
Abstract
Background
While a connection has been established between serum interleukin-6 (IL-6) levels and the IL-6 gene (− 174G/C) polymorphism in allergic diseases such as asthma, its specific association with severe asthma remains unexplored. This study examined the relationship between the IL-6 (− 174G/C) gene polymorphism and mild and severe asthma, focusing on its influence on type 2 inflammation.
Methods
Our study comprised 98 patients with mild asthma and 116 with severe asthma. Additionally, we recruited 121 healthy participants to serve as controls for comparative analyses. The IL-6 gene (− 174G/C) polymorphism was assessed utilizing the polymerase chain reaction-restriction fragment length polymorphism (PCR–RFLP) method.
Results
In our study, the risk of mild asthma exhibited a significant fourfold increase in individuals with the GG genotype pattern compared to healthy controls, yielding an odds ratio (OR) of 4.4 (p < 0.001). Conversely, we found no significant correlation between the IL-6 − 174G/C gene polymorphism and severe asthma when compared to the healthy control group. However, a noteworthy pattern emerged when we compared subgroups of mild and severe asthma. The risk of severe asthma increased fivefold in individuals with the GC polymorphism pattern, with an OR of 4.99 (p < 0.001), while the likelihood of mild asthma showed a similar fourfold increase with the GG polymorphism pattern, OR = 4.4 (p < 0.001). Consequently, we observed a significantly higher frequency of the C allele in patients with severe asthma, whereas the G allele was more prevalent in individuals with mild asthma (p = 0.05). Additionally, the correlation between markers of type 2 inflammation and the dominant model of the IL-6 gene -174G/C polymorphism (CC + CG vs GG) revealed a significant increase in total serum immunoglobulin E (IgE), Blood Eosinophil Counts (BEC), and Fractional Exhaled Nitric Oxide (FeNO) levels in asthmatic patients with the CC + CG gene pattern compared to those with GG, with p-values of 0.04, 0.03, and 0.04, respectively. Furthermore, after adjusting for other risk factors, the likelihood of developing severe asthma increased from fourfold to eightfold, with an OR of 8.12 (p = 0.01) with (CC + CG) gene pattern. Other predictors for severe asthma included older age and childhood-onset disease (OR = 1.13 and 19.19, p < 0.001). Allergic rhinitis (AR) and nasal polyps (NP) also demonstrated a substantial association with an increased risk of severe asthma, with odds ratios of 5 and 32.29 (p = 0.01 and < 0.001), respectively. Additionally, elevated Body Mass Index (BMI), BEC, and FeNO were linked to severe asthma, with ORs of 1.11, 1.00, and 1.04, respectively (p = 0.04, 0.05, and 0.001).
Conclusion
This study illuminated the intricate relationship between the IL-6 gene polymorphism, type 2 inflammation markers, and diverse risk factors in shaping asthma severity. As a significant association between the GG polymorphism of the IL-6 gene (− 174G/C) and mild asthma was found, while possessing at least one C allele, whether in a homozygous (CC) or heterozygous (CG) combination, independently predicts the likelihood of severe asthma.
Similar content being viewed by others


Introduction
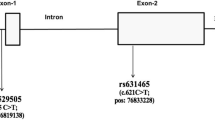
The IL-6 gene is located on chromosome 7p21 and has many single nucleotide polymorphisms (SNPs) in the database. However, only three SNPs (− 597 G/A (rs1800797), − 572 G/C (rs1800796), and − 174 G/C (rs1800795) have been extensively studied for their role in different diseases [1]. Individuals may exhibit varying cytokine production levels due to SNPs in crucial regulatory regions, including promoters, introns, and the 5′-UTR and 3′-UTR regulatory regions. These SNPs can impact cytokine expression levels. Genetic polymorphisms in gene-coding regions may also lead to the loss or modification of function in the expressed proteins [2]. The − 174 G/C SNP is located at position 174 within the promoter region of the IL-6 gene [3]. It has been found to influence the expression of IL-6, particularly its G allele, and regulate different inflammatory responses [4,5,6].
IL-6 is a typical immune-modulating cytokine with a potential pro-inflammatory ability that stimulates both the innate and adaptive immune systems [7, 8]. The role of this cytokine exceeds that of immune-mediated disease to play a pivotal role in hematopoiesis and the endocrine system [9,10,11]. Some innate immune cells are responsible for producing IL-6 as B-cells and CD-4 Th cells [7]. Nevertheless, other non-leukocyte cells secrete this cytokine, such as astrocyte cells, fibroblasts, endothelial cells, and some malignant cells [12,13,14,15,16].
Research has shown that the airway epithelium in lung tissue secretes a significant amount of IL-6, along with T-lymphocytes and macrophages. This secretion of IL-6 is believed to be involved in cellular signaling during the development of asthma [17, 18]. The findings suggest that IL-6 can activate the immune system in response to acute phase reactions caused by viral invasion, bacterial infection, and tissue damage. It also plays a role in differentiating T-cells, increasing the production and function of cytokines and mediators, and creating hyper-inflammatory states in severe COVID-19 and septic shock cases [19,20,21,22,23,24].
New studies have revealed a correlation between heightened levels of IL-6 in the bloodstream and specific allergic conditions, including asthma, allergic rhinitis, and atopic dermatitis [1, 25]. Nevertheless, additional research has indicated that individuals with the IL-6 -174G/C gene polymorphism and corresponding cytokine levels are significantly linked to chronic inflammatory disorders such as coronary heart disease [3], arthritis [26, 27], and metabolic syndromes like diabetes mellitus, hypertension, and obesity [28,29,30,31].
Individuals who suffer from allergic diseases such as asthma, allergic rhinitis, and atopic dermatitis experience multiple conditions and frequently possess a shared genetic predisposition. Nevertheless, specific asthma phenotypes deviate from the standard allergy-related procedure, and their pathogenesis varies, often resulting in a more severe ailment [32]. The traditional classification of asthma into extrinsic and intrinsic subtypes also reflects cellular and molecular mechanisms. Hence, extrinsic asthma mainly passes in the allergic pathway and is attributed to Th2 inflammation; however, intrinsic asthma is mainly triggered by other factors, including infections, obesity, exercise, stress, cold, and other causes, and may be passed through different pathogenic mechanisms [33, 34]. However, asthma’s molecular and cellular pathogenesis is complex and overlaps due to different arrays of precipitating factors with dissimilar mechanisms, which could reflect the discrepancy between the traditional classifications of asthma phenotypes. Allergic asthma is initiated in early childhood, continues to adulthood, and is often persistent, with significant variations in disease severity [35, 36].
