
Abstract
Background
Nasopharyngeal carcinoma (NPC) is prevalent in South China, including Hong Kong and Southeast Asia, constantly associated with Epstein-Barr virus (EBV) infection. Epigenetic etiology attributed to EBV plays a critical role in NPC pathogenesis. Through previous CpG methylome study, we identified Disheveled-associated binding antagonist of beta-catenin 2 (DACT2) as a methylated target in NPC. Although DACT2 was shown to regulate Wnt signaling in some carcinomas, its functions in NPC pathogenesis remain unclear.
Methods
RT-PCR, qPCR, MSP, and BGS were applied to measure expression levels and promoter methylation of DACT2 in NPC. Transwell, flow cytometric analysis, colony formation, and BrdU-ELISA assay were used to assess different biological functions affected by DACT2. Immunofluorescence, Western blot, and dual-luciferase reporter assay were used to explore the mechanisms of DACT2 functions. Chemosensitivity assay was used to measure the impact of DACT2 on chemotherapy drugs.
Results
We found that DACT2 is readily expressed in multiple normal adult tissues including upper respiratory tissues. However, it is frequently downregulated in NPC and correlated with promoter methylation. DNA methyltransferase inhibitor 5-aza-2′-deoxycytidine restored its expression in NPC cells. DACT2 methylation was further detected in 29/32 (91%) NPC tumors but not in any (0/8) normal nasopharyngeal tissue samples. Ectopic expression of DACT2 in NPC cells suppressed their proliferation, migration, and invasion through downregulating matrix metalloproteinases. DACT2 expression also induced G2/M arrest in NPC cells through directly suppressing β-catenin/Cdc25c signaling, which sensitized NPC cells to paclitaxel and 5-FU, but not cisplatin.
Conclusion
Our results demonstrate that DACT2 is frequently inactivated epigenetically by CpG methylation in NPC, while it inhibits NPC cell proliferation and metastasis via suppressing β-catenin/Cdc25c signaling. Our study suggests that DACT2 promoter methylation is a potential epigenetic biomarker for the detection and chemotherapy guidance of NPC.
Similar content being viewed by others


Background
Unlike other malignancies, the incidence of nasopharyngeal carcinoma (NPC) has great ethnic and geographic differences. Its incidence is high in Chinese and Malay populations in Southeast Asia and North Africa [1]. Specific biomarkers would be helpful in populations with a high incidence of NPC, but few are available. Study of NPC pathogenesis should aim to identify diagnostic biomarkers [2]. The Disheveled-associated binding antagonist of β-catenin (DACT) family, also known as Dapper/Frodo, are small intracellular scaffold proteins. There are three family members, DACT1, 2, and 3 [3]. DACT2 is repressed by promoter methylation in various cancers, including breast [3, 4], colon [5], lung [6], and gastric cancers [7], but the mechanisms differ. In breast cancer, our findings demonstrated that DACT2 antagonizes Akt/GSK-3 and Wnt/β-catenin signaling to suppress epithelial-to-mesenchymal transition (EMT) [3]. In glioma cells, DACT2 interacts with Wnt/β-catenin signaling to prevent Yes-associated protein translocation to the nucleus, resulting in its sequestration and degradation in the cytoplasm [8]. In esophageal squamous cell cancer, DACT2 suppresses TGFβ/SMAD2/3 activity via both the proteasome and lysosomal degradation pathways [9]. Zebrafish DACT2 was reported to inhibit TGF-β/Nodal signaling during mesoderm induction by interacting with type 1 receptors ALK5 and ALK4 and further promoting lysosomal degradation [10, 11].
In our recent study, DACT2 gene was identified to be a methylated target in NPC [2], but its molecular functions and mechanism were not determined. Here, we intend to investigate the expression and methylation of DACT2 in NPC cells and tissues. The effect of DACT2 on the cell cycle was evaluated to explore the influence of DACT2 overexpression on drug treatment.
Results
DACT2 was downregulated in NPC by promoter methylation
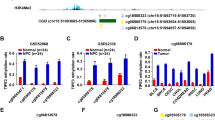
Reverse transcription (RT)-PCR confirmed that DACT2 was expressed in the majority of normal adult tissues (Fig. 1a). To investigate the expression of DACT2 in NPC, we analyzed the gene expression data of DACT2 in Oncomine online database (https://www.oncomine.org/), and it clearly shows that its expression is suppressed in the T1 and N0 stage NPC, which means DACT2 has potential to be an early diagnosed biomarker (Fig. 1b). DACT2 expression was downregulated in HNE1 and HONE1 NPC cells and was restored by 5-aza-2′-deoxycytidine (Aza) without or with trichostatin A (TSA). Following treatment, quantitative methylation-specific PCR (qMSP) showed a decrease of methylated level and an increase in un-methylated level (Fig. 1c). Thus, DACT2 expression was downregulated in these NPC cell lines by promoter methylation.

