
Abstract
Background
The environmental role of carbapenemase-producing Enterobacteriaceae (CPE) acquisition and infection in human disease has been described but not thoroughly investigated. We aimed to assess the occurrence of CPE in nearshore aquatic bodies.
Methods
Enterobacteriaceae were cultured from coastal and estuary water near Netanya, Israel in June and July of 2018. Bacteria were identified by VITEK2® and their antimicrobial susceptibility was tested according to the CLSI guidelines. Enterobacteriaceae genomes were sequenced to elucidate their resistome and carbapenemase types.
Results
Among other clinically relevant bacteria, four CPE (three Enterobacter spp and one Escherichia coli isolate) were isolated from two river estuaries (Poleg and Alexander Rivers) and coastal water at a popular recreational beach (Beit Yanai). Molecular analysis and genome sequencing revealed the persistent presence of rare beta-lactamase resistance genes, including blaIMI-2 and a previously unknown blaIMI-20 allele, which were not found among the local epidemiological strains. Genome comparisons revealed the high identity of riverine and marine CPE that were cultivated one month apart.
Conclusions
We show that CPE contamination was widespread in nearshore marine and riverine habitats. The high genome-level similarity of riverine and marine CPEs, isolated one month apart, hints at the common source of infection. We discuss the clinical implications of these findings and stress the urgent need to assess the role of the aquatic environment in CPE epidemiology.
Similar content being viewed by others


Background
Carbapenem-resistant Enterobacteriaceae (CRE), in particular, carbapenemase- and extended-spectrum β-lactamase-producing Enterobacteriaceae (CPE and ESBL-PE) endanger global health, as they have spread worldwide during the last two decades. The risk factors for the acquisition of these multidrug-resistant organisms (MDROs) are usually associated with healthcare [1] and overseas travel [2]. Potential modes of environmental transmission of these bacteria to humans outside of healthcare facilities following exposure to wildlife, livestock and pet animals have been reported [3]. The environmental routes of CRE infection may play a role in causing human disease and spreading it globally, however, they have not been thoroughly investigated yet. In particular, little is known about the epidemiology of CRE in the aquatic environment.
Recent studies show that CRE may contaminate aquatic environments such as marine surface water [4,5,6,7], rivers [5, 8,9,10,11,12,13,14,15,16,17], estuaries [18] and polluted drinking water [19]. Riverine CRE often harbor several carbapenemase types [8, 10, 11, 15]. A case of blaIMI-2 CP-Enterobacter asburiae bacteremia following a river near-drowning accident was described, and the infecting pathogen was isolated from the river one month later [16]. In two studies that reported the isolation of CPE from seawater, Enterobacter spp. was prominent and comprised of 69–76% of the Enterobacteriaceae isolated [6, 7]. Assessing the link between the clinical and aquatic epidemiology of CRE is often challenging. While some studies showed that clinical strains may be found in aquatic bodies [7, 9, 16], none such link was demonstrated in others [17, 20].
CRE may reach aquatic bodies as a result of organic contamination from multiple sources [21], including hospital effluents [22,23,24], wastewater treatment plants (WWTPs) [24,25,26], discharge of livestock farms and agriculture [3, 27], seepage water [28] as well as others [21]. Once mixed with the aquatic body, these effluents may introduce not only foreign MDROs, but also high doses of antibiotics, which likely trigger resistance propagation [29]. In Israel, the main sources of aquatic contamination comprise of regional councils that are not connected to WWTPs, controlled discharges from fish farms and WWTPs [30] and untreated sewage from the Palestinian Authority [31]. Israeli governmental programs that monitor the quality of coastal seawater and rivers report coliform concentrations, but not the presence of MDROs. MDRO infection, however, poses a real threat to those engaged in recreational water activities.
A young patient who suffered a near-drowning experience in one of Netanya's beaches was admitted to the Laniado medical center, Netanya, Israel, in June 2014. Bacterial screening upon his admission to the intensive care unit found he was a carrier of two CPE species: Enterobacter cloacae and Klebsiella oxytoca, both carrying blaKPC. We assumed that seawater ingestion and aspiration led to this infection, as no other risk factors could explain the high rates of CPE colonization. We hypothesized that these CPEs originated from rivers supplying water to the shoreline in the vicinity of Netanya. To test this hypothesis, we aimed to identify CPE in coastal waters and two river estuaries in the Sharon district.
