
Abstract
Background
Phosphocreatine dynamics provide the gold standard evaluation of in-vivo mitochondrial function and is tightly coupled with oxygen availability. Low mitochondrial oxidative capacity has been associated with health issues and low exercise performance.
Methods
To evaluate the relationship between near-infrared spectroscopy-based muscle oxygen dynamics and magnetic resonance spectroscopy-based energy-rich phosphates, a systematic review of the literature related to muscle oxygen dynamics and energy-rich phosphates was conducted. PRISMA guidelines were followed to perform a comprehensive and systematic search of four databases on 02-11-2021 (PubMed, MEDLINE, Scopus and Web of Science). Beforehand pre-registration with the Open Science Framework was performed. Studies had to include healthy humans aged 18–55, measures related to NIRS-based muscle oxygen measures in combination with energy-rich phosphates. Exclusion criteria were clinical populations, laboratory animals, acutely injured subjects, data that only assessed oxygen dynamics or energy-rich phosphates, or grey literature. The Effective Public Health Practice Project Quality Assessment Tool was used to assess methodological quality, and data extraction was presented in a table.
Results
Out of 1483 records, 28 were eligible. All included studies were rated moderate. The studies suggest muscle oxygen dynamics could indicate energy-rich phosphates under appropriate protocol settings.
Conclusion
Arterial occlusion and exercise intensity might be important factors to control if NIRS application should be used to examine energetics. However, more research needs to be conducted without arterial occlusion and with high-intensity exercises to support the applicability of NIRS and provide an agreement level in the concurrent course of muscle oxygen kinetics and muscle energetics.
Trial registration
Key points
1. NIRS derived measures of muscle oxygenation agree with gold-standard measures of high energy phosphates when assessed in an appropriate protocol setting.
2. At rest when applying the AO protocol, in the absence of muscle activity, an initial disjunction between the NIRS signal and high energy phosphates can been seen, suggesting a cascading relationship.
3. During exercise and recovery a disruption of oxygen delivery is required to provide the appropriate setting for evaluation through either an AO protocol or high intensity contractions.
Similar content being viewed by others
Introduction
Mitochondria are complex filamentous organelles that are the primary structures responsible for cellular energetics [1, 2]. Mitochondria use electrons from substrate oxidation to shift protons within the electron transport system, causing a chemiosmotic gradient and driving ATP production as an energy currency. The last electron acceptor is oxygen, concluding that there is no substrate oxidation and no energy flux during energy demands without oxygen [3, 4]. Near-infrared spectroscopy (NIRS) can be used in vivo as a non-invasive and simple assessment to measure the oxygen consumption rate in skeletal muscle [5,6,7].
As shown, after muscle activity or ischemic states, phosphocreatine (PCr) recovery depends on oxidative ATP production by mitochondria [8,9,10]. Consequently, the dependence of PCr recovery on oxidative ATP production and overall oxygen availability may indicate a close correlation between oxygen availability and energy-rich phosphates. During activity, ATP levels in the muscle are buffered through PCr, dynamically depleted and synthesized through creatine kinase. Under high ATP demands, mitochondrial respiration is stimulated by rising ADP concentrations, reflected by PCr concentrations in the muscle [11, 12]. Assuming creatine kinase equilibrium, the recovery rate of PCr after exercise represents mitochondrial ATP production [13, 14]. These changes can be measured via phosphorus nuclear magnetic resonance spectroscopy (P-MRS) and provide quantifiable information about phosphocreatine, ATP, and inorganic phosphate (Pi). Overall, mitochondrial function [15, 16] and a higher oxidative capacity [17,18,19,20] are associated with enhanced performance, from patients with chronic illness to endurance athletes. Since magnetic resonance spectroscopy (MRS) is expensive, limited in availability, and complex movements of healthy individuals are not feasible, NIRS has garnered more interest in assessing oxidative capacity, comparing muscle oxygen kinetics and changes in energy-rich phosphates [6, 7]. Moreover, muscle oxygen kinetics are increasingly measured in both laboratory and applied sports settings to assess muscle performance and training status [21]. However, studies regarding oxygen dynamics differ in their approaches and show contradictory results that do not consistently demonstrate the connection between muscle oxygen dynamics and energy-rich phosphates [6, 7, 21, 22].
Perrey & Ferrari [21] previously conducted a systematic review on muscle oximetry in sports science, presenting the development of the application of oximetry in sports during the past 35 years. The authors highlighted the need for a more in-depth comparison of physiological parameters to show the advantages of the routine use of muscle oximetry [21]. Nonetheless, Campbell and Marcinek [23] discussed nuclear MRS and optical measurements for in vivo evaluations of mitochondrial biogenetics in their narrative review and provided an overview of some possible approaches. The authors submitted that NIRS could represent an alternative to P-MRS for measuring mitochondrial capacity in skeletal muscle and emphasize this novel approach’s development, validation, and application.
A better and more comprehensive understanding of the relationship between corresponding muscle oxygen values measured by NIRS and energy-rich phosphates during rest, activity, or ischemic assessments will be helpful in the development of non-invasive continuous examinations of integrated energetics in athletes.
