
Abstract
Background
Pulp refining is an energy consuming, but integral part of paper production with the aim to increase tensile strength and smoothness of paper sheets. Commercial enzyme formulations are used to lower the energy requirements by pre-treatment of pulp before refining. However, a high number of different commercial enzyme products are available on the market containing enzymes of varying origin and composition, which complicates the prediction of their behavior, especially using different pulp types.
Results
Endoglucanase-rich enzyme formulations were characterized regarding enzyme activity at different temperatures, resulting in a significant decrease of activity above 70 °C. Some enzyme preparations additionally contained arabinosidase, xylanase and β-glucosidase activity consequently resulting in a release of xylose and glucose from pulp as determined by high-performance liquid chromatography. Interestingly, one enzyme formulation even showed lytic polysaccharide monooxygenase (LPMO) activity of 3.05 nkat mg−1. A correlation between enzyme activity using the endoglucanase specific derivatized cellopentaose (CellG5) substrate and enzyme performance in laboratory PFI (Papirindustriens forskningsinstitut) refining trials was observed on softwood pulp resulting in a maximum increase in the degree of refining values from 27.7°SR to 32.7°SR. When added to a purified endoglucanase enzyme (31.6°SR), synergistic effects were found for cellobiohydrolase II (34.7°SR) or β-glucosidase enzymes (35.7°SR) in laboratory refining. Comparison with previously obtained laboratory refining results on hardwood pulp allowed differences in enzyme performance based on varying pulp types to be elucidated.
Conclusions
Interestingly, the individual enzymes indeed showed different refining effects on softwood and hardwood pulp. This difference could be predicted after development of an adapted enzyme activity assay by combination of the derivatized cellopentaose CellG5 substrate with either softwood or hardwood sulfate pulp.
Similar content being viewed by others

Background
The refining of pulp is a crucial step during paper production, as it gives paper its defined properties such as increased tensile properties and improved smoothness. The refining process consists of a mechanical treatment, which generates internal and external fibrillation. Internal fibrillation leads to breakage of hydrogen bonds between cellulose microfibrils, thus enabling water molecules to enter the interior of the fibers, causing swelling and an increase in fiber flexibility and plasticity, resulting in increased fiber bonding. During external fibrillation, fibrils are peeled off from the surface, which enhances the tensile strength through mechanical interlocking [1,2,3]. However, the refining process is energy-demanding, requiring up to 30% of the total energy needed for paper production. Therefore, any means to reduce energy consumption are appreciated, considering a total amount of 412.5 million tons of paper and board products were produced in 2019, which required a total amount of 94 354 GWh of electricity [4, 5]. Enzymes like cellulases increasingly came into focus to lower the power consumption by pre-treatment of pulp before the refining process, thereby allowing the implementation of sustainable production systems. Depending on the used enzyme, between 10 and 40% of energy can be saved, however, the high number of different enzyme formulations on the market containing enzymes of different origin and composition, makes the prediction of the enzyme behavior among various formulations and pulp types difficult and thereby the selection of the most efficient enzymes a challenge [6, 7].
Cellulase is an umbrella term for several individual enzymes, which in combination synergistically degrade cellulose into the main building block glucose. Cellulases comprise endoglucanases (EC 3.2.1.4), cellobiohydrolases (EC 3.2.1-), β-glucosidases (EC 3.2.1.21) as well as oxidative enzymes such as lytic polysaccharide monooxygenases (EC 1.14.99-). Endoglucanases (endo-1–4-β-glucanases) are cutting cellulose chains randomly and internally by hydrolysis of β-1–4 bonds in the amorphous region of cellulose. Cellobiohydrolases function as exo-glucanases and attack the cellulose chain at reducing and non-reducing ends, ultimately releasing cellobiose units, while β-glucosidases transform cellobiose units released by endoglucanases and cellobiohydrolases into glucose [8,9,10].
Endoglucanases are primarily the enzymes used for the reduction of energy consumption during the refining process. Endoglucanases introduce weak points within the cellulose fiber, thereby causing the loosening of the entire cellulose structure, which enables water molecules to enter and break the hydrogen bond network connecting the fibers [11,12,13,14]. However, β-glucosidases may still be required to prevent product inhibition of endoglucanases and cellobiohydrolases (CBH) through accumulation of cellobiose [15]. The role of cellobiohydrolases in pulp refining is still contradictory. For example, a study examining the effect of CBH I on pulp refining showed no effect on hardwood pulp [16]. Similar observations were made in a study investigating the cellobiohydrolases CBH I and CBH II, resulting in no effect in refining, while the combination of CBH I with endoglucanase EG I had similar effects as the treatment with endoglucanase alone [17]. However, it was also reported in another study that the addition of cellobiohydrolase I can indeed lead to energy savings during pulp refining [18]. Lytic polysaccharide monooxygenases (LPMOs) are enzymes that aid other cellulase enzymes in the degradation of cellulose fibers through oxidative action on the tightly packed crystalline structure, which is usually recalcitrant to enzymatic degradation. Through oxidative cleavage of cellulose by these copper containing enzymes, the highly ordered crystalline structure is disrupted, becomes less ordered and para-crystalline, which improves the accessibility to other enzymes like endoglucanases that act mainly in the amorphous region of cellulose [19, 20].
The origin of pulp (e.g., softwood or hardwood species) is expected to have a strong influence on the effect of enzymes. For example, softwood pulps comprise conifer species such as spruce (Picea abies), while hardwood pulps are derived from species like birch (Betula spp.) or eucalyptus (e.g., Eucalyptus grandis) [21,22,23]. However, also the pulping process has an effect, which removes lignin from wood, thereby enriching the cellulose content [24]. Today two different pulping processes are established, the sulfate (kraft) and the sulfite process. The sulfate process uses sodium hydroxide and sodium sulfide to cook the wood chips and is the dominant form because of improved tensile properties [25, 26]. The sulfite process, on the other hand, utilizes sulfite in the cooking liquor such as sodium sulfite or calcium, magnesium, or ammonium bisulfite and can be used over a broad pH range [27,28,29].
In our previous studies, endoglucanase activity on derivatized cellopentaose (CellG5) was successfully correlated to enzyme performance in laboratory refining using hardwood pulp [30,31,32]. However, the enzyme behavior on other pulp types such as softwood pulp still needs to be studied when dosed according to CellG5. In addition, considerably less is known about the individual effect of different endoglucanases and the necessity of accessory enzymes (e.g., arabinosidases, cellobiohydrolases and β-glucosidases) in refining, which was further elucidated in this study. Finally, an adapted activity assay was developed and evaluated with the aim to improve the prediction of enzyme behavior in pulp refining by combination of the endoglucanase specific derivatized cellopentaose substrate (CellG5) with either softwood or hardwood pulp.
Materials and methods
Pulps, enzyme formulations and chemicals
Bleached softwood sulfate pulp (approx. 80% spruce, 20% pine and larch) and bleached hardwood sulfate pulp (eucalyptus) as well as commercial enzyme formulations (EnzA, EnzB, EnzC) were provided from the Austrian pulp and paper industry. Bleached softwood sulfate pulp is composed of 6.6% hemicelluloses, while bleached hardwood sulfate pulp contains 12.2% hemicelluloses based on 13C NMR. The commercial endoglucanase FiberCare R was kindly provided by Novozymes A/S (Bagsværd, Denmark). All other chemicals were obtained from Sigma-Aldrich (Austria) in HPLC grade if not stated otherwise. The β-glucosidase E-BGLUC from Aspergillus niger, the microbial cellobiohydrolase II (E-CBHIIM) as well as the CellG5 cellulase and XylX6 xylanase enzyme activity assays were obtained from Megazyme (Bray, Ireland).
Endoglucanase purification
The endoglucanases EndoA, EndoB and EndoC were purified from the enzyme formulations EnzA, EnzB and EnzC using a combination of hydrophobic interaction (HIC) and anion exchange chromatography (AEX) as described in previous studies [30]. In brief: enzyme formulations were suspended in 70 ml binding buffer A (10 mM acetate buffer + 1.5 M (NH4)2SO4 at pH 4.8) and applied to the HIC column. Endoglucanases were eluted after column washing with buffer A. EndoA was eluted by a 30% buffer B (10 mM acetate buffer pH 4.8) step followed by a linear gradient of 30–80% B. EndoB was obtained by using a 50% B step, followed by a linear gradient from 50 to 100% B and EndoC by a 45% B step, followed by a linear gradient of 45%-90% B. Desired enzyme fractions were identified by SDS-PAGE, concentrated using Vivaflow 50 membranes (Sartorius, Germany) and buffer exchanged to anion exchange buffer A (10 mM Tris pH 7.5) with PD-10 columns (Cytiva, USA). AEX was performed using a HiTrap DEAE FF 5 ml column. A total sample volume of 25 ml was applied, and elution started after a column wash with buffer A. EndoA and EndoB were eluted with a 0–7% buffer B (10 mM Tris pH 7.5 + 1 M NaCl) and EndoC using a 0–1.5% B gradient. Desired fractions were concentrated after SDS-PAGE using Vivaspin 20 centrifugal concentrators (Sartorius, Germany) and buffer exchanged to 50 mM citrate buffer (pH 4.8) with PD-10 columns (Cytiva, USA).
