
Abstract
Background
Staphylococcus aureus (S. aureus) is a pathogen responsible for a wide range of clinical manifestations and potentially fatal conditions. There is a paucity of information on the influence of androgens in the immune response to S. aureus infection. In this study, we evaluated the influence of the hormone 5α-dihydrotestosterone (DHT) on mouse peritoneal macrophages (MPMs) and human peripheral blood monocytes (HPBMs) induced by S. aureus.
Methods
An in vitro model of MPMs from BALB/c sham males, orchiectomised (OQX) males, and females was used. Cells were inoculated with 10 μL of S. aureus, phage-type 80 or sterile saline (control) for 6 h. The MPMs of OQX males and females were pre-treated with 100 μL of 10–2 M DHT for 24 h before inoculation with S. aureus. The concentration of the cytokines TNF-α, IL-1α, IL-6, IL-8, and IL-10; total nitrites (NO−2); and hydrogen peroxide (H2O2) were measured in the supernatant of MPM cultures. In addition, the toll-like receptor 2 (TLR2) and nuclear factor kappa B (NF-kB) genes that are involved in immune responses were analysed. For the in vitro model of HPBMs, nine men and nine women of childbearing age were selected and HPBMs were isolated from samples of the volunteers’ peripheral blood. In women, blood was collected during the periovulatory period. The HPBMs were inoculated with S. aureus for 6 h and the supernatant was collected for the analysis of cytokines TNF-α, IL-6, IL-12; and GM-CSF, NO−2, and H2O2. The HPBMs were then removed for the analysis of 84 genes involved in the host’s response to bacterial infections by RT-PCR array. GraphPad was used for statistical analysis with a p value < 0.05.
Results
Our data demonstrated that MPMs from sham males inoculated with S. aureus displayed higher concentrations of inflammatory cytokines and lower concentrations of IL-10, NO−2, and H2O2 when compared with MPMs from OQX males and females. A similar result was observed in the HPBMs of men when compared with those of women. Previous treatment with DHT in women HPBMs increased the production of pro-inflammatory cytokines and decreased the levels of IL-10, NO−2, and H2O2. The analysis of gene expression showed that DHT increased the activity of the TLR2 and NF-kB pathways in both MPMs and HPBMs.
Conclusions
We found that DHT acts as an inflammatory modulator in the monocyte/macrophage response induced by S. aureus and females exhibit a better immune defence response against this pathogen.
Highlights
-
A sex difference in the immune response to S. aureus was observed.
-
DHT stimulated a greater release of inflammatory cytokines.
-
DHT promoted the activation of genes involved in this response.
-
DHT suppressed the release of inflammatory markers.
Similar content being viewed by others


Introduction
Staphylococcus aureus (S. aureus) is a pathogen responsible for a variety of clinical manifestations ranging from relatively benign skin infections to potentially life-threatening conditions, such as endocarditis and osteomyelitis. S. aureus is also a commensal bacterium that colonises approximately 30% of the human population, with a higher risk of complications and mortality in men [1,2,3]. According to some studies, the sex of an individual serves as a contributory factor in the incidence and progression of disorders associated with the immune response. Accumulated evidence shows that sex can also play an important role in susceptibility to infectious diseases. In general, females generate more robust humoral and cell-mediated immune responses after exposure to an antigen when compared with males. Furthermore, males frequently mount heightened inflammatory immune responses to microbial stimuli, thereby worsening disease prognosis [4, 5].
In general, while oestrogen has an immune-stimulating effect, testosterone has an immunosuppressive effect, thereby increasing susceptibility to infection and colonisation by S. aureus [6,7,8,9]. Men are more susceptible to sepsis, acute respiratory distress, and multiple organ failure following soft tissue traumatic haemorrhagic shock and thermal injury, in part as a result of immune suppression and abnormal neutrophil activation [10]. A correlation exists between the parasitic load, strength of immune responses, and levels of sex hormones in men, and it has been observed that both low parasitic loads and weakened immune responses are caused by the immunosuppressive effects of testosterone on immune cells [11].
Women also display reduced infection rates for a variety of bacteria, viruses, and parasites, such as Helicobacter pylori [12], Mycobacterium tuberculosis [13], hepatitis B virus [14], and Aspergillus fumigatus [5]. In addition, an analysis of COVID-19 virus-related deaths among 17 million adults showed that females were more protected against severe COVID-19 infection and mortality. Males have a 1.59 times greater risk ratio of developing virus-related complications than females [15]. However, the molecular mechanisms underlying sexually dimorphic immune responses are not fully understood.
Sex hormones such as oestrogen and testosterone have been reported to modulate the differentiation, maturation, lifespan, and effector functions of innate immune cells, including neutrophils, macrophages, natural killer (NK) cells, and dendritic cells [5]. Macrophages perform several functions in the immune system. They facilitate the elimination of invading pathogens by phagocytosis, production of reactive oxygen/nitrogen species, and release of several inflammatory mediators, such as cytokines. These cytokines are fundamental to the initiation and propagation of the inflammatory process to contain pathogens and promote homeostatic regulation [4, 16]. There are few studies regarding the role of testosterone in S. aureus infection of macrophages.
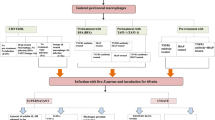
To better understand the role of androgens in the immune response against S. aureus, this study assessed the effects of 5α-dihydrotestosterone (DHT) on the profile of cytokines, inflammatory mediators as NO−2 and H2O2, genes associated with this response to S. aureus mediated by murine peritoneal macrophages (MPMs), and human peripheral blood monocytes (HPBMs). There is limited information concerning the role of DHT in the response to infections caused by S. aureus. Despite the impressive advances in many medical fields, the pathophysiology, diagnostic procedures, and appropriate therapeutic interventions for S. aureus infections are still a matter of debate [17,18,19]. The model stimulated by S. aureus developed in this study was based on the studies cited.
Materials and methods
Microorganisms
The phage-type used in this study was Staphylococcus aureus 80 from the microbiology laboratory of the Federal University of Bahia, Campos Anísio Teixeira, Vitória da Conquista-Ba. This is the same strain used in previous studies [20, 21] and is derived from the nasal swabs of children attending day care centres in the municipality. Brain heart infusion (BHI) and mannitol salt agar were used to activate, culture, and subculture S. aureus strains for subsequent inoculation of MPMs and HPBMs. In addition, Gram staining, catalase and coagulase tests, and PCR (nuc gene) were performed to confirm the identity and purity of the bacterial strain. The inoculum of S. aureus was obtained by direct suspension, removing 3–5 colonies (same morphological type) from the plates containing the reference strain. The colonies were then transferred to tubes containing sterile saline solution, vortexed, and analysed using a spectrophotometer to obtain the following parameters: absorbance of 0.135 at 660 nm that is equivalent to 1 × 108 colony forming units (CFU) [22].
In vitro model of MPMs
Animals
Six male and three female specific pathogen-free (SPF) Balb/c mice aged between 6 and 8 weeks were used. The mice used in the in vitro experiments were obtained from the Animal Laboratory of the Multidisciplinary Centre for Biological Research of the State University of Campinas (CEMIB/UNICAMP-SP) and kept under controlled conditions of light (light from 7 am to 7 pm) and temperature (23 ± 3 °C), with free access to water and food in the animal facility of the Multidisciplinary Institute of Health at the Federal University of Bahia (IMS/UFBA).