A preliminary literature search showed that there is currently no systematic summary regarding the relationship between muscle oxygenation measured by NIRS and phosphate synthesis and resynthesis. This review will favor a practical perspective on an integrated and dynamic evaluation of energetics and analyses of oxygenation data solely from muscle oxygen kinetics and phosphate-related measurement methods. Therefore, this review examines how muscle oxygenation data measured by NIRS represents energy-rich phosphates according to published research to address whether NIRS can be useful in determining energy-rich phosphates. As will become evident, apart from a systematic literature review the results are narrative in nature, as large methodical and technical variation makes any systemic analysis or modeling difficult.
Methods
The study was conducted according to the Preferred Reporting Items for Systematic Reviews and Meta-Analysis (PRISMA) statement [24]. The protocol was pre-registered on the Open Science Framework before full searches and analysis (https://osf.io/py32n). Since the current study reviewed published studies, ethical approval or patient consent was unnecessary.
Search strategy and study identification
Literature was searched in PubMed/MEDLINE, Scopus, and Web of Science. All searches were conducted on February 11, 2021. The Boolean method was used with the operators AND/OR/NOT to limit the results to relevant studies containing the following search terms: “near-infrared spectroscopy”, “NIRS”, “SmO2”, “HHb”, “O2Hb”, “TSI”, “mVO2” or “tHb” in combination with “phosphate”, “phosphorus”, “creatine phosphate”, “phosphocreatine”, “adenosinetriphosphate”, “adenosine triphosphate”, and “phosph*”. Reference lists from the selected studies were screened for other relevant studies; duplicates and studies known to the authors were also included. Two authors (KM and AF) have independently screened the titles and abstracts for relevance. The two mentioned authors examined the remaining full texts based on the inclusion and exclusion criteria. In the event of discrepancies arising, a third author (RJ) provided an examination of the relevant articles to reach a consensus decision. The detailed search strategies for each database are available in a supplementary file (Supplemental material, https://osf.io/7u4ym/?view_only=1048d187f51c4cdb970bb6dc6632738d).
Selection criteria
Inclusion
Studies eligible for the present review article had to [1] be published in the English language, [2] have abstracts available for screening, [3] include relevant data concerning the relationship of oxygenation data and energy-rich phosphates, [4] include human subjects at age 18–55 years. There was no limitation to publication date since the first NIRS studies were published in 1977 and contributed to the understanding of the development of NIRS measurements [25].
Exclusion
Studies were excluded if one of the following criteria were met: (1) acute injured and other clinical population, (2) laboratory animals, (3) in vitro or in situ experiments (muscle biopsies, high-resolution respirometry, measuring enzymatic activity), (4) data only assessing NIRS, (5) dietary intervention, (6) non-research letters and editorials, case studies, case series, grey literature - such as theses and dissertations -, abstracts or congress communications were excluded, as well as epidemiological, commentaries, literature, narrative, and systematic reviews.
Data extraction and quality assessment
One author (KM) extracted the relevant data from each included study using predesigned table forms on Microsoft Excel. A second author (AF) confirmed the results. Discrepancies between authors were resolved by the third author (RJ). For examining the methodological quality of all included studies, the Effective Public Health Practice Project Quality Assessment Tool (EPHPP) (Armijo-Olivo et al., 2012) was used and performed independently by two reviewers (KM & AF). The EPHPP assess the analytical cross-sectional and intervention studies, which will include the comparisons of the measurement tools. All questions had to be rated as strong (3 points), moderate (2 points), or weak (1 point), and domain scores were averaged to provide the total score. The maximum total score per study was 3.00. Based on their total score, studies were assigned a quality rating of weak (1.00–1.50), moderate (1.51–2.50) or strong (2.51–3.00).
Results
Study selection
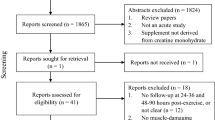
As shown in Fig. 1, 2722 records were identified in the four electronic databases (MEDLINE/Pubmed, Scopus, and Web of Science). One thousand two hundred forty-seven duplicates were removed, and 1483 remained for screening. Forty-one studies were considered relevant after assessing the title and abstract inspection. Full-text screening showed 28 articles included in the systematic review (Fig. 1).

PRISMA flow diagram of the selection process of the journal articles included in the systematic review
Study characteristics
The main characteristics of included studies are presented in Table 1. Studies were published between 1994 and 2020. Exercise intensity in the studies varied from resting to maximal intensity. Muscles of the lower and upper extremities and lumbar were included. Moreover, arterial occlusion (AO) was applied in 15 studies [6, 26, 28,29,30, 32, 34,35,36, 38, 45, 49, 50, 52]. Two studies included data on ischemia in the absence of exercise [28, 29]. All other studies involve training and/or post-exercise recovery. The study populations were heterogeneous, ranging from sedentary to highly active participants. The sample size ranged from four to 50 participants.
Quality of studies
EPHPP tool was conducted to assess the cross-sectional and intervention studies. The complete analysis is reported in Table 2. The mean quality score for the 28 articles was 2.15 ± 0.23, thereby moderate. All articles achieved a moderate score. The withdrawals and dropouts category did not apply to most studies because a lower dropout risk exists in crossover study designs. Studies have been rated between weak and strong when other techniques were applied.