Several epidemiological studies have found that atopic asthma tends to be less severe than non-atopic asthma [37]. Additionally, patients with mild to moderate asthma tend to have a more robust sensitization response than those with severe asthma [38].
The variations in markers of type 2 inflammations, such as IgE, BEC, and FeNO, differ among various phenotypes/endotypes of asthma. In allergic asthma, a persistent Th2-type inflammatory process is initiated upon exposure to inhaled allergens. Susceptible individuals are more predisposed to activating the airway epithelium and dendritic cells, ultimately leading to the synthesis of specific IgE antibodies [39]. Upon re-exposure to the allergen, cross-linking of FcεRI on tissue mast cells results in immediate bronchoconstriction, eosinophil recruitment, and a late-phase inflammatory response [40]. Detection of specific IgE in the serum through serology or skin-prick testing is a key feature of allergic asthma. Blood eosinophils are typically moderately elevated, and other atopy-related disorders, such as allergic rhinitis and atopic dermatitis, are commonly associated with allergic asthma [41]. The atopic phenotype is genetically determined and influenced by single nucleotide polymorphisms at Th2 genes, and 17q12 loci. While total IgE values overlap between atopic and nonatopic subjects, serum IgE levels closely correlate with asthma risk and airway hyperresponsiveness (AHR), regardless of allergen specificity [42, 43]. High serum IgE levels are more commonly observed in severe and early-onset asthma, and very high levels, greater than 2000 kU L−1, correlate with dermatitis severity [44, 45].
On the other hand, around 25–30% of people with asthma experience the “nonallergic” phenotype, which is characterized by asthma onset in adulthood without any known cause. Within this phenotype, there is a subset called intrinsic asthma that is different from allergic asthma because there are no detectable specific IgE antibodies in the blood, and skin-prick tests for common allergens are negative [46]. Clinical features of intrinsic asthma include a later onset, no family history of asthma, higher prevalence in women, and often being associated with chronic rhinosinusitis with nasal polyps, which can lead to a more severe form of the disease [47].
People with nonallergic asthma may have higher levels of total serum IgE than healthy individuals [43]. Although there are differences between allergic and intrinsic asthma, there is some overlap in immune cell infiltration and local IgE synthesis [43]. However, the allergen specificity of the IgE remains unknown. A subset of intrinsic asthma known as “hyper-eosinophilic asthma,” which occurs later in life and is not caused by allergies; this type is associated with nasal polyps and high levels of eosinophils in the blood (over 1000 μL–1 or over 500 μL–1 when treated with oral corticotherapy). Biological therapy that targets IL-5, such as mepolizumab and reslizumab, are effective treatments for this subtype [48]. Severe cases are characterized by high levels of eosinophils in both blood and sputum, more commonly found in males, airflow limitation, elevated FeNO, and frequent severe exacerbations [49]. Isolated sputum eosinophilia is less linked with poor control but is associated with higher serum IgE levels [43, 50]. There is no identified genetic predisposition, but IgE responses to enterotoxin-producing S. aureus colonizing the upper airways are widespread [51]. Staphylococcal superantigens can cause polyclonal T- and B-cell activation [52]. Anti-IgE therapy has shown to be beneficial in asthma with nasal polyps, regardless of atopic status, supporting the role of IgE in hypereosinophilic nonallergic asthma [53].
Moreover, a distinct subgroup without evidence of Th2 eosinophilic inflammation was reported and characterized by severe corticosteroid resistance shares features with chronic obstructive pulmonary disease. This phenotype may be more accurately termed “type 2-low” asthma, given the involvement of alternative producers like ILC2s in “type 2” cytokine production [43]. The role of IgE in this phenotype is unclear, but IL-17, IL-33, and ADAM8 have been implicated [54].
To date, there is a notable gap in research addressing the inflammatory implications of IL-6 gene -174G/C polymorphism in adult asthma, particularly within Kuwait and the Gulf region. This study aims to fill this gap by investigating the association between the IL-6 gene − 174G/C polymorphism and asthma severity in adults, distinguishing between mild and severe asthma and assessing its influence on type 2 inflammation while focusing on total serum IgE, BEC and FeNO levels as markers of type 2 inflammations.
Patients and methods
Patients and study design
This case–control study was conducted et al.-Rashed Allergy Center, Kuwait, from December 2022 to April 2023. The study enrolled 98 mild and 116 severe asthma patients based on clinical diagnosis and reversibility of FEV1% [55]. The study also included 121 healthy controls for comparison. Patients with mild asthma were characterized by infrequent symptoms and relatively well-preserved lung function, which minimally impacted their daily activities. In contrast, severe asthma manifested persistent symptoms, frequent exacerbations, frequent use of corticosteroid courses, and significant lung function limitations, often requiring intensive treatment and medical intervention [55, 56]
Ethics approval and consent to participate
Ethical approval has been obtained from Kuwait University and the Ministry of Health, Kuwait (Research project number MI02/22, 2022/2010), and to ensure that the research is conducted ethically and in compliance with the internationally recognized standards, Helsinki Declaration protocol were applied.
Informed consent has been obtained from all participants involved in the study and their legal guardians to ensure that they are fully aware of the nature and purpose of the research; they have given their informed consent voluntarily to participate.
Sample size
The sample size required for this study was calculated using Minitab 17.1.0.0 for Windows software (Minitab Inc., 2013, Pennsylvania, USA). To achieve 80% power, we considered an odds ratio of 2, a disease prevalence of about 1%, a minor allele frequency of 5%, complete linkage disequilibrium (LD), and a 5% error rate in an allelic test. This study’s minimum total sample size was 248, with a 1:1 case/control ratio. Additionally, to ensure a robust representation of the subgroup of cases with varying degrees of asthma severity (mild and severe), we included an equal number of mild asthma cases, assuming a case: case ratio similar to the control: case ratio, to achieve 80% study power.
Sample collection
In a sterile setting, 10 mL of venous blood was obtained from each participant via a plastic syringe. The blood was then split into two tubes—one without anticoagulant and the other with EDTA to aid DNA extraction. After centrifugation at 4000 g for 10 min, the serum, plasma, and buffy coat with leukocytes were separated. The samples were then frozen at − 20 °C for DNA extraction. Genomic DNA was extracted using the QIAamp Blood Kit (QIAGEN, Germany) following the manufacturer's protocol. The purity and quantity of the extracted DNA were evaluated using a Nanodrop 8000 spectrophotometer (Thermo-Scientific, Delaware, USA), measuring absorbance at wavelengths 260 and 280 nm. DNA purity was assessed by the A260/A280 ratio, with the target range set between 1.8 and 2.0. To estimate DNA concentration, the optical density (O.D.) at 260 nm was measured, and the concentration (mg/ml) was calculated using the formula: Concentration = O.D. 260 × 50 x dilution factor (× 100). The final DNA concentration was directly determined using the Nanodrop 8000 spectrophotometer software, falling within the optimal range of 107–552 ng/μL.