Orchiectomy and sham surgery
The animals underwent surgery to remove the gonads (orchiectomy) to significantly decrease their levels of sex hormones. Following administration of anaesthesia, the orchiectomy or simulated surgery was performed using a trans scrotal approach. A 1-cm midline scrotal incision was made, through which the testes and epididymis were exposed and removed. The incision was then sutured after the resection of both testes. For sham mice, the testes were visualised but not removed [24]. The sham surgery was performed as the control [25]. The animals were anesthetised with ketamine and xylazine at doses of 5 and 50 mg/kg, respectively. After surgery, the animals received prophylactic treatment with 2.5% of the antibiotic enrofloxacin in a single dose of 2.5 mg/kg [22]. OQX animals remained under observation for 14 days to confirm the reduction of testosterone levels from the gonads using enzyme-linked immunosorbent assay (ELISA) (Cayman Chemical’s ACE™, Ann Arbor, MI, USA). After this period, macrophage recruitment was induced in the peritoneal cavity of OQX and sham males.
Females
The oestrous cycles of females were monitored by vaginal lavage, which was visualised on a slide under an optical microscope. When females were in diestrum, intraperitoneal macrophages were stimulated and proestrum, the phase of the cycle with the highest levels of 17b-estradiol (E2), was confirmed after 72 h. No surgery was performed (ovariectomy) in females. Females were used to assess the effect of sex on the immune response of HPBMs to S. aureus.
Isolation of male and female MPMs
To induce macrophages recruitment into the peritoneal cavity, 2 mL of thioglycolate medium (3%) was injected into the peritoneal cavity of OQX (n = 3), sham (n = 3), and female (n = 3) mice for increasing monocyte migration into the peritoneum, thereby increasing macrophages yield. Peritoneal macrophages are more mature after isolation and more stable in their functions and phenotype [23]. The animals were euthanized after 3 days by decapitation with the aid of a guillotine. Cells were isolated from the peritoneal lavage and cultured in RPMI 1640 medium (Gibco™) supplemented with 10% foetal bovine serum (FBS) (Sigma Aldrich) and ciprofloxacin (20 µg/mL) (Sigma Aldrich). The MPM cell suspension (2 × 105/mL) was seeded in 24-well plates [24].
Inoculation of MPMs with S. aureus or sterile saline solution and treatment with DHT
MPMs from male sham, male OQX, and female mice were cultured with 10 μL of the inoculum of S. aureus (108 CFU) for 6 h at 37 °C in 5% CO2 and 95% humidified air [24]. Part of the cells were pre-treated with 100 μL of 10–2 M DHT for 24 h at 37 °C before inoculation with S. aureus or addition of 10 μL of sterile saline solution. Male and female MPMs were plated as follows: cells with saline solution (control) (6 h); Cells with S. aureus (6 h); Cells pre-treated with DHT (24 h) and then infected with S. aureus (6 h). DHT was used in place of testosterone, since it is the active androgen that is not converted to E2 [25]. The DHT concentration used was determined by following the manufacturer's instructions (Sigma Aldrich Brazil LTDA-5α-Dihydrotestosterone-D3 (16, 16, 17-D3) Solution). For dose selection, we conducted a pilot study based on different doses (DHT 10–1 M–10–9 M—data not shown) and the dose of choice was 10–2 M. The incubation time was similar to that of other studies using macrophages cell cultures treated in vitro with testosterone [26, 27].
Dosage of cytokines
The levels of cytokines TNF-α, IL-1α, IL-6, IL-8, and IL-10 were measured in the supernatant of MPM cell cultures by ELISA using the commercial ProcartaPlex™ Simplex Kit Invitrogen (Thermo Fisher, São Paulo, Brazil). The assay was performed following the manufacturer’s instructions and the cytokine levels are expressed in absolute values (pg/mL). All groups were analysed in triplicate.
Gene expression analysis of receptors and transcription factors by RT-qPCR
Cells were detached from the wells using trypsin and stored at -80 °C for gene expression analysis. The relative gene expressions of TLR2 and NF-κB were determined. Gene expression was analysed by RT-qPCR arrays. The gene expression assay was performed on plates with the NF-κB and TLR2 target genes using the SYBR® PCR Master Mix (Applied Biosystems™). Amplification was performed using the StepOnePlus thermal cycler with the following cycles: 50 °C for 10 min, 95 °C for 10 min, 45 cycles of 95 °C for 15 s, and 60 °C for 1 min. Data were analysed using the comparative method (2ΔΔCt) and normalised based on GAPDH gene expression according to the manufacturer’s instructions.
In vitro model of HPBMs
Selection of study participants
Peripheral blood samples were obtained from men and women aged 18–25 years, who volunteered to be included in the study. Volunteers were screened using questionnaires, which were provided. Information on the participants’ date of birth, sex, clinical history, and lifestyle habits, such as alcohol use or smoking, and medication use, was collected. The inclusion criteria were: (1) healthy, young volunteers who had not used antibiotics, antifungals, or anti-inflammatory drugs in the last 90 days and (2) women of childbearing age with a regular menstrual cycle in the fertile period of their menstrual cycle. The exclusion criteria were: (1) women who used hormonal contraceptives, (2) breastfeeding, pregnant, and puerperal women, (3) volunteers who consume alcohol or use tobacco, (4) volunteers who had performed physical activity 24 h prior to blood collection, and (5) volunteers with human immunodeficiency virus (HIV) infection or other infections and immune inflammatory diseases including diabetes, cardiopathies, psychiatric diseases, cancer, and kidney disease, and volunteers presenting with clinical signs and symptoms, such as fever, sore throat, skin rash, allergy, or flu. This study was approved by the Ethics Committee in Research on Human Beings (CEP) of the same institution (CAAE:95158218.3.0000.5556). The study met ethical requirements by complying with the guidelines of Resolution 466/12.
Sampling procedure
Phlebotomies for the collection of peripheral blood samples were performed by qualified professionals using sterile materials and personal protective equipment (PPE). All blood samples were kept at 6–10 °C, and monocytes were isolated within 2 h of blood collection. A portion of the collected blood was used to determine the levels of hormones, including testosterone, E2, progesterone, follicle-stimulating hormone (FSH), and luteinizing hormone (LH). Furthermore, the general health status of the individual was evaluated by measuring glucose, total serum cholesterol, glutamic–oxaloacetic transaminase (GOT), glutamic pyruvic transaminase (GPT), and complete blood count. For women, sample collection dates were determined based on their menstrual cycle.
Isolation of HPBMs
Peripheral blood mononuclear cells (PBMC) were extracted from the blood using Ficoll, a hydrophilic polysaccharide density gradient medium to separate the blood into its components. Once PBMCs were isolated, it was possible to separate the monocytes from lymphocytes using techniques based on different properties of cells, such as their unique size and density compared to lymphocytes, and their ability to adhere to glass or plastic. Owing to the risk of lymphocyte contamination, Percoll, a hyperosmolar percolator gradient (density = 1064 g/mL), was used. Peripheral blood was mixed with PBS in a 1:1 ratio. PBMCs were separated by Ficoll column centrifugation (400 × g for 20 min), washed, and centrifuged twice with 1 × PBS at 100 × g for 10 min. Subsequently, the mononuclear cells were separated by Percoll column centrifugation at 400 × g for 35 min and resuspended in RPMI 1640 medium containing 10% foetal bovine serum, 2 mM glutamine, 10 mM HEPES, and 20 μl/mL ciprofloxacin. After the evaluation of viability and adequate confluence (2 × 105/mL), the cells were plated in 24-well polystyrene plates and incubated for 24 h at 37 °C with 5% CO2 before inoculation with S. aureus [28]. This procedure allowed us to obtain a homogeneous population of HPBMs, which appeared as adherent cells [24].