Material and methods
Sampling sites
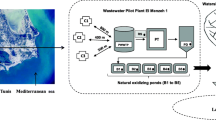
Two river estuaries were sampled in the vicinity of the Laniado hospital: Poleg (32°16′11.7"N 34°49′55.7"E) ~ 13 Km south to Laniado hospital and the Alexander River (32°23′46.7"N 34°51′57.0"E) ~ 5 Km north to the hospital. Two popular public recreational beaches along the coastline were also sampled: Beit Yanai (32°23′15.7"N 34°51′48.3"E) ~ 4.6 Km north of the hospital and 1 Km south of the Alexander river estuary; and Sironit (32°19′51.4"N 34°50′53.8"E) 1.7 Km south of the hospital and 6.5 Km north of the Poleg River estuary (Fig. 1).

Map of the coastline near Netanya and areas of water samplings (red asterisks)
Sampling and bacterial isolation
On two occasions (June, July 2018), water samples were collected with sterile-glass containers (pre-combusted at 500 °C) and delivered to the laboratory at room temperature. 200–1300 ml of freshwater or seawater was filtered, using a vacuum pump, through a 0.2 µm sterile cellulose acetate membrane (Sartorius Stedim®). Distinct volumes of water were filtered at each site (Table 1), due to differences in turbidity and membrane clogging. Control membranes were obtained by filtering room air for the same duration. The membranes were soaked in an enrichment medium (thioglycolate broth (Novamed®)), that served as the source of cultures: first after overnight incubation at room temperature, and after 24 h of incubation at 37 °C. The media were cultured on TSBA + Blood/Chocolate agar, MacConkey agar, CNA agar, Chromagar Orientation, Chromagar MSupercarba and Streptococci Select medium plates (all from HyLabs®). Colonies were picked, isolated and then identified using VITEK2® (bioMérieux). Antimicrobial susceptibility testing was performed by VITEK2® (bioMérieux) and Etest (bioMérieux) on Mueller–Hinton agar according to the Clinical & Laboratory Standards Institute (CLSI) guidelines. Enterobacteriaceae growing on CRE plates were subjected to the modified Hodge test or CARBA NP hydrolysis. Following species isolation and identification, the carbapenemase type was identified using Cepheid® GeneXpert Carba-R for blaKPC, blaNDM, blaVIM, blaIMP, blaOXA-48 genes. The blaIMI was identified using the polymerase chain reaction (PCR) [32].
We repeated the sampling from the same sites 3 weeks later. During the second sampling session, we mainly attempted to identify CREs, and used 60 ml of sterile media as a negative control.
Genome sequencing, assembly and bioinformatics
DNA was extracted from three Enterobacter isolates: nEC133, nEC134 and nEC135 using the Presto Mini gDNA Bacteria kit (GeneAid). DNA libraries were prepared using the Illumina Nextera XT kit (Illumina). Sequencing was performed using the Illumina MiSeq platform with 2 × 250 bp paired-end reads aiming at > 100 × coverage, MiSeq Reagent Kit v2.
Bioinformatic analyses
Sequencing was performed using the Illumina MiSeq platform with 2 × 250 bp paired-end reads aiming at > 100 × coverage, MiSeq Reagent Kit v2. Genomes were de-novo assembled with SPAdes V3.12 [33] and their quality was estimated with checkM [34]. The whole-genome multilocus sequence typing (wgMLST [35]) for Enterobacter cloacae complex was performed using the BioNumerics version 7.6.3 cloud-based calculation engine. wgMLST alleles were determined using assembly-free and assembly-based approaches. The resistome was identified using a standalone Resistance Gene Identifier (RGI) application with the Comprehensive Antibiotic Resistance Database (CARD [36]). Plasmid sequences were predicted with plasFlow [37] because we were unable to assemble complete plasmid sequences with PlasmidSPAdes [38]. Average nucleotide identity (ANI) was calculated as orthoANIu [39] using EzBioCloud [40]. The maximum likelihood tree of BlaIMI amino acid sequences was constructed with MEGA7 [41], based on the LG model [42].