Laboratory refining trials using softwood and hardwood pulp
Laboratory refining experiments were conducted using 130 g dry softwood or hardwood sulfate pulp. Pulps were pre-soaked overnight and disintegrated for 10 min. Enzyme treatment was conducted using a 4% w/w pulp concentration in 50% tap water + 50% deionized water (adjusted to pH 7) at 45 °C for 30 min under continuous agitation and stopped through addition of 30% hydrogen peroxide. Enzyme treatment using enzyme formulations and pure endoglucanases was performed at an enzyme dosage of 5.06 nkat according to the Megazyme CellG5 activity assay. For the study of enzyme synergisms, cellobiohydrolase II from Megazyme was added at a dosage of 0.17 nkat, while β-glucosidase from Megazyme was added using a dosage of 16.7 nkat. Refining experiments were subsequently performed utilizing a PFI (Papirindustriens forskningsinstitut) laboratory mill type Mark IV (Hamjern Maskin AS, Norway) at 4000 and 6000 revolutions according to ISO 5264–2:2011. The achieved degree of refining was determined using the Schopper–Riegler method (ISO 5267–1:1999). Handsheets were formed using a RK4-KWT sheet former from Frank PTI (Austria) according to Rapid–Köthen (ISO 5269–2:2004). The tensile strength of the paper sheets was evaluated according to ISO 1924–2:2008 and air permeability Bendtsen according to ISO 5636–3:2013 after conditioning the samples for at least 24 h in a climate room at 23 °C and 50% relative humidity. A total number of 13–15 paper samples were tested. Tensile index values are available in Additional file 1: Fig. S1. Error bars indicate the measured standard deviation.
Enzyme activity assays
Endoglucanase activity on derivatized cellopentaose (CellG5)
Endoglucanase activity was measured at 45 °C, 55 °C and 70 °C. For this purpose, the endoglucanase specific CellG5 assay from Megazyme (Bray, Ireland) was used. The CellG5 substrate comprised 4,6-O-(3-ketobutylidene)-4-nitrophenyl-β-D-cellopentaoside (BPNPG5) and a thermostable β-glucosidase. The assay was conducted similarly as described in [33]. A volume of 100 µl CellG5 substrate was placed in a 15-ml glass tube. Enzymes were diluted in 50 mM citrate buffer (pH 4.8) or 50% tap water + 50% deionized water (adjusted to pH 7) and 100 µl of diluted enzyme solution was added after equilibration of the glass tube at the respective temperature. The glass tube was subsequently incubated for 10 min. The enzyme reaction was stopped using 3 ml of 2% Tris solution (pH 10). The reaction mixture was then transferred to 3 ml cuvettes and the absorbance measured at 400 nm using a Hitachi U2900 spectrophotometer (Chiyoda, Japan). Enzyme activity was calculated in nkat ml−1. Measurements were performed in duplicates and error bars indicate the measured standard deviation.
β-Glucosidase activity
ß-Glucosidase activity was determined at 45 °C, 55 °C and 70 °C using the substrate 4-nitrophenyl β-d-glucopyranoside (Sigma-Aldrich, Austria) similar as described in [34]. Therefore, a 2 mM 4-nitrophenyl β-D-glucopyranoside substrate solution was prepared. Enzymes were diluted in 50 mM citrate buffer (pH 4.8) and a volume of 50 µl was added to 200 µl of substrate solution in 15-ml glass tubes, which were equilibrated in a water bath at the respective temperature before enzyme addition. The enzyme reactions were stopped after 0, 10, 20, 30, 40, 50 and 60 min using 500 µl of methanol. A volume of 750 µl of 500 mM NaPO4 buffer (pH 7) was added for colorization and 200 µl were transferred into a 96-well plate. The absorbance was measured at 410 nm using an Infinite M200 Pro plate reader (Tecan, Switzerland). Enzyme activity was calculated in nkat ml−1. Measurements were performed in duplicates and error bars indicate the standard deviation.
Filter paper cellulase activity (FPU)
The cellulase activity using Whatman No. 1 filter paper was determined according to the standard procedure of the International Union of Pure and Applied Chemistry (IUPAC) with minor modifications [35]. Therefore, 7.5 cm × 0.75 mm strips of filter paper were prepared and suspended in 800 µl citrate buffer (50 mM, pH 4.8) using 15-ml glass tubes. The glass tubes were equilibrated at the respective temperature and a volume of 200 µl enzyme solution was added. The enzyme reaction was stopped by addition of 1000 µl 1 M NaOH after 0, 5, 10, 20, 40 and 60 min. For colorization, 1000 µl of DNS (3,5-dinitrosalicylic acid) solution was added and the glass tubes were boiled in a boiling water bath for 5 min. A volume of 200 µl was then transferred to a 96-well plate and the absorbance was measured at 540 nm using an Infinite M200 Pro plate reader (Tecan, Switzerland). Measurements were performed in duplicates and enzyme activity was calculated in nkat ml−1. Error bars indicate the standard deviation.
Xylanase activity
Xylanase activity was determined using the xylanase-specific XylX6 activity assay from Megazyme (Bray, Ireland) similar to that described in [36]. Therefore, the XylX6 substrate was used, which comprised 4,6-O-(3-ketobutylidene)-4-nitrophenyl-β-D-45-glucosyl-xylopentaoside and a β-xylosidase. The sample was diluted in 50 mM citrate buffer (pH 4.8) and 50 µl diluted enzyme solution was added to 50 µl of XylX6 substrate in a 15-ml glass tube. The reaction was incubated for 10 min at 45 °C in a water bath and stopped by addition of 1.5 ml of 2% Tris solution (pH 10). The solution was then transferred to 3-ml cuvettes and the absorbance measured at 400 nm using a Hitachi U2900 photometer (Chiyoda, Japan). Measurements were performed in duplicates and enzyme activity was calculated in nkat ml−1. Error bars indicate the standard deviation.
Arabinosidase activity
Arabinosidase activity was determined using 4-nitrophenyl-α-L arabinopyranoside as substrate similar as described in [37], but with some modifications. Therefore, a 2 mM 4-nitrophenyl-α-L arabinopyranoside substrate solution was prepared in 50 mM citrate buffer (pH 4.8) or 50% tap water + 50% deionized water at pH 7 (refining conditions). A volume of 200 µl substrate solution was placed in 15-ml glass tubes and equilibrated at 45 °C in a water bath. The enzyme reaction was started by addition of 50 µl of diluted enzyme solution and stopped after 0, 5, 10, 20, 40 and 60 min by addition of 500 µl methanol. For colorization, 750 µl of 500 mM NaPO4 buffer (pH 7) was added and 200 µl were transferred into a 96-well plate. Absorbance was measured at 410 nm using an Infinite M200 Pro plate reader (Tecan, Switzerland). Measurements were performed in duplicates and the enzyme activity was subsequently calculated in nkat ml−1.
Lytic polysaccharide monooxygenase (LPMO) activity
The activity of lytic polysaccharide monooxygenases was determined using a 10 mM 2,6-dimethoxyphenol substrate solution similar to that described in [38]. Therefore, a volume of 1720 µl of 50% tap water and 50% deionized water (adjusted to pH 7) was placed in a 3-ml cuvette and 200 µl of 2,6-dimethoxyphenol substrate solution was added. The cuvette was incubated at 45 °C for 15 min after addition of 40 µl of 5 mM H2O2 solution. Enzymes were diluted in 50% tap water + 50% deionized water (adjusted to pH 7) and the reaction was started by addition of 40 µl of enzyme solution. A blank with 50% tap water and 50% deionized water (pH 7) instead of enzyme solution was measured along with all samples. The absorbance at 469 nm was recorded using a Hitachi U2900 spectrophotometer (Chiyoda, Japan) for 300 s and the enzyme activity was calculated from the recorded slope using the extinction coefficient of the produced product coerulignone (53.200 M−1 cm−1). Measurements were performed in duplicates.
