, Dzuricky MJ, Isaacs FJ, Brustad EM, Chilkoti A. Active Targeting of Cancer Cells by Nanobody Decorated Polypeptide Micelle with Bio-orthogonally Conjugated Drug. Nano Lett. 2019;19(1):247–54. [cited 2021 May 1]. Available from:. https://doi.org/10.1021/acs.nanolett.8b03837.
Altintas I, Heukers R, Van Der Meel R, Lacombe M, Amidi M, Van Bergen En Henegouwen PMP, et al. Nanobody-albumin nanoparticles (NANAPs) for the delivery of a multikinase inhibitor 17864 to EGFR overexpressing tumor cells. J Control Release. 2013;165(2):110–8 [cited 2021 May 1]. Available from: https://pubmed.ncbi.nlm.nih.gov/23159529/.
Heukers R, Altintas I, Raghoenath S, De Zan E, Pepermans R, Roovers RC, et al. Targeting hepatocyte growth factor receptor (Met) positive tumor cells using internalizing nanobody-decorated albumin nanoparticles. Biomaterials. 2014;35(1):601–10 [cited 2021 May 1]. Available from: https://pubmed.ncbi.nlm.nih.gov/24139763/.
Dou S, Yang XZ, Xiong MH, Sun CY, Yao YD, Zhu YH, et al. ScFv-Decorated PEG-PLA-Based Nanoparticles for Enhanced siRNA Delivery to Her2+ Breast Cancer. Adv Healthc Mater. 2014;3(11):1792–803 [cited 2021 May 1]. Available from: https://pubmed.ncbi.nlm.nih.gov/24947820/.
Saqafi B, Rahbarizadeh F. Polyethyleneimine-polyethylene glycol copolymer targeted by anti-HER2 nanobody for specific delivery of transcriptionally targeted tBid containing construct. Artif Cells, Nanomedicine Biotechnol. 2019;47(1):501–11 [cited 2021 May 1]. Available from: https://pubmed.ncbi.nlm.nih.gov/30810413/.
Eichhoff AM, Börner K, Albrecht B, Schäfer W, Baum N, Haag F, et al. Nanobody-Enhanced Targeting of AAV Gene Therapy Vectors. Mol Ther - Methods Clin Dev. 2019;15:211–20. [cited 2021 May 1]. Available from. https://doi.org/10.1016/j.omtm.2019.09.003.
Ahani R, Roohvand F, Cohan RA, Etemadzadeh MH, Mohajel N, Behdani M, et al. Sindbis Virus-Pseudotyped Lentiviral Vectors Carrying VEGFR2-Specific Nanobody for Potential Transductional Targeting of Tumor Vasculature. Mol Biotechnol. 2016;58(11):738–47 Available from: https://pubmed.ncbi.nlm.nih.gov/27647452/.
Goyvaerts C, De Groeve K, Dingemans J, Van Lint S, Robays L, Heirman C, et al. Development of the Nanobody display technology to target lentiviral vectors to antigen-presenting cells. Gene Ther. 2012;19(12):1133–40 [cited 2021 May 1]. Available from: https://pubmed.ncbi.nlm.nih.gov/22241177/.
Dreja H, Piechaczyk M. The effects of N-terminal insertion into VSV-G of an scFv peptide. Virol J. 2006;3:69 [cited 2021 May 1]. Available from: /pmc/articles/PMC1564393/.
Frank AM, Buchholz CJ. Surface-Engineered Lentiviral Vectors for Selective Gene Transfer into Subtypes of Lymphocytes. Mol Therapy Methods Clin Dev. 2019;12:19–31.
Poulin KL, Lanthier RM, Smith AC, Christou C, Risco Quiroz M, Powell KL, et al. Retargeting of Adenovirus Vectors through Genetic Fusion of a Single-Chain or Single-Domain Antibody to Capsid Protein IX. J Virol. 2010;84(19):10074–86 [cited 2021 May 1]. Available from: /pmc/articles/PMC2937758/.
Marschall ALJ, Dübel S. Antibodies inside of a cell can change its outside: Can intrabodies provide a new therapeutic paradigm? Computational and Structural Biotechnology Journal. 2016;14:304–8 [cited 2021 May 1]. Available from: /pmc/articles/PMC4990636/.
Lo ASY, Zhu Q, Marasco WA. Intracellular antibodies (intrabodies) and their therapeutic potential. Handb Exp Pharmacol. 2008;181(181):343–73 [cited 2021 May 1]. Available from: https://pubmed.ncbi.nlm.nih.gov/18071953/.