Discussion
Our systematic review of 28 studies identified research investigating energy-rich phosphate changes and NIRS during rest, exercise, and ischemia. All articles achieved a moderate score (Range = 1.80–2.50). Since the relationship between two physiological variables were compared in the studies, most included studies assessed the variables within a cross-sectional study design. This led 18 out of 28 studies to a low grading concerning the study design. Although observational study design is appropriate to infer a relationship [53], the quality appraisal tool tended to produce lower quality scores for those studies.
Overall, the results of this review were inconclusive. No consistent relationship between oxygen levels and PCr were identified. Measurement protocols were heterogeneous and had a low sample size. Additionally, heterogeneous formulas for NIRS-based variables were used, making it challenging to compare oxygen kinetics between studies. Because of the high heterogeneity of NIRS-based variables, the categorization of variables was not applied. Nevertheless, considering the current state of knowledge, the application of NIRS to determine changes in energy-rich phosphates would appear appropriate when certain conditions are met.
Energy rich phosphates and NIRS at rest with and without AO
Differences in oxygen kinetics and the time course of high-energy phosphates during rest with AO have been observed. In general, the absence of blood flow with an AO was used to decouple muscle oxygen delivery from oxygen consumption and focus on muscle oxygen consumption. Hamaoka et al. [26] described the first observations at rest. Muscle oxygen declines in a linear fashion attenuating at 240 sec and leading to a plateau at 360 sec. Particularly at rest, these observations can vary with low metabolic rate and high individual variation; for this reason, muscle activation is included in the protocol to stimulate metabolism. When O2 was measured to be insufficient to maintain ATP level, PCr decreased, broken down to maintain ATP level for resting metabolism [26]. This can only be examined if a sufficient gradient is reached between blood vessels and the tissue or mitochondria. Furthermore, Sako et al. [32] observed a decrease in PCr as soon as Hb reached a plateau in AO resting measurements 5 min after the start of AO. Notably, when AO was applied for 5 min periods during rest in the study by Binzoni & Hildebrand [28], PCr and ATP concentrations did not change. In contrast to the constant PCr and ATP levels at rest, a constant change in desaturation was observed during the applied ischemic periods. It can be concluded that at rest under an extended period of ischemia (i.e., no longer than 5 min), no change in PCr occurs, even if changes in NIRS-related values are observed. This can be extrapolated because if sufficient oxygen levels are available to support oxidative metabolism, high resting PCr can be maintained. As oxygen levels decline, this is no longer the case. Therefore, during short periods of ischemia at rest, NIRS will not reflect PCr.
The deoxygenation rate of NIRS-O2 has been shown to be a function of work rate [54], and during ischemia at rest and exercise, 2 to 8-fold increases have been documented [6, 30]. Since pH in skeletal muscle decreases only when coupled to muscular contraction, the glycolysis rate is assumed to be low at rest [55] and increases exponentially with the onset of exercise. Hence, the authors attempted to explain that oxygen stores, namely Hb and Mb, could have the capabilities to maintain tissue metabolism during AO at rest [30].
Energy-rich phosphates and NIRS during exercise
A few studies measured NIRS and MRS simultaneously during exercise. Without AO rate of deoxygenation (time constant: 42 ± 12.5 sec) was in line with the time constant of the decrease in PCr (48.2 ± 10.2 sec) at the onset of intermitted contraction exercise (5 sec isometric contractions and 5 sec relaxation) at 50% of MVC [33]. Additionally, to show that at 50% of MVC exercise can potentially occlude arterial blood flow, Hamaoka et al. [33] compared a 10 sec occluded isometric hold with a non-occluded trial. The rate of deoxygenation was similar in both trials (2.23 ± 1.25%/sec versus 2.13 ± 1.18%/sec), hence in their preliminary study, Hamaoka and colleagues [33] were the first that demonstrates that the rate of muscle deoxygenation by NIRS during an intermittent exercise at 50% of MVC without AO indicates only muscle oxygen consumption.
The biochemical effect of extensive ATP hydrolysis from exercise resulting in increasing proton accumulation and consequently a shift in pH [27, 37, 47, 48, 56] is an important factor to consider when looking at NIRS and PCr response during exercise. In a study by Willcocks and colleagues [43] using constant-work rates without pH changes, HHb and PCr were not associated; PCr decreased from rest to end-exercise regardless of HHb responses. However, when exercise intensity increases and thus significant changes in pH occur, ATP production increased approximately twofold when MVC was elevated from 15 to 30% in rhythmic handgrip exercise when using either MRS or NIRS data to calculate high energy phosphates [30]. In a ramp test, energy-rich phosphate changes and HHb show similar behavior [49]. During 10, 20, and 30% of MVC, NIRS data can predict the course of PCr [49]. Nevertheless, no direct comparison between PCr and HHb was undertaken. From a physiological perspective, at higher intensities above 50% of MVC, an increased glycolytic rate could decrease the ADP effect on oxidative phosphorylation and, therefore, the availability of intramuscular O2 in mitochondria [37, 38, 46]. Accordingly, when lower intensity exercise is performed (e.g., at 30% of MVC), intramuscular oxidative metabolism matters more and curtails both PCr and glycolysis as energy suppliers [31, 38, 44]. Thus, visual inspection of data, quantitative analysis, and physiological explanations support the assumption that exercise intensity similarly impacts the rate of decline of muscle oxygenation and PCr. Considering this fact, we support the hypothesis that the change of PCr is greatly influenced oxygen availability [8,9,10].