Cellobiohydrolase II activity
Cellobiohydrolase II activity was measured using a 2 mM 1,4-β-D-cellopentaitol substrate solution. A volume of 200 µl substrate solution was placed in a 15-ml glass tube and equilibrated at 45 °C in a water bath. Enzymes were diluted in 50% tap water + 50% deionized water (adjusted to pH 7) and a volume of 50 µl was added to the glass tube to start the enzyme reaction. The reaction was stopped after 0, 10, 20, 40 and 60 min by addition of 250 µl 1 M NaOH. For colorization, 250 µl of DNS (3,5-dinitrosalicylic acid) solution was added and the glass tube boiled in a boiling water bath for 5 min. A volume of 200 µl was transferred to a 96-well plate and the absorbance was measured at 540 nm using an Infinite M200 Pro plate reader (Tecan, Switzerland). Measurements were performed in duplicates and the enzyme activity was calculated in nkat ml−1.
Activity assay after adsorption of derivatized cellopentaose (CellG5) on pulp
Endoglucanase activity was measured using the endoglucanase specific activity assay CellG5 from Megazyme, which is based on derivatized cellopentaose [33]. To test if this substrate can also be used in combination with pulp to consider potential sorption effects, a volume of 100 µl of CellG5 substrate was mixed with 5 mg of hardwood or softwood pulp in 15-ml glass tubes and incubated for 15 min. The CellG5 substrate was subsequently removed, and the pulps were washed 3 × with MQ water. After suspension of the pulps in 100 µl 50% tap water + 50% deionized water (pH 7), the glass tubes were equilibrated in a water bath at 45 °C and a volume of 100 µL diluted enzyme solution was added to start the enzyme reaction. The reaction was conducted for 10 min and stopped by addition of 3 ml of Tris solution (pH 10). The solution was then transferred to 3-ml cuvettes and the absorbance measured at 400 nm using a Hitachi U2900 spectrophotometer (Chiyoda, Japan). Enzyme activity was calculated in nkat ml−1. Measurements were performed in duplicates and error bars indicate the standard deviation.
HPLC analysis of sugar release
An amount of 50 mg of hardwood or softwood sulfate pulp was incubated with enzyme formulations at a dosage of 166.7 nkat for 48 h in 1250 µl of 50 mM citrate buffer (pH 4.8). For the study of the synergistic effect of cellobiohydrolase II, a CBH II activity of 0.83 nkat was added to 0.83 nkat of EndoC endoglucanase activity and the reaction subsequently incubated for 4 h in 1250 µl of 50 mM citrate buffer (pH 4.8). The supernatant was removed and diluted 1:10 with MQ water after inactivation of the enzyme reaction by incubation at 99 °C for 5 min on a thermomixer. Proteins and lipids were removed by Carrez precipitation. Therefore, 20 µl of 2% potassium hexacyanoferrate(II) trihydrate solution and 20 µl of 2% zinc sulfate heptahydrate solution was added and the samples were centrifuged for 30 min at 12,500 rpm using a Eppendorf 5427 R centrifuge equipped with an FA-45–48-11 rotor (Hamburg, Germany). Afterwards, the supernatant was filtrated through a 0.45-µm filter into glass vials, which were subsequently applied to an Agilent 1260 Infinity LC system (Santa Clara, USA), equipped with an ICSep ION-300 (Transgenomic Inc) column and a refraction index detector. HPLC analysis was performed for 45 min at a flow rate of 0.325 ml min−1 using 0.01 M H2SO4 as the mobile phase at a temperature of 45 °C. Glucose and xylose standards ranging between 0–0.96 mg ml−1 were measured along with the samples for calculation of the sample concentration in mg ml−1. Measurements were performed in duplicates and error bars indicate the standard deviation.
Scanning electron microscopy of refined sample sheets
Formed paper handsheets were analyzed using scanning electron microscopy (SEM) after laboratory refining. Therefore, small pieces were cut-off and sputter coated with 1 nm platinum using the Leica EM ACE 200 sputter coater (Wetzlar, Germany). Sputter-coated samples were subsequently visualized using the Hitachi 3030 scanning electron microscope (Chiyoda, Japan) at 1000 × magnification. SEM pictures of refined handsheets after 4000 and 6000 revolutions are shown in Additional file 1: Figs. S2–S5.
Determination of protein content
Protein content was determined by measuring the absorbance at 280 nm using the NanoDrop NP80 (Implen, Germany). A volume of 2 µl of 50 mM citrate buffer (pH 4.8) was used as a blank, after which the samples were measured in duplicates. For the calculation of the protein concentration in mg ml−1, the extinction coefficient of BSA was used: 44.289 [M−1*cm−1].
Results and discussion
Enzyme activity of enzyme formulations
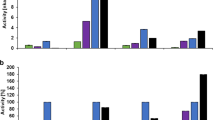
A high number of different commercial enzyme formulations is available on the market while knowledge about individual enzyme activities is essential for their correct dosing in refining. This is achieving the desired effect (e.g., energy savings) while, on the other hand, avoiding fiber damage due to excessive hydrolysis. For pulp refining, endoglucanase activity is of major importance, as these enzymes are responsible for reduction of energy consumption [30]. Endoglucanase activity on the endoglucanase-specific derivatized cellopentaose substrate CellG5 showed a high variation between the commercial enzyme formulations (Fig. 1a). The highest activity at 45 °C was determined for FiberCare R (13.08 nkat mg−1), which is reasonable, considering it is marketed as an endoglucanase only formulation. EnzC showed a slightly lower activity at 45 °C, while both EnzA and EnzB, exhibited the lowest activities on derivatized cellopentaose (CellG5). Endoglucanase activity increased at 55 °C for most enzyme formulations (except EnzB), while at 70 °C enzyme activities decreased considerably (lowest for EnzA: 0.52 nkat mg−1). However, for Fiber Care R, this decrease was only minor, which suggests a high temperature stability for this enzyme, while activity of EnzC decreased to a large extent (Fig. 1a). Similar behavior is known for cellulases such as EGI of Trichoderma reesei, which exhibits only a short half-life above 50 °C. On the other hand, endoglucanases of organisms like Thermoascus aurantiacus exhibit high thermostability even at 70 °C and above [39,40,41].β-Glucosidase activity was highest for the EnzC formulation (1.08 nkat mg−1) at 45 °C (Fig. 1b), while the lowest activity was determined for EnzB (0.21 nkat mg−1). β-Glucosidase activity increased at 55° for most formulations, with EnzC having more than a two times higher activity (2.47 nkat mg−1). At 70 °C, the β-glucosidase activity of EnzA decreased, while that of EnzC resulted in the highest measured value (3.43 nkat mg−1). Therefore, more cello-oligosaccharides released from endoglucanases would be hydrolyzed to glucose at higher temperatures, which is not desired in refining since it potentially requires extensive wastewater treatments. On the other hand, high β-glucosidase activity could enhance endoglucanase performance by preventing product inhibition through processing released cellobiose units into glucose [31, 42].

Endoglucanase, β-glucosidase, filter paper as well as xylanase activity in nkat mg−1 of commercial enzyme formulations EnzA, EnzB, EnzC and the commercial endoglucanase formulation FiberCare R at 45 °C, 55 °C and 70 °C using 50 mM citrate buffer (pH 4.8). Statistical significance was tested using a single factor ANOVA test with a significance level of 0.05 (significantly different results are indicated by the letter s, not significantly different results by the letter n)
The filter paper activity assay is a common determination method for cellulase activity and was therefore previously used for dosage in pulp refining or recycling processes [11, 43]. Interestingly, the highest activity was measured for FiberCare R, as this assay is also influenced by synergistic action in conjunction with β-glucosidases, which are only present in neglectable amounts in this formulation (Fig. 1c). EnzC and EnzB exhibited similar activity at 45 °C on filter paper (1.56 and 1.31 nkat mg−1, respectively) and EnzA the lowest effect. However, the activity of EnzC increased considerably at 70 °C, despite of having only a slightly higher endoglucanase activity, which suggests the increase is mainly related to the elevated β-glucosidase activity (Fig. 1b). This is also apparent for 70 °C, with EnzC still showing a high activity. This synergistic nature of the filter paper assay complicates prediction of processes that require only endoglucanase activity like pulp refining. Inaccuracies of the filter paper assay through varying amounts of β-glucosidases were previously reported with the recommendation to add additional β-glucosidases for complete conversion to glucose. However, the addition of β-glucosidases also influences the activity of endoglucanases by causing a synergistic effect [15, 44, 45]. Comparison of the filter paper assay with determined endoglucanase activity also confirms the synergistic nature of the filter paper assay caused by additional enzymes as the activity of EnzB was much higher on filter paper possibly through contribution of xylanases and ß-glucosidases (Fig. 1a vs. c).