Messer A, Butler DC. Optimizing intracellular antibodies (intrabodies/nanobodies) to treat neurodegenerative disorders. Neurobiol Dis. 2020;134(September 2019):104619. Available from. https://doi.org/10.1016/j.nbd.2019.104619.
Butler DC, McLear JA, Messer A. Engineered antibody therapies to counteract mutant huntingtin and related toxic intracellular proteins. Prog Neurobiol. 2012;97(2):190–204. Available from:. https://doi.org/10.1016/j.pneurobio.2011.11.004.
Scheller L, Fussenegger M. From synthetic biology to human therapy: engineered mammalian cells. Curr Opin Biotechnol. 2019;58:108–16 [cited 2021 Mar 15]. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0958166918302167.
Teixeira PA, Fussenegger M. Engineering mammalian cells for disease diagnosis and treatment. Curr Opin Biotechnol. 2019;55:87–94.
Lim WAWA, June CH. Review the principles of engineering immune cells to treat cancer. Cell. 2017;168(4):724–40. [cited 2017 Nov 4]. Available from. https://doi.org/10.1016/j.cell.2017.01.016.
Rahbarizadeh F, Ahmadvand D, Moghimi SM. CAR T-cell bioengineering: Single variable domain of heavy chain antibody targeted CARs. Adv Drug Deliv Rev. 2019;141:41–6. Available from. https://doi.org/10.1016/j.addr.2019.04.006.
Xie YJ, Dougan M, Jailkhani N, Ingram J, Fang T, Kummer L, et al. Nanobody-based CAR T cells that target the tumor microenvironment inhibit the growth of solid tumors in immunocompetent mice. Proc Natl Acad Sci U S A. 2019;116(33):16656. https://doi.org/10.1073/pnas.1817147116.
Khaleghi S, Ahmadvand D, Pognonec P, Rahbarizadeh F, Rasaee MJ. A caspase 8-based suicide switch induces apoptosis in nanobody-directed chimeric receptor expressing T cells. Int J Hematol. 2012;95(4):434–44.
Jamnani FR, Rahbarizadeh F, Shokrgozar MA, Mahboudi F, Ahmadvand D, Sharifzadeh Z, et al. T cells expressing VHH-directed oligoclonal chimeric HER2 antigen receptors: Towards tumor-directed oligoclonal T cell therapy. Biochim Biophys Acta - Gen Subj. 2014;1840(1):378–86.
Hajari Taheri F, Hassani M, Sharifzadeh Z, Behdani M, Arashkia A, Abolhassani M. T cell engineered with a novel nanobody-based chimeric antigen receptor against VEGFR2 as a candidate for tumor immunotherapy. IUBMB Life. 2019;71(9):1259–67 [cited 2021 Mar 15]. Available from: https://onlinelibrary.wiley.com/doi/abs/10.1002/iub.2019.
De Munter S, Van Parys A, Bral L, Ingels J, Goetgeluk G, Bonte S, et al. Rapid and Effective Generation of Nanobody Based CARs using PCR and Gibson Assembly. Int J Mol Sci. 2020;21(3):883 [cited 2021 Mar 15]. Available from: https://www.mdpi.com/1422-0067/21/3/883.
Rajabzadeh A, Ahmadvand D, Salmani MK, Rahbarizadeh F, Hamidieh AA. A VHH-based anti-MUC1 chimeric antigen receptor for specific retargeting of human primary T cells to MUC1-positive cancer cells. Cell J. 2021;22(4):502–13.
Bao C, Gao Q, Li L-L, Han L, Zhang B, Ding Y, et al. The Application of Nanobody in CAR-T Therapy. Biomolecules. 2021;11(2):238 [cited 2021 Mar 15]. Available from: https://www.mdpi.com/2218-273X/11/2/238.
De Munter S, Ingels J, Goetgeluk G, Bonte S, Pille M, Weening K, et al. Nanobody Based Dual Specific CARs. Int J Mol Sci. 2018;19(2):403 [cited 2021 Mar 15]. Available from: http://www.mdpi.com/1422-0067/19/2/403.
Morsut L, Roybal KT, Xiong X, Gordley RM, Coyle SM, Thomson M, et al. Engineering Customized Cell Sensing and Response Behaviors Using Synthetic Notch Receptors. Cell. 2016;164(4):780–91.