PCr and muscle oxygen kinetics can be affected by prior ‘priming’ exercise, dependent on intensity. In particular, shifts in pH influence PCr kinetics [57]. PCr recovery follows a biexponential function with two distinct phases, an initial and a slow component [58]. While it is unclear what the exact mechanisms of control (i.e., cytosolic ADP) of the initial phase are, the slow component would appear to be tightly coupled to pH. This is a reasonable outcome considering the role of H+ in creatine kinase equilibrium and, therefore, the mass-action ratio. Priming can increase muscle oxygenation in a subsequent high intensity exercise bout with pH level and muscle efficiency influencing PCr hydrolysis. This may lead to a larger reduction in PCr [39]. Though an increase in muscle oxygen could be seen, PCr decreased faster and thereby dissociated PCr kinetics from muscle oxygen kinetics. Forbes and colleagues [40] examined the influence of the recovery time between two high intensity exercise bouts with 3 min, 6 min, and 15 min. PCr slow component was similarly reduced in all three bouts, whereas HHb onset kinetics were only slowed in the 3 min bouts. During the mid to later (approx. 2–6 minute) course of the second exercise bout, independent of the recovery time, PCr was significantly higher than in the first, indicating changed metabolism regardless of the recovery time. HHb was elevated only in the exercise bout after 3 min recovery. As a result, this led to the assumption that PCr consumption is not influenced by O2 delivery, fatigue mechanisms, PCr level or intracellular acidosis at the onset of the second exercise bout [40].
Contrastingly, Layec et al. [42] reported an increased deoxygenation amplitude, a decrease in PCr breakdown, and a decrease in glycolysis in consecutive bouts of high-intensity knee-extension exercise, illustrating a shift from glycolytic to oxidative ATP production in the second exercise bout. A reduced ADP stimulus was proposed as the reason for the shift since, at the end of the second exercise bout oxygenation level was increased, and the ADP level decreased. This could indicate higher intracellular O2 extraction for higher ATP production rates with concurrent lower ADP levels [42]. Although a coupling of NIRS data with PCr behavior was observed in this study, the authors argued that the control of oxidative phosphorylation could rely on the more complex relationship between PCr and muscle oxygen kinetics and essentially be influenced by additional factors such as phosphate potential, pH, and ADP [42]. Given these contradictions, there appears to be room for interpretation regarding how exercise and priming influence muscle oxygen kinetics and phosphate metabolism.
Further, when exercise is introduced under AO, metabolic stress is indicated by both PCr and NIRS parameters [41, 50]. Blood flow restriction intervention from Yanagisawa et al. [50] revealed that both PCr and Hb were significantly changed when AO occurred. Moreover, during the four-minute course of the exercise, the comparison of restricted versus non-restricted blood flow constituted a significant difference in the decrease of Hb from the very first minute onward; in fact, PCr decreases were significantly different from the second minute onward. These findings support the assumptions that insufficient O2 supply as a result of AO occurs, ATP supply could increasingly depend on oxygen stores and glycolytic metabolism. Nevertheless, the study did not directly compare the course of Hb and PCr and, thus, limits further conclusions.
Energy rich phosphates and NIRS during recovery
Most studies in the present review included recovery measurements. McCully et al. [22] were the first to assess the relationship between PCr and Hb saturation. Their results indicated a similar course of PCr and Hb after exercise. Additionally, their results demonstrated a decoupling of Hb and PCr when exercise intensity was increased to maximal bouts intensity [22]. Consequently, the potential benefit of the function of reoxygenation kinetics as an indicator of PCr recovery kinetics during recovery periods may be limited to low intensity exercise with controlled pH (approximate pH of 7.0).
Moreover, in high oxidative capacity muscle, it was shown that after a short duration, high intensity isometric exercise, the reoxygenation rate during the recovery phase is slower due to greater O2 utilization capacity [34, 35]. In particular, these observations were made when O2 demand was maximized at high intensity exercise. Results of Kime et al. [34, 35] and Hamaoka et al. [33] show, contrary to McCully et al. [22], that higher intensities are necessary to create a bridge between NIRS and MRS-based PCr. This is likely due to the degree to which NIRS is removed from the direct measurement of PCr, and oxygen dynamics functions as a limited surrogate. It was shown that muscle oxygen consumption at 90% of MVC was significantly higher than at 30 and 60% of MVC. Additionally, the correlation between Tc of PCr and HbO2 reoxygenation rate could only be found at 90% of MVC and not at 30 and 60% of MVC [35]. Consequently, at higher intensities above approximately 50% of MVC, an increase in glycolytic metabolism could decrease the ADP effect on oxidative phosphorylation and, therefore, the availability of intramuscular O2 in mitochondria [38].