Inoculation of HPBMs with S. aureus or sterile saline solution and pre-treatment with DHT
HPBMs were grown using 10 μl of S. aureus inoculum (CFU 108) for 6 h in 5% CO2 and 95% humidified air at 37 °C [24]. Additional cells in triplicate were pre-treated with 100 μl DHT (10–2 M) for 24 h at 37 °C before stimulation with S. aureus or 10 μl of sterile saline solution. The HPBMs were divided into plaques: cells with S. aureus (6 h), cells with sterile saline solution (control) (6 h), and cells pre-treated with DHT (24 h), and subsequently infected with S. aureus (6 h).
Cytokine concentrations of HPBMs
To determine cytokine concentrations, the cell culture supernatants were obtained from the culture plate wells and stored at – 80 °C. The concentrations of TNF-α, IL-12 and GM-CSF were determined using Luminex xMAP® with the Procartaplex Immunoassay kit (Affymetrix eBioscience) according to the manufacturer’s instructions.
Gene expression analysis of HPBMs
With the aid of trypsin, the HPBMs were removed from the cell culture wells and stored in RNAlater at − 80 °C for gene expression. The expression of inflammatory markers was evaluated using quantitative reverse transcription PCR (RT-qPCR). The mRNA was extracted using TRIzol® LS (Life Technologies™) following the manufacturer’s protocol. The cDNA was obtained from mRNA by retro-transcription (RT), using the SuperScript® III Reverse Transcriptase kit with additional complementary oligonucleotides to the mRNA poly-A tail (Oligo Dt) and RNAse inhibitor. The obtained cDNA was subjected to analysis using the Human Innate and Adaptive Immune Responses PCR Array (Qiagen-SABioscience) for the evaluation of genes involved in the response of the host’s monocytes to bacterial infection. The qPCR array kit was used to analyse the expression of 84 genes involved in the host response to bacterial infection, including genes related to TLR signalling, and genes involved in the acute phase response, complement system activation, inflammatory response, and acquired immune response against bacteria. All procedures and data analyses were performed in accordance with the manufacturer’s instructions and software.
Inflammatory markers
Measurement of NO−2 concentration in MPMs and HPBMs
Nitric oxide (NO) levels were quantified using the Griess assay. Briefly, 50 µL aliquots of the test solution (standards and experimental samples) were added to 0.1% Griess reagent solution (50 µL of 3.9 mM N-(1-naphthyl)ethylenediamine in 5% (v/v) phosphoric acid) and incubated in the dark at room temperature for 10 min. Sulphanilamide solution (1% in phosphoric acid) was added to the mixture, and after 10 min of incubation the absorbance of the coloured product was measured at 540 nm using a microplate reader. Standard sodium nitrate solutions (1.65–100 mM) were used to construct a standard curve to determine the concentrations of the samples [29]. Since nitric oxide is a difficult gas to detect, the concentration of nitrites was determined by indirect measurement of the levels of total nitrites by this reaction in a 96-well plate.
Measurement of H2O2 concentration in MPMs and HPBMs
For determining H2O2 concentration, an aliquot of each cell culture supernatant was added to a 96-well plate, in triplicates, using the Amplex® Red Hydrogen Peroxide/Peroxidase (Invitrogen) kit, according to the manufacturers’ instructions. The absorbance was measured at 560 nm using a spectrophotometer [30].
Statistical analysis
Statistical analyses were performed using the Graphpad-Prism 5.0 (Graphpad Software, San Diego, CA-USA). The results were expressed as mean ± standard deviation (SD). The Shapiro Wilk test was used to evaluate data normality. To determine statistical differences in values between two different groups with normal distribution and variance homogeneity the Student’s T test was used; otherwise, the Mann Whitney test was used. Statistical differences were considered significant at p < 0.05, using a 95% confidence interval.
Results
In vitro model of MPMs
MPMs from sham males express more inflammatory cytokines (TNF-α, IL-1α, IL-8) than those from OQX males, while those from females express more IL-10
MPMs from sham males inoculated with S. aureus exhibited higher concentrations of inflammatory cytokines TNF-α, (Fig. 1A1) IL-1α (Fig. 1B1), IL-6 (Fig. 1C1), and IL-8 (Fig. 1D1) when compared with MPMs from females. However, those of females expressed higher IL-10 (Fig. 1E1).

TNF-α (A1), IL-1α (B1), IL-6 (C1), IL-8 (D1), IL-10 (E1) concentrations in the supernatant of MPMs from sham males, OQX males and females. MPMs inoculated with 10 µL of S. aureus [6 h (108 CFU)]. TNF-α (A2), IL-1α (B2), IL-6 (C2), IL-8 (D2), IL-10 (E2) concentrations in the supernatant of MPMs from sham males, OQX males and females: pre-treated with 100 µL of DHT [24 h (10–2 M)] and inoculated with 10 µL of S. aureus [6 h (108 CFU)]. Data are expressed as mean ± SDM. *p < 0.05, **p < 0.01, ***p < 0.001
DHT increases the expression of inflammatory cytokines and decreases levels of IL-10 in MPMs of OQX males and females
MPMs from OQX males and females inoculated with S. aureus and pre-treated with DHT exhibited a significant increase in the production of inflammatory cytokines TNF-α (Fig. 1A2), IL-1α (Fig. 1B2), IL-6 (Fig. 1C2), and IL-8 (Fig. 1D2) when compared with untreated MPMs from OQX males and females. However, DHT decreased IL-10 levels in MPMs from OQX males and females (Fig. 1E2) inoculated with S. aureus.
Concentration of NO−2 and H2O2 in MPMs from sham males, OQX males, and females
MPMs from sham males inoculated with S. aureus exhibited lower concentrations of NO−2 (Fig. 2A1) and H2O2 (Fig. 2B1) when compared with MPMs from OQX males and females. MPMs from OQX males and females inoculated with S. aureus and pre-treated with DHT exhibited a significant reduction in NO−2 (Fig. 2A2) and H2O2 (Fig. 2B2) production when compared with untreated MPMs from OQX males and females.

Total nitrite- NO−2 (A) concentration in the supernatant of MPMs from sham males, OQX males, and females. (A1): Sham males, OQX males and females: inoculated with 10 µL of S. aureus [6 h (108 CFU)]. A2 Sham males, OQX males, and females: pre-treated with 100 µL of DHT [24 h (10–2 M)] and inoculated with 10 µL of S. aureus [6 h (108 CFU)]. H2O2 (B) concentration in the supernatant of MPMs from sham males, OQX males, and females. B1 Sham males, OQX males, and females: inoculated with 10 µL of S. aureus [6 h (108 CFU)]. B2 Sham males, OQX males, and females: pre-treated with 100 µL of DHT [24 h (10–2 M)] and inoculated with 10 µL of S. aureus [6 h (108 CFU)]. Data are expressed as mean ± SDM. *p < 0.05, **p < 0.01, ***p < 0.001
Relative gene expressions of TLR2 and NF-κB in MPMs from sham males, OQX males, and females inoculated with S. aureus and pre-treated with DHT
No differences were observed in TLR2 and NF-κB expression between MPMs from sham and OQX males. However, MPMs from sham males exhibited higher TLR2 (Fig. 3A1) and NF-κB (Fig. 3B1) expression levels when compared with MPMs from females. MPMs from OQX males pre-treated with DHT exhibited a significant increase in TLR2 (Fig. 3A2) and NF-κB (Fig. 3B2) expression. However, no significant differences were observed in expression of these genes in MPMs from females pre-treated with DHT.