Genomic sequences are available under the NCBI BioProject accession number PRJNA578038. The blaIMI-20 sequence was submitted to GeneBank with the accession number MN619794.
Results
Water-borne bacterial isolates
Enterobacteriaceae, non-fermenter bacilli and Gram-positive cocci were isolated from the coastal aquatic environment (Table 1). Four strains of CPE were found in the two sampling sessions: three isolates of CP-Enterobacter spp and CP-Escherichia coli blaOXA-48. The three Enterobacter isolates, which were isolated from seawater and freshwater, had remarkably similar antibiogram phenotype, different from that of E. coli blaOXA-48 (Table 2).
Resistome of aquatic Enterobacter revealed by genome analysis
We sequenced and assembled three high-quality genomes (completeness > 99%, contamination < 1%) of the aquatic Enterobacter isolates. nEC134 and nEC135 genomes were highly similar to each other, based both on the 99.9% ANI and 5 out of 15,612 allele differences on wgMLST analysis. These genomes were classified as E. bugandensis (98.7% ANI against the GenBank genomes). nEC133 genome was more diverged when compared to nEC134 and nEC135 genomes (91.5–91.6% ANI, 2600 distinct alleles), and was classified as E. asburiae (97.5% ANI against the GenBank genomes). The genome-derived resistomes of all the three isolates were similar and included IMI, ACT and ampC-type beta-lactamases, as well as numerous genes that encode components of antibiotic efflux pumps, and other potential antibiotic resistance mechanisms (Table 3). E. asburiae nEC133 blaIMI was classified as IMI-2, based on 100% sequence identity (Fig. 2, Additional file 1: Fig. 1). E. bugandensis nEC134 and nEC135 carried a previously undescribed IMI-20 allele, whose sequence was distinguished from that of IMI-2 by a single nucleotide polymorphism (SNP), resulting in cysteine to phenylalanine substitution (Fig. 2, Additional file 1: Fig. 1). As in other Enterobacteriaceae, imiR gene that encodes a LysR-type regulator was found upstream of the blaIMI gene [8, 43]. We were not able to assemble complete plasmids using short reads, and only 3943–6219 bp long scaffolds included the blaIMI and imiR genes. However, plasFlow analysis suggested these scaffolds may be plasmid-related. The total length of scaffolds that were assigned to proteobacterial plasmids in E. asburiae nEC133 genome was 74,645 bp, which is comparable to that of the plasmid p3442-IMI-2 (78,374 bp). In E. bugandensis nEC134 and nEC135 genomes, the cumulative length of plasmid sequences was 142,368–176,174 bp, hinting at the presence of larger, or more than one plasmid. Genome assembly graphs of these isolates revealed a close linkage between the blaIMI-20 and plasmid scaffolds that carry the traXIDTHBFNCUWICVBKEL genes of the IncF conjugal transfer system, providing further evidence that blaIMI-20 is encoded on an IncF plasmid (Additional file 2: Fig. 2).

Phylogenetic tree of IMI type carbapenem-hydrolyzing class A beta-lactamases (alignment of 21 sequences of 292 amino acids). The tree is drawn to scale, with branch lengths representing the number of substitutions per site. The percentage of trees in which the associated taxa clustered together was determined based on 100 bootstrap resamples. Sequences from this study are marked in blue
Discussion
Our results suggest that CPE, as well as other MDR pathogens, occur in the coastal waters of a popular public recreational beach offshore Netanya, Israel and nearby river estuaries. These CPE appear to be abundant and well-acclimated to freshwater and seawater, as they were easy to isolate and at least on one occasion, the same E. bugandensis was found twice, in distant sampling sites, one month apart. It is important to state that at the time of our sampling (June and July 2018) no considerable fluctuations in seawater quality were reported [44]. Residents and tourists visiting popular recreation and leisure sites may be exposed to these MDROs.