Scheller L, Strittmatter T, Fuchs D, Bojar D, Fussenegger M. Generalized extracellular molecule sensor platform for programming cellular behavior article. Nat Chem Biol. 2018;14(7):723–9. [cited 2021 Mar 15]. Available from. https://doi.org/10.1038/s41589-018-0046-z.
Bojar D, Scheller L, El Hamri GC, Xie M, Fussenegger M. Caffeine-inducible gene switches controlling experimental diabetes. Nat Commun. 2018;9(1):1–10 [cited 2021 Mar 15]. Available from: www.nature.com/naturecommunications.
Huang L, Muyldermans S, Saerens D. Nanobodies®: proficient tools in diagnostics. Expert Rev Mol Diagn. 2010;10(6):777–85. Available from. https://doi.org/10.1586/erm.10.62.
De Groeve K, Deschacht N, De Koninck C, Caveliers V, Lahoutte T, Devoogdt N, et al. Nanobodies as tools for in vivo imaging of specific immune cell types. J Nucl Med. 2010;51(5):782–9 [cited 2021 May 6]. Available from: https://pubmed.ncbi.nlm.nih.gov/20395332/.
Hilderbrand SA, Weissleder R. Near-infrared fluorescence: application to in vivo molecular imaging. Curr Opin Chem Biol. 2010;14:71–9 [cited 2021 May 6]. Available from: https://pubmed.ncbi.nlm.nih.gov/19879798/.
Capala J, Bouchelouche K. Molecular imaging of HER2-positive breast cancer: A step toward an individualized “image and treat” strategy. Curr Opin Oncol. 2010;22:559–66 [cited 2021 May 6]. Available from: https://pubmed.ncbi.nlm.nih.gov/20842031/.
Chakravarty R, Goel S, Cai W. Nanobody: The “magic bullet” for molecular imaging? Theranostics. 2014;4(4):386–98.
Fatehi D, Baral TN, Abulrob A. In vivo imaging of brain cancer using epidermal growth factor single domain antibody bioconjugated to near-infrared quantum dots. J Nanosci Nanotechnol. 2014;14(7):5355–62 [cited 2021 May 6]. Available from: https://pubmed.ncbi.nlm.nih.gov/24758031/.
Deckers N, Saerens D, Kanobana K, Conrath K, Victor B, Wernery U, et al. Nanobodies, a promising tool for species-specific diagnosis of Taenia solium cysticercosis. Int J Parasitol. 2009;39(5):625–33 [cited 2021 May 7]. Available from: https://pubmed.ncbi.nlm.nih.gov/19041315/.
Saerens D, Stijlemans B, Baral TN, Nguyen Thi GT, Wernery U, Magez S, et al. Parallel selection of multiple anti-infectome Nanobodies without access to purified antigens. J Immunol Methods. 2008;329(1–2):138–50 [cited 2021 May 7]. Available from: https://pubmed.ncbi.nlm.nih.gov/17996887/.
Abeijon C, Dilo J, Tremblay JM, Viana AG, Bueno LL, Carvalho SFG, et al. Use of VHH antibodies for the development of antigen detection test for visceral leishmaniasis. Parasite Immunol. 2018;40(11) [cited 2021 May 7]. Available from: /pmc/articles/PMC6220836/.
Melli LJ, Zylberman V, Hiriart Y, Lauche CE, Baschkier A, Pardo R, et al. Development and evaluation of a novel VHH-based immunocapture assay for high-sensitivity detection of Shiga toxin type 2 (Stx2) in stool samples. J Clin Microbiol. 2020;58(3) [cited 2021 May 7]. Available from: http://jcm.asm.org/.
Pinto Torres JE, Goossens J, Ding J, Li Z, Lu S, Vertommen D, et al. Development of a Nanobody-based lateral flow assay to detect active Trypanosoma congolense infections. Sci Rep. 2018;8(1):1–15 [cited 2021 May 7]. Available from: www.nature.com/scientificreports/.
Wting Z, Tting L, Wu M, Xchen C, Han L, Zzhong S, et al. Development of a nanobody-based immunoassay for the sensitive detection of fibrinogen-like protein 1. Acta Pharmacol Sin. 2021:1–9. [cited 2021 May 7]. Available from. https://doi.org/10.1038/s41401-020-00574-4.