Genotyping
The genotypes for IL6 gene (− 174G/C; rs1800795) polymorphism were identified by PCR–RFLP (polymerase chain reaction-restriction fragment length polymorphism). The details about primers and PCR method used are given below:
Forward primer: 5ʹ-GGAGTCACACACTCCACCT-3ʹ,
Reverse primer: 5ʹ-GTGGGGCTGATTGGAAACC-3ʹ.
The PCR reactions were carried out in a total volume of 25 ml containing 100 ng of genomic DNA, 10 pmol of each primer, 2 mM MgCl2, 0.2 mM deoxynucleotides (dNTPs), 1 × buffer, and 2U of Taq DNA polymerase. For IL-6 gene (− 174G/C) polymorphism, the amplification was performed for 35 cycles with an initial denaturation step at 94 °C for 5 min followed by the PCR cycles of 94 °C for 1 min, annealing at 65 °C for 1 min, 72 °C for 1 min and then an extension step at 72 °C for 10 min. The polymorphism was identified by cleavage of the PCR products with restriction enzyme SfaN1. The GàC change at position 174 creates a restriction site for this enzyme and consequently, G-allele produces an un-cleaved 532 bp product while the C-allele yields two products of 474 and 58 bp, respectively upon digestion of the PCR products with SfaN1.The DNA cleavage fragments were resolved by electrophoresis on a 3% agarose gel and were visualized under UV light after Ethidium bromide staining.
Statistical analysis
Demographic data of patients and control groups were collected in an Excel sheet, along with the genotype of each study participant. Minitab for Windows (Minitab Inc, 2013, version 17.1.0.0, Pennsylvania, USA) was used for statistical analysis. The data is represented as mean and standard deviation for numerical data and number (%) for categorical data. Comparison between two means was performed using an independent t-test and between two frequencies through a Chi-square test. Logistic regression analysis with adjusted and non-adjusted methods was applied to find the predictive ability of IL-6 -174G/C gene polymorphism and severe asthma. All tests were two-sided, and p < 0.05 was considered significant.
Results
In Table 1, a comparison of demographic characteristics and the frequency of IL-6 -174G/C gene polymorphism was conducted between control subjects and asthma subgroups. The patients with severe asthma exhibited a significantly higher age (52.8 ± 11.5 years) compared to the control (37.5 ± 16.1 years) and mild asthma subgroups (38.3 ± 16.6 years), with a p-value of < 0.001. However, the three groups were matched with respect to gender and BMI.
Moreover, the polymorphism pattern of IL-6 -174G/C gene subgroup analysis showed that the risk of mild asthma increased fourfold with the GG genotype pattern compared to healthy control, OR = 4.4, p < 0.001. However, the GC pattern showed a protective ability against mild asthma, OR = 0.31, p = 0.002. Additionally, the frequency of the G allele was significantly higher in mild asthmatic patients, OR = 1.67, p = 0.006. Conversely, the frequency of the C allele was significantly observed in the control subject, OR = 0.41, p = 0.001. The dominant model of IL-6 -174G/C gene (CC + CG vs GG) showed that the risk of mild asthma decreased fourfold, OR = 4.4, p < 0.001, while the recessive model (CC Vs CG + GG) showed insignificant association, p = 0.32. On the other hand, no correlation was found between the IL-6 -174G/C gene polymorphism and severe asthma compared to healthy controls. However, in comparing mild and severe asthma subgroups, the risk of severe asthma increased fivefold with the GC polymorphism pattern, OR = 4.99, p < 0.001, while the likelihood of mild asthma increased fourfold with the GG polymorphism pattern, OR = 4.4, p < 0.001. Thus, the frequency of the C allele was significantly higher in patients with severe asthma, while the G allele was higher in mild asthma patients, p = 0.05. The use of dominant model of IL-6 gene − 174G/C polymorphism (CC + CG vs GG) showed that (CC + CG) increased the likelihood of severe asthma fourfold, OR = 4.4, p < 0.001. Yet, in the recessive model (CC vs CG + GG) revealed an insignificant association, p = 0.65.
In Table 2, a comparison between mild and severe asthma regarding clinical features and markers of type 2 inflammation revealed notable differences. Patients with severe asthma exhibited significantly higher prevalence of comorbidities, specifically allergic rhinitis (AR) and nasal polyp (NP), at 73.28% and 49.14%, respectively, with a p-value of < 0.001. Additionally, levels of total serum IgE, BEC, and FeNo were significantly elevated, with p-values of < 0.001, 0.001, and 0.002, respectively. Conversely, the FEV1% predicted was significantly lower in the severe asthma group, demonstrating a p-value of 0.03. Furthermore, the frequency of exacerbations requiring an oral corticosteroid (OCS) course was significantly higher in the severe asthma group, with a p-value of 0.001. Additionally, the correlation between markers of type 2 inflammation and the dominant model of the IL-6 gene -174G/C polymorphism (CC + CG vs GG) was presented in Table 3. The results revealed a significant increase in total serum IgE, BEC, and FeNO levels in asthmatic patients with the CC + CG gene pattern compared to those with GG, with p-values of 0.04, 0.03, and 0.04, respectively.
Moreover, the predictors of severe asthma are outlined in Table 4, with the prominent IL-6 gene -174GC polymorphism (CC + CG) model emerging as an independent predictor. Upon adjusting for other risk factors, the likelihood of severe asthma significantly increased, ranging from fourfold to eightfold, presenting an odds ratio (OR) of 8.12 with a p-value of 0.01. Other independent predictors for severe asthma encompassed older age and childhood-onset disease, yielding odds ratios of 1.13 and 19.19, respectively, with p-values less than 0.001. Additionally, AR and NP substantially heightened the risk of severe asthma, with odds ratios of 5 and 32.29 (p < 0.01 and < 0.001, respectively). Furthermore, elevated BMI, BEC, and FeNO levels were also associated with severe asthma, exhibiting odds ratios of 1.11, 1.00, and 1.04, and corresponding p-values of 0.04, 0.05, and 0.001, respectively.
Discussion
Various research studies have attempted to examine the role of genetic factors in the development and progression of severe asthma [57]. However, a comprehensive understanding of this topic still needs to be improved. Given the diversity of asthma, various endo/phenotypes have arisen, each indicative of a unique pathogenic process [58]. The main objective of genetic investigations in a multifaceted condition such as asthma is to pinpoint the link between a cluster of genes or their variations and the onset or progression of the disease toward a severe state [59].