There may be room here for a confounder in the relationship between PCr breakdown and recovery, and muscle oxygenation kinetics, which may clarify some of the contradiction. Difference in muscle oxygen on and off-set kinetics as a result of fitness may be the factor determining the goodness of fit between muscle oxygenation and PCr [22]. The higher the fitness the smaller the O2 supply and demand gap at the onset and offset of exercise, as shown by VO2 kinetics [59], and therefore the better the relationship between deoxygenation and reoxygenation and PCr kinetics. Finally, Heskamp and colleagues [51] showed that the close relation between PCr and oxygen supply remains independent of spatial variation. The oxygen supply increased from distal to proximal along the tibialis anterior muscle, just as the PCr recovery rate did. A strong correlation between Hb recovery rate constant and PCr recovery rate constant confirmed the relationship (r = .956 and r = .852).
A calculation of ATP change based on NIRS variables was demonstrated by Hamaoka et al. [26] and Sako et al. [32]. Hamaoka et al. [26] and Sako et al. [32] calculated muscle metabolic rate based on NIRS values and expressed it as millimolar ATP per second. Significant correlation occurred between NIRS-based muscle O2 consumption in absolute terms with ADP and PCr [26] and between NIRS-based muscle oxidative metabolic rate with PCr resynthesis rate post exercise [32]. The results provide evidence that NIRS could quantitively indicate the rate of muscle oxidative metabolism. The calculation of muscle consumption and muscle oxidative metabolic rate in both studies was similarly achieved (Hb decline rate with AO after exercise)/(Hb rate with AO at rest) × (Resting metabolic rate measured by P-MRS). The equation illustrates the dependence of these methods on P-MRS data. In the absence of P-MRS data, NIRS data alone cannot complete these calculations therefore limiting their applications.
In contrast, recovery constants only by NIRS have been experimentally validated against P-MRS independent of P-MRS [6, 36]. These time constants correlated well (r2 = .88–.95) and agreed in Bland-Altman Plots. The authors demonstrated that the indices solely based on NIRS could provide information about the muscle oxidative capacity.
Limitations
The present study has two major limitations. First, many of the studies evaluated applied time constants or complex calculated indices to provide information about muscle metabolism rather than to compare the direct course of PCr and muscle oxygen kinetics during exercise. The heterogeneous data offered in the review provides evidence for a relationship between PCr and muscle oxygen kinetics when appropriate protocol settings are used, as correlations confirm (Table 1). Essential conditions for study protocols are measurement methods, intensity of priming exercise, recovery time, and intracellular pH level. Thus, comparison and interpretation of the results are strongly limited. Second, this study aimed to obtain information about the application of NIRS to examine integrated energetics in athletes. However, NIRS can only measure the balance between O2 consumption and delivery. This limits its scope of application, and the authors mainly cited used the AO to calculate oxygen consumption and derive from that phosphate levels. AO restricts the practical application of NIRS to be integrated into dynamic measurement and diagnostics. Hamaoka et al. [33] were the only ones to examine the necessity of AO above 50% of MVC. Based on their assumptions, it could be possible to solely measure the muscle oxygen consumption with high intensity exercises without AO and, thus, indicate the course of PCr during everyday sport science settings. Because of the significant correlations between PCr and the rate of deoxygenation (r = .96, p > .05) and the agreement of the time constant of the rise of the rate of deoxygenation and the time constant of the decrease in PCr, it seems that NIRS has the potential to indicate PCr during exercise. Though first quantitative measures were provided, further investigations would be beneficial to support those findings and strengthen the evidence of muscle oxygen kinetics as an indicator of energy rich phosphates in an applied sport science context.
Future directions
In future studies a greater emphasis should be placed on the integration of NIRS with P-MRS to leverage the strengths of both measurements. Exercise, in particular strenuous exercise, involves low oxygen conditions and NIRS is uniquely placed to characterize oxygen levels during such exercise. P-MRS on the other hand is uniquely designed to measure changes in glycolysis and muscle pH, which is important in understanding strenuous exercise. While not covered in this review, joining NIRS measures with other MRS or MRI techniques like 1H-MRS may provide a more comprehensive evaluation of muscle energetics, by exploiting the different techniques inherent strengths and weaknesses [60,61,62]. In this way future studies can be classified into three general categories, those that include NIRS as a necessity for context with MRS and MRI data collection, those that include NIRS as an extension to the findings of the MRS/MRI findings, and those that apply NIRS independently or as surrogate of MRS/MRI findings. While true for all three categories, the third category in particular lends itself to combining NIRS and other noninvasive assessments to answering very practical and in situ questions, given the increasing portability of NIRS and other noninvasive tools. For example, the combination of NIRS to assess muscle metabolism and triaxial accelerometry to measure muscle movements and position in space; or with electromyography (EMG) to measure muscle activation and metabolism. Including a multi-site NIRS measurements with assessments of whole-body oxygen consumption would seem to greatly advance our understanding of whole body movement and metabolism. In summary, muscle responses to exercise are a fundamental part of the how the body addresses questions exercise and intensity, and future research should include both comprehensive assessments of muscle function with multimodal approaches, as well as assessments of local and whole body responses to exercise.
Conclusion
In conclusion, 28 studies have included both NIRS derived muscle oxygen kinetics and energy rich phosphates. The results suggest that the application of NIRS can indicate the change in energy rich phosphates when assessed in an appropriate protocol setting. High intensity exercise may be necessary, such that oxygen delivery is disrupted. For this reason, the AO method or high intensity exercise should be included in NIRS-based studies evaluating high energy phosphates. The heterogeneity of the data, protocols and interoperations limit the interpretation of the data. More research needs to be conducted using MRS and NIRS without AO and with high intensity exercise to demonstrate an agreement in the concurrent course of muscle oxygen kinetics and muscle energetics.