Ji Y, Li X, Lu Y, Guo P, Zhang G, Wang Y, et al. Nanobodies Based on a Sandwich Immunoassay for the Detection of Staphylococcal Enterotoxin B Free from Interference by Protein A. J Agric Food Chem. 2020;68(21):5959–68 [cited 2021 May 7]. Available from: https://pubs.acs.org/doi/abs/10.1021/acs.jafc.0c00422.
Doerflinger SY, Tabatabai J, Schnitzler P, Farah C, Rameil S, Sander P, et al. Development of a Nanobody-Based Lateral Flow Immunoassay for Detection of Human Norovirus. mSphere. 2016;1(5):219–35 [cited 2021 May 7]. Available from: http://msphere.asm.org/.
Chen J, He QH, Xu Y, Fu JH, Li YP, Tu Z, et al. Nanobody medicated immunoassay for ultrasensitive detection of cancer biomarker alpha-fetoprotein. Talanta. 2016;147:523–30 [cited 2021 May 7]. Available from: https://pubmed.ncbi.nlm.nih.gov/26592642/.
Shi FS, Zhang L, Xia WQ, Liu J, Zhang HC, Wang JP. Production and evolution of a ScFv antibody for immunoassay of residual phenothiazine drugs in meat based on computational simulation. Anal Methods. 2017;9(30):4455–63 [cited 2021 May 7]. Available from: https://pubs.rsc.org/en/content/articlehtml/2017/ay/c7ay01103b.
Li D, Cui Y, Morisseau C, Gee SJ, Bever CS, Liu X, et al. Nanobody Based Immunoassay for Human Soluble Epoxide Hydrolase Detection Using Polymeric Horseradish Peroxidase (PolyHRP) for Signal Enhancement: The Rediscovery of PolyHRP? Anal Chem. 2017;89(11):6248–56 [cited 2021 May 7]. Available from: /pmc/articles/PMC5611449/.
Yang H, Zhong Y, Wang J, Zhang Q, Li X, Ling S, et al. Screening of a ScFv antibody with high affinity for application in human IFN-γ immunoassay. Front Microbiol. 2018;9(MAR):261 [cited 2021 May 7]. Available from: www.frontiersin.org.
Saerens D, Huang L, Bonroy K, Muyldermans S. Antibody fragments as probe in biosensor development. Sensors. 2008;8(8):4669–86.
Kaiser PD, Maier J, Traenkle B, Emele F, Rothbauer U. Recent progress in generating intracellular functional antibody fragments to target and trace cellular components in living cells. Biochim Biophys Acta. 2014;1844(11):1933–42 Available from: https://www.sciencedirect.com/science/article/pii/S1570963914001113.
Williams AF, Barclay AN. The immunoglobulin superfamily--domains for cell surface recognition. Annu Rev Immunol. 1988;6:381–405.
Glockshuber R, Schmidt T, Plueckthun A. The disulfide bonds in antibody variable domains: effects on stability, folding in vitro, and functional expression in Escherichia coli. Biochemistry. 1992;31(5):1270–9. Available from. https://doi.org/10.1021/bi00120a002.
Proba K, Honegger A, Plückthun A. A natural antibody missing a cysteine in VH: consequences for thermodynamic stability and folding11Edited by I. A Wilson Mol Biol. 1997;265(2):161–72 Available from: https://www.sciencedirect.com/science/article/pii/S0022283696907263.
Beghein E, Gettemans J. Nanobody Technology: A Versatile Toolkit for Microscopic Imaging, Protein–Protein Interaction Analysis, and Protein Function Exploration. Front Immunol. 2017;8:771 Available from: https://www.frontiersin.org/article/10.3389/fimmu.2017.00771.
Keller B-M, Maier J, Secker K-A, Egetemaier S-M, Parfyonova Y, Rothbauer U, et al. Chromobodies to Quantify Changes of Endogenous Protein Concentration in Living Cells. Mol Cell Proteomics. 2018;17(12):2518–33.
de Bruin RCG, Lougheed SM, van der Kruk L, Stam AG, Hooijberg E, Roovers RC, et al. Highly specific and potently activating Vγ9Vδ2-T cell specific nanobodies for diagnostic and therapeutic applications. Clin Immunol. 2016;169:128–38.
Peyrassol X, Laeremans T, Gouwy M, Lahura V, Debulpaep M, Van Damme J, et al. Development by Genetic Immunization of Monovalent Antibodies (Nanobodies) Behaving as Antagonists of the Human ChemR23 Receptor. J Immunol. 2016;196(6):2893–901.