Relative expression of TLR2 (A) of MPMs from sham males, OQX males, and females. A1 Sham males, OQX males, and females: inoculated with 10 µL of S. aureus [6 h (108 CFU)]; A2 Sham males, OQX males, and females: pre-treated with 100 µL of DHT [24 h (10–2 M)] and inoculated with 10 µL of S. aureus [6 h (108 CFU)]. Relative expression of NF-κB (B) of MPMs from sham males, OQX males, and females. B1 Sham males, OQX males, and females: inoculated with 10 µL of S. aureus [6 h (108 CFU)]; (B2): Sham males, OQX males, and females: pre-treated with 100 µL of DHT [24 h (10–2 M)] and inoculated with 10 µL of S. aureus [6 h (108 CFU)]. Data are expressed as mean ± SDM. *p < 0.05, **p < 0.01, ***p < 0.001
In vitro model of HPBMs
HPBMs from men express more inflammatory cytokines than HPBMs of women, while those from women express more IL-10
The secretion of TNF-α (Fig. 4A1), IL-6 (Fig. 4B1), IL-12 (Fig. 4C1), and GM-CSF (Fig. 4D1) in HPBMs of men was significantly higher than that in women. However, the secretion of IL-10 (Fig. 4E1) in HPBMs of men was significantly lower than that in women.

TNF-α (A1), IL-6 (B1), IL-10 (C1), IL-12 (D1), GM-CSF (E1) concentrations in the supernatant of HPBMs from men and women. HPBMs inoculated with 10 µL of S. aureus [6 h (108 CFU)]. TNF-α (A2), IL-6 (B2), IL-10 (C2), IL-12 (D2), GM-CSF (E2) concentrations in the supernatant of HPBMs from men and women: pre-treated with 100 µL of DHT [24 h (10–2 M)] and inoculated with 10 µL of S. aureus [6 h (108 CFU)]. Data are expressed as mean ± SDM. *p < 0.05, **p < 0.01, ***p < 0.001
DHT increases the secretion of cytokines TNF-α, IL-12, and GM-CSF in HPBMs of women
The HPBMs of women pre-treated with DHT exhibited a significant increase in the secretion of TNF-α (Fig. 4A2), IL-12 (Fig. 4C2), and GM-CSF (Fig. 4D2) when compared with the HPBMs of women. However, the HPBMs of women pre-treated with DHT exhibited a significantly reduced secretion of IL-10 when compared with the HPBMs of women (Fig. 4E2).
Total NO−2 and H2O2 concentrations in HPBMs
The HPBMs of men produced significantly less NO−2 (Fig. 5A1) and H2O2 (Fig. 5B1) than women. Reduced production of NO−2 (Fig. 5A2) and H2O2 (Fig. 5B2) was observed in HPBMs of women pre-treated with DHT when compared with women.

Total nitrite (A) concentration in the supernatant of HPBMs from men and women. A1 HPBMs inoculated with 10 µL of S. aureus [6 h (108 CFU)]. A2 HPBMs pre-treated with 100 µL of DHT [24 h (10–2 M)] and inoculated with 10 µL of S. aureus [6 h (108 CFU)]. H2O2 (B) concentration in the supernatant of HPBMs from men and women. B1 HPBMs inoculated with 10 µL of S. aureus [6 h (108 CFU)]. B2 HPBMs pre-treated with 100 µL of DHT [24 h (10–2 M)] and inoculated with 10 µL of S. aureus [6 h (108 CFU)]. Data are expressed as mean ± SDM. *p < 0.05, **p < 0.01, ***p < 0.001
Comparison of the gene expression of HPBMs of men and women inoculated with S. aureus or sterile saline
Among the 84 genes analysed, a statistically significant upregulation of 25 genes, i.e., BTK, CD80, CLEC4E, CSF2, EIF2AK2, ELK1, HRAS, IL1A, IL2, IL6, CXCL8, IRF3, LTA, LY86, MAP2K4, NKKB1, NFKBIA, NFRKB, NR2C2, RELA, SIGIRR, TLR3, TLR8, TNF, and UBE2N, was observed HPBMs from men and women inoculated with S. aureus when compared to HPBMs inoculated with sterile saline (p < 0.05) (Fig. 6A).

Analysis of 84 genes involved in the host response to bacterial infections. A HPBMs inoculated with S. aureus when compared to HPBMs inoculated with sterile saline of men and women. B HPBMs inoculated with S. aureus from men when compared to HPBMs from women. C HPBMs inoculated with S. aureus from DHT-treated women compared to HPBMs from Women. Analysis performed with the Human Innate and Adaptive Immune Responses PCR Array kit (Qiagen—SABioscience). Upregulated genes (green). Data are expressed as mean ± DPM. *p < 0.05 (non-parametric Mann–Whitney one-tailed test)
Comparison of the gene expressions of HPBMs of men and women
Among the 84 genes analysed, a statistically significant upregulation of 22 genes, i.e., CSF3, CXCL10, FOS, HRAS, IL12A, IL1A, IRAK1, IRAK4, JUN, LTA, LY96, NFKB1, NFKB2, NFKBIA, NR2C2, RIPK2, SARM1, SIGIRR, TAB1, TLR2, TLR8, and UBE2N, was observed HPBMs inoculated with S. aureus from men and women (p < 0.05) (Fig. 6B).
Comparison of the gene expressions of HPBMs of women and those of women treated with DHT
Among the 84 genes analysed, a statistically significant upregulation of 19 genes, i.e., BTK, CD80, EIF2AK2, IL12A, IL1A, IL1B, IL6, IRAK1, IRAK4, IRF3, JUN, LTA, NFKB2, NFKBIA, NR2C2, TLR2, TLR3, TLR8, and TNF, was observed in the women HPBMs inoculated with S. aureus and pre-treated with DHT when compared to HPBMs from women (p < 0.05) (Fig. 6C).
Discussion
The results of the present study provide evidence in support of the inflammatory effects of DHT in innate immunity. Using MPMs from male sham, male OQX, and female mice, and HPBMs from men and women healthy donors, we have shown that MPMs from male and HPBMs from men have higher expression and production of inflammatory cytokines than those from females and women respectively. This response is related to the action of the hormone testosterone. Supporting our hypothesis, a group of males castrated (orchiectomised-OQX) displayed lower concentrations of TNF-α, IL-1α, and IL-8 when compared with Sham males. When comparing sham males and females, sham males displayed higher concentrations of TNF-α, IL-1α, IL-6, and IL-8 and lower concentrations of IL-10. Similar results were observed with HPBMs; men displayed higher concentrations of TNF-α, IL-6, IL-12, and GM-CSF and lower concentrations of IL-10 when compared with women. A study in humans by Schröder et al. [31] showed that men display higher concentrations of TNF and IL-6 and women produce higher levels of anti-inflammatory cytokines and IL-10. This cytokine, as has been documented, plays an important beneficial role in suppressing inflammatory responses and improving the inflammatory profile in severe infections [32, 33]. A study by Von Aulock et al. [34] showed that the blood of men produced significantly more TNF-α, IL-1β, IL-6, and IL-8 than that of women in response to a high concentration of lipopolysaccharides (LPS) or lipoteichoic acid (LTA), corroborating with our results.