The fact that isolates with highly similar genomes were isolated from both the riverine and marine environments, implies a mutual source, however, its origin is still unclear. Rivers are putative reservoirs and sources of MDROs, which were shown to be virulent [16, 45] and MDROs are less frequently encountered in patients drowning in seawater [46]. The pollution sources of Poleg River include discharge of urban sewage [47, 48], authorized dumping of treated sewage water [49] and cattle herding as seen on site. One of the main sources of Alexander River is the Nablus River, running from Nablus city through Tul-Karem located in the West Bank (Fig. 1). Other possible contamination sources include effluents from the adjacent WWTPs, a sea-turtles rescue center, an algae plant, an agricultural catchment basin and reservoir waters [30, 50]. Becker et al. (2013) reported that since 1995 several polluting sources were successfully treated, yet Wadi Zeimar, a major contamination source concentrating pollutants from Tul-Karem to Nablus River, remained [31]. The polluted rivers are the likely sources of marine contamination. Alternatively, coastal water can migrate up to e few kilometers inland, as shown by the daily salinity profiles measured by Ruppin Estuarine and Coastal Observatory [50], potentially reaching the possible contamination sources. Despite the limited mixing of water masses, bacterial cross-contamination may be substantial, hence bidirectional contamination is feasible.
Gut microbes may be adapted to the aquatic environment, in which their survival rates are poorly understood. Yet, cultivation-based studies demonstrate that common gut bacteria, such as Enterobacteriaceae, are frequently detected in freshwater [8, 10, 11, 15, 51, 52] and seawater [5,6,7]. These bacteria may be able to cope with different salinities because the human gut environment is characterized by spatial and temporal heterogeneity in osmolarity, based on the kinds of meals consumed [53]. The fitness of microbes and their growth rates depends not only on osmolarity but also on taxon-specific physiology and additional external factors such as nutrient abundance, pH and oxygen levels [54]. The three sequenced CP Eneterobacter spp. genomes encoded proteins that are involved in halotolerance, including the osmosensitive K+ channel histidine kinase KdpD, NhaA type Na+/H+ antiporter DNA-binding protein H-NS [55]. These sequences were found in scaffolds that were > 400 kbp in length and were defined as chromosomal by plasFlow. This indicates that at least some of the salt tolerance-related traits in these strains are not linked to plasmids as in other bacteria, and therefore not to blaIMI genes, which most likely are encoded on plasmids. Salinity has been recently shown to be the most important factor modulating the distribution patterns of antibiotic resistance genes in oceans and river beach soils [56], although other studies of aquatic ecosystems failed to show this [57]. Thus, it is feasible that less-studied traits that mediate salinity tolerance may be linked to antibiotic resistance genes.
Most importantly, these aquatic isolates are only remotely associated with the local clinical epidemiology. CP Enterobacter spp., are uncommon in our hospital’s clinical settings: between August 2013 and February 2019, we identified 129 Enterobacter spp. out of 798 CRE rectal screening isolates (16%). Carbapenemase-producing Enterobacter spp. were detected in 65 isolates. blaIMI genes were found only in three of these CPE isolates, while blaKPC (50 isolates) and blaNDM (10 isolates) were more common. However, the IMI mechanism can often go undetected, because only the five major enzymes (KPC, NDM, VIM, OXA-48 and IMP) are routinely tested. The clinical implication is that Enterobacter spp. carrying an unidentified blaIMI could have been misidentified as non-CP CREs. Since patients carrying non-CP CRE isolates are not cohorted in Israeli hospitals as CPE carriers, such misidentifications increase the potential for hospital cross-infection and outbreaks.
OXA-48 has also not been frequently encountered during this period in our facility (19 isolates out of 798 CREs, mostly seen in E. coli spp). Nevertheless, IMI carbapenemases appear to be emerging in clinical practice [58] as well as causing nosocomial outbreaks [59].
Conclusions
The widespread occurrence of CPE contamination in popular recreational beaches of the Sharon district is alarming and has major ramifications for environmental and public health, as well as to the public perceptions and awareness. However, occupation- or recreation-related seawater or river exposure is currently not considered a risk factor for CPE contamination, as it should probably be. It is important to consider MDROs (including CPE) when providing empiric antibiotic therapy for aspiration pneumonia during near-drowning as well as other injuries occurring during water-related activities, such as surfing and fishing.
Future large scale studies that will investigate the roles of the contamination sources along the river route to the sea, as well as resistome-related monitoring programs based on the frequent sampling of water and sediments, are crucial to mitigate aquatic CPE infection risks.