Chamma I, Letellier M, Butler C, Tessier B, Lim K-H, Gauthereau I, et al. Mapping the dynamics and nanoscale organization of synaptic adhesion proteins using monomeric streptavidin. Nat Commun. 2016;7(1):10773.
Guignet EG, Hovius R, Vogel H. Reversible site-selective labeling of membrane proteins in live cells. Nat Biotechnol. 2004;22(4):440–4.
Ezzine A, el Adab S, Bouhaouala-Zahar B, Hmila I, Baciou L, Marzouki MN. Efficient expression of the anti-AahI’ scorpion toxin nanobody under a new functional form in a Pichia pastoris system. Biotechnol Appl Biochem. 2012;59(1):15–21 Available from: https://iubmb.onlinelibrary.wiley.com/doi/abs/10.1002/bab.67.
Pucca MB, Bertolini TB, Barbosa JE, Galina SVR, Porto GS. Therapeutic monoclonal antibodies: scFv patents as a marker of a new class of potential biopharmaceuticals. Brazilian J Pharm Sci [Internet]. 2011;47:31–8 Available from: http://www.scielo.br/scielo.php?script=sci_arttext&pid=S1984-82502011000100005&nrm=iso.
Traenkle B, Rothbauer U. Under the Microscope: Single-Domain Antibodies for Live-Cell Imaging and Super-Resolution Microscopy. Front Immunol. 2017;8:1030 Available from: https://www.frontiersin.org/article/10.3389/fimmu.2017.01030.
Ries J, Kaplan C, Platonova E, Eghlidi H, Ewers H. A simple, versatile method for GFP-based super-resolution microscopy via nanobodies. Nat Methods. 2012;9(6):582–4. Available from. https://doi.org/10.1038/nmeth.1991.
Sheetz T, Mills J, Tessari A, Pawlikowski M, Braddom AE, Posid T, et al. NCL Inhibition Exerts Antineoplastic Effects against Prostate Cancer Cells by Modulating Oncogenic MicroRNAs. Cancers (Basel). 2020;12(7) Available from: https://www.mdpi.com/2072-6694/12/7/1861.
Embrione V, Scarpati M, Uzzel ZD, Ahmad R, de FA S, Sheetz TJ, et al. Abstract 476: A human scFv as a tool to understand the biogenesis of a subset of oncogenic microRNAs. Cancer Res. 2018;78(13 Supplement):476 Available from: https://cancerres.aacrjournals.org/content/78/13_Supplement/476.
Harmansa S, Affolter M. Protein binders and their applications in developmental biology. Development. 2018;145(2):dev148874.
Gettemans J, de Dobbelaer B. Transforming nanobodies into high-precision tools for protein function analysis. Am J Physiol Cell Physiolo. 2021;320:C195–215.
Romer T, Leonhardt H, Rothbauer U. Engineering antibodies and proteins for molecular in vivo imaging. Curr Opin Biotechnol. 2011;22(6):882–7.
Jobling SA, Jarman C, Teh M-M, Holmberg N, Blake C, Verhoeyen ME. Immunomodulation of enzyme function in plants by single-domain antibody fragments. Nat Biotechnol. 2003;21(1):77–80.
De Clercq S, Zwaenepoel O, Martens E, Vandekerckhove J, Guillabert A, Gettemans J. Nanobody-induced perturbation of LFA-1/L-plastin phosphorylation impairs MTOC docking, immune synapse formation and T cell activation. Cell Mol Life Sci. 2013;70(5):909–22.
Tanaka T, Sewell H, Waters S, Phillips SEV, Rabbitts TH. Single domain intracellular antibodies from diverse libraries: emphasizing dual functions of LMO2 protein interactions using a single VH domain. J Biol Chem. 2011;286(5):3707–16.
Caussinus E, Kanca O, Affolter M. Fluorescent fusion protein knockout mediated by anti-GFP nanobody. Nat Struct Mol Biol. 2012;19(1):117–21. Available from. https://doi.org/10.1038/nsmb.2180.
Joshi SN, Butler DC, Messer A. Fusion to a highly charged proteasomal retargeting sequence increases soluble cytoplasmic expression and efficacy of diverse anti-synuclein intrabodies. MAbs. 2012;4(6):686–93.
Wang Y, Fan Z, Shao L, Kong X, Hou X, Tian D, et al. Nanobody-derived nanobiotechnology tool kits for diverse biomedical and biotechnology applications. Int J Nanomedicine. 2016;11:3287–303 Available from: https://pubmed.ncbi.nlm.nih.gov/27499623.