Our study showed that hormonal treatment with DHT increased the production of TNF-α, IL-1α, IL-6, and IL-8 and decreased the production of IL-10 in OQX males. In corroborating these findings, HPBMs from women pre-treated with DHT displayed higher concentrations of TNF, IL-12, and GM-CSF and a lower concentration of IL-10. As reported, Th2 responses are usually activated and high levels of interleukins IL-4, IL-5, and IL-10 are usually produced in women in response to infections [31, 35]. Conversely, men predominantly display Th1 responses and overproduce TNF-α, IL-1β, IL-2, IL-6, and IL-8, which in turn is often associated with poor outcomes, such as septicaemia and bacteremia [36, 37]. Matalka et al. [38] observed that IL-10 levels were associated with the presence of oestrogen in women and were a protective factor. Since the concentration of 17-β estradiol (E2) increased, reaching concentrations corresponding to those found during pregnancy, IL-10 levels increased significantly. IL-10 is also released into the circulation during human sepsis and controls the production of TNF, IL-1, and IL-8 in vitro [36]. In this case, it is an advantage for the females.
Another salient finding of this study is that the MPMs of sham animals displayed lower concentrations of NO−2 and H2O2 when compared with OQX males. The orchiectomy can thus increase the release of free radicals by MPM-associated responses by S. aureus. In contrast, OQX males showed lower concentrations of these markers in cases of pre-treatment with DHT. When comparing genders, MPMs from sham males showed lower concentrations of NO−2 and H2O2 when compared with MPMs from females. We found similar results in HPBMs; men displayed lower concentrations of NO−2 and H2O2 when compared with women and, in relation to pre-treatment with DHT, the women's cells displayed lower concentrations of NO−2 and H2O2.
Although testosterone has pro-oxidant properties, as shown by Azevedo et al. [37], this hormone inhibited the free radical-mediated response in our study. This effect of steroid hormones is, in part performed through the transcriptional regulation of certain enzymes involved in free radical metabolism [38]. Androgens are also known to have a role in inhibiting the activity of the NADPH enzyme complex, an important system as a microbicidal, free radical generating mechanism [37].
A study by Scotland et al. [39] found that the antibacterial activity of intracellular NADPH oxidase was significantly elevated in resident peritoneal macrophages in females when compared to males. Females should respond better to infection and show greater control of inflammatory mediators, such as free radicals. A study by Azevedo et al. [37] found that males produced lower levels of antioxidant enzymes in resident intraperitoneal macrophages when compared with females. Our hypothesis is that such an antioxidant response is harmful in the defence against S. aureus infections. Aerobic bacteria such as S. aureus develop antioxidant enzymes that inhibit free radical production by macrophages and prevent bacterial clearance and efficient defence responses of the immune system. S. aureus uses several strategies for its survival and proliferation in the host organism, such as complement opsonisation, neutralising phagocytosis, and inhibiting both humoral and cellular immune responses. Even the catalase enzyme produced by S. aureus converts H2O2, which would have a toxic effect on the bacteria, into oxygen and water [40]. In this case, both the dose and the inflammation model influenced the release of these markers [41,42,43]. A study by Xiao et al. [26] found that treatment with testosterone reduced the concentration of H2O2 by modulating the testosterone AR receptor in animal cardiac cells. Damage reduction was testosterone dose-dependent via the NF-κB pathway.
Considering the NF-κB pathway, our study did not find significant differences in TLR2 and NF-κB expression between MPMs from sham and OQX males infected with S. aureus. However, pre-treatment with DHT resulted in an increase in TLR-2 and NF-κB expression in MPMs from OQX males. One hypothesis for these findings is that the exogenous hormone may act through different pathways compared to the biological hormone. It is known that androgens can act via both the cellular and/or nuclear hormone receptor [44, 45] and directly on specific membrane-binding sites (SMBS). Testosterone, particularly exogenous testosterone, may have a non-genomic action via SMBS, an action not mediated by classical AR, but rather through the activation of membrane kinase signalling via the ERK and PK pathways, resulting in a series of biological activities dependent on this pathway, including expression of NF-κB, RelA (p65), Caspase-3, serine threonine kinase (Akt), extracellular signal-regulated kinase (ERK) 1/2, Bcl-2 and Bcl-xL, and protein kinases (PK) [20, 46].
In addition, our study showed that MPMs from sham males display higher expression levels of TLR2 and NF-κB when compared to those of females. In this case, the strong expression of these receptors provides a more uncontrolled response to S. aureus infection. An imbalance of TLR2, NF-κB, and pro- and anti-inflammatory cytokines usually results in an exacerbated inflammatory response, multiple organ dysfunction syndrome, and septic shock in humans [47, 48]. The analysis of the response in MPMs from females pre-treated with DHT showed no significant differences in TLR2 and NF-κB gene expression. However, this response in females is likely a consequence of the protective role of oestrogen by modulating the expression of TLR2 and NF-κB and providing greater balance in the release of cytokines and free radicals [49, 50]. Other studies pre-treated monocytes with E2 in vitro for 24 h before stimulation with LPS and found that NF-κB activation was suppressed in macrophages [51, 52]. These findings suggest that E2 exerts anti-inflammatory effects by modulating the response of receptors and cytokines [53], which can partially account for the sex differences in severe infections [31]. A study by Souza et al. [24] found that E2 reduced the production of TNF-α and decreased the expression of TLR2 and NF-κB in macrophages from mice infected with S. aureus. In our results, the hormone DHT had the opposite effect. The stimuli of expression of CD80 and CD86 probably promotes the expressions of TLR2 and transcription factors and DHT acts by increasing the expression of TLR2 and the transcription factors IRAK1, IRAK4, IKKβ and NFκB. Several extracellular stimuli, such as pro-inflammatory cytokines (IL-1, interleukin 1; TNF, tumor necrosis factor), cause phosphorylation (P) of IkB, which can be mediated by IkB kinase (IKK) and the TAB1 protein, known to mediate several intracellular signaling pathways, such as those induced by interleukin-1. Phosphorylation of IkB releases NF-kB which can act on a target gene in the nucleus, while IkB is degraded. However, there is a limitation in our trials to conclude this claim. New assays are needed to directly quantify the activation of the NF-kB pathway.