Availability of data and materials
Genomic sequences are available under the NCBI BioProject accession number PRJNA578038. The blaIMI-20 sequence was submitted to GeneBank with the accession number MN619794.
Abbreviations
- CRE:
-
Carbapenem-resistant enterobacteriaceae
- CPE:
-
Carbapenemase-producing enterobacteriaceae
- ESBL-PE:
-
Extended-spectrum β-lactamase-producing enterobacteriaceae
- MDRO:
-
Multidrug-resistant organism
- WWTP:
-
Wastewater treatment plants
- CLSI:
-
Clinical and Laboratory Standards Institute
- PCR:
-
Polymerase chain reaction
- wgMLST:
-
Whole-genome multilocus sequence typing
- RGI:
-
Resistance gene identifier
- CARD:
-
Comprehensive antibiotic resistance database
- ANI:
-
Average nucleotide identity
- SNP:
-
Single nucleotide polymorphism
References
van Loon K, In V, ’t Holt AF, Vos MC, . A systematic review and meta-analyses of the clinical epidemiology of carbapenem-resistant enterobacteriaceae. Antimicrob Agents Chemother. 2018;62:1.
Blyth DM, Mende K, Maranich AM, Beckius ML, Harnisch KA, Rosemann CA, et al. Antimicrobial resistance acquisition after international travel in US travelers. Trop Dis Travel Med Vacc. 2016;2:4.
Kock R, Daniels-Haardt I, Becker K, Mellmann A, Friedrich AW, Mevius D, et al. Carbapenem-resistant Enterobacteriaceae in wildlife, food-producing, and companion animals: a systematic review. Clin Microbiol Infect. 2018;24:1421.
Mahon BM, Brehony C, Cahill N, McGrath E, O’Connor L, Varley A, et al. Detection of OXA-48-like-producing Enterobacterales in Irish recreational water. Sci Total Environ. 2019;10(690):1–6.
Mahon BM, Brehony C, McGrath E, Killeen J, Cormican M, Hickey P, et al. Indistinguishable NDM-producing Escherichia coli isolated from recreational waters, sewage, and a clinical specimen in Ireland, 2016 to 2017. Euro Surveill. 2017;22:15.
Montezzi LF, Campana EH, Correa LL, Justo LH, Paschoal RP, da Silva IL, et al. Occurrence of carbapenemase-producing bacteria in coastal recreational waters. Int J Antimicrob Agents. 2015;45(2):174–7.
Paschoal RP, Campana EH, Correa LL, Montezzi LF, Barrueto LRL, da Silva IR, et al. Concentration and variety of carbapenemase producers in recreational coastal waters showing distinct levels of pollution. Antimicrob Agents Chemother. 2017;61:12.
Aubron C, Poirel L, Ash RJ, Nordmann P. Carbapenemase-producing Enterobacteriaceae, U. S. rivers. Emerg Infect Dis. 2005;11(2):260–4.
Khan FA, Hellmark B, Ehricht R, Soderquist B, Jass J. Related carbapenemase-producing Klebsiella isolates detected in both a hospital and associated aquatic environment in Sweden. Eur J Clin Microbiol Infect Dis. 2018;37(12):2241–51.
Kieffer N, Poirel L, Bessa LJ, Barbosa-Vasconcelos A, da Costa PM, Nordmann P. VIM-1, VIM-34, and IMP-8 carbapenemase-producing Escherichia coli strains recovered from a Portuguese river. Antimicrob Agents Chemother. 2016;60(4):2585–6.
Kittinger C, Lipp M, Folli B, Kirschner A, Baumert R, Galler H, et al. Enterobacteriaceae isolated from the river Danube: Antibiotic resistances, with a focus on the presence of ESBL and carbapenemases. PLoS ONE. 2016;11(11):e0165820.
Nascimento T, Cantamessa R, Melo L, Fernandes MR, Fraga E, Dropa M, et al. International high-risk clones of Klebsiella pneumoniae KPC-2/CC258 and Escherichia coli CTX-M-15/CC10 in urban lake waters. Sci Total Environ. 2017;15(598):910–5.