This study aimed to explore the possible association between the IL-6 gene (− 174G/C) polymorphism and adult patients with bronchial asthma, emphasizing assessing its correlation with the disease severity. The finding revealed that in comparing mild asthma with control subjects, the risk of mild asthma increased four and two-fold in homogenous (GG) and allele comparison (G), respectively, and reduced in dominant (CC + GC), heterogeneous (GC), and allele comparison (C), respectively. However, in comparing mild asthma with severe one, the risk of severe asthma increased four, five, and two-fold in dominant (CC + GC), heterogeneous (GC), and allele (C) comparisons, respectively, and reduced in homogeneous (GG) comparisons.
In previous literature [60, 61], the relationship between the IL-6 gene (− 174G/C) polymorphism and bronchial asthma remains inconclusive. A comprehensive meta-analysis study examined the potential correlation between the IL-6 gene (− 174G/C) polymorphism and allergic diseases in a sample of 1282 patients with allergic ailments and 1902 control subjects [1]. The findings revealed that, in the general population, there was no notable link between the IL-6 gene − 174G/C polymorphism and the overall risk of allergic diseases. However, trends were contrasted when the analysis was based on ethnicity. Among Caucasians, the IL-6 gene (− 174G/C) polymorphism was correlated with a reduced risk of overall allergic diseases in dominant, allele, and heterozygote comparisons. Conversely, in Asian populations, the IL-6 gene (− 174G/C) polymorphism was associated with an increased risk of overall allergic diseases in dominant, allele, and homozygote comparisons. Additionally, the study found that the IL-6 (− 174G/C) variant significantly correlates with a lower risk of childhood allergic diseases across different age groups, observed in multiple comparisons, including dominant, allele, heterozygote, and homozygote comparisons. Furthermore, the homozygote comparison remarkably reduced the risk of allergic diseases.
Investigating a connection between IL-6 gene variations and specific allergic conditions is crucial due to the contribution of various factors and epigenetic influences on different allergic diseases. This approach is necessary to assess the effectiveness of the study for each category individually. A more extensive analysis included 746 asthmatic patients and 1145 controls and investigated the link between the IL-6 gene (− 174G/C) polymorphism variant and asthma risk [1]. The results reported a significant association between IL-6 gene (− 174G/C) polymorphism and asthma risk, specifically in the recessive and homozygote comparisons. Moreover, a subgroup analysis by ethnicity and age revealed that among Caucasians, the IL-6 gene (− 174G/C) polymorphism decreased asthma risk in both the recessive and homozygote comparisons. However, in children, it reduced asthma risk in the dominant, heterozygous, and homozygous comparisons. A similar trend was observed in adults under the homozygote comparison [62]. In another meta-analysis, the data also discovered that CC genotype carriers exhibited a protective effect against asthma in a study of Caucasian populations [60, 62]. However, no evidence was found to support this in the Asian population [63]. Nevertheless, further analysis revealed that the CC genotype may offer protection against asthma in adults [60, 64]. These findings suggest that the IL-6 gene (− 174G/C) polymorphism may play a role in asthma pathogenesis by altering transcriptional regulation and serum IL-6 levels [60, 64, 65].
On the other hand, other studies have refuted any association between IL-6 cytokines and its gene -174G/C polymorphism with bronchial asthma and have considered IL-6 a product of airway inflammation [66, 67].
In a different aspect, a supportive finding came with Noss et al., the study revealed the relationship between genetic variation, protein expression, and IL-6 regulation in rheumatoid arthritis [68]. They identified a specific association between the IL-6 proximal promoter polymorphism rs1800795 and heightened IL-6 production in fibroblasts; the highest IL-6 levels were linked to a genetic variant (minor allele C), thus emphasizing the importance of examining diverse cell types. The significance of his study lies in its ability to offer valuable insights for interpreting genetic associations and unraveling the complexities of gene regulation in chronic inflammatory diseases such as rheumatoid arthritis.
In a smaller study, patients with infantile asthma were shown to have a strong correlation with IL-6 gene (-174G/C) polymorphism; the likelihood of asthma increased up to fourfold, especially in the atopic phenotype [69]. The inconsistency among genetic studies regarding asthma may be attributed to the complex nature of its origin and the diverse forms it can take.
Our results demonstrate that patients with severe asthma were generally older patients than in those with mild asthma. Notably, many of these patients also suffered from associated AR and NP. Additionally, total serum IgE and BEC were significantly elevated, along with FeNO; these pointed to the presence of type 2 inflammation in patients with severe asthma. Moreover, this study showed a connection between the dominant pattern of the IL-6 gene (-174G/C) and markers of type 2 inflammation. Specifically, it demonstrated that the dominant variation (CC + CG) is associated with increased levels of total serum IgE, BEC, and FeNO compared to the GG variation. Additionally, the study found that predictors of severe asthma, besides the dominant variation of IL-6 gene (-174G/C) polymorphism, were being older with childhood-onset disease. We also report that the likelihood of severe asthma increased two-fold every year and 19-fold in patients with childhood-onset disease. In addition, the AR and NP increased the risk of severe asthma five and 32-fold.
Individuals with severe asthma often experience persistent and challenging symptoms that are difficult to manage [70]. Unlike typical asthma cases, this refractory condition is characterized by an inadequate response to traditional treatments and heightened, prolonged symptoms [71]. Asthma involves lung inflammation, airway hyper-responsiveness, airway remodeling, and excessive mucus production, all contributing to airflow issues. It is not one disease but a combination of variations in these features [72]. Severe asthma significantly impairs patients' quality of life and managing the disease can be daunting [73]. Despite extensive research to understand its causes and treatment responses, many aspects of severe asthma are still unknown. Our patients with severe asthma exhibited steroid resistance and required additional biological therapy to control the disease, in addition to managing comorbidities such as nasal polyps and allergic rhinitis. A recent study in Kuwait found a high prevalence of asthma in patients with nasal polyps [74], and the GG genotype was significantly associated with both nasal polyps and bronchial asthma [75]. However, the study did not determine the severity of asthma. Another study found that GG genotype was significantly reported in 67% of patients with NP [76]. The development of polyps in the nasal cavity is attributed mainly to chronic inflammation, closely resembling the pathogenesis of severe asthma. Various types of inflammatory cells, including eosinophils, neutrophils, plasma cells, lymphocytes, and mast cells, contribute to this process [77]. The inflammatory pathways Th1 and Th2 are activated by signaling molecules, such as IL-6, further exacerbating the inflammatory response [78].