The promoter methylation causes DACT2 low expression in nasopharyngeal carcinoma cells. a DACT2 expression in human adult normal tissues detected by RT-PCR. b Expression of DACT2 was shown in the nasopharynx, and NPC is classified by T or N stage. Data was provided by Oncomine website. c The expression and methylation status of DACT2 were detected in HNE1 and HONE1 cells treated with Aza (A) without or with TSA (T) by qPCR and qMSP. d, e The methylation status of DACT2 in eight normal nasopharyngeal tissues (SD) and 32 nasopharyngeal cancer (NPC) tissues measured by MSP. M, methylated; U, unmethylated. f Methylation alleles of DACT2 measured by BGS in two normal nasopharyngeal tissues (SD) and two nasopharyngeal cancer (NPC) tissues
The methylation status of eight normal nasopharyngeal tissues and 32 NPC tissues was assayed by methylation-specific polymerase chain reaction (MSP), which found that the DACT2 promoter was not methylated in any of the normal nasopharyngeal tissues but was methylated in 29 of 32 (91%) NPC tissues (Fig. 1d, e). Bisulfite genomic sequencing (BGS) was used to assay methylated DACT2 promoter alleles in two normal nasopharyngeal tissue and two NPC tissue samples to confirm the result of MSP and found that DACT2 methylation was more frequent in NPC than in normal nasopharyngeal tissues (Fig. 1f).
Overexpression of DACT2 inhibited NPC cell proliferation, viability, and colony formation
The overexpression of DACT2 after DACT2 plasmid transfection was confirmed using RT-PCR and Western blot by comparing to empty control (Fig. 2a, b). The MTS assay (Fig. 2c) showed that cell viability was significantly reduced in DACT2-expressing cells. Colony formation (Fig. 2d) was also significantly suppressed compared with the control cells. These results indicated that DACT2 suppressed both viability and growth of NPC cells.

Overexpressed DACT2 suppress nasopharyngeal carcinoma cell proliferation ability. a, b The expression of DACT2 was detected in HNE1 and HONE1 cells stably transfected with vector (pcDNA3.1) or DACT2 by RT-PCR and Western blot. c The ability of cell proliferation was measured in vector- and DACT2-expressing HNE1 and HONE1 cells by MTS. d Representative images and the histogram statistics of the colony-formation assay in vector- and DACT2-expressed HNE1 and HONE1 cells. Mean ± SD, *p < 0.05, **p < 0.01, ***p < 0.001. All experiments were performed in triplicate, respectively
DACT2 induced G2/M cell cycle arrest and apoptosis in NPC cells
The influence of DACT2 on tumor cell proliferation might be mediated by its effects on the cell cycle and apoptosis. Flow cytometry of HONE1 and HNE1 cells found that the percentage of cells in the G2/M phase was increased in those that overexpressed DACT2 compared with controls transfected with an empty vector (Fig. 3a, b), accompanied by the increased cell population of S phase. Furthermore, the BrdU-ELISA assay, which reflects active DNA synthesis, revealed that the cell proliferation rate was decreased in DACT2-expressing cells (Fig. 3c). DACT2 overexpression also promoted cell apoptosis compared with controls (Fig. 3d, e). These results indicated that DACT2 inhibited cell proliferation by blocking the cell cycle in G2/M and by inducing cell apoptosis.

Overexpressed DACT2 induced G2/M cell cycle arrest and promote cell apoptosis in nasopharyngeal carcinoma cells. a, b The effect of DACT2 on cell cycle in vector- and DACT2-expressing HNE1 and HONE1 cells was detected by flow cytometry analysis. Representative flow cytometry plots (a) and the histogram statistics of cell cycle changes (b). c, BrdU-ELISA assay at 24 h in vector- or DACT2-transfected HNE1 and HONE1 cells. d, e The proportion of apoptotic cells was detected in vector- and DACT2-expressing HNE1 and HONE1 cells by flow cytometry analysis. Representative flow cytometry plots (d) and the histogram statistics of apoptosis changes (e)
DACT2 inhibited NPC cell migration and invasion
Wound healing and Transwell assays were used to investigate the influence of DACT2 expression on NPC cell migration and invasiveness. In the Transwell assay, significantly fewer DACT2-overexpressing cells passed through the membrane than control cells (p < 0.001) (Fig. 4a). Further wound healing assay revealed that scratches made in confluent layers of DACT2-overexpressing cells healed significantly slower than control cell layers over 24 h for HNE1 (p < 0.001) or 33 h for HONE1 (p < 0.01, Fig. 4b), which showed that DACT2 inhibited NPC cell migration. In the Transwell assay including a Matrigel barrier, DACT2 overexpression was associated with significant inhibition of NPC cancer cell invasion through the Matrigel before traversing the Transwell chamber membrane (p < 0.01, p < 0.001 at 24 h, Fig. 4c). qPCR and Western blot assays assessed the effect of DACT2 on expression of matrix metalloproteinases (MMPs) 2 and 9 (Fig. 4d, e), which are essential for cell migration and invasion [12, 13]. Overall, the results indicate that DACT2 suppressed cell migration and invasion in NPC cells.