Oliveira S, Moura RA, Silva KC, Pavez M, McCulloch JA, Dropa M, et al. Isolation of KPC-2-producing Klebsiella pneumoniae strains belonging to the high-risk multiresistant clonal complex 11 (ST437 and ST340) in urban rivers. J Antimicrob Chemother. 2014;69(3):849–52.
Zarfel G, Lipp M, Gurtl E, Folli B, Baumert R, Kittinger C. Troubled water under the bridge: Screening of River Mur water reveals dominance of CTX-M harboring Escherichia coli and for the first time an environmental VIM-1 producer in Austria. Sci Total Environ. 2017;1(593–594):399–405.
Zurfluh K, Hachler H, Nuesch-Inderbinen M, Stephan R. Characteristics of extended-spectrum beta-lactamase- and carbapenemase-producing Enterobacteriaceae Isolates from rivers and lakes in Switzerland. Appl Environ Microbiol. 2013;79(9):3021–6.
Laurens C, Jean-Pierre H, Licznar-Fajardo P, Hantova S, Godreuil S, Martinez O, et al. Transmission of IMI-2 carbapenemase-producing Enterobacteriaceae from river water to human. J Glob Antimicrob Resist. 2018;15:88–92.
Poirel L, Barbosa-Vasconcelos A, Simoes RR, Da Costa PM, Liu W, Nordmann P. Environmental KPC-producing Escherichia coli isolates in Portugal. Antimicrob Agents Chemother. 2012;56(3):1662–3.
Diab M, Hamze M, Bonnet R, Saras E, Madec JY, Haenni M. Extended-spectrum beta-lactamase (ESBL)- and carbapenemase-producing Enterobacteriaceae in water sources in Lebanon. Vet Microbiol. 2018;217:97–103.
Tanner WD, VanDerslice JA, Goel RK, Leecaster MK, Fisher MA, Olstadt J, et al. Multi-state study of Enterobacteriaceae harboring extended-spectrum beta-lactamase and carbapenemase genes in U.S. drinking water. Sci Rep. 2019;9(1):3938.
Piedra-Carrasco N, Fabrega A, Calero-Caceres W, Cornejo-Sanchez T, Brown-Jaque M, Mir-Cros A, et al. Carbapenemase-producing enterobacteriaceae recovered from a Spanish river ecosystem. PLoS ONE. 2017;12(4):e0175246.
Mills MC, Lee J. The threat of carbapenem-resistant bacteria in the environment: evidence of widespread contamination of reservoirs at a global scale. Environ Pollut. 2019;255(Pt 1):113143.
Picao RC, Cardoso JP, Campana EH, Nicoletti AG, Petrolini FV, Assis DM, et al. The route of antimicrobial resistance from the hospital effluent to the environment: focus on the occurrence of KPC-producing Aeromonas spp and Enterobacteriaceae in sewage. Diagn Microbiol Infect Dis. 2013;76(1):80–5.
Zhang X, Lu X, Zong Z. Enterobacteriaceae producing the KPC-2 carbapenemase from hospital sewage. Diagn Microbiol Infect Dis. 2012;73(2):204–6.
Chagas TP, Seki LM, da Silva DM, Asensi MD. Occurrence of KPC-2-producing Klebsiella pneumoniae strains in hospital wastewater. J Hosp Infect. 2011;77(3):281.
Mathys DA, Mollenkopf DF, Feicht SM, Adams RJ, Albers AL, Stuever DM, et al. Carbapenemase-producing Enterobacteriaceae and Aeromonas spp present in wastewater treatment plant effluent and nearby surface waters in the US. PLoS ONE. 2019;14(6):e0218650.
Zurfluh K, Bagutti C, Brodmann P, Alt M, Schulze J, Fanning S, et al. Wastewater is a reservoir for clinically relevant carbapenemase- and 16s rRNA methylase-producing Enterobacteriaceae. Int J Antimicrob Agents. 2017;50(3):436–40.
He T, Wang Y, Sun L, Pang M, Zhang L, Wang R. Occurrence and characterization of blaNDM-5-positive Klebsiella pneumoniae isolates from dairy cows in Jiangsu, China. J Antimicrob Chemother. 2017;72(1):90–4.