Moreover, the xylanase activity is of interest as xylanases have shown to also increase pulp refining performance [46, 47]. The highest xylanase activity using the xylanase-specific XylX6 activity assay was by far found for EnzB at 45 °C (45.06 nkat mg−1), the lowest for EnzA with a value of 1.69 nkat mg−1 (Fig. 1d). Interestingly, xylanase activity decreased considerably at 55 °C for all enzyme formulations with EnzB showing the highest activity (10.04 nkat mg−1). At 70 °C, xylanase activity decreased even further with EnzA having the highest residual activity (0.42 nkat mg−1). Therefore, the xylanase enzymes in the enzyme formulations exhibit almost no activity at higher temperatures, which needs to be considered in the selection of enzyme formulations for pulp refining.
Additional enzymes that could be present in complex formulations are arabinosidases which are able to liberate arabinose from side groups of xylans [48, 49]. The highest arabinosidase activity using 4-nitrophenyl-⍺-L arabinopyranoside was measured for EnzC (0.12 nkat mg−1), the lowest for EnzA (0.017 nkat mg−1) in citrate buffer pH 4.8 (Table 1). The activity was lower under refining conditions (50% tap water + 50% deionized water pH 7) for most enzyme formulations and a different pattern could be observed, with EnzA being the enzyme with the highest activity (0.019 nkat mg−1) and EnzB with the lowest (0.0061 nkat mg−1). It was reported that some hemicellulases such as the xylosidase Dictyoglomus thermophilum have an arabinosidase side activity, while a GH43 ⍺-L-arabinofuranosidase showed activity towards arabinopyranoside as well as arabinofuranoside. However, also an individual GH27 family arabinosidase enzyme was isolated from the bacteria Streptomyces avermitilis exhibiting high activity on arabinopyranoside, which was able to hydrolyse arabinogalactan that is present in plants such as coniferous woods [37, 50, 51].
The presence of enzymes such as arabinosidases could explain differences between the effects of enzyme formulations in pulp refining, as these enzymes could improve accessibility for endoglucanases through synergistic degradation of xylans in conjunction with xylanases. Moreover, it was also described, that the presence of arabinosyl residues is impairing adsorption of xlyan to cellulose and therefore, the reduction of xylan substitutions is increasing pulp strength through improvement of xylan–cellulose interaction via rearrangement of the connecting hydrogen bond network [52,53,54].
In addition, the activity of lytic polysaccharide monooxygenases (LPMOs) was determined. These enzymes were until now mainly associated with lignocellulose degradation, during which they oxidatively cleave glycosidic bonds of cellulose and act as auxiliary enzymes to cellulases [55]. Thereby, the presence of such enzymes in enzyme formulations could enhance or decrease the refining performance. Indeed, no lytic polysaccharide monooxygenase activity was measured for most enzyme formulations and endoglucanases, however, an LPMO activity of 3.05 nkat mg−1 was determined for EnzC. LPMOs assist cellulases by disrupting the crystalline cellulose packaging, which results in the transformation of these areas into less-ordered or para-crystalline regions that are better accessible to enzymatic treatment. The presence of LPMOs could therefore be beneficial for pulp refining, as a higher susceptibility of LPMO-treated cellulose fibers to mechanical treatment was reported in a previous study [19].
Laboratory refining of enzyme formulations on softwood sulfate pulp
In contrast to hardwood pulp, there is only little known about the effect of individual enzymes in refining of softwood pulp, especially when dosed according to derivatized cellopentaose (CellG5). Nevertheless, more information would be important considering that softwood pulps have different characteristics such as varying types and amounts of hemicelluloses [6, 56]. Laboratory PFI refining trials on softwood sulfate pulp were therefore conducted to elucidate differences compared to refining on hardwood pulp. One of the major properties for the assessment of the outcome of the refining process is the drainability of pulp, which is characterized by the Schopper–Riegler degree (°SR). The Schopper–Riegler degree is expected to increase during refining due to the increased fiber bonding. Laboratory refining was performed at two different intensities, 4000 and 6000 PFI revolutions, while the highest effect was observed using 6000 PFI revolutions.
At 6000 PFI revolutions, the reference without enzyme addition resulted in a degree of refining value of 27.7°SR. Indeed, enzyme addition increased the degree of refining, with EnzC achieving the highest effect (32.7°SR), followed by FiberCare R (31.2°SR), although to an already much lower extent. Thereby, a lower amount of refiner revolutions is required to reach a certain degree of refining using these two enzymes, which corresponds to a lower energy consumption. However, the enzyme formulations EnzB (29.9°SR) and EnzA (27.2°SR) showed no, or only minor improvement compared to the reference, which emphasizes the need for more accurate prediction methods (Fig. 2a).

Laboratory refining data examining commercial enzyme formulations as well as enzyme synergisms using endoglucanase, β-glucosidase and cellobiohydrolase II enzymes after 4000 and 6000 PFI refiner revolutions. a–d Show the results of the degree of refining and air permeability Bendtsen measurements of the complex enzyme formulations EnzA, EnzB, EnzC and the commercial endoglucanase FiberCare R on softwood pulp. In addition, the pure endoglucanases EndoA, EndoB, EndoC and synergistic effects through addition of β-glucosidase and cellobiohydrolase II to EndoC were investigated, e–h show the results of the degree of refining and air permeability Bendtsen of enzyme formulations and pure endoglucanases on hardwood sulfate pulp. Degree of refining measurements were conducted according to ISO 5267–1:1999 and therefore no standard deviation is given. Data of hardwood pulp were modified from [30]. Statistical significance was tested using a single factor ANOVA test with a significance level of 0.05 (significantly different results are indicated by the letter s, not significantly different results by the letter n)
EnzC is also the only enzyme formulation with LPMO activity; therefore, these enzymes could explain the high refining performance of this formulation. Another property for the characterization of refined paper sheets is the air permeability. The air permeability is expected to decrease during pulp refining as the sheet density and compactness is increased through enhanced fiber bonding, which limits air flow [57, 58]. EnzC and FiberCare R decreased the measured air permeability the most by showing a value of 222 ml min−1 and 254 ml min−1 (Fig. 2b), respectively, while the reference achieved a value of 262 ml min−1. This in accordance with the measured degree of refining value after 6000 revolutions, however, for EnzB (284 ml min−1) and EnzA (351 ml min−1), the obtained values were even higher than the reference (262 ml min−1). This suggests a negative impact by the enzymes present in these formulations, possibly through disruption and degradation of the cellulose structure, which decreases paper density.
Interestingly, the comparison with reported data on laboratory refining of hardwood sulfate pulp revealed differences in enzyme behavior between both pulps (Fig. 2e.). It is apparent that EnzC performed most efficient on both pulps, followed by FiberCare R, although the latter was already much less effective on softwood pulp. Moreover, EnzA and EnzB, which exhibited a minor effect on hardwood pulp, were almost not effective at all on softwood pulp [30]. The air permeability according to Bendtsen decreased significantly as well for most of the enzyme formulations on hardwood pulp with EnzC exhibiting the lowest value (149 ml min−1) and EnzA the highest (213 ml min−1). However, all enzyme formulations achieved a better result than the reference on hardwood pulp, which was not the case for softwood pulp for EnzA and EnzB (Fig. 2b). This emphasizes the need for prediction methods that can project the diverging enzyme behavior on different pulp types.
Differences in enzymatic behavior between pulp types can also be related to varying content of hemicelluloses or lignin. Indeed, used bleached hardwood sulfate pulp contains a higher fraction of hemicelluloses (12.2%), while bleached softwood sulfate pulp only exhibits 6.6%. Usually, a higher content of hemicelluloses corresponds to lower enzyme activity of cellulases because of inhibitory effects. However, despite hardwood pulp having a higher hemicellulose content, the increase in degree of refining and pulp density was higher on hardwood pulp compared to softwood pulp, therefore, varying hemicellulose content cannot explain the differences in enzymatic behavior for pulp refining. For example, the EnzB enzyme formulation exhibited the highest xylanase activity, but could not achieve a higher refining performance on softwood pulp. This could be explained by excessive degradation of hemicelluloses. This is supported by a study investigating the effect of xylanases on refining, which concluded that the best paper properties are obtained by xylanases causing the smallest decrease in hemicellulose content [46].
Another factor that can inhibit enzyme activity is the lignin content, which is usually higher for softwood plants compared to hardwood plants [59,60,61]. However, for bleached pulp, residual lignin is only present at neglectable levels, as any remaining lignin from the pulping process is almost completely removed by the pulp bleaching process. For example, bleached softwood sulfate pulp only contains a maximum lignin fraction of 0.30%, while industrial bleached hardwood eucalyptus pulp typically contains 0.27% lignin based on kappa number of 1.8% [6, 62, 63].