CD80/CD86 signalling contributes to the pro-inflammatory response of S. aureus. This pathway is associated with cytokine release and macrophages activation [54]. T cells are activated through the interaction of peptides presented by the major histocompatibility complex (MHC), which interact with the T cell receptor complex, and lead to the initiation of several signalling cascades. This signalling complex is also induced through several co-receptors and their respective ligands. The CD28 T cell co-receptor interacts with CD80 (B7-1) and CD86 (B7-2) that are expressed on activated antigen-presenting cells in response to pathogens, such as S. aureus, which leads to the induction of several signalling pathways, such as those controlled by NF-κB, MAPK, PI3K, and AKT [55, 56]. In a study by Parker [54], CD80 and CD86-deficient mice displayed significant reductions in several pro-inflammatory cytokines and significantly improved survival rates in a murine model of pneumonia. This need for CD80 and CD86 in the pro-inflammatory response to S. aureus was evident early in the infection and in vitro using bone-marrow-derived macrophages. The work supported the hypothesis that much of the morbidity and mortality associated with S. aureus infection is the result of excessive production of cytokines. This brings us to the idea that DHT stimulates a more robust response of inflammatory cytokines, promoting a more intense response to S. aureus infection. Our results showed higher expression of genes such as CD80, TNF, IL1A, and IL6 in HPBMs inoculated with S. aureus compared to in HPBMs inoculated with sterile saline (control). In addition to this response, we observed higher expressions of IL12A and TLR2 in HPBMs from men inoculated with S. aureus when compared to HPBMs from women. Corroborating our results, HPBMs of men displayed higher expression of TLR2 when compared with women. Our study also showed genes associated with upregulation in HPBMs inoculated with S. aureus from women treated with DHT compared to in HPBMs from women. We observed that DHT canupregulate genes that are involved in monocyte functions, such as the BTK gene [57], in addition to genes that encode proteins that phosphorylate the NF-κB pathway, such as: CD80, TNF, IL12A, IL1A, IL1B, IL6, NFKB2, NFKBIA, and TLR2. S. aureus can activate monocytes, whose activation is necessary for the initiation and modulation of the immune response, mainly through the transcription of genes related to the NF-κB pathway, consequently leading to the production/secretion of inflammatory pathway mediators and pro-inflammatory [58, 59] cytokines, such as TNF-α, IL-12, and IL6. Regardless of the stimulus, there seems to be a participation of ROS (oxidative stress) and an increase in intracellular calcium for the activation of NF-kB [60]. In a study by Yamamoto et al. [61] using cell culture, it was confirmed that LPS induced expression of the co-stimulatory molecules CD80 and CD86 macrophages. NF-kB is a master regulator of immunity and inflammation, and controls adhesion molecules, chemokines, growth factors, inflammatory enzymes, and pro-inflammatory. NF-kB is a heterodimer consisting of two subunits: p65 (also called RelA) and p50 [62, 63]. In the literature, the term authentic NF-kB designates the p50/RelA combination. In addition to these, other subunits have been described, such as c-Rel, RelB, and p52, and it is likely that different types of combinations are capable of activating different genes or blocking the transcription of p50/RelA [64]. In our results, we observed higher expression of RelA in HPBMs inoculated with S. aureus when compared to in those of the control. As given above, this study proposes that DHT stimulates the NF-κB pathway in the monocyte immune response induced by S. aureus, as outlined in Fig. 7.

Effect of DHT on the immune response induced by S. aureus. DHT acts by increasing the expression of TLR2 and that of transcription factors IRAK1, IRAK4, IKKβ, and NFκB (represented by red arrows). Several extracellular stimuli, such as pro-inflammatory cytokines (IL-1, interleukin 1; TNF, tumour necrosis factor), cause IkB phosphorylation (P), which can be mediated by IkB kinase (IKK) and mediated by the TAB1 protein, known to mediate several intracellular signaling pathways, such as those induced by interleukin-1. The IkB phosphorylation releases NFκB which acts on a target gene in the nucleus, whereas IkB is degraded
Perspectives and significance
The data show that females produce lower (potentially harmful) inflammatory cytokines, but produce more products to increase anti-inflammatory cytokines (IL-10). Although our model of study involved monocytes, we emphasize that these cells are applicable to what is seen in all diseases for sex differences, not just those associated with S. aureus infection. One of the most interesting results of our study is the data showing that TLR-2 increases with testosterone in men but not in women. This indicates that testosterone enhances pathogen recognition and is, in part the mechanism for the increase in dysregulated inflammation. Certainly, a protective response in HPBMs of women compared to men, and clearly show that the differences are related to testosterone. Thus, further studies are required to better understand these mechanisms of action of DHT in this inflammatory response pathway. We intend to analyze the profiles of other cells of the immune system in treatment with DHT in the future. It is expected that a better understanding of the role of testosterone in the body’s defense response against microorganisms may propose strategies for an effective, safe, and useful treatment. In addition, it may contribute to actions in promoting the population’s health, resulting in the reduction of costs with treatments, hospitalizations, complications, and improving the quality of life of people with this condition.
Availability of data and materials
The data sets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- BHI:
-
Brain heart infusion
- CAAE:
-
Certificado de Apresentação para Apreciação Ética (Certificate of Submission for Ethical Consideration)
- CFU:
-
Colony Forming Units
- DHT:
-
Dihydrotestosterone
- E2:
-
17β-Oestradiol
- ELISA:
-
Enzyme-linked immunosorbent assay
- FBS:
-
Foetal bovine serum
- GAPDH:
-
Glyceraldehyde 3-phosphate dehydrogenase
- GM-CSF:
-
Granulocyte and macrophage stimulating factor
- HPBMs:
-
Human peripheral blood monocytes
- H2O2 :
-
Hydrogen peroxide
- IFN-γ:
-
Interferon-gamma
- IL-1α:
-
Interleukin 1 alpha
- IL-6:
-
Interleukin 6
- IL-8:
-
Interleukin 8
- IL-10:
-
Interleukin 10
- IL-12:
-
Interleukin 12
- JNK:
-
C-Jun N-terminal kinase
- MAPK:
-
Mitogen-activated protein kinase
- MHC:
-
Major histocompatibility complex
- MPMs:
-
Mouse peritoneal macrophages
- NF-κB:
-
Nuclear factor kappa B
- NFKBIA:
-
NFKB inhibitor alpha
- NO− 2 :
-
Total nitrites
- OQX:
-
Orchiectomised males
- P38:
-
Mitogen-activated protein kinases
- ROS:
-
Reactive oxygen species
- RPMI:
-
Roswell Park Memorial Institute (culture medium)
- RT-qPCR:
-
Reverse transcription quantitative PCR
- SAPK:
-
Interacting protein
- SHAM:
-
Males submitted to simulated surgery (sham surgery)
- SPF:
-
Specific-pathogen-free
- TLR2:
-
Toll-like receptor 2
- TNF-α:
-
Tumour necrosis factor alpha
References
Souli M, et al. Changing characteristics of Staphylococcus aureus bacteremia: results from a 21-Year, prospective, longitudinal study. Clin Infect Dis. 2019;13:1868–77. https://doi.org/10.1093/cid/ciz112.
Tong SY, et al. Staphylococcus aureus infections: epidemiology, pathophysiology, clinical manifestations, and management. Clin Microbiol Rev. 2015;28:603–61. https://doi.org/10.1128/CMR.00134-14.
Mansur N, et al. Does sex affect 30-day mortality in Staphylococcus aureus bacteremia? Gend Med. 2012;9:463–70. https://doi.org/10.1016/j.genm.2012.10.009.
Marriott I, Huet-Hudson YM. Sexual dimorphism in innate immune responses to infectious organisms. Immunol Res. 2006;34:177–92. https://doi.org/10.1385/IR:34:3:177.
Jaillon S, Berthenet K, Garlanda C. Sexual dimorphism in innate immunity. Clin Rev Allergy Immunol. 2019;56:308–21. https://doi.org/10.1007/s12016-017-8648-x.
Taneja V. Sex hormones determine immune response. Front Immunol. 2018;27:1931. https://doi.org/10.3389/fimmu.2018.01931.
Angele MK, Pratschke S, Hubbard WJ, Chaudry IH. Gender differences in sepsis: cardiovascular and immunological aspects. Virulence. 2014;5:12–9. https://doi.org/10.4161/viru.26982.
Olsen K, et al. Staphylococcus aureus nasal carriage is associated with serum 25-hydroxyvitamin D levels, gender and smoking status. The Tromsø Staph and Skin Study. Eur J Clin Microbiol Infect Dis. 2012;31:465–73. https://doi.org/10.1007/s10096-011-1331-x.
Sollid JU, Furberg AS, Hanssen AM, Johannessen M. Staphylococcus aureus: determinants of human carriage. Infect Genet Evol. 2014;21:531–41. https://doi.org/10.1016/j.meegid.2013.03.020.