Unlike NP, research findings indicate that possessing the CC genotype of the IL-6 gene (-174G/C) polymorphism may increase the likelihood of developing allergic rhinitis two-fold in the Chinese population [62]. Likewise, studies conducted in Iran suggest that the IL-6 gene (− 174G/C) polymorphism could contribute to the onset of AR [67].
While patients with mild and severe asthma were initially matched in this study for BMI during univariate analysis, the multivariate analysis revealed that the probability of severe asthma doubled with every one-point increase in BMI. Surprisingly, this factor did not correlate with the dominant polymorphism pattern of the IL-6 gene (− 174G/C). In contrast, a study investigating specific genetic variations in the IL6 promoter, including − 174 G/C and − 572 G/C polymorphisms, found associations with insulin resistance, dyslipidemia, and increased serum insulin release [79]. Composite genotype and haplotype analyses involving all three IL6 promoter variants were linked to type 2 diabetes, obesity, and metabolic syndrome in Caucasians. These findings align with a parallel study in children [80], which explored the association between the G-174C polymorphism of the IL-6 gene and obesity as well as the incidence of metabolic syndrome (MetS). Interestingly, carriers of the C allele, in both homozygotic and heterozygotic genotypes, were more prevalent among obese children. However, despite a 33% incidence of MetS in the obese group, the analysis did not establish a significant association between the 174G > C polymorphism and either obesity or the occurrence of MetS in children. This discrepancy suggests the presence of other influencing factors beyond the IL-6 174G > C gene polymorphism in patients with inflammatory diseases like metabolic syndrome or asthma.
Our study has certain limitations that need to be acknowledged. Firstly, the patients and control subjects were all selected from a single center, which might have caused a selection bias. However, we took measures to address this issue by carefully matching the control group with the patients, especially those with mild asthma. Secondly, we only investigated polymorphisms within the IL-6 gene without examining other genetic variants in asthma development.
In conclusion, patients with severe asthma were notably older. They had significant comorbidities such as AR and NP, accompanied by high levels of BEC, total serum IgE, and FeNO. The dominant model (CC + GC) of the IL-6 gene (-174G/C) polymorphism was significantly associated with severe asthma and influencing type 2 inflammatory markers.
Availability of data and materials
The datasets used during the current study are available from the corresponding author on reasonable request.
References
Yang Y, Xiao J, Tang L, Wang B, Sun X, Xu Z, Liu L, Shi S. Effects of IL-6 polymorphisms on individual susceptibility to allergic diseases: a systematic review and meta-analysis. Front Genet. 2022;16(13):822091.
Haukim N, Bidwell JL, Smith AJ, Keen LJ, Gallagher G, Kimberly R, Huizinga T, McDermott MF, Oksenberg J, McNicholl J, Pociot F, Hardt C, D’Alfonso S. Cytokine gene polymorphism in human disease: on-line databases, supplement 2. Genes Immun. 2002;3(6):313–30. https://doi.org/10.1038/sj.gene.6363881.
Rai H, Colleran R, Cassese S, Joner M, Kastrati A, Byrne RA. Association of interleukin 6–174 G/C polymorphism with coronary artery disease and circulating IL-6 levels: a systematic review and meta-analysis. Inflamm Res. 2021;1:1–3.
Forbester JL, Humphreys IR. Genetic influences on viral-induced cytokine responses in the lung. Mucosal Immunol. 2021;14(1):14–25.
Chair SY, Law BM, Chan JY, So WK, Waye MM. Association of genetic polymorphisms with psychological symptoms in cancer: a systematic review. Asia Pac J Oncol Nurs. 2022;9(1):12–20.
Rajabi-Moghaddam M, Abbaszadeh H. Gene polymorphisms and risk of head and neck squamous cell carcinoma: a systematic review. Rep Pract Oncol Radiother. 2022. https://doi.org/10.5603/RPOR.a2022.0115.
Tanaka T, Narazaki M, Kishimoto T. IL-6 in inflammation, immunity, and disease. Cold Spring HarbPerspect Biol. 2014;6(10):a016295. https://doi.org/10.1101/cshperspect.a016295.
Rose-John S, Winthrop K, Calabrese L. The role of IL-6 in host defence against infections: immunobiology and clinical implications. Nat Rev Rheumatol. 2017;13(7):399–409. https://doi.org/10.1038/nrrheum.2017.83.
Groza Y, Jemelkova J, Kafkova LR, Maly P, Raska M. IL-6 and its role in IgA nephropathy development. Cytokine Growth Factor Rev. 2022;66:1–14. https://doi.org/10.1016/j.cytogfr.2022.04.001.
Hauser SP, Kajkenova O, Lipschitz DA. The pivotal role of interleukin 6 in formation and function of hematopoietically active murine long-term bone marrow cultures. Stem Cells. 1997;15(2):125–32. https://doi.org/10.1002/stem.150125.
Papanicolaou DA. Interleukin-6: the endocrine cytokine. J Clin Endocrinol Metab. 2000;85(3):1331–3.
Zhang JM, An J. Cytokines, inflammation, and pain. Int Anesthesiol Clin. 2007;45(2):27–37. https://doi.org/10.1097/AIA.0b013e318034194e.
Van Wagoner NJ, Oh JW, Repovic P, Benveniste EN. Interleukin-6 (IL-6) production by astrocytes: autocrine regulation by IL-6 and the soluble IL-6 receptor. J Neurosci. 1999;19(13):5236–44. https://doi.org/10.1523/JNEUROSCI.19-13-05236.1999.
Zongfei J, Rongyi C, Xiaomeng C, Lili M, Lingying M, Xiufang K, Xiaomin D, Zhuojun Z, Huiyong C, Ying S, Lindi J. In vitro IL-6/IL-6R trans-signaling in fibroblasts releases cytokines that may be linked to the pathogenesis of IgG4-related disease. Front Immunol. 2020;8(11):1272. https://doi.org/10.3389/fimmu.2020.01272.
Chonov DC, Ignatova MMK, Ananiev JR, Gulubova MV. IL-6 activities in the tumour microenvironment. Part 1. Open Access Maced J Med Sci. 2019;7(14):2391–8. https://doi.org/10.3889/oamjms.2019.589.
Ene CV, Nicolae I, Geavlete B, Geavlete P, Ene CD. IL-6 signaling link between inflammatory tumor microenvironment and prostatic tumorigenesis. Anal Cell Pathol. 2022;12(2022):5980387. https://doi.org/10.1155/2022/5980387.
Rincon M, Irvin CG. Role of IL-6 in asthma and other inflammatory pulmonary diseases. Int J Biol Sci. 2012;8(9):1281–90. https://doi.org/10.7150/ijbs.4874.