Laboratory refining of pure endoglucanases and possible enzyme synergisms
The refining effect of the purified endoglucanases EndoA, EndoB and EndoC was studied on softwood pulp to potentially identify the contribution of other enzymes present in the complex formulations that can limit refining performance (Fig. 2c). The purified endoglucanase EndoC reached the highest effect in terms of the degree of refining after 6000 revolutions with a value of 33.7°SR (Fig. 2b), while the endoglucanases EndoA (28.2°SR) and EndoB (29.0°SR) showed almost no improvement when compared to the reference without enzyme addition (27.7°SR), which is in accordance with their respective enzyme formulations. The obtained results are also in line with the prediction of enzyme behavior using carbohydrate binding modules, which were used to monitor changes in amorphous and crystalline regions upon enzyme treatment, resulting in EndoC and FiberCare R being the most effective enzymes on softwood pulp fibers [32]. However, the enzyme formulation EnzC achieved a higher degree of refining than its purified endoglucanase EndoC (32.7 vs. 31.6°SR after 6000 revolutions), which might be related to the presence of rather high xylanase (1.72 nkat mg−1), arabinosidase (0.0096 nkat mg−1) or LPMO activity (3,05 nkat mg−1) in this formulation. The purified endoglucanases resulted in a decrease of the air permeability on softwood pulp as well for some enzymes (Fig. 2d). The air permeability was least affected by EndoA and EndoB (both: 288 ml min−1), which showed no significant difference to the reference. However, using the endoglucanase EndoC yielded the highest effect (191 ml min−1). This suggests the formation of a higher paper density by EndoC, nonetheless a lower tensile index was achieved when compared to the reference (Additional file 1: Fig. S1b).
Comparison of the degree of refining results with reported hardwood pulp values (Fig. 2g) showed an even higher effect of EndoC on hardwood pulp compared to the respective EnzC formulation after 6000 PFI revolutions (45.0 vs. 47.8°SR).
However, on softwood pulp the formulation EnzC resulted in a slightly higher degree of refining value than its purified endoglucanase EndoC (32.7 vs. 31.6°SR). Nevertheless, EndoC still achieved a better refining performance on softwood pulp by decreasing the air permeability value to 187 ml min−1 compared to 222 ml min−1 by EnzC (Fig. 2d). This could be explained by additional enzymes such as LPMOs or ß-glucosidases in the complex enzyme formulation EnzC, which work synergistically in conjunction with endoglucanases, but result in the degradation and destabilization of cellulose, a process that is not beneficial for paper production. Interestingly, the purified endoglucanases EndoA and EndoB increased the degree of refining on hardwood pulp, although to a much lower extent than FiberCare R and EndoC, however, on softwood pulp no positive effects were achieved by these enzymes. Furthermore, the EndoA endoglucanase resulted in a higher degree of refining on hardwood pulp than its corresponding enzyme formulation EnzA (42.7°SR vs 40.7°SR), which might be related to additional enzymes in the formulation as well, thereby affecting the drainability of pulp negatively. Indeed, EnzA exhibited the highest arabinosidase activity at refining conditions, which could have led to degradation of stabilizing xylan in conjunction with xylanases.
For the investigation of synergistic enzyme effects on softwood pulp, β-glucosidase or cellobiohydrolase II (CBH II) enzymes were added individually to EndoC. Indeed, addition of β-glucosidase from Aspergillus niger increased the degree of refining value to 35.7°SR when compared to treatment with EndoC alone (31.6°SR) after 6000 PFI revolutions on softwood pulp (Fig. 2c). Similar results were achieved after addition of CBH II, leading to a degree of refining value of 34.7°SR.
However, treatment with only cellobiohydrolase II resulted in no effect and even led to lower the degree of refining value (26.4°SR) than the reference (Fig. 2c). This confirms the synergistic nature of this enzyme requiring endoglucanases to boost refining performance.
A similar observation was made in a previous study, showing no effect after CBH II treatment, with even a slight decrease in paper strength [17]. The air permeability according to Bendtsen was decreased significantly by either ß-glucosidase or cellobiohydrolase II addition on softwood pulp (Fig. 2d). The highest effect was achieved by addition of β-glucosidase to EndoC (132 ml min−1) after 6000 PFI revolutions, suggesting that the paper density was increased even further compared to the treatment using only EndoC endoglucanase (187 ml min−1), while the addition of CBH II improved the density only slightly (141 ml min−1). Analogous to the degree of refining, treatment with CBH II alone showed an even higher air permeability value (290 ml min−1) than the reference (262 ml min−1), suggesting a disruption of the cellulose structure.
Release of sugars from pulps at high enzyme dosage
Analysis of the sugar concentration at high enzyme dosage resulted in high levels of released glucose and xylose by most complex enzyme formulations, suggesting the degradation of surface layers by the combination of endoglucanases, xylanases, and β-glucosidases (Fig. 3). The improvement of paper smoothness by partial degradation of the surface layers of the fibers is also one of the main advantages of pulp refining. Scanning electron microscopy pictures of refined handsheets indeed suggest a higher smoothness of the cellulose fibers through fiber shortening (Additional file 1: Fig. S2–S5). However, excessive fiber degradation can decrease pulp strength. Therefore, the selection of the appropriate enzyme mixture is important to avoid excessive fiber degradation, which can be assessed through the release of glucose and xylose monomers cause by synergistic degradation of cellulose. [3, 11, 64]. The highest release of glucose was found for EnzC on both pulp types (hardwood: 2.98 mg ml−1, softwood: 2.81 mg ml−1). The commercial endoglucanase formulation FiberCare R also released low levels of glucose (hardwood: 0.44 mg ml−1, softwood: 0.52 mg ml−1), which might be related to a low level of residual β-glucosidase in this formulation. Interestingly, the release of glucose was higher on softwood pulp compared to hardwood pulp using a lower enzyme dosage of 0.83 nkat, as determined in a previous study [32]. This suggests a major influence of ß-glucosidases at higher enzyme dosages, which usually prevent product inhibition of endoglucanase enzymes by converting cellobiose to glucose [65]. This is beneficial for recycling processes, depending on the complete degradation of cellulose to glucose, but can be disadvantageous for processes that relay mainly on endoglucanases such as pulp refining, as a high amount of ß-glucosidases can cause excessive degradation of cellulose fibers. This highlights the importance of correct enzyme dosage and accurate prediction methods.

Glucose and xylose concentrations determined by HPLC in mg ml−1. Statistical significance was tested using a single factor ANOVA test with a significance level of 0.05 (significantly different results are indicated by the letter s, not significantly different results by the letter n)
While the glucose release was similar between hardwood and softwood pulp at high enzyme dosage, the pattern was different for xylose. The highest xylose concentration on hardwood pulp was caused by EnzC (3.14 mg ml−1) and EnzB (3.10 mg ml−1), while the xylose concentration was slightly lower after treatment with EnzA (2.12 mg ml−1). As expected, on softwood pulp the released xylose concentrations were much lower (highest for EnzC: 1.56 mg ml−1). The lower amount of released xylose from softwood can be related to lower xylan content in this pulp (6.6% compared to 12.2% present in hardwood pulp). Generally, hardwood pulp is composed of a higher xylan content, although it also depends on the used organism und pulping method [66, 67]. The high levels of glucose emphasize the need for wastewater treatments due to the conversion of cello-oligosaccharides to glucose by β-glucosidases due to machine malfunctions or wrong dosage during pulp refining. This is clearly avoided by formulations containing only endoglucanases, but may come with a cost increase as an additional purification step is required [68].
Interestingly, the high sugar release did correlate to the refining performance of EnzC, which showed the highest effect as well, however, this was not the case for all the other enzymes, as both EnzA and EnzB exhibited poor refining performance.
Similarly, FiberCare R with the lowest sugar release had a comparably high refining performance as EnzC. Therefore, the use of reducing sugar assays is not suitable for enzyme dosage in pulp refining. Nonetheless, analysis of the sugar release can give important information about synergistic enzyme effects, as glucose and xylose are released by the combined action of endoglucanases, ß-glucosidases, and xylanases.
The synergistic effect of CBH II was investigated through its addition to purified endoglucanase EndoC to mimic the conducted laboratory refining trials on softwood pulp. Indeed, treatment with only CBH II resulted in a low cellobiose formation (0.025 mg ml−1). Similarly, treatment of softwood pulp using only EndoC resulted in a low amount of cellobiose (0.019 mg ml−1). However, the combination of both enzymes synergistically enhanced the action of both enzymes, as the release of cellobiose from softwood increased considerably to 0.071 mg ml−1. Thereby, the addition of cellobiohydrolases can boost the performance of endoglucanases, which might benefit industrial refining or saccharification processes. Indeed, the refining performance was increased after addition of cellobiohydrolase II to EndoC endoglucanase. This synergistic effect is referred to as exo–endo synergism and it is proposed to be based on the action of endoglucanases on the cellulose fibers leading to the creation of new free ends that can be attacked by cellobiohydrolases [69]. A synergism between endoglucanases and cellobiohydrolases was reported for the degradation of cotton in the case of EG I and CBH II of Trichoderma reesei, resulting in an increased release of reducing sugars by combined action, which was not observed with individual CBH II treatment [70].