Sheth SU, et al. Testosterone depletion or blockade in male rats protects against trauma hemorrhagic shock-induced distant organ injury by limiting gut injury and subsequent production of biologically active mesenteric lymph. J Trauma. 2011;71:1652–8. https://doi.org/10.1097/TA.0b013e31823a06ea.
Folstad I, Karter AJ. Parasites, bright males, and the immunocompetence. Handicap. 2015;139:603–22. https://doi.org/10.1086/285346.
Ibrahim A, et al. Sex-differences in the prevalence of Helicobacter pylori infection in pediatric and adult populations: systematic review and meta-analysis of 244 studies. Dig Liver Dis. 2017;49:742–9. https://doi.org/10.1016/j.dld.2017.03.019.
Neyrolles O, Quintana-Murci L. Sexual inequality in tuberculosis. PLoS Med. 2009;6(12):e1000199. https://doi.org/10.1371/journal.pmed.1000199.
Baig S. Gender disparity in infections of Hepatitis B virus. J Coll Physicians Surg Pak. 2009;19:598–600.
Williamson EJ, et al. Factors associated with COVID-19-related death using OpenSAFELY. Nature. 2020;584:430–6. https://doi.org/10.1038/s41586-020-2521-4.
Dimitrijević M, et al. The influence of aging and estradiol to progesterone ratio on rat macrophage phenotypic profile and NO and TNF-α production. Exp Gerontol. 2013;48:1243–54. https://doi.org/10.1016/j.exger.2013.07.001.
Guo Y, Song G, Sun M, Wang J, Wang Y. Prevalence and therapies of antibiotic-resistance in Staphylococcus aureus. Front Cell Infect Microbiol. 2020;10:107. https://doi.org/10.3389/fcimb.2020.00107.
Mehta Y, et al. Methicillin-resistant Staphylococcus aureus in intensive care unit setting of India: a review of clinical burden, patterns of prevalence, preventive measures, and future strategies. Indian J Crit Care Med. 2020;24:55–62. https://doi.org/10.5005/jp-journals-10071-23337.
Reddy P, Srirama K, Dirisala V. An update on clinical burden, diagnostic tools, and therapeutic options of Staphylococcus aureus. Infect Dis (Auckl). 2017. https://doi.org/10.1177/1179916117703999.
de Almeida JB, de Carvalho SP, da Silva L. Molecular characterization of methicillin-resistant and methicillin-sensitive Staphylococcus aureus isolates from human milk samples in Brazil. Braz J Microbiol. 2020;51:1813–7. https://doi.org/10.1007/s42770-020-00367-1.
de Carvalho SP, et al. Genomic profile of Brazilian methicillin-resistant Staphylococcus aureus resembles clones dispersed worldwide. J Med Microbiol. 2019;68:693–702. https://doi.org/10.1099/jmm.0.000956.
de Oliveira THC, et al. Sepsis induced by Staphylococcus aureus: participation of biomarkers in a murine model. Med Sci Monit. 2015;21:345–55. https://doi.org/10.12659/msm.892528.
Layoun A, Samba M, Santos M. Isolation of murine peritoneal macrophages to carry out gene expression analysis upon toll-like receptors stimulation. J Vis Exp. 2015. https://doi.org/10.3791/52749.
Souza CLSE, et al. Ovarian hormones influence immune response to Staphylococcus aureus infection. Braz J Infect Dis. 2020;24:534–44. https://doi.org/10.1016/j.bjid.2020.10.004.
Durrani F, et al. Gonadal hormones and oxidative stress interaction differentially affects survival of male and female mice after lung Klebsiella pneumoniae infection. Exp Lung Res. 2012;38:165–72. https://doi.org/10.3109/01902148.2011.654045.
Xiao FY, Nheu L, Komesaroff P, Ling S. Testosterone protects cardiac myocytes from superoxide injury via NF-κB signalling pathways. Life Sci. 2015;133:45–52. https://doi.org/10.1016/j.lfs.2015.05.009.
Rettew JA, Huet-hudson YM, Marriott I. Testosterone reduces macrophage expression in the mouse of toll-like receptor 4, a trigger for inflammation and innate immunity. Biol Reprod. 2008;78:432–7. https://doi.org/10.1095/biolreprod.107.063545.
Campesi I, et al. Oral contraceptives modify DNA methylation and monocyte-derived macrophage function. Biol Sex Differ. 2012. https://doi.org/10.1186/2042-6410-3-4.
Green LC, et al. Analysis of nitrate, nitrite, and [15N] nitrate in biological fluids. Anal Biochem. 1982;126:131–8. https://doi.org/10.1016/0003-2697(82)90118-X.
das Neves Selis N, et al. Gardnerella vaginalis and Neisseria gonorrhoeae are effectively inhibited by lactobacilli with probiotic properties isolated from Brazilian Cupuaçu (Theobroma grandiflorum) Fruit. BioMed Res Int. 2021;2021:6626249. https://doi.org/10.1155/2021/6626249.
Schröder J, Kahlke V, Staubach K, Zabel P, Stüber F. Gender differences in human sepsis. Arch Surg. 1998;133:1200–5. https://doi.org/10.1001/archsurg.133.11.1200.
Fournier B, Philpott D. Recognition of Staphylococcus aureus by the innate immune system. Clin Microbiol Rev. 2005;18:521–40. https://doi.org/10.1128/cmr.18.3.521-540.2005.
Lee M, et al. JQ1, a BET inhibitor, controls TLR4-induced IL-10 production in regulatory B cells by BRD4-NF-κB axis. BMB Rep. 2017;50:640–6. https://doi.org/10.5483/bmbrep.2017.50.12.194.
Aulock S, et al. Gender difference in cytokine secretion on immune stimulation with LPS and LTA. J Interferon Cytokine Res. 2006;26:887–92. https://doi.org/10.1089/jir.2006.26.887.
Marriott I, Bost K, Huet-Hudson Y. Sexual dimorphism in expression of receptors for bacterial lipopolysaccharides in murine macrophages: a possible mechanism for gender-based differences in endotoxic shock susceptibility. J Reprod Immunol. 2006;71:12–27. https://doi.org/10.1016/j.jri.2006.01.004.
Blackwell T, Christman J. Sepsis and cytokines: current status. Br J Anaesth. 1996;77:110–7. https://doi.org/10.1093/bja/77.1.110.
Azevedo RB, et al. Regulation of antioxidant enzyme activities in male and female rat macrophages by sex steroids. Braz J Med Biol Res. 2001;34:683–7. https://doi.org/10.1590/s0100-879x2001000500018.
Adler I, et al. The effect of certain steroid hormones on the expression of genes involved in the metabolism of free radicals. Gynecol Endocrinol. 2012;28:912–6. https://doi.org/10.3109/09513590.2012.683067.
Scotland RS, Stables MJ, Madalli S, Watson P, Gilroy DW. Sex differences in resident immune cell phenotype underlie more efficient acute inflammatory responses in female mice. Blood. 2011;118:5918–27. https://doi.org/10.1182/blood-2011-03-340281.
Santos ALD, et al. Staphylococcus aureus: visiting a strain of clinical importance. J Bras Patol Med Lab. 2007;43:413–23. https://doi.org/10.1590/S1676-24442007000600005.
Lee GT, et al. Dihydrotestosterone increases cytotoxic activity of macrophages on prostate cancer cells via trail. Endocrinology. 2019;160:2049–60. https://doi.org/10.1210/en.2019-00367.
Kajihara T, et al. Androgen signaling in decidualizing human endometrial stromal cells enhances resistance to oxidative stress. Fertil Steril. 2012;97:185–91. https://doi.org/10.1016/j.fertnstert.2011.10.017.