Overexpressed DACT2 suppress nasopharyngeal carcinoma cell migration and invasion ability. a Representative image (left) and the histogram statistics (right) of Transwell cell migration assay in vector- and DACT2-expressing HNE1 and HONE1 cells, × 100 magnification (***p < 0.001). b Representative image (left) of wound healing assay in vector- and DACT2-expressed HNE1 and HONE1 cells. The distance of cell migration was collected to make the line chart statistics (right). c Representative image (left) and the histogram statistics (right) of Transwell cell invasion assay in vector- and DACT2-expressing HNE1 and HONE1 cells, × 100 magnification (**p < 0.01, ***p < 0.001). d, e Expression of MMP2 and MMP9 in vector- and DACT2-expressed HNE1 and HONE1 cells was detected by qRT-PCR and Western blot
DACT2 induced G2/M cell cycle arrest through the β-catenin/Cdc25c signaling pathway in nasopharyngeal cancer cells
Investigation of the mechanism of G2/M cell cycle arrest was based on a previous report that DACT2 suppressed β-catenin activity in colon cancer by competition for LEF1 binding [5]. In addition, the gene coding for cell division control protein 25C (Cdc25c), a regulator of G2/M cell cycle progression, has a LEF1 binding site on its promoter region. The function of that protein may thus be responsive to Wnt/β-catenin signaling activity [14, 15]. It is thus reasonable that DACT2 inhibits the activity of β-catenin/LEF1 complex by competitively binding with β-catenin followed by downregulation of Cdc25c, which ultimately blocks cell division in the G2/M phase. DACT2 regulation of the β-catenin/Cdc25c pathway to produce G2/M cell cycle arrest was assessed by immunofluorescent staining of β-catenin. As shown in Fig. 5a, the morphology of NHE1 cells overexpressing DACT2 differed from that of control cells (Fig. 5a), and the expression of total β-catenin did not significantly change but active β-catenin decreased in the cell nuclear location (Fig. 5b, c). qRT-PCR showed that DACT2 overexpression was associated with decreased expression of Cdc25c and cyclin B1 (Fig. 6a). Western blots revealed that DACT2 suppressed the expression of active β-catenin, Cdc25c, and the downstream target genes of β-catenin/Cdc25c signaling (Fig. 6b). A dual-luciferase reporter assay was performed to confirm whether DACT2 inhibited the activity of the β-catenin/LEF1 complex. It showed that DACT2 downregulated the induced TOPflash luciferase activities (Fig. 6c). Overall, the results indicate that DACT2 led to G2/M cell cycle arrest by inhibiting the β-catenin/Cdc25c signaling pathway.

Overexpressed DACT2 led HNE1 cell morphological alteration and decreased the expression of active-β-catenin. a HNE1 cell morphological alteration caused by DACT2 overexpression was observed by immunofluorescence. Upper—the cell shape became round and fewer thin fibers in DACT2-expressing cells but longer cell processes in control cells. Lower—double immunofluorescent staining also showed that DACT2-expressing cell became rounder and its pseudopods decreased. Photos were taken under × 400 magnification. b Localization and expression of endogenous total β-catenin was showed in DACT2-expressing HNE1 and HONE1 by immunofluorescence. c Immunofluorescence showed the localization and expression of active β-catenin in HNE1 transfected with DACT2 plasmid

Overexpressed DACT2 suppressed β-catenin/Cdc25c pathway in nasopharyngeal carcinoma cells. a Expression of CDC25C and CyclinB1 in vector- and DACT2-expressed HNE1 and HONE1 cells was detected by qPCR. b Expression of β-catenin and its target genes was measured by Western blot in vector- and DACT2-expressing HNE1 and HONE1 cells. c The activity of β-catenin/LEF1 complex was examined by dual-luciferase reporter assay in vector- and DACT2-expressed HNE1 and HONE1 cells
DACT2 induced sensitivity of NPC cells to paclitaxel and 5-FU but not cisplatin
As DACT2 expression led to G2/M cell cycle arrest, the impact of DACT2 overexpression on the sensitivity of NPC cells to cell cycle phase-specific and phase-nonspecific chemotherapy drugs was tested. Paclitaxel, 5-fluorouracil (5-FU), and cisplatin were selected. Paclitaxel acts by arrest of the cell cycle in G2/M [16]. 5-FU is an atypical periodic chemotherapy drug targeting on the S and other phases [17]. Cisplatin is cell cycle phase-nonspecific chemotherapy drug [18]. As shown in Fig. 7, DACT2 expression increased the sensitivity of HNE1 and HONE1 cells to paclitaxel and 5-FU compared with controls, but had no effect on cisplatin sensitivity (Fig. 7).

Overexpressed DACT2 sensitized nasopharyngeal carcinoma cells to paclitaxel, 5-FU rather than cisplatin. The cell viability was detected by CCK8 in vector- and DACT2-expressed HNE1 and HONE1 cells after the treatment of chemotherapy drugs paclitaxel or cisplatin for 48 h or 5-FU for 24 h. All experiments were performed in triplicate, respectively. *p < 0.05, **p < 0.01, ***p < 0.001
Discussion and conclusion
Promoter CpG methylation, which downregulates the expression of tumor suppressor genes, is essential to the pathogenesis of malignancies including NPC [19, 20]. Specific epigenetic therapy may increase the effectiveness of NPC treatment [21]. In papillary thyroid cancer [22] and hepatocellular cancer [23, 24], the expression of DACT2 is downregulated by promoter methylation. In this study, DACT2 was strongly expressed in normal adult tissues but weakly expressed and hyper-methylated in NPC cell lines. DACT2 expression was restored in NPC cell lines by Aza and TSA demethylation. Promoter methylation was detected in 29 of 32 (91%) NPC tissue samples but was not detected in any of the normal nasopharyngeal tissue samples. The results indicated that the low expression of DACT2 in NPC was caused by promoter CpG methylation. The function of DACT2 was investigated in HONE1 and HNE1 cells transfected with DACT2 gene. The restoration of DACT2 expression inhibited NPC cell proliferation, migration, and invasiveness and induced G2/M cell cycle arrest.