Neveu WA, Allard JB, Dienz O, Wargo MJ, Ciliberto G, Whittaker LA, Rincon M. IL-6 is required for airway mucus production induced by inhaled fungal allergens. J Immunol. 2009;183(3):1732–8. https://doi.org/10.4049/jimmunol.0802923.
Velazquez-Salinas L, Verdugo-Rodriguez A, Rodriguez LL, Borca MV. The role of interleukin 6 during viral infections. Front Microbiol. 2019;10(10):1057.
Leemans JC, Vervoordeldonk MJ, Florquin S, van Kessel KP, van der Poll T. Differential role of interleukin-6 in lung inflammation induced by lipoteichoic acid and peptidoglycan from Staphylococcus aureus. Am J Respir Crit Care Med. 2002;165(10):1445–50.
Hirano T. IL-6 in inflammation, autoimmunity, and cancer. Int Immunol. 2021;33(3):127–48. https://doi.org/10.1093/intimm/dxaa078.
Gubernatorova EO, Gorshkova EA, Polinova AI, Drutskaya MS. IL-6: relevance for immunopathology of SARS-CoV-2. Cytokine Growth Factor Rev. 2020;53:13–24. https://doi.org/10.1016/j.cytogfr.2020.05.009.
Sutherland AM, Walley KR, Manocha S, Russell JA. The association of interleukin 6 haplotype clades with mortality in critically ill adults. Arch Intern Med. 2005;165(1):75–82. https://doi.org/10.1001/archinte.165.1.75.
McElvaney OJ, Curley GF, Rose-John S, McElvaney NG. Interleukin-6: obstacles to targeting a complex cytokine in critical illness. Lancet Respir Med. 2021;9(6):643–54.
Pan R, Kuai S, Li Q, Zhu X, Wang T, Cui Y. Diagnostic value of IL-6 for patients with asthma: a meta-analysis. Allergy Asthma Clin Immunol. 2023;19(1):39. https://doi.org/10.1186/s13223-023-00794-3.
Favalli EG. Understanding the role of interleukin-6 (IL-6) in the joint and beyond: a comprehensive review of IL-6 inhibition for the management of rheumatoid arthritis. Rheumatol Ther. 2020;7(3):473–516.
Honsawek S, Deepaisarnsakul B, Tanavalee A, Yuktanandana P, Bumrungpanichthaworn P, Malila S, Saetan N. Association of the IL-6-174G/C gene polymorphism with knee osteoarthritis in a Thai population. Genet Mol Res. 2011;10(3):1674–80.
Pal M, Febbraio MA, Whitham M. From cytokine to myokine: the emerging role of interleukin-6 in metabolic regulation. Immunol Cell Biol. 2014;92(4):331–9.
Zafar U, Khaliq S, Ahmad HU, Lone KP. Serum profile of cytokines and their genetic variants in metabolic syndrome and healthy subjects: a comparative study. 2019. Biosci Rep. https://doi.org/10.1042/BSR20181202.
Lehrskov LL, Christensen RH. The role of interleukin-6 in glucose homeostasis and lipid metabolism. Semin Immunopathol. 2019;41(4):491–9.
Hastuti P, Martantiningtyas DC, Karita D, Tasmini SA, Sadewa A. Association of-174 G> C interleukin-6 gene polymorphism with interleukin-6 and c-reactive protein levels and obesity: a case–control study among people/residents of western Indonesia. Med J Malaysia. 2019;74(5):400.
Ferreira MA, Vonk JM, Baurecht H, Marenholz I, Tian C, Hoffman JD, Helmer Q, Tillander A, Ullemar V, van Dongen J, Lu Y, Rüschendorf F, Esparza-Gordillo J, Medway CW, Mountjoy E, Burrows K, Hummel O, Grosche S, Brumpton BM, Witte JS, Hottenga JJ, Willemsen G, Zheng J, Rodríguez E, Hotze M, Franke A, Revez JA, Beesley J, Matheson MC, Dharmage SC, Bain LM, Fritsche LG, Gabrielsen ME, Balliu B, Nielsen JB, Zhou W, Hveem K, Langhammer A, Holmen OL, Løset M, Abecasis GR, Willer CJ, Arnold A, Homuth G, Schmidt CO, Thompson PJ, Martin NG, Duffy DL, Novak N, Schulz H, Karrasch S, Gieger C, Strauch K, Melles RB, Hinds DA, Hübner N, Weidinger S, Magnusson PKE, Jansen R, Jorgenson E, Lee YA, Boomsma DI, Almqvist C, Karlsson R, Koppelman GH, Paternoster L. Shared genetic origin of asthma, hay fever and eczema elucidates allergic disease biology. Nat Genet. 2017;49(12):1752–7. https://doi.org/10.1038/ng.3985.
Wang J, Zhou Y, Zhang H, Hu L, Liu J, Wang L, Wang T, Zhang H, Cong L, Wang Q. Pathogenesis of allergic diseases and implications for therapeutic interventions. Signal Transduct Target Ther. 2023;8(1):138.
Sadeghnejad A, Bleecker E, Meyers D. Principles of genetics in allergic diseases and asthma. Middlet Allergy Princ Pract. 2008;7:59–73.
Holt PG, Upham JW, Sly PD. Contemporaneous maturation of immunologic and respiratory functions during early childhood: implications for development of asthma prevention strategies. J Allergy Clin Immunol. 2005;116(1):16–24. https://doi.org/10.1016/j.jaci.2005.04.017.
Lotvall J, Akdis CA, Bacharier LB, Bjermer L, Casale TB, Custovic A, et al. Asthma endotypes: a new approach to classification of disease entities within the asthma syndrome. J Allergy Clin Immunol. 2011;127:355–60.
Siroux V, Oryszczyn MP, Paty E, Kauffmann F, Pison C, Vervloet D, Pin I. Relationships of allergic sensitization, total immunoglobulin E and blood eosinophils to asthma severity in children of the EGEA study. Clin Exp Allergy. 2003;33(6):746–51. https://doi.org/10.1046/j.1365-2222.2003.01674.x.
The ENFUMOSA cross-sectional European multicentre study of the clinical phenotype of chronic severe asthma. European network for understanding mechanisms of severe asthma. Eur Respir J. 2003;22(3):470–7. https://doi.org/10.1183/09031936.03.00261903.
Platts-Mills TA, Woodfolk JA. Allergens and their role in the allergic immune response. Immunol Rev. 2011;242(1):51–68. https://doi.org/10.1111/j.1600-065X.2011.01021.x.
Martinez FD, Vercelli D. Asthma. Lancet. 2013;382(9901):1360–72. https://doi.org/10.1016/S0140-6736(13)61536-6.