Lee SH, Heo JS, Lee MY, Han HJ. Effect of dihydrotestosterone on hydrogen peroxide-induced apoptosis of mouse embryonic stem cells. J Cell Physiol. 2008;216:269–75. https://doi.org/10.1002/jcp.21402.
Lai JJ, et al. Androgen receptor influences on body defense system via modulation of innate and adaptive immune systems: lessons from conditional AR knockout mice. Am J Pathol. 2012;181:1504–12. https://doi.org/10.1016/j.ajpath.2012.07.008.
Ruszkiewicz JA, et al. Sex-specific differences in redox homeostasis in brain norm and disease. J Mol Neurosci. 2019;67:312–42. https://doi.org/10.1007/s12031-018-1241-9.
Shepherd R, Cheung A, Pang K, Saffery R, Novakovic B. Sexual dimorphism in innate immunity: the role of sex hormones and epigenetics. Front Immunol. 2021. https://doi.org/10.3389/fimmu.2020.604000.
Kim NH, et al. Toll-like receptor 2 downregulation and cytokine dysregulation predict mortality in patients with Staphylococcus aureus bacteremia. BMC Infect Dis. 2020;20:1–10. https://doi.org/10.1186/s12879-020-05641-z.
Yimin, et al. Contribution of toll-like receptor 2 to the innate response against Staphylococcus aureus infection in mice. PLoS ONE. 2013;8:e74287. https://doi.org/10.1371/journal.pone.0074287.
Chandrasekaran VRM, Periasamy S, Liu LL, Liu MY. 17β-Estradiol protects against acetaminophen-overdose-induced acute oxidative hepatic damage and increases the survival rate in mice. Steroids. 2011;76:118–24. https://doi.org/10.1016/j.steroids.2010.09.008.
Wagner AH, Schroeter MR, Hecker M. 17β-Estradiol inhibition of NADPH oxidase expression in human endothelial cells. FASEB J. 2001;15:2121–30. https://doi.org/10.1096/fj.01-0123.
Murphy AJ, Guyre PM, Pioli PA. Estradiol suppresses NF-κB activation through coordinated regulation of let-7a and miR-125b in primary human macrophages. J Immunol. 2010;184:5029–37. https://doi.org/10.4049/jimmunol.0903463.
Pioli PA, et al. Estradiol attenuates lipopolysaccharide-induced CXC chemokine ligand 8 production by human peripheral blood monocytes. J Immunol. 2007;179:6284–90. https://doi.org/10.4049/jimmunol.179.9.6284.
Aomatsu M, Kato T, Kasahara E, Kitagawa S. Gender difference in tumor necrosis factor-α production in human neutrophils stimulated by lipopolysaccharide and interferon-γ. Biochem Biophys Res Commun. 2013;441:220–5. https://doi.org/10.1016/j.bbrc.2013.10.042.
Parker D. CD80/CD86 signaling contributes to the pro-inflammatory response of Staphylococcus aureus in the airways. Cytokine. 2018;107:130–6. https://doi.org/10.1016/j.cyto.2018.01.016.
Chen L, Flies DB. Molecular mechanisms of co-stimulation and co-inhibition of T cells. Nat Rev Immunol. 2013;13(4):227–42. https://doi.org/10.1038/nri3405.
Yao S, Zhu Y, Chen L. Advances in targeting cell surface signalling molecules for immune modulation. Nat Rev Drug Discov. 2013;12(2):130–46. https://doi.org/10.1038/nrd3877.
Cavaliere FM, et al. The lack of BTK does not impair monocytes and polymorphonuclear cells functions in Xlinked agammaglobulinemia under treatment with intravenous immunoglobulin replacement. PLoS ONE. 2017;12(4):1–21. https://doi.org/10.1371/journal.pone.0175961.
Giannoni E, et al. Estradiol and progesterone strongly inhibit the innate immune response of mononuclear cells in newborns. Infect Immun. 2011;79(7):2690–8. https://doi.org/10.1128/IAI.00076-11.
Pelekanou V, et al. Estrogen anti-inflammatory activity on human monocytes is mediated through cross-talk between estrogen receptor ERa36 and GPR30/GPER1. J Leukoc Biol. 2016;99(2):333–47. https://doi.org/10.1189/jlb.3A0914-430RR.
Glezer I, et al. The role of the transcription factor NF-kappaB in the molecular mechanisms of action of psychoactive drugs. Braz J Psychiatry. 2000;22(1):26–30. https://doi.org/10.1590/S1516-44462000000100008.
Yamamoto S, et al. Haloperidol suppresses NF-kappaB to inhibit lipopolysaccharide-induced pro-inflammatory response in RAW 264 cells. Med Sci Monit. 2016;4(22):367–72. https://doi.org/10.12659/msm.895739.
Baeuerle PA, Baltimore D. NF-kB: ten years after. Cell. 1996;87(1):13–20. https://doi.org/10.1016/s0092-8674(00)81318-5.
Siebenlist U. NF-kB/IkB proteins. Their role in cell growth, differentiation and development. Biochim Biophys Acta. 1997;1332(1):R7–13. https://doi.org/10.1016/s0304-419x(96)00038-8.
Ghosh S, May MJ, Kopp EB. NF-kB and rel proteins: evolutionary conserved mediators of immune responses. Annu Rev Immunol. 1998;16(1):225–60. https://doi.org/10.1146/annurev.immunol.16.1.225.
Acknowledgements
Jim Hesson proofread the manuscript (https://www.academicenglishsolutions.com).
Funding
This study was supported by Foundation for Research Support of the State of Bahia Fapesb n. 3055/2018, cota 2018, BOLO 755/2018 and Higher Education Personnel Improvement Coordination (Code 001). The funding agency played no role in the design nor conclusions of the study.
Author information
Authors and Affiliations
Contributions
Conceived and designed the experiments: DCS, RSB, CLSS and LMM. Orchiectomy: SIS, IDA and DCS. Collection, isolation and culture of MPMs and HPBMs: DCS, RSDMD, IDA, LSCP, CFA, LLBM, TCS and MNSJ. Cytokine dosage: DCS, RSDMD, RSB, BCBG. Dosage of nitrites and hydrogen peroxide: DCS, TMLC, LSCP, IDA. Gene expression: DCS, GBC and LMM. Analysed the data: DCS, RSB and LMM. Contributed reagents/materials/analysis tools: DCS, RSB, TJS, RY, GBC and LMM. Wrote the paper: DCS and LMM. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Ethics approval and consent were deemed unnecessary in this research, according to the Animal Ethics Committee (AEC) and approved by the Human Ethics Committee (Protocol-95158218300005556) of the Multidisciplinary Health Institute, Federal University of Bahia. The strain used was obtained during other studies after approval by the Ethics Committee of Research with Human Beings of the Multidisciplinary Health Institute Campus Anísio Teixeira under CAAE nº08731912.5.0000.5556 (nasal strain). Informed consent was obtained from a parent or guardian. All methods were performed in accordance with the relevant guidelines and regulations.
Consent to publication
Not applicable.
Competing interests
The authors declare that there are no competing interests regarding the publication of this paper.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Dos Santos, D.C., de Souza Bittencout, R., Arêas, I.D. et al. Effects of 5α-dihydrotestosterone on the modulation of monocyte/macrophage response to Staphylococcus aureus: an in vitro study. Biol Sex Differ 14, 15 (2023). https://doi.org/10.1186/s13293-023-00501-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13293-023-00501-2