The Wnt signaling pathway is active in tumorigenesis, cell differentiation, and cell proliferation [25]. β-catenin is a transcription cofactor that induces target gene expression by binding to T cell factor/lymphoid enhancer factor (TCF/LEF) in the activated Wnt pathway [26]. The DACT family members are inhibitors of Disheveled, an important Wnt pathway component that suppresses c-Jun N-terminal kinase (JNK) signaling and the β-catenin cascades [27, 28]. DACT2 has been reported to decrease LEF1-β-catenin binding in colon cancer by competing with β-catenin [5]. In this study, DACT2 decreased the expression of active β-catenin and its downstream genes in NPC cells and suppressed the activity of the β-catenin/LEF1 complex. Cdc25c, which has been shown to regulate the G2/M checkpoint, has also been reported to have a functional LEF binding site [14, 15, 29, 30]. In NPC cells, DACT2 was found to suppress the expression of Cdc25c and its downstream genes related to G2/M arrest. We conclude that DACT2 downregulated the activity of the β-catenin/LEF1 complex by binding to β-catenin and that the decreased expression of Cdc25c induced G2/M arrest (Fig. 8). DACT2 overexpression increased the sensitivity of NPC cells to the cell cycle phase-specific chemotherapy drugs, paclitaxel and 5-FU.

Proposed models how DACT2 affects the β-catenin/Cdc25c pathway in nasopharyngeal carcinoma. When β-catenin/LEF1 complex is activated, it promotes the development of nasopharyngeal carcinoma by targeting its downstream genes including Cdc25c, which plays a crucial role in regulating cell cycle G2/M stage. DACT2 inhibits the activity of β-catenin/LEF1 complex by competitively binding to β-catenin and downregulates the expression of Cdc25c and its downstream genes, which can suppress nasopharyngeal carcinoma growth
In summary, DACT2 was silenced by promoter methylation as a tumor suppressor in NPC cells and induced G2/M phase arrest by the regulating β-catenin/Cdc25c signaling pathway. The results show that DACT2 is a tumor suppressor in nasopharyngeal cancer and support continuing evaluation of its value for early diagnosis and for targeted therapy of nasopharyngeal cancer.
Methods
Tumor cell lines and tissues
HNE1 and HONE1, two poorly differentiated nasopharyngeal squamous carcinoma cell lines, were used [19, 31]. Cells were maintained in RPMI 1640 (Gibco BRL, MD, USA) with 10% fetal bovine serum (Gibco, CA, USA) and 1% penicillin and streptomycin (Gibco BRL). Thirty-two nasopharyngeal cancer and eight normal nasopharyngeal tissue samples were obtained from Department of Otolaryngology of the First Affiliated Hospital of Chongqing Medical University between December 2010 and July 2013 [32,33,34]. All samples were verified by histology. All patients gave written informed consent.
RNA isolation, reverse transcription-PCR, and quantitative real-time PCR
Total RNA was extracted from cell lines and tissues using TRIzol Reagent (Invitrogen, Carlsbad, CA, USA) following the manufacturer’s instructions, and aliquots containing 1 μg of total RNA were reverse-transcribed to 20 μl cDNA. PCR was performed using Go-Taq (Promega, Madison, WI, USA) with initial denaturation at 95 °C for 2 min, followed by 32 cycles (95 °C for 30 s, 55 °C for 30 s, and 72 °C for 30 s) of amplification, with a final extension at 72 °C for 3 min [35] with β-actin used as a control. Twenty-three cycles of amplification were performed. The primer sequences are shown in Table 1. qPCR was performed using SYBR Green (Thermo Fisher) following the manufacturer’s instructions (7500 Real-Time PCR System, Applied Biosystems, Foster City, CA, USA). Each sample was tested in triplicate. Gene expression level was calculated by the 2−ΔΔCt method.
5-Aza-2′-deoxycytidine and trichostatin A treatment
Cell lines were treated with 10 μmol/L 5-aza-2′-deoxycytidine (Aza, Sigma-Aldrich, Steinheim, Germany), a DNA methyltransferase (DNMT) inhibitor, for 3 days and then without or with 100 nmol/L trichostatin A (TSA, Sigma-Aldrich) for 24 h as previously described [36].
DNA isolation, bisulfite modification of DNA, methylation-specific PCR, and quantitative methylation-specific PCR
Genome DNA was extracted from tissues using a QIAamp DNA Mini kit (Qiagen, Hilden, Germany) following the manufacturer’s instructions. Bisulfite modification of DNA was performed as previously described [37, 38]. The MSP primers (annealing temperature of 60 °C, 40 cycles) are shown in Table 1, and have been confirmed not amplify any nonbisulfited DNA [3]. MSP was performed using AmpliTaq-Gold DNA Polymerase (Applied Biosystems). The PCR products were identified on 2% agarose gels. qMSP was performed with the 7500 Real-Time PCR System (Applied Biosystems, Foster City, CA, USA) [39].