Endoglucanase activity assay after adsorption of derivatized cellopentaose CellG5 on pulp
Despite the use of the endoglucanase-specific derivatized cellopentaose (CellG5) substrate for enzyme dosage in pulp refining, minor differences between the enzyme formulations remained, especially if applied on different pulp types. Therefore, an adapted activity assay was developed by combining the CellG5 substrate with either softwood or hardwood pulp through the formation of hydrogen bonds between the derivatized cellopentaose substrate and pulp fibers. This could result in an enhanced activity assay that also takes pulp characteristics such as varying pulp morphology, accessible surface area or hemicelluloses into account, which are known to impact enzyme adsorption and therefore enzyme activity [71, 72].
For comparison of the adapted assay, the endoglucanase activity needed to be determined using the standard assay protocol at refining conditions (50% tap water + 50% deionized water, pH 7). The highest activities were observed with EnzB (741.22 nkat ml−1) and EnzC (581.33 nkat ml−1), while EnzA exhibited the lowest activity of 11.13 nkat ml−1 (Fig. 4a). Indeed, the behavior of the enzyme formulations was different after combination of the CellG5 substrate with softwood pulp (Fig. 4b). The highest activity was measured with EnzC (4.82 nkat ml−1), while the lowest activity was determined for EnzA as well (0.56 nkat ml−1). However, the activities of EnzB and FiberCare R were significantly lower after combination of CellG5 with softwood pulp (0.87 nkat ml−1 and 1.43 nkat ml−1, respectively). This would also fit to the results of laboratory refining trials on this pulp, with EnzC showing the highest degree of refining value after 6000 PFI revolutions (32.7°SR), while all the other enzyme formulations had a lower refining performance with FiberCare R being the second-best enzyme formulation (31.2°SR) and EnzB and EnzA having almost no effect (29.1 and 27.2°SR). The adapted enzyme assay would therefore indeed result in an improved correlation of enzyme activity with pulp refining outcome, although the CellG5 pulp combination did not entirely fit to EnzA, which showed a higher enzyme activity on softwood pulp as expected.

Comparison of the results of the standard CellG5 activity assay a with the results of the combination of the CellG5 substrate with softwood b or hardwood pulp c under refining conditions (50% tap water and 50% deionized water at pH 7.0). The obtained degree of refining values (°SR) after refining of softwood or hardwood pulp using 6000 PFI revolutions are indicated above each bar for comparison. Statistical significance was tested using a single factor ANOVA test with a significance level of 0.05 (significantly different results are indicated by the letter s, not significantly different results by the letter n)
The adapted assay was also tested on hardwood pulp (Fig. 4c). The highest activities were measured for EnzC (3.04 nkat ml−1) and FiberCare R (1.38 nkat ml−1) as well. This fits to the results of the refining trials, with both enzymes showing the highest performance (45.0 and 44.8°SR, respectively), although a higher activity of FiberCare R on the adapted assay was expected, considering the similar refining performance. However, the degree of refining is not the only factor for the assessment of refining performance. Indeed, the measured air permeability value of FiberCare R was higher than that of EnzC, thereby indicating a lower paper density (EnzC: 149 ml min−1, FiberCare R: 173 ml min−1 after 6000 PFI revolutions). EnzA and EnzB both exhibited the lowest refining effect and the lowest enzyme activities using the adapted CellG5 assay on hardwood pulp. Therefore, the use of the new adapted activity assay should translate to an improved enzyme dosage for refining especially for EnzB and thereby enhance energy savings by enabling the selection of the most efficient enzyme [30].
Despite the adapted assay not fitting entirely to the performance of EnzA, it could achieve improved correlation for all the other enzyme formulations. The discrepancy might be related to derivatized cellopentaose (CellG5) being bound to the pulp surface and therefore being accessible to all enzymes as part of a “background” activity. Moreover, the endoglucanase activity on softwood pulp was higher than on hardwood pulp, suggesting that indeed varying pulp characteristics between softwood and hardwood pulp are now considered by the adapted assay (Fig. 4b vs. c). Similar observations were conducted in a previous study after analysis of released sugars via HPLC from softwood and hardwood pulp upon enzyme treatment, resulting in a higher sugar release from softwood pulp at a lower enzyme dosage [32]. This could be explained by a generally higher binding of derivatized cellopentaose to softwood pulp, better accessibility to deeply bound CellG5 substrate or increased enzymatic separation of cellulose fibers with bound CellG5 from pulp surface, the latter would also fit to the increased sugar release measured by HPLC. A similar mechanism was proposed for cellulases that remove ink from wastepaper by hydrolyzation of cellulose fibers on the paper surface, leading to loosening of fibers with attached ink [73].
Conclusion
The characterization of commercial enzyme formulations revealed a varying enzyme composition. Identified enzymes include endoglucanases, β-glucosidases, xylanases and even arabinosidases. In case of the most effective enzyme formulation EnzC even LPMO activity was detected, the enzymes that are proposed to support endoglucanases in their function to attack cellulose. Knowledge about enzyme behavior is therefore essential for most efficient pulp refining. Indeed, endoglucanase activity was at the highest level at 55 °C and decreased considerably at 70 °C. Interestingly, the β-glucosidase activity increased at higher temperatures with EnzC even having the highest measured activity at 70 °C, which would result in enhanced glucose formation. In addition, laboratory refining on softwood pulp showed a diverging enzyme behavior compared to hardwood pulp. Enzymes exhibiting a high efficiency on hardwood pulp were also very active on softwood pulp, while enzymes with low efficiency on hardwood pulp showed almost no reaction on softwood pulp. This emphasizes the need for investigation of each pulp type despite the use of the endoglucanase specific derivatized cellopentaose substrate (CellG5) for enzyme dosage. Therefore, the CellG5 substrate was combined with either softwood or hardwood pulp, which indeed improved predictability, while simultaneously taking specific pulp characteristics such as enzyme accessibility into account. The adapted method thus offers the selection of the most effective enzymes for each pulp type, thereby maximizing energy savings in pulp refining.
Availability of data and materials
The data sets used in the current study are available from the corresponding author on reasonable request.
References
Przybysz P, Dubowik M, Małachowska E, Kucner M, Gajadhur M, Przybysz K (2020) Effect of refining intensity on the progress of internal fibrillation and shortening of cellulose fibers. BioResources 15:1482–1499
Wang X, Paulapuro H, Maloney TC (2005) Chemical pulp refining for optimum combination of dewatering and tensile strength. Nord Pulp Paper Res J 20:442–447. https://doi.org/10.3183/npprj-2005-20-04-p442-447
Reza H, Armin M, Artem EH, Fines SÁ, Fiber ÁDÁ (2019) Mechanisms of strength and stiffness improvement of paper after PFI refining with a focus on the effect of fines. Cellulose 26:4099–4124. https://doi.org/10.1007/s10570-019-02349-5
Singh R, Bhardwaj NK, Choudhury B (2015) Cellulase-assisted refining optimization for saving electrical energy demand and pulp quality evaluation. J Sci Ind Res (India) 74:471–475
Confederation of European Paper Industries (Cepi). Key Statistics 2020 European Pulp & Paper industry. 2021.