Jiang H, Hener P, Li J, Li M. Skin thymic stromal lymphopoietin promotes airway sensitization to inhalant house dust mites leading to allergic asthma in mice. Allergy. 2012;67(8):1078–82. https://doi.org/10.1111/j.1398-9995.2012.02857.x.
Burrows B, Martinez FD, Halonen M, Barbee RA, Cline MG. Association of asthma with serum IgE levels and skin-test reactivity to allergens. N Engl J Med. 1989;320(5):271–7. https://doi.org/10.1056/NEJM198902023200502.
Froidure A, Mouthuy J, Durham SR, Chanez P, Sibille Y, Pilette C. Asthma phenotypes and IgE responses. Eur Respir J. 2016;47(1):304–19. https://doi.org/10.1183/13993003.01824-2014.
Borish L, Chipps B, Deniz Y, Gujrathi S, Zheng B, Dolan CM. Total serum IgE levels in a large cohort of patients with severe or difficult-to-treat asthma. Ann Allergy Asthma Immunol. 2005;95(3):247–53. https://doi.org/10.1016/S1081-1206(10)61221-5.
Joshi AY, Iyer VN, Boyce TG, Hagan JB, Park MA, Abraham RS. Elevated serum immunoglobulin E (IgE): when to suspect hyper-IgE syndrome-A 10-year pediatric tertiary care center experience. Allergy Asthma Proc. 2009;30(1):23–7. https://doi.org/10.2500/aap.2009.30.3193.
Ilmarinen P, Tuomisto LE, Kankaanranta H. Phenotypes, risk factors, and mechanisms of adult-onset asthma. Med Inflamm. 2015. https://doi.org/10.1155/2015/514868.
Peters SP. Asthma phenotypes: nonallergic (intrinsic) asthma. J Allergy Clin Immunol Pract. 2014;2(6):650–2. https://doi.org/10.1016/j.jaip.2014.09.006.
Dean K, Niven R. Asthma phenotypes and endotypes: implications for personalised therapy. BioDrugs. 2017;31(5):393–408. https://doi.org/10.1007/s40259-017-0242-5.
Carr TF, Kraft M. Use of biomarkers to identify phenotypes and endotypes of severeasthma. Ann Allergy Asthma Immunol. 2018;121(4):414–20. https://doi.org/10.1016/j.anai.2018.07.029.
Manise M, Holtappels G, Van Crombruggen K, Schleich F, Bachert C, Louis R. Sputum IgE and cytokines in asthma: relationship with sputum cellular profile. PLoS ONE. 2013;8(3):e58388. https://doi.org/10.1371/journal.pone.0058388.
Huvenne W, Hellings PW, Bachert C. Role of staphylococcal superantigens in airway disease. Int Arch Allergy Immunol. 2013;161(4):304–14. https://doi.org/10.1159/000350329.
Sørensen M, Klingenberg C, Wickman M, Sollid JUE, Furberg AS, Bachert C, Bousquet J. Staphylococcus aureus enterotoxin sensitization is associated with allergic poly-sensitization and allergic multimorbidity in adolescents. Allergy. 2017;72(10):1548–55. https://doi.org/10.1111/all.13175.
Kariyawasam HH, James LK. Chronic rhinosinusitis with nasal polyps: targeting IgE with anti-IgE omalizumab therapy. Drug Des Dev Ther. 2020;10:5483–94.
Calvén J, Ax E, Rådinger M. The airway epithelium-a central player in asthma pathogenesis. Int J Mol Sci. 2020;21(23):8907. https://doi.org/10.3390/ijms21238907.
Louis R, Satia I, Ojanguren I, Schleich F, Bonini M, Tonia T, Rigau D, Ten Brinke A, Buhl R, Loukides S, Kocks JWH, Boulet LP, Bourdin A, Coleman C, Needham K, Thomas M, Idzko M, Papi A, Porsbjerg C, Schuermans D, Soriano JB, Usmani OS. European respiratory society guidelines for the diagnosis of asthma in adults. Eur Respir J. 2022. https://doi.org/10.1183/13993003.01585-2021.
Ibrahim MA, Ismail AI, Rani MF. A brief review of severe asthma. J Clin Health Sci. 2021;6(2):4–12.
Slager RE, Hawkins GA, Li X, Postma DS, Meyers DA, Bleecker ER. Genetics of asthma susceptibility and severity. Clin Chest Med. 2012;33(3):431–43. https://doi.org/10.1016/j.ccm.2012.05.005.
Habib N, Pasha MA, Tang DD. Current understanding of asthma pathogenesis and biomarkers. Cells. 2022;11(17):2764. https://doi.org/10.3390/cells11172764.
Torgerson DG, Ampleford EJ, Chiu GY, Gauderman WJ, Gignoux CR, Graves PE, Himes BE, Levin AM, Mathias RA, Hancock DB, Baurley JW, Eng C, Stern DA, Celedón JC, Rafaels N, Capurso D, Conti DV, Roth LA, Soto-Quiros M, Togias A, Li X, Myers RA, Romieu I, Van Den Berg DJ, Hu D, Hansel NN, Hernandez RD, Israel E, Salam MT, Galanter J, Avila PC, Avila L, Rodriquez-Santana JR, Chapela R, Rodriguez-Cintron W, Diette GB, Adkinson NF, Abel RA, Ross KD, Shi M, Faruque MU, Dunston GM, Watson HR, Mantese VJ, Ezurum SC, Liang L, Ruczinski I, Ford JG, Huntsman S, Chung KF, Vora H, Li X, Calhoun WJ, Castro M, Sienra-Monge JJ, del Rio-Navarro B, Deichmann KA, Heinzmann A, Wenzel SE, Busse WW, Gern JE, Lemanske RF Jr, Beaty TH, Bleecker ER, Raby BA, Meyers DA, London SJ, Gilliland FD, Martinez FD, Weiss ST, Williams LK, Barnes KC, Ober C, Nicolae DL. Meta-analysis of genome-wide association studies of asthma in ethnically diverse North American populations. Nat Genet. 2011;43(9):887–92. https://doi.org/10.1038/ng.888.
Li F, Xie X, Li S, Ke R, Zhu B, Yang L, Li M. Interleukin-6 gene -174G/C polymorphism and bronchial asthma risk: a meta-analysis. Int J Clin Exp Med. 2015;8(8):12601–8.
Zhu Q, Zhang H, Wang J, Wu Y, Chen X. Associations of TNF-α-238G/A, TNF-α-308G/A, and IL-6-174G/C polymorphisms with the risk of asthma: Evidence from a meta-analysis. Pediatr Pulmonol. 2020;55(11):2893–900.