Bisulfite genomic sequencing
BGS primers (Table 1) were used to amplify bisulfite-treated DNA, and the PCR products were cloned into a pCR4-Top vector (Invitrogen). Eight to 12 colonies were randomly chosen and sequenced.
Construction of vector- and DACT2-expressed stable cell lines
Stable cell lines were constructed by transfecting cell lines with plasmids using Lipofectamine 2000 (Invitrogen) following the manufacturer’s instructions. pcDNA3.1 and pcDNA3.1–DACT2 plasmids were transfected at a concentration of 4 μg and selection by geneticin at 48 h after transfection [3]. Ectopic expression of DACT2 was assayed by RT-PCR and Western blotting prior to the other experimental procedures.
Cell viability assay
Cell viability was evaluated with a CellTiter 96 AQueous One Solution Cell Proliferation Assay (MTS, Promega) following the manufacturer’s instructions. HONE1 and HNE1 cells were cultured in 96-well plates after transient transfection with DACT2 and vector (pcDNA3.1) plasmids. Cell viability was measured at 0, 24, 48, and 72 h. Absorbance was read in a microplate reader at 490 nm. All experiments were performed in triplicate.
Colony formation assay
Cell proliferation was assayed by colony formation assay [40]. DACT2- and vector-expressing cells were plated in 6-well plates at densities of 200, 400, or 600 cells/well with geneticin. Surviving colonies (≥ 50 cells/colony) were counted on day 10 of culture after fixation and staining with Gentian violet. All experiments were performed in triplicate.
BrdU cell proliferation enzyme-linked immunosorbent assay
Cells were seeded in 96-well plates at 1 × 104 cells per well after transfection with DACT2 and vector (pcDNA3.1) plasmids for 48 h. After 24-h culture, BrdU (bromodeoxyuridine) was added to the wells to incorporate into proliferating cells for 4 h. The BrdU-ELISA assay was performed by BrdU Cell Proliferation ELISA Kit (colorimetric) (Abcam, Cambridge, UK) following the kit manufacturer’s instructions. The results were read at 450 nm using a microplate reader.
Flow cytometry
Flow cytometry was used for cell cycle analysis and to assay apoptosis [36, 41]. To assess cell cycle status, cells were stained with propidium iodide (PI) following transfection and fixation. For apoptosis, cells were double-stained with annexin V-fluorescein isothiocyanate and PI. The flow cytometry results were evaluated using a Cell Quest kit (BD Biosciences, CA, USA) and were performed in triplicate.
Wound healing, Transwell, and Matrigel assays
Cell migration was evaluated by wound healing and Transwell assays [42]. Stably transfected DACT2- and vector-(pcDNA3.1) HONE1 and HNE1 cells were plated in 6-well plates and were wounded when confluent by scratching with a sterile pipette tip. Migration was measured on phase-contrast micrographs (Leica DMI4000B, Milton Keynes, Buckinghamshire, UK) at 0, 12, and 24 h for HNE1 and 0, 11, 24, and 33 h for HONE1. Transwell chambers (Corning Life Sciences, Corning, NY, USA) with a pore size of 8 μm were used to evaluate cell migration and cell invasion. To assay cell invasiveness, a Matrigel (BD Biosciences) barrier was added on top of the Transwell membrane. Cells on the lower surface of the chamber at 24 h were photographed using a phase-contrast microscope (Leica) after fixation and staining and were then counted. All experiments were performed in triplicate.
Chemosensitivity assay
The effect of DACT2 on the cytotoxicity of paclitaxel, cisplatin, and 5-fluorouracil (5-FU) was assayed using a Cell Counting Kit-8 (CCK-8). Briefly, HONE1 and HNE1 cells transfected with DACT2 or vector (pcDNA3.1) plasmids were plated at 5000/well in 96-well plates. After cell attachment, culture media containing different concentrations of the tested drugs were added. Cells were then counted at 24 or 48 h with CCK-8 (Dojindo, Shanghai, China) following the manufacturer’s instructions. Absorbance was read with a microplate reader at 450 nm. The half-maximal inhibitory concentration (IC50) was calculated for each drug concentration. All experiments were performed in triplicate.
Dual-luciferase reporter assay
The effect of DACT2 on TCF/LEF transcriptional activities was investigated by a dual-luciferase reporter assay. pTopflash and pFopflash were used in our previous work [41]. pTopflash was constructed with TCF/LEF binding sites but pFopflash containing a mutant TCF/LEF binding sites as a control. HONE1 and HNE1 cells were transiently co-transfected with a pTOPflash or pFOPflash and DACT2 or vector (pcDNA3.1) with a Renilla luciferase reporter pRL-TK (Promega) as an internal control. Luciferase activity was measured after 48 h transfection using a dual-luciferase reporter assay kit (Promega) following the manufacturer’s instructions. All experiments were performed in triplicate.