Haske-Cornelius O, Hartmann A, Brunner F, Pellis A, Bauer W, Nyanhongo GS et al (2020) Effects of enzymes on the refining of different pulps. J Biotechnol 320:1–10. https://doi.org/10.1016/j.jbiotec.2020.06.006
Fleiter T, Fehrenbach D, Worrell E, Eichhammer W (2012) Energy efficiency in the German pulp and paper industry—a model-based assessment of saving potentials. Energy 40:84–99. https://doi.org/10.1016/j.energy.2012.02.025
Tuveng TR, Jensen MS, Fredriksen L, Vaaje-Kolstad G, Eijsink VGH, Forsberg Z (2020) A thermostable bacterial lytic polysaccharide monooxygenase with high operational stability in a wide temperature range. Biotechnol Biofuels. https://doi.org/10.1186/s13068-020-01834-5
Obeng EM, Adam SNN, Budiman C, Ongkudon CM, Maas R, Jose J (2017) Lignocellulases: a review of emerging and developing enzymes, systems, and practices. Bioresour Bioprocess. https://doi.org/10.1186/s40643-017-0146-8
Song W, Han X, Qian Y, Liu G, Yao G, Zhong Y et al (2016) Proteomic analysis of the biomass hydrolytic potentials of Penicillium oxalicum lignocellulolytic enzyme system. Biotechnol Biofuels. https://doi.org/10.1186/s13068-016-0477-2
Buzala KP, Przybysz P, Kalinowska H, Derkowska M (2016) Effect of cellulases and xylanases on refining process and kraft pulp properties. PLoS ONE 11:1–14. https://doi.org/10.1371/journal.pone.0161575
Cui L, Meddeb-Mouelhi F, Laframboise F, Beauregard M (2015) Effect of commercial cellulases and refining on kraft pulp properties: correlations between treatment impacts and enzymatic activity components. Carbohydr Polym 115:193–199. https://doi.org/10.1016/j.carbpol.2014.08.076
Lecourt M, Meyer V, Sigoillot JC, Petit-Conil M (2010) Energy reduction of refining by cellulases. Holzforschung 64:441–446. https://doi.org/10.1515/HF.2010.066
Wang M, Du J, Zhang D, Li X, Zhao J (2017) Modification of different pulps by homologous overexpression alkali-tolerant endoglucanase in Bacillus subtilis Y106. Sci Rep. https://doi.org/10.1038/s41598-017-03215-9
Zhang Z, Wang M, Gao R, Yu X, Chen G (2017) Synergistic effect of thermostable β-glucosidase TN0602 and cellulase on cellulose hydrolysis. 3 Biotech. https://doi.org/10.1007/s13205-017-0672-2
Kamaya Y (1996) Role of endoglucanase in enzymatic modification of bleached kraft pulp. J Ferment Bioeng 82:549–553
Oksanen T, Pere J, Buchert J, Viikari L (1997) The effect of Trichoderma reesei cellulases and hemicellulases on the paper technical properties of never-dried bleached kraft pulp. Cellulose 4:329–339
Pere JLSSMGJVL. Use of purified enzymes in mechanical pulping. Tappi Pulping Conference, 1996, p. 693–9.
Villares A, Moreau C, Bennati-Granier C, Garajova S, Foucat L, Falourd X et al (2017) Lytic polysaccharide monooxygenases disrupt the cellulose fibers structure. Sci Rep 7:1–9. https://doi.org/10.1038/srep40262
Eibinger M, Sattelkow J, Ganner T, Plank H, Nidetzky B (2017) Single-molecule study of oxidative enzymatic deconstruction of cellulose. Nat Commun. https://doi.org/10.1038/s41467-017-01028-y
Saijonkari-Pahkala K (2001) Non-wood plants as raw material for pulp and paper. Agric Food Sci Finland. https://doi.org/10.23986/afsci.5707
Pirralho M, Flores D, Sousa VB, Quilhó T, Knapic S, Pereira H (2014) Evaluation on paper making potential of nine Eucalyptus species based on wood anatomical features. Ind Crops Prod 54:327–334. https://doi.org/10.1016/j.indcrop.2014.01.040
Barneto AG, Vila C, Ariza J (2011) Eucalyptus kraft pulp production: thermogravimetry monitoring. Thermochim Acta 520:110–120. https://doi.org/10.1016/j.tca.2011.03.027
Ainun ZMA, Muhammad KI, Rasmina H, Hazwani HA, Sharmiza A, Naziratulasikin AK et al (2018) Effect of chemical pretreatment on pulp and paper characteristics of bamboo Gigantochloa scorthechinii kraft fibers. IOP Conf Ser Mater Sci Eng. https://doi.org/10.1088/1757-899X/368/1/012044
Kamali M, Khodaparast Z (2015) Review on recent developments on pulp and paper mill wastewater treatment. Ecotoxicol Environ Saf 114:326–342. https://doi.org/10.1016/j.ecoenv.2014.05.005
Demuner IF, Colodette JL, Demuner AJ, Jardim CM (2019) Biorefinery review: wide-reaching products through kraft lignin. BioResources 14:7543–7581
Mboowa D (2021) A review of the traditional pulping methods and the recent improvements in the pulping processes. Biomass Convers Biorefin. https://doi.org/10.1007/s13399-020-01243-6
Llano T, García-Quevedo N, Quijorna N, Viguri JR, Coz A (2015) Evolution of lignocellulosic macrocomponents in the wastewater streams of a sulfite pulp mill: a preliminary biorefining approach. J Chem. https://doi.org/10.1155/2015/102534
Liu Z, Wang H, Hui L (2018) Pulping and papermaking of non-wood fibers. Pulp and Paper Processing, InTech; https://doi.org/10.5772/intechopen.79017.
Nagl M, Haske-Cornelius O, Skopek L, Pellis A, Bauer W, Nyanhongo GS et al (2021) Biorefining: the role of endoglucanases in refining of cellulose fibers. Cellulose 28:7633–7650. https://doi.org/10.1007/s10570-021-04022-2
Nagl M, Haske-Cornelius O, Bauer W, Csarman F, Ludwig R, Nyanhongo GS et al (2022) Towards a better understanding of synergistic enzyme effects during refining of cellulose fibers. Carbohydr Polym Technol App. https://doi.org/10.1016/j.carpta.2022.100223
Nagl M, Haske-Cornelius O, Skopek L, Bausch F, Pellis A, Bauer W et al (2022) Mechanistic investigation of the effect of endoglucanases related to pulp refining. Cellulose 29:2579–2598. https://doi.org/10.1007/s10570-021-04386-5
Mangan D, Cornaggia C, McKie V, Kargelis T, McCleary BV (2016) A novel automatable enzyme-coupled colorimetric assay for endo-1,4-β-glucanase (cellulase). Anal Bioanal Chem 408:4159–4168. https://doi.org/10.1007/s00216-016-9507-y
Parry NJ, Beever DE, Owen E, Vandenberghe I, van Beeumen J, Bhat MK (2001) Biochemical characterization and mechanism of action of a thermostable β-glucosidase purified from Thermoascus aurantiacus. Biochem J 353:117–127. https://doi.org/10.1042/bj3530117
Ghose TK (1987) Measurement of cellulase activities. Pure App Chem 59:257–268. https://doi.org/10.1351/pac198759020257
Mangan D, Cornaggia C, Liadova A, Mccormack N, Ivory R, Mckie VA et al (2017) Novel substrates for the automated and manual assay of endo-1, 4-b-xylanase. Carbohydr Res 445:14–22. https://doi.org/10.1016/j.carres.2017.02.009
Ichinose H, Fujimoto Z, Honda M, Harazono K, Nishimoto Y, Uzura A et al (2009) A β-L-arabinopyranosidase from Streptomyces avermitilis is a novel member of glycoside hydrolase family 27. J Biol Chem 284:25097–25105. https://doi.org/10.1074/jbc.M109.022723
Breslmayr E, Hanžek M, Hanrahan A, Leitner C, Kittl R, Šantek B et al (2018) A fast and sensitive activity assay for lytic polysaccharide monooxygenase. Biotechnol Biofuels. https://doi.org/10.1186/s13068-018-1063-6
Chokhawala HA, Roche CM, Kim TW, Atreya ME, Vegesna N, Dana CM et al (2015) Mutagenesis of Trichoderma reesei endoglucanase I: Impact of expression host on activity and stability at elevated temperatures. BMC Biotechnol. https://doi.org/10.1186/s12896-015-0118-z
Akcapinar GB, Venturini A, Martelli PL, Casadio R, Sezerman UO (2015) Modulating the thermostability of endoglucanase I from Trichoderma reesei using computational approaches. Protein Eng Des Sel 28:127–135. https://doi.org/10.1093/protein/gzv012
Jain KK, Kumar S, Bhardwaj KN, Kuhad RC (2018) Functional expression of a thermostable endoglucanase from Thermoascus aurantiacus RCKK in Pichia pastoris X-33 and its characterization. Mol Biotechnol 60:736–748. https://doi.org/10.1007/s12033-018-0106-3
Teugjas H, Väljamäe P (2013) Product inhibition of cellulases studied with 14C-labeled cellulose substrates. Biotechnol Biofuels. https://doi.org/10.1186/1754-6834-6-104
Zhang Q, Bao J (2017) Industrial cellulase performance in the simultaneous saccharification and co-fermentation (SSCF) of corn stover for high-titer ethanol production. Bioresour Bioprocess. https://doi.org/10.1186/s40643-017-0147-7
Coward-Kelly G, Aiello-Mazzari C, Kim S, Granda C, Holtzapple M (2003) Suggested improvements to the standard filter paper assay used to measure cellulase activity. Biotechnol Bioeng 82:745–749. https://doi.org/10.1002/bit.10620
Ghio S, Bradanini MB, Garrido MM, Ontañon OM, Piccinni FE, Marrero Diaz de Villegas R, et al (2020) Synergic activity of Cel8Pa β-1,4 endoglucanase and Bg1Pa β-glucosidase from Paenibacillus xylanivorans A59 in beta-glucan conversion. Biotechnol Rep 28:1–9. https://doi.org/10.1016/j.btre.2020.e00526.