Xiaomin L, Fenglin C, Jianmin H, Yuzhi S, Binsheng G, Yingmei Z. Correlation between genetic polymorphism of cytokine genes, plasma protein levels and bronchial asthma in the Han people in northern China. J Asthma. 2008;45(7):583–9. https://doi.org/10.1080/02770900802032925.
Settin A, Zedan M, Farag M, Ezz El Regal M, Osman E. Gene polymorphisms of IL-6(-174) G/C and IL-1Ra VNTR in asthmatic children. Indian J Pediatr. 2008;5(10):1019–23. https://doi.org/10.1007/s12098-008-0161-z.
Fishman D, Faulds G, Jeffery R, Mohamed-Ali V, Yudkin JS, Humphries S, Woo P. The effect of novel polymorphisms in the interleukin-6 (IL-6) gene on IL-6 transcription and plasma IL-6 levels, and an association with systemic-onset juvenile chronic arthritis. J Clin Invest. 1998;102(7):1369–76. https://doi.org/10.1172/JCI2629.
Deetz DC, Jagielo PJ, Quinn TJ, Thorne PS, Bleuer SA, Schwartz DA. The kinetics of grain dust-induced inflammation of the lower respiratory tract. Am J Respir Crit Care Med. 1997;155(1):254–9. https://doi.org/10.1164/ajrccm.155.1.9001321.
Huang M, Wang L, Ma H, Wang J, Xiang M. Lack of an association between interleukin-6 -174G/C polymorphism and circulating interleukin-6 levels in normal population: a meta-analysis. DNA Cell Biol. 2013;32(11):654–64. https://doi.org/10.1089/dna.2013.2148.
Alizadeh-Navaei R, Rafiei A, Hedayatizadeh-Omran A, Mohammadzadeh I, Arabi M. Gene susceptibility in Iranian asthmatic patients: a narrative review. Ann Med Health Sci Res. 2014;4(6):837–40. https://doi.org/10.4103/2141-9248.144871.
Noss EH, Nguyen HN, Chang SK, Watts GF, Brenner MB. Genetic polymorphism directs IL-6 expression in fibroblasts but not selected other cell types. Proc Natl Acad Sci USA. 2015;112(48):14948–53. https://doi.org/10.1073/pnas.1520861112.
Babusikova E, Jurecekova J, Jesenak M, Evinova A. Association of gene polymorphisms in interleukin 6 in infantile bronchial asthma. Arch Bronconeumol. 2017;53(7):381–6. https://doi.org/10.1016/j.arbres.2016.09.012.
Withers AL, Green R. Transition for adolescents and young adults with asthma. Front Pediatr. 2019;23(7):301. https://doi.org/10.3389/fped.2019.00301.
Menzies-Gow A, Moore WC, Wechsler ME. Difficult-to-control asthma management in adults. J Allergy Clin Immunol Pract. 2022;10(2):378–84. https://doi.org/10.1016/j.jaip.2021.12.007.
Joseph C, Tatler AL. Pathobiology of airway remodeling in asthma: the emerging role of integrins. J Asthma Allergy. 2022;11(15):595–610. https://doi.org/10.2147/JAA.S267222.PMID:35592385;PMCID:PMC9112045.
Stubbs MA, Clark VL, McDonald VM. Living well with severe asthma. Breathe. 2019;15(2):e40–9. https://doi.org/10.1183/20734735.0165-2019.
Al-Ahmad M, Ali A, Khalaf M, Alterki A, Rodriguez-Bouza T. Comorbid asthma in patients with chronic rhinosinusitis with nasal polyps: did dupilumab make a difference? BMC Pulm Med. 2023;23(1):266. https://doi.org/10.1186/s12890-023-02556-8.
Kosugi EM, de Camargo-Kosugi CM, Hirai ER, Mendes-Neto JA, Gregorio LC, Guerreiro-da-Silva ID, Weckx LL. Interleukin-6 -174 G/C promoter gene polymorphism in nasal polyposis and asthma. Rhinology. 2013;51(1):70–6. https://doi.org/10.4193/Rhino12.166.
Kosugi EM, de Camargo-Kosugi CM, Weckx LL, Guerreiro-da-Silva ID, Gregorio LC. Interleukin-6-174 G/C promoter polymorphism and nasal polyposis. Rhinology. 2009;47(4):400.
Schneider AL, Schleimer RP, Tan BK. Targetable pathogenic mechanisms in nasal polyposis. Int Forum Allergy Rhinol. 2021;11(8):1220–34. https://doi.org/10.1002/alr.22787.
Palacios-García J, Porras-González C, Moreno-Luna R, Maza-Solano J, Polo-Padillo J, Muñoz-Bravo JL, Sánchez-Gómez S. Role of fibroblasts in chronic inflammatory signalling in chronic rhinosinusitis with nasal polyps-a systematic review. J Clin Med. 2023;12(9):3280. https://doi.org/10.3390/jcm12093280.
Hamid YH, Rose CS, Urhammer SA, Glümer C, Nolsøe R, Kristiansen OP, Mandrup-Poulsen T, Borch-Johnsen K, Jorgensen T, Hansen T, Pedersen O. Variations of the interleukin-6 promoter are associated with features of the metabolic syndrome in Caucasian Danes. Diabetologia. 2005;48(2):251–60. https://doi.org/10.1007/s00125-004-1623-0.
Pyrzak B, Wisniewska A, Majcher A, Popko K, Wasik M, Demkow U. Association between metabolic disturbances and G-174C polymorphism of interleukin-6 gene in obese children. Eur J Med Res. 2009;14(Suppl 4):196–200. https://doi.org/10.1186/2047-783x-14-s4-196.
Acknowledgements
Not applicable.
Funding
This research received funding from Kuwait University research sector, project number (M102/22).
Author information
Authors and Affiliations
Contributions
MA and AA have contributed equally to this study. AM worked on data collection, AA analyzed and interpreted the results and helped in writing up the paper. MH did the laboratory part and helped in writing up the paper. MA and AA were a major contributor in writing up the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The study was approved by the Ethics committee of Kuwait University and the Ministry of Health (Research project number MI02/22, 2022/2010). Informed consent has been obtained from all participants involved in the study, as well as their legal guardians, to ensure that they are fully aware of the nature and purpose of the research, and have given their voluntary and informed consent to participate.
Consent for publication
Not applicable.
Competing interests
All authors declare no competing interests. Each author has revised and approved the final version of the manuscript independently.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Al-Ahmad, M., Ali, A., Maher, A. et al. Association between interleukin-6-174G/C gene polymorphism and asthma severity: exploring the role of total serum IgE, blood eosinophils, and FeNO as markers of type 2 inflammation. Allergy Asthma Clin Immunol 20, 15 (2024). https://doi.org/10.1186/s13223-024-00880-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13223-024-00880-0