Immunofluorescence staining
Cells were seeded in 24-well plates containing glass coverslips and then transfected with pcDNA3.1-DACT2 or pcDNA3.1 plasmid for 48 h. After transfection, cells were fixed with 4% paraformaldehyde in pH 7.4 PBS for 10 min, permeabilized for 10 min in 0.5% Triton X-100, and blocked with blocking buffer for 1 h at room temperature. Cells were incubated with primary antibodies against DACT2 (TA306668, Origene) and β-catenin (#2677, Cell Signaling Technology, Danvers, MA, USA) or β-actin (sc-8432, Santa Cruz Biotechnology, CA, USA), Flag-M2 (F3165, Sigma-Aldrich, Darmstadt, Germany) and non-p-β-catenin (#19807, Cell Signaling Technology) overnight at 4 °C. After primary antibody binding, cells were incubated with Alexa Fluor 594- or 488-conjugated goat anti-rabbit or anti-mouse secondary antibody (Jackson ImmunoResearch, West Grove, PA, USA) for an additional 30 min. Nuclei were counterstained with 4, 6-diamidino-2-phenylindole (DAPI, Roche, Palo Alto, CA, USA). The slides were observed with a confocal laser scanning microscope and photographed. Phalloidin staining was performed as our previous work [43]. Stable cells were used. Phalloidin-iFluor™ 594 Conjugate (23122, AAT Bioquest, CA, USA) at room temperature for 1 h.
Western blot assay
Western blotting was performed as previously described [31]. Aliquots of 40 μg of protein lysate were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred onto polyvinylidene difluoride (PVDF) membranes (Bio-Rad, Hercules, CA, USA). Membranes were incubated with DACT2 (TA306668, Origene), active β-catenin (#4270; Cell Signaling Technology), total β-catenin (#9562; Cell Signaling Technology), MMP9 (ab76003, Abcam), MMP2 (ab86607, Abcam), c-Myc (#13987, Cell Signaling Technology), Cyclin D1(sc-450), p-GSK3β (sc-373800), Cdc25c (sc-13138), Cdc2 (sc-54), p-Cdc2 (pY15.44) (sc-136014), β-actin (sc-8432) (all from Santa Cruz Biotechnology, CA, USA), or CyclinB1 (ab32053, Abcam) primary antibodies. Proteins were visualized using an Immobilon Western Chemiluminescent HRP Substrate kit (Millipore Corporation, Billerica, MA, USA).
Statistical analysis
SPSS16 (SPSS, Chicago, IL, USA) was used to perform the statistical analysis. Differences were evaluated for significance with the χ2 test and Fisher’s exact test. p values < 0.05 were considered statistically significant.
Abbreviations
- 5-FU:
-
5-Fluorouracil
- Aza:
-
5-Aza-2-deoxycytidine
- BGS:
-
Bisulfite genomic sequencing
- BrdU:
-
Bromodeoxyuridine
- CCK-8:
-
Cell Counting Kit-8
- Cdc25c:
-
Cell division control protein 25C
- DACT2:
-
Disheveled-associated binding antagonist of beta-catenin 2
- DAPI:
-
4, 6-diamidino-2-phenylindole
- DNMT:
-
DNA methyltransferase
- EBV:
-
Epstein-Barr virus
- ELISA:
-
Enzyme-linked immunosorbent assay
- EMT:
-
Epithelial-to-mesenchymal transition
- IC50:
-
Half-maximal inhibitory concentration
- JNK:
-
c-Jun N-terminal kinase
- MMPs:
-
Matrix metalloproteinases
- MSP:
-
Methylation-specific polymerase chain reaction
- NPC:
-
Nasopharyngeal carcinoma
- PI:
-
Propidium iodide
- PVDF:
-
Polyvinylidene difluoride
- qMSP:
-
Quantitative methylation-specific polymerase chain reaction
- qPCR:
-
Quantitative real-time polymerase chain reaction
- RT-PCR:
-
Reverse transcription polymerase chain reaction
- SDS-PAGE:
-
Sodium dodecyl sulfate polyacrylamide gel electrophoresis
- TCF/LEF:
-
T Cell factor/lymphoid enhancer factor
- TSA:
-
Trichostatin A
References
Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. 2015;65:87–108.
Li L, Zhang Y, Fan Y, Sun K, Su X, Du Z, Tsao SW, Loh TK, Sun H, Chan AT, et al. Characterization of the nasopharyngeal carcinoma methylome identifies aberrant disruption of key signaling pathways and methylated tumor suppressor genes. Epigenomics. 2015;7:155–73.
Xiang T, Fan Y, Li C, Li L, Ying Y, Mu J, Peng W, Feng Y, Oberst M, Kelly K, et al. DACT2 silencing by promoter CpG methylation disrupts its regulation of epithelial-to-mesenchymal transition and cytoskeleton reorganization in breast cancer cells. Oncotarget. 2016;7:70924–35.
Li J, Zhang M, He T, Li H, Cao T, Zheng L, Guo M. Methylation of DACT2 promotes breast cancer development by activating Wnt signaling. Sci Rep. 2017;7:3325.
Wang S, Dong Y, Zhang Y, Wang X, Xu L, Yang S, Li X, Dong H, Xu L, Su L, et al. DACT2 is a functional tumor suppressor through inhibiting Wnt/beta-catenin pathway and associated with poor survival in colon cancer. Oncogene. 2015;34:2575–85.
Jia Y, Yang Y, Brock MV, Zhan Q, Herman JG, Guo M. Epigenetic regulation of DACT2, a key component of the Wnt signalling pathway in human lung cancer. J Pathol. 2013;230:194–204.
Yu Y, Yan W, Liu X, Jia Y, Cao B, Yu Y, Lv Y, Brock MV, Herman JG, Licchesi J, et al. DACT2 is frequently methylated in human gastric cancer and methylation of DACT2 activated Wnt signaling. Am J Cancer Res. 2014;4:710–24.