Walsh TR, Weeks J, Livermore DM, Toleman MA. Dissemination of NDM-1 positive bacteria in the New Delhi environment and its implications for human health: an environmental point prevalence study. Lancet Infect Dis. 2011;11(5):355–62.
Aydin S, Aydin ME, Ulvi A, Kilic H. Antibiotics in hospital effluents: occurrence, contribution to urban wastewater, removal in a wastewater treatment plant, and environmental risk assessment. Environ Sci Pollut Res Int. 2019;26(1):544–58.
Israeli Ministry of Environmental Protection. https://www.sviva.gov.il/InfoServices/LicencesPerMissions/Rivers/Pages/PermissionToStream.aspx?WPID=WPQ7&PN=1. Accessed 8 August 2020.
Becker N, Friedler E. Integrated hydro-economic assessment of restoration of the Alexander-Zeimar River (Israel-Palestinian Authority). Reg Environ Change. 2013;13(1):103–14.
Schechner V, Straus-Robinson K, Schwartz D, Pfeffer I, Tarabeia J, Moskovich R, et al. Evaluation of PCR-based testing for surveillance of KPC-producing carbapenem-resistant members of the Enterobacteriaceae family. J Clin Microbiol. 2009;47(10):3261–5.
Bankevich A, Nurk S, Antipov D, Gurevich AA, Dvorkin M, Kulikov AS, et al. SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. J Comput Biol. 2012;19(5):455–77.
Parks DH, Imelfort M, Skennerton CT, Hugenholtz P, Tyson GW. CheckM: assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015;25(7):1043–55.
Miyoshi-Akiyama T, Hayakawa K, Ohmagari N, Shimojima M, Kirikae T. Multilocus sequence typing (MLST) for characterization of Enterobacter cloacae. PLoS ONE. 2013;8(6):e66358.
Jia B, Raphenya AR, Alcock B, Waglechner N, Guo P, Tsang KK, et al. CARD 2017: expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res. 2017;45(D1):D566–73.
Krawczyk PS, Lipinski L, Dziembowski A. PlasFlow: predicting plasmid sequences in metagenomic data using genome signatures. Nucleic Acids Res. 2018;46(6):e35.
Antipov D, Hartwick N, Shen M, Raiko M, Lapidus A, Pevzner PA. plasmidSPAdes: assembling plasmids from whole genome sequencing data. Bioinformatics. 2016;32(22):3380–7.
Yoon SH, Ha SM, Lim J, Kwon S, Chun J. A large-scale evaluation of algorithms to calculate average nucleotide identity. Antonie Van Leeuwenhoek. 2017;110(10):1281–6.
Yoon SH, Ha SM, Kwon S, Lim J, Kim Y, Seo H, et al. Introducing EzBioCloud: a taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int J Syst Evol Microbiol. 2017;67(5):1613–7.
Kumar S, Stecher G, Tamura K. MEGA7: molecular evolutionary genetics analysis version 7.0 for vigger datasets. Mol Biol Evol. 2016;33(7):1870–4.
Le SQ, Gascuel O. An improved general amino acid replacement matrix. Mol Biol Evol. 2008;25(7):1307–20.
Boyd DA, Mataseje LF, Davidson R, Delport JA, Fuller J, Hoang L, et al. Enterobacter cloacaecomplex isolates harboring blaNMC-A or blaIMI-type class A carbapenemase genes on novel chromosomal integrative elements and plasmids. Antimicrob Agents Chemother. 2017;61:5.
Israeli Ministry Of Health. https://www.health.gov.il/Subjects/Environmental_Health/bathing_sites/beach/Pages/sea_tests.aspx. Accessed 9 August 2020. 2018.
Tadie JM, Heming N, Serve E, Weiss N, Day N, Imbert A, et al. Drowning associated pneumonia: a descriptive cohort. Resuscitation. 2012;83(3):399–401.
Robert A, Danin PE, Quintard H, Degand N, Martis N, Doyen D, et al. Seawater drowning-associated pneumonia: a 10-year descriptive cohort in intensive care unit. Ann Intensive Care. 2017;7(1):45.
Israeli Ministry of Foreign Affairs. https://mfa.gov.il/MFA/PressRoom/1994/Pages/MARINE%20POLLUTION%20CONTROL.aspx. Accessed 9 August 2020.