Rothbauer U, Zolghadr K, Muyldermans S, Schepers A, Cardoso MC, Leonhardt H. A versatile nanotrap for biochemical and functional studies with fluorescent fusion proteins. Mol Cell Proteomics. 2008;7(2):282–9.
Harmansa S, Hamaratoglu F, Affolter M, Caussinus E. Dpp spreading is required for medial but not for lateral wing disc growth. Nature. 2015;527(7578):317–22.
Harmansa S, Alborelli I, Bieli D, Caussinus E, Affolter M. A nanobody-based toolset to investigate the role of protein localization and dispersal in Drosophila. Elife. 2017;6:e22549.
Klooster R, Eman MR, le Duc Q, Verheesen P, Verrips CT, Roovers RC, et al. Selection and characterization of KDEL-specific VHH antibody fragments and their application in the study of ER resident protein expression. J Immunol Methods. 2009;342(1–2):1–12.
Tang JCY, Szikra T, Kozorovitskiy Y, Teixiera M, Sabatini BL, Roska B, et al. A nanobody-based system using fluorescent proteins as scaffolds for cell-specific gene manipulation. Cell. 2013;154(4):928–39.
Koide S. Engineering of recombinant crystallization chaperones. Curr Opin Struct Biol. 2009;19:449–57 [cited 2021 May 19]. Available from: /pmc/articles/PMC2736338/.
Kromann-Hansen T, Louise Lange E, Peter Sørensen H, Hassanzadeh-Ghassabeh G, Huang M, Jensen JK, et al. Discovery of a novel conformational equilibrium in urokinase-type plasminogen activator. Sci Rep. 2017;7(1):3385.
Rasmussen SGF, Devree BT, Zou Y, Kruse AC, Chung KY, Kobilka TS, et al. Crystal structure of the β 2 adrenergic receptor-Gs protein complex. Nature. 2011;477(7366):549–57.
Dumoulin M, Last AM, Desmyter A, Decanniere K, Canet D, Larsson G, et al. A camelid antibody fragment inhibits the formation of amyloid fibrils by human lysozyme. Nature. 2003;424(6950):783–8. Available from. https://doi.org/10.1038/nature01870.
Domanska K, Vanderhaegen S, Srinivasan V, Pardon E, Dupeux F, Marquez JA, et al. Atomic structure of a nanobody-trapped domain-swapped dimer of an amyloidogenic beta2-microglobulin variant. Proc Natl Acad Sci U S A. 2011;108(4):1314–9.
De Meyer T, Muyldermans S, Depicker A. Nanobody-based products as research and diagnostic tools. Trends Biotechnol. 2014;32:263–70.
De Genst EJ, Guilliams T, Wellens J, O’Day EM, Waudby CA, Meehan S, et al. Structure and properties of a complex of α-synuclein and a single-domain camelid antibody. J Mol Biol. 2010;402(2):326–43.
Muyldermans S. Applications of Nanobodies. Annu Rev Anim Biosci. 2021;9(1):401–21. Available from. https://doi.org/10.1146/annurev-animal-021419-083831.
Klooster R, Maassen BTH, Stam JC, Hermans PW, Ten Haaft MR, Detmers FJM, et al. Improved anti-IgG and HSA affinity ligands: clinical application of VHH antibody technology. J Immunol Methods. 2007;324(1–2):1–12.
Nguyen-Duc T, Peeters E, Muyldermans S, Charlier D, Hassanzadeh-Ghassabeh G. Nanobody(R)-based chromatin immunoprecipitation/micro-array analysis for genome-wide identification of transcription factor DNA binding sites. Nucleic Acids Res. 2013;41(5):e59.
Acknowledgements
Not applicable.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
Yasaman Asaadi: Conceptualization, Investigation, Writing-original draft, Writing-review & editing, Validation, Supervision. Fatemeh Fazlollahi: Writing-original draft, Writing-review & editing, Validation Sara Janani: Writing original draft, Writing-review & editing, Validation. Fatemeh Rahbarizadeh: Writing-review & editing, Validation, Supervision. The author(s) read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Asaadi, Y., Jouneghani, F.F., Janani, S. et al. A comprehensive comparison between camelid nanobodies and single chain variable fragments. Biomark Res 9, 87 (2021). https://doi.org/10.1186/s40364-021-00332-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40364-021-00332-6