Tan Y, Li QM, Huang N, Cheng S, Zhao GJ, Chen H, Chen S, Tang ZH, Zhang WQ, Huang Q, Cheng Y. Upregulation of DACT2 suppresses proliferation and enhances apoptosis of glioma cell via inactivation of YAP signaling pathway. Cell Death Dis. 2017;8:e2981.
Hou J, Liao LD, Xie YM, Zeng FM, Ji X, Chen B, Li LY, Zhu MX, Yang CX, Qing Z, et al. DACT2 is a candidate tumor suppressor and prognostic marker in esophageal squamous cell carcinoma. Cancer Prev Res (Phila). 2013;6:791–800.
Su Y, Zhang L, Gao X, Meng F, Wen J, Zhou H, Meng A, Chen YG. The evolutionally conserved activity of Dapper2 in antagonizing TGF-beta signaling. FASEB J. 2007;21:682–90.
Zhang L, Zhou H, Su Y, Sun Z, Zhang H, Zhang L, Zhang Y, Ning Y, Chen YG, Meng A. Zebrafish Dpr2 inhibits mesoderm induction by promoting degradation of nodal receptors. Science. 2004;306:114–7.
Sun B, Xu M. Matrine inhibits the migratory and invasive properties of nasopharyngeal carcinoma cells. Mol Med Rep. 2015;11:4158–64.
Bjorklund M, Koivunen E. Gelatinase-mediated migration and invasion of cancer cells. Biochim Biophys Acta. 2005;1755:37–69.
Suzuki A, Pelikan RC, Iwata J. WNT/β-catenin signaling regulates multiple steps of myogenesis by regulating step-specific targets. Mol Cell Biol. 2015;35:1763–76.
de Gooijer MC, van den Top A, Bockaj I, Beijnen JH, Wurdinger T, van Tellingen O. The G2 checkpoint-a node-based molecular switch. FEBS Open Bio. 2017;7:439–55.
Liebmann J, Cook JA, Lipschultz C, Teague D, Fisher J, Mitchell JB. The influence of Cremophor EL on the cell cycle effects of paclitaxel (Taxol) in human tumor cell lines. Cancer Chemother Pharmacol. 1994;33:331–9.
Takeda H, Haisa M, Naomoto Y, Kawashima R, Satomoto K, Yamatuji T, Tanaka N. Effect of 5-fluorouracil on cell cycle regulatory proteins in human colon cancer cell line. Jpn J Cancer Res. 1999;90:677–84.
Siddik ZH. Cisplatin: mode of cytotoxic action and molecular basis of resistance. Oncogene. 2003;22:7265–79.
Li LL, Shu XS, Wang ZH, Cao Y, Tao Q. Epigenetic disruption of cell signaling in nasopharyngeal carcinoma. Chin J Cancer. 2011;30:231–9.
Jones PA, Baylin SB. The epigenomics of cancer. Cell. 2007;128:683–92.
Tao Q, Chan AT. Nasopharyngeal carcinoma: molecular pathogenesis and therapeutic developments. Expert Rev Mol Med. 2007;9:1–24.
Zhao Z, Herman JG, Brock MV, Sheng J, Zhang M, Liu B, Guo M. Methylation of DACT2 promotes papillary thyroid cancer metastasis by activating Wnt signaling. PLoS One. 2014;9:e112336.
Zhang X, Yang Y, Liu X, Herman JG, Brock MV, Licchesi JD, Yue W, Pei X, Guo M. Epigenetic regulation of the Wnt signaling inhibitor DACT2 in human hepatocellular carcinoma. Epigenetics. 2013;8:373–82.
Gao S, Yang Z, Zheng ZY, Yao J, Zhang F, Wu LM, Xie HY, Zhou L, Zheng SS. Reduced expression of DACT2 promotes hepatocellular carcinoma progression: involvement of methylation-mediated gene silencing. World J Surg Oncol. 2013;11:57.
Nelson WJ, Nusse R. Convergence of Wnt, beta-catenin, and cadherin pathways. Science. 2004;303:1483–7.
Cadigan KM, Nusse R. Wnt signaling: a common theme in animal development. Genes Dev. 1997;11:3286–305.
Cheyette BN, Waxman JS, Miller JR, Takemaru K, Sheldahl LC, Khlebtsova N, Fox EP, Earnest T, Moon RT. Dapper, a Dishevelled-associated antagonist of beta-catenin and JNK signaling, is required for notochord formation. Dev Cell. 2002;2:449–61.
Waxman JS, Hocking AM, Stoick CL, Moon RT. Zebrafish Dapper1 and Dapper2 play distinct roles in Wnt-mediated developmental processes. Development. 2004;131:5909–21.
Deibler RW, Kirschner MW. Quantitative reconstitution of mitotic CDK1 activation in somatic cell extracts. Mol Cell. 2010;37:753–67.
Perdiguero E, Nebreda AR. Regulation of Cdc25C activity during the meiotic G2/M transition. Cell Cycle. 2004;3:733–7.
Li L, Tao Q, Jin H, van Hasselt A, Poon FF, Wang X, Zeng MS, Jia WH, Zeng YX, Chan AT, Cao Y. The tumor suppressor UCHL1 forms a complex with p53/MDM2/ARF to promote p53 signaling and is frequently silenced in nasopharyngeal carcinoma. Clin Cancer Res. 2010;16:2949–58.