Israel Ministry of Environment Protection, Rivers Pollutant Loads (2009). https://www.sviva.gov.il/InfoServices/ReservoirInfo/DocLib2/Publications/P0501-P0600/p0581.pdf. Accessed 17 August 2020.
Israeli Ministry of Environmental Protection. https://www.gov.il/he/Departments/dynamiccollectors/permission_to_stream?skip=0&limit=10&name_nachal=78. Accessed 9 August 2020.
Ruppin Estuarine and Coastal Observatory. https://reco.ruppin.ac.il. Accessed 9 August 2020.
Amaya E, Reyes D, Paniagua M, Calderon S, Rashid MU, Colque P, et al. Antibiotic resistance patterns of Escherichia coli isolates from different aquatic environmental sources in Leon, Nicaragua. Clin Microbiol Infect. 2012;18(9):E347–54.
Xu H, Wang X, Yu X, Zhang J, Guo L, Huang C, et al. First detection and genomics analysis of KPC-2-producing Citrobacter isolates from river sediments. Environ Pollut. 2018;235:931–7.
Fordtran JS, Locklear TW. Ionic constituents and osmolality of gastric and small-intestinal fluids after eating. Am J Dig Dis. 1966;11(7):503–21.
Brock TD. Microbial growth rates in nature. Bacteriol Rev. 1971;35(1):39–58.
Broaders E, O’Brien C, Gahan CG, Marchesi JR. Evidence for plasmid-mediated salt tolerance in the human gut microbiome and potential mechanisms. FEMS Microbiol Ecol. 2016;92:3.
Zhang YJ, Hu HW, Yan H, Wang JT, Lam SK, Chen QL, et al. Salinity as a predominant factor modulating the distribution patterns of antibiotic resistance genes in ocean and river beach soils. Sci Total Environ. 2019;1(668):193–203.
Bergeron S, Brown R, Homer J, Rehage S, Boopathy R. Presence of antibiotic resistance genes in different salinity gradients of freshwater to saltwater marshes in southeast Louisiana, USA. Int Biodeterior Biodegrad. 2016;113:80–7.
Miltgen G, Bonnin RA, Avril C, Benoit-Cattin T, Martak D, Leclaire A, et al. Outbreak of IMI-1 carbapenemase-producing colistin-resistant Enterobacter cloacae on the French island of Mayotte (Indian Ocean). Int J Antimicrob Agents. 2018;52(3):416–20.
Beyrouthy R, Barets M, Marion E, Dananche C, Dauwalder O, Robin F, et al. Novel enterobacter lineage as leading cause of nosocomial outbreak involving carbapenemase-producing strains. Emerg Infect Dis. 2018;24(8):1505–15.
Acknowledgements
None
Funding
Sanz Medical Center funded the sequencing costs.
Author information
Authors and Affiliations
Contributions
RC and PA conceived the study; SP cultivated bacteria, identified and tested antimicrobial susceptibility; AR and MRB analyzed the genomes; PA conducted environmental sampling. RC wrote the manuscript with contributions from all coauthors. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Additional file 1
. Figure 1: Alignment of IMI type carbapenem-hydrolyzing class A beta-lactamase amino acid sequences (see Figure 2 in the main text for accession numbers). Names of novel IMI alleles are marked in blue, cysteine to phenylalanine substitution is shown in the frame.
Additional file 2
. Figure 2: The link between blaMI-20 and traXIDTHBFNCUWICVBKEL genes of the IncF plasmid, based on the simplified de Bruijn assembly graph of the E. bugandensis nEC134 genome, generated by the SPAdes assembler and viewed in Bandage (the whole graph in the top panel and zoom in into the paths that contain the target genes in the bottom panel). The target genes are shown (blaIMI-20-red, lysR-yellow, traXIDTHBFNCUWICVBKEL-shades of blue).
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Cohen, R., Paikin, S., Rokney, A. et al. Multidrug-resistant enterobacteriaceae in coastal water: an emerging threat. Antimicrob Resist Infect Control 9, 169 (2020). https://doi.org/10.1186/s13756-020-00826-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13756-020-00826-2