Availability of data and materials
All data generated or analysed during this study are included in this published article.
Abbreviations
- AO :
-
Arterial occlusion
- BFR :
-
Blood flow restriction
- HbO 2 /O 2 Hb/oxy-Hb/Oxyhemoglobin :
-
Oxygenated hemoglobin/myoglobin
- HHb/deoxy-Hb :
-
Deoxygenated hemoglobin/myoglobin
- HbO 2 Tc :
-
Time constant for oxygenated hemoglobin/myoglobin
- Hb-Mg :
-
Hemoglobin and myoglobin
- Hb :
-
Hemoglobin
- Hb tot :
-
Total hemoglobin
- ∆HbO 2 :
-
Change in oxygenated hemoglobin
- ∆Hb tot :
-
Change in total hemoglobin
- MRI:
-
Magnetic resonance imaging
- MRS:
-
Magnetic resonance spectroscopy
- mVO 2 :
-
Muscle oxygen consumption
- NIRS :
-
Near-infrared spectroscopy
- PCr:
-
Phosphocreatine
- PCrTc :
-
Time constant for phosphocreatine
- Pi :
-
Inorganic phosphate
- P-MRS :
-
Phosphorus nuclear magnetic resonance spectroscopy
- Reoxy-rate :
-
Rate of oxygenated hemoglobin/myoglobin increase
- Tau :
-
Time constant when value has reached 63% of the resting value
- Tc :
-
Time constant
- TOI :
-
Tissue oxygenation
- VO 2mus :
-
Muscle oxygen consumption measured with deoxygenation rate of hemoglobin and myoglobin
- VIB :
-
Vibration
References
Popov V, Medvedev NI, Davies HA, Stewart MG. Mitochondria form a filamentous reticular network in hippocampal dendrites but are present as discrete bodies in axons: a three-dimensional ultrastructural study. J Comp Neurol. 2005;492(1):50–65.
Willingham TB, Ajayi PT, Glancy B. Subcellular specialization of mitochondrial form and function in skeletal muscle cells. Front cell Dev Biol. 2021;9:757305.
Connett RJ, Honig CR, Gayeski TE, Brooks GA. Defining hypoxia: a systems view of VO2, glycolysis, energetics, and intracellular PO2. J Appl Physiol. 1990;68(3):833–42.
Shulman RG, Rothman DL. The “glycogen shunt” in exercising muscle: a role for glycogen in muscle energetics and fatigue. Proc Natl Acad Sci U S A. 2001;98(2):457–61.
Hamaoka T, McCully KK, Quaresima V, Yamamoto K, Chance B. Near-infrared spectroscopy/imaging for monitoring muscle oxygenation and oxidative metabolism in healthy and diseased humans. J Biomed Opt. 2007;12(6):062105.
Ryan TE, Southern WM, Reynolds MA, McCully KK. A cross-validation of near-infrared spectroscopy measurements of skeletal muscle oxidative capacity with phosphorus magnetic resonance spectroscopy. J Appl Physiol. 2013;115(12):1757–66.
Ryan TE, Brophy P, Te LC, Hickner RC, Neufer PD. Assessment of in vivo skeletal muscle mitochondrial respiratory capacity in humans by near-infrared spectroscopy: a comparison with in situ measurements. J Physiol. 2014;592(15):3231–41.
Grassi B, Quaresima V, Marconi C, Ferrari M, Cerretelli P. Blood lactate accumulation and muscle deoxygenation during incremental exercise. J Appl Physiol. 1999;87(1):348–55.
Kemp GJ, Ahmad RE, Nicolay K, Prompers JJ. Quantification of skeletal muscle mitochondrial function by 31 P magnetic resonance spectroscopy techniques: a quantitative review. Acta Physiol. 2015;213(1):107–44.
Haseler LJ, Hogan MC, Richardson RS. Skeletal muscle phosphocreatine recovery in exercise-trained humans is dependent on O2 availability. J Appl Physiol. 1999;86(6):2013–8.
Mader A, Heck H. A theory of the metabolic origin of “anaerobic thresold.” Int J Sports Med. 1986;7(1):45–65.
Brand MD. Regulation analysis of energy metabolism. J Exp Biol. 1997;200(Pt 2):193–202.
Mahler M. First-order kinetics of muscle oxygen consumption, and an equivalent proportionality between QO2 and phosphorylcreatine level. Implications for the control of respiration. J Gen Physiol. 1985;86(1):135–65.
Meyer RA. A linear model of muscle respiration explains monoexponential phosphocreatine changes. Am J Phys. 1988;254(4):548–53.
Holloszy JO, Coyle EF. Adaptations of skeletal muscle to endurance exercise and their metabolic consequences. J Appl Physiol. 1984;56(4):831–8.
Gollnick PD, Armstrong RB, Saltin B, Saubert CW, Sembrowich WL, Shepherd RE. Effect of training on enzyme activity and fiber composition of human skeletal muscle. J Appl Physiol. 1973;34(1):107–11.
Fryer S, Stoner L, Stone K, Giles D, Sveen J, Garrido I, et al. Forearm muscle oxidative capacity index predicts sport rock-climbing performance. Eur J Appl Physiol. 2016;116(8):1479–84.
Nakada M, Demura S, Yamaji S, Minami M, Kitabayashi T, Nagasawa Y. Relationships between force curves and muscle oxygenation kinetics during repeated handgrip. J Physiol Anthropol Appl Hum Sci. 2004;23(6):191–6.
Philippe M, Wegst D, Müller T, Raschner C, Burtscher M. Climbing-specific finger flexor performance and forearm muscle oxygenation in elite male and female sport climbers. Eur J Appl Physiol. 2012;112(8):2839–47.
van der Zwaard S, Jaspers RT, Blokland IJ, Achterberg C, Visser JM, den Uil AR, et al. Oxygenation Threshold Derived from Near-Infrared Spectroscopy: Reliability and Its Relationship with the First Ventilatory Threshold. Fisher G, editor. PLoS One. 2016;11(9).
Perrey S, Ferrari M. Muscle oximetry in sports science: a systematic review. Sport Med. 2018;48(3):597–616.
McCully KK, Iotti S, Kendrick K, Wang Z, Posner JD, Leigh JJ, et al. Simultaneous in vivo measurements of HbO2 saturation and PCr kinetics after exercise in normal humans. J Appl Physiol. 1994;77(1):5–10.
Campbell MD, Marcinek DJ. Evaluation of in vivo mitochondrial bioenergetics in skeletal muscle using NMR and optical methods. Biochim Biophys Acta - Mol Basis Dis. 2016;1862(4):716–24.
Liberati A, Altman DG, Tetzlaff J, Mulrow C, Gøtzsche PC, Ioannidis JPA, et al. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: explanation and elaboration. PLoS Med. 2009;6.
Hamaoka T, McCully KK. Review of early development of near-infrared spectroscopy and recent advancement of studies on muscle oxygenation and oxidative metabolism. J Physiol Sci. 2019;69(6):799–811.
Hamaoka T, Iwane H, Shimomitsu T, Katsumura T, Murase N, Nishio S, et al. Noninvasive measures of oxidative metabolism on working human muscles by near-infrared spectroscopy. J Appl Physiol. 1996;81(3):1410–7.
Yoshida T, Watari H, Tagawa K. Effects of active and passive recoveries on splitting of the inorganic phosphate peak determined by 31P-nuclear magnetic resonance spectroscopy. NMR Biomed. 1996;9(1):13–9.
Binzoni T, Hiltbrand E. Influence of repeated ischaemia/reperfusion cycles (ischaemic preconditioning) on human calf energy metabolism by simultaneous near infrared spectroscopy,31P-NMR and 23Na-NMR measurements. Adv Exp Med Biol. 1997;428:533–8.
Binzoni T, Quaresima V, Barattelli G, Hiltbrand E, Gürke L, Terrier F, et al. Energy metabolism and interstitial fluid displacement in human gastrocnemius during short ischemic cycles. J Appl Physiol. 1998;85(4):1244–51.
Boushel R, Pott F, Madsen P, Rådegran G, Nowak M, Quistorff B, et al. Muscle metabolism from near infrared spectroscopy during rhythmic handgrip in humans. Eur J Appl Physiol Occup Physiol. 1998;79(1):41–8.
Kutsuzawa T, Shioya S, Kurita D, Haida M, Yamabayashi H. Effects of age on muscle energy metabolism and oxygenation in the forearm muscles. Med Sci Sports Exerc. 2001;33(6):901–6.
Sako T, Hamaoka T, Higuchi H, Kurosawa Y, Katsumura T. Validity of NIR spectroscopy for quantitatively measuring muscle oxidative metabolic rate in exercise. J Appl Physiol. 2001;90(1):338–44.
Hamaoka T, Katsumura T, Murase N, Sako T, Higuchi H, Murakami M, et al. Muscle oxygen consumption at onset of exercise by near infrared spectroscopy in humans. In: Advances in experimental medicine and biology. Kluwer Academic/Plenum Publishers; 2003. p. 475–83.
Kime R, Hamaoka T, Sako T, Murakami M, Homma T, Katsumura T, et al. Delayed reoxygenation after maximal isometric handgrip exercise in high oxidative capacity muscle. Eur J Appl Physiol. 2003;89(1):34–41.
Kime R, Katsumura T, Hamaoka T, Osada T, Sako T, Murakami M, et al. In: Dunn JF, Swartz HM, editors., et al., Muscle Reoxygenation rate after isometric exercise at various intensities in relation to muscle oxidative capacity BT - oxygen transport to tissue XXIV. Boston, MA: Springer US; 2003. p. 497–507.
Nagasawa T, Hamaoka T, Sako T, Murakami M, Kime R, Homma T, et al. A practical indicator of muscle oxidative capacity determined by recovery of muscle O2 consumption using NIR spectroscopy. Eur J Sport Sci. 2003;3(2):1–10.
Forbes SC, Kowalchuk JM, Thompson RT, Marsh GD. Effects of hyperventilation on phosphocreatine kinetics and muscle deoxygenation during moderate-intensity plantar flexion exercise. J Appl Physiol. 2007;102(4):1565–73.
Kimura N, Hamaoka T, Kurosawa Y, Katsumura T. Contribution of intramuscular oxidative metabolism to Total ATP production during forearm isometric exercise at varying intensities. Tohoku J Exp Med. 2006;208(4):307–20.
Jones AM, Fulford J, Wilkerson DP. Influence of prior exercise on muscle [phosphorylcreatine] and deoxygenation kinetics during high-intensity exercise in men. Exp Physiol. 2008;93(4):468–78.
Forbes SC, Raymer GH, Kowalchuk JM, Thompson RT, Marsh GD. Effects of recovery time on phosphocreatine kinetics during repeated bouts of heavy-intensity exercise. Eur J Appl Physiol. 2008;103(6):665–75.
Zange J, Beisteiner M, Müller K, Shushakov V, Maassen N. Energy metabolism in intensively exercising calf muscle under a simulated orthostasis. Pflugers Arch Eur J Physiol. 2008;455(6):1153–63.
Layec G, Bringard A, Le Fur Y, Vilmen C, Micallef JP, Perrey S, et al. Effects of a prior high-intensity knee-extension exercise on muscle recruitment and energy cost: a combined local and global investigation in humans. Exp Physiol. 2009;94(6):704–19.
Willcocks RJ, Williams CA, Barker AR, Fulford J, Armstrong N. Age- and sex-related differences in muscle phosphocreatine and oxygenation kinetics during high-intensity exercise in adolescents and adults. NMR Biomed. 2010;23(6):569–77.
Layec G, Bringard A, Yashiro K, Le Fur Y, Vilmen C, Micallef JP, et al. The slow components of phosphocreatine and pulmonary oxygen uptake can be dissociated during heavy exercise according to training status. Exp Physiol. 2012;97(8):955–69.
Layec G, Haseler LJ, Trinity JD, Hart CR, Liu X, Le Fur Y, et al. Mitochondrial function and increased convective O-2 transport: implications for the assessment of mitochondrial respiration in vivo. J Appl Physiol. 2013;115(6):803–11.
Fulford J, Liepa A, Barker AR, Meakin J. The reliability of 31P-MRS and NIRS measurements of spinal muscle function. Int J Sports Med. 2014;35(13):1078–83.
Hart CR, Layec G, Trinity JD, Liu X, Kim S-E, Groot HJ, et al. Evidence of preserved oxidative capacity and oxygen delivery in the plantar flexor muscles with age. J Gerontol Ser A Biol Sci Med Sci. 2015;70(9):1067–76.
Layec G, Trinity JD, Hart CR, Kim SE, Groot HJ, Le Fur Y, et al. In vivo evidence of an age-related increase in ATP cost of contraction in the plantar flexor muscles. Clin Sci. 2014;126(8):581–92.
Bendahan D, Chatel B, Jue T. Comparative NMR and NIRS analysis of oxygen-dependent metabolism in exercising finger flexor muscles. Am J Physiol - Regul Integr Comp Physiol. 2017;313(6):R740–53.
Yanagisawa O, Sanomura M. Effects of low-load resistance exercise with blood flow restriction on high-energy phosphate metabolism and oxygenation level in skeletal muscle. Interv Med Appl Sci. 2017;9(2):67–75.
Heskamp L, Lebbink F, van Uden MJ, Maas MC, Claassen JAHR, Froeling M, et al. Post-exercise intramuscular O2 supply is tightly coupled with a higher proximal-to-distal ATP synthesis rate in human tibialis anterior. J Physiol. 2020;599(5):1533–50.
Zange J, Haller T, Müller K, Liphardt AM, Mester J. Energy metabolism in human calf muscle performing isometric plantar flexion superimposed by 20-Hz vibration. Eur J Appl Physiol. 2009;105(2):265–70.
Mann CJ. Observational research methods. Research design II: cohort, cross sectional, and case-control studies. Emerg Med J. 2003;20(1):54–60.
Chuang M-L, Ting H, Otsuka T, Sun X-G, Chiu FY-L, Hansen JE, et al. Muscle deoxygenation as related to work rate. Med Sci Sports Exerc. 2002;34(10):1614–23.
Connett RJ, Gayeski TE, Honig CR. Energy sources in fully aerobic rest-work transitions: a new role for glycolysis. Am J Physiol Circ Physiol. 1985;248(6):922–9.
Robergs RA, Ghiasvand F, Parker D. Biochemistry of exercise-induced metabolic acidosis. Am J Physiol Integr Comp Physiol. 2004;287(3):502–16.
Barbiroli B, Iotti S, Lodi R. Aspects of human bioenergetics as studied in vivo by magnetic resonance spectroscopy. Biochimie. 1998;80(10):847–53.
McMahon S, Jenkins D. Factors affecting the rate of phosphocreatine Resynthesis following intense exercise. Sport Med. 2002;32(12):761–84.
Chilibeck PD, Paterson DH, Petrella RJ, Cunningham DA. The influence of age and cardiorespiratory fitness on kinetics of oxygen uptake. Can J Appl Physiol. 1996;21(3):185–96.
Ren J, Lakoski S, Haller RG, Sherry AD, Malloy CR. Dynamic monitoring of carnitine and acetylcarnitine in the trimethylamine signal after exercise in human skeletal muscle by 7T 1H-MRS. Magn Reson Med. 2013;69(