Chen T, Long B, Ren G, Xiang T, Li L, Wang Z, He Y, Zeng Q, Hong S, Hu G. Protocadherin20 acts as a tumor suppressor gene: epigenetic inactivation in nasopharyngeal carcinoma. J Cell Biochem. 2015;116:1766–75.
He D, Zeng Q, Ren G, Xiang T, Qian Y, Hu Q, Zhu J, Hong S, Hu G. Protocadherin8 is a functional tumor suppressor frequently inactivated by promoter methylation in nasopharyngeal carcinoma. Eur J Cancer Prev. 2012;21:569–75.
Sung FL, Cui Y, Hui EP, Li L, Loh TK, Tao Q, Chan AT. Silencing of hypoxia-inducible tumor suppressor lysyl oxidase gene by promoter methylation activates carbonic anhydrase IX in nasopharyngeal carcinoma. Am J Cancer Res. 2014;4:789–800.
Ying J, Li H, Seng TJ, Langford C, Srivastava G, Tsao SW, Putti T, Murray P, Chan AT, Tao Q. Functional epigenetics identifies a protocadherin PCDH10 as a candidate tumor suppressor for nasopharyngeal, esophageal and multiple other carcinomas with frequent methylation. Oncogene. 2006;25:1070–80.
Xiang T, Li L, Yin X, Yuan C, Tan C, Su X, Xiong L, Putti TC, Oberst M, Kelly K, et al. The ubiquitin peptidase UCHL1 induces G0/G1 cell cycle arrest and apoptosis through stabilizing p53 and is frequently silenced in breast cancer. PLoS One. 2012;7:e29783.
Tao Q, Swinnen LJ, Yang J, Srivastava G, Robertson KD, Ambinder RF. Methylation status of the Epstein-Barr virus major latent promoter C in iatrogenic B cell lymphoproliferative disease: application of PCR-based analysis. Am J Pathol. 1999;155:619–25.
Tao Q, Huang H, Geiman TM, Lim CY, Fu L, Qiu GH, Robertson KD. Defective de novo methylation of viral and cellular DNA sequences in ICF syndrome cells. Hum Mol Genet. 2002;11:2091–102.
Husseiny MI, Kuroda A, Kaye AN, Nair I, Kandeel F, Ferreri K. Development of a quantitative methylation-specific polymerase chain reaction method for monitoring beta cell death in type 1 diabetes. PLoS One. 2012;7:e47942.
Yin X, Xiang T, Li L, Su X, Shu X, Luo X, Huang J, Yuan Y, Peng W, Oberst M, et al. DACT1, an antagonist to Wnt/beta-catenin signaling, suppresses tumor cell growth and is frequently silenced in breast cancer. Breast Cancer Res. 2013;15:R23.
Li L, Ying J, Tong X, Zhong L, Su X, Xiang T, Shu X, Rong R, Xiong L, Li H, et al. Epigenetic identification of receptor tyrosine kinase-like orphan receptor 2 as a functional tumor suppressor inhibiting beta-catenin and AKT signaling but frequently methylated in common carcinomas. Cell Mol Life Sci. 2014;71:2179–92.
Hu XT, Zhang FB, Fan YC, Shu XS, Wong AH, Zhou W, Shi QL, Tang HM, Fu L, Guan XY, et al. Phospholipase C delta 1 is a novel 3p22.3 tumor suppressor involved in cytoskeleton organization, with its epigenetic silencing correlated with high-stage gastric cancer. Oncogene. 2009;28:2466–75.
Mu H, Wang N, Zhao L, Li S, Li Q, Chen L, Luo X, Qiu Z, Li L, Ren G, et al. Methylation of PLCD1 and adenovirus-mediated PLCD1 overexpression elicits a gene therapy effect on human breast cancer. Exp Cell Res. 2015;332:179–89.
Acknowledgements
This study was supported by the National Natural Science Foundation of China (#81572769, #31420103915), Natural Science Foundation of Chongqing.
Municipal Commission of Health and Family Planning (#2016ZDXM006) and VC special research fund from The Chinese University of Hong Kong.
Availability of data and materials
The datasets analyzed during the current study are available in the Oncomine repository (https://www.oncomine.org/).
Author information
Authors and Affiliations
Contributions
YZ, TX, and QT contributed to the conception and design of the study. YZ, JF, DZ, XS, and JM performed the experiments and analyzed the data. XH and QX contributed to the RNA and DNA extraction. YY collected the samples. YZ and TX prepared the figures and drafted the manuscript. GR, LL, and YF reviewed the manuscript. TX and QT finalized the manuscript. All authors reviewed and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
This research was approved by the Institutional Ethics Committees of the First Affiliated Hospital of Chongqing Medical University (#20130306) and conformed to the tenets of the Declaration of Helsinki.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Zhang, Y., Fan, J., Fan, Y. et al. The new 6q27 tumor suppressor DACT2, frequently silenced by CpG methylation, sensitizes nasopharyngeal cancer cells to paclitaxel and 5-FU toxicity via β-catenin/Cdc25c signaling and G2/M arrest. Clin Epigenet 10, 26 (2018). https://doi.org/10.1186/s13148-018-0459-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13148-018-0459-2