Przybysz Buzała K, Kalinowska H, Borkowski J, Przybysz P (2018) Effect of xylanases on refining process and kraft pulp properties. Cellulose 25:1319–1328. https://doi.org/10.1007/s10570-017-1609-y
Tao W, Guo L, Meng A, Wang L, Ren H, Zhai H (2019) Effects of xylanase pretreatment on the quality of refiner mechanical mulberry branch fibers. Adv Polym Technol 2019:1–10. https://doi.org/10.1155/2019/6252013
Whitehead TR, Cotta MA (2001) Identification of a broad-specificity xylosidase/arabinosidase important for xylooligosaccharide fermentation by the ruminal anaerobe Selenomonas ruminantium GA192. Curr Microbiol 43:293–298. https://doi.org/10.1007/s002840010304
Kotake T, Yamanashi Y, Imaizumi C, Tsumuraya Y (2016) Metabolism of l-arabinose in plants. J Plant Res 129:781–792. https://doi.org/10.1007/s10265-016-0834-z
Li Q, Wu T, Zhao L, Pei J, Wang Z, Xiao W (2019) Highly efficient biotransformation of astragaloside IV to cycloastragenol by sugar-stimulated β-glucosidase and β-xylosidase from Dictyoglomus thermophilum. J Microbiol Biotechnol 29:1882–1893. https://doi.org/10.4014/jmb.1807.07020
Ahmed S, Luis AS, Bras JLA, Ghosh A, Gautam S, Gupta MN et al (2013) A novel α-L-arabinofuranosidase of family 43 glycoside hydrolase (Ct43Araf) from Clostridium thermocellum. PLoS ONE. https://doi.org/10.1371/journal.pone.0073575
Wohlert M, Benselfelt T, Wågberg L, Furó I, Berglund LA, Wohlert J (2022) Cellulose and the role of hydrogen bonds: not in charge of everything. Cellulose. https://doi.org/10.1007/s10570-021-04325-4
Bosmans TJ, Stépán AM, Toriz G, Renneckar S, Karabulut E, Wågberg L et al (2014) Assembly of debranched xylan from solution and on nanocellulosic surfaces. Biomacromol 15:924–930. https://doi.org/10.1021/bm4017868
Kabel MA, van den Borne H, Vincken JP, Voragen AGJ, Schols HA (2007) Structural differences of xylans affect their interaction with cellulose. Carbohydr Polym 69:94–105. https://doi.org/10.1016/j.carbpol.2006.09.006
Ogunyewo OA, Randhawa A, Gupta M, Kaladhar VC, Verma PK, Yazdani SS (2020) Synergistic action of a lytic polysaccharide monooxygenase and a cellobiohydrolase from Penicillium funiculosum in cellulose saccharification under high-level substrate loading. Appl Environ Microbiol 86:1–21. https://doi.org/10.1128/AEM.01769-20
Stagno V, Ricci S, Longo S, Verticchio E, Frasca F, Siani MA et al (2022) Discrimination between softwood and hardwood based on hemicellulose content obtained with portable nuclear magnetic resonance. Cellulose 29:7917–7934
Krysztof M, Olejnik K, Kulpinski P, Erdman A, Sasiadek E (2021) A comparative study of the effect of cellulose-based deep coating and pulp refining on the structural and mechanical properties of paper. BioResources 16:5376–5389. https://doi.org/10.15376/biores.16.3.5376-5389
Hobisch MA, Zabler S, Bardet SM, Zankel A, Nypelö T, Eckhart R et al (2021) How cellulose nanofibrils and cellulose microparticles impact paper strength—a visualization approach. Carbohydr Polym. https://doi.org/10.1016/j.carbpol.2020.117406
Shackford LD (2003) A Comparison of Pulping and Bleaching of Kraft Softwood and Eucalyptus Pulps. 36th International Pulp and Paper Congress and Exhibition, São Paulo
Vermaas JV, Petridis L, Qi X, Schulz R, Lindner B, Smith JC (2015) Mechanism of lignin inhibition of enzymatic biomass deconstruction. Biotechnol Biofuels. https://doi.org/10.1186/s13068-015-0379-8
Rowell RM (2012) Cell wall chemistry. Handbook of wood chemistry and wood composites, 2nd edn, CRC Press; p 75–98. https://doi.org/10.1201/b12487.
Afsahi G, Rojalin T, Vuorinen T (2019) Chemical characteristics and stability of eucalyptus kraft pulps bleached with tertiary amine catalyzed hypochlorous acid. Cellulose 26:2047–2054. https://doi.org/10.1007/s10570-018-2172-x
Aksenov AS, Tyshkunova I, Poshina DN, Guryanova AA, Chukhchin DG, Sinelnikov IG et al (2020) Biocatalysis of industrial kraft pulps: similarities and differences between hardwood and softwood pulps in hydrolysis by enzyme complex of penicillium verruculosum. Catalysts. https://doi.org/10.3390/catal10050536
Žnidaršič-Plazl P, Rutar V, Ravnjak D (2009) The effect of enzymatic treatments of pulps on fiber and paper properties. Chem Biochem Eng 23:497–506
Singh G, Verma AK, Kumar V (2016) Catalytic properties, functional attributes and industrial applications of β-glucosidases. 3 Biotech 6:1–14. https://doi.org/10.1007/s13205-015-0328-z
Ahmad Z, Dajani WW, Paleologou M, Xu C (2020) Sustainable process for the depolymerization/oxidation of softwood and hardwood kraft lignins using hydrogen peroxide under ambient conditions. Molecules. https://doi.org/10.3390/molecules25102329
Surup GR, Trubetskaya A, Tangstad M (2020) Charcoal as an alternative reductant in ferroalloy production: a review. Processes 8:1–41. https://doi.org/10.3390/pr8111432
Sakhuja D, Ghai H, Rathour RK, Kumar P, Bhatt AK, Bhatia RK (2021) Cost-effective production of biocatalysts using inexpensive plant biomass: a review. 3 Biotech. https://doi.org/10.1007/s13205-021-02847-z
Lemos MA, Teixeira JA, Domingues MRM, Mota M, Gama FM (2003) The enhancement of the cellulolytic activity of cellobiohydrolase I and endoglucanase by the addition of cellulose binding domains derived from Trichoderma reesei. Enzyme Microb Technol 32:35–40
Kleman-Leyer KM, Siika-Aho M, Teeri TT, Kirk TK (1996) The cellulases endoglucanase I and cellobiohydrolase II of Trichoderma reesei act synergistically to solubilize native cotton cellulose but not to decrease its molecular size. Appl Environ Microbiol 62:2883–2887
Baig KS (2020) Interaction of enzymes with lignocellulosic materials: causes, mechanism and influencing factors. Bioresour Bioprocess. https://doi.org/10.1186/s40643-020-00310-0
Siqueira G, Arantes V, Saddler JN, Ferraz A, Milagres AMF (2017) Limitation of cellulose accessibility and unproductive binding of cellulases by pretreated sugarcane bagasse lignin. Biotechnol Biofuels. https://doi.org/10.1186/s13068-017-0860-7
Verma N, Bansal MC, Kumar V (2011) Enzymatic deinking with cellulases: a review. J Solid Waste Technol Manag 37:297–306. https://doi.org/10.5276/JSWTM.2011.297
Acknowledgements
The authors are grateful to the participating companies of the pulp and paper industry of Austria for their scientific contribution. In addition, the authors would like to thank the BioMatInt doctoral school for their support.
Funding
Open access funding provided by University of Natural Resources and Life Sciences Vienna (BOKU). This work was supported by the Austrian Research Promotion Agency (FFG) within the scope of the FibreZyme Project and the Government of Lower Austria.
Author information
Authors and Affiliations
Contributions
MN: conceptualization, methodology, investigation, data curation, formal analysis, writing—original draft, visualization, funding acquisition. OH-C: conceptualization, methodology, project administration. Funding acquisition. GSN: project administration, funding acquisition, supervision. GMG and WB: conceptualization, methodology, formal analysis, writing—review and editing, supervision. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1:
Tensile index data of refined softwood pulp. Cellobiohydrolase II activity results. Scanning electron microscopy pictures of refined paper handsets.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Nagl, M., Haske-Cornelius, O., Bauer, W. et al. Enhanced energy savings in enzymatic refining of hardwood and softwood pulp. Energ Sustain Soc 13, 19 (2023). https://doi.org/10.1186/s13705-023-00398-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13705-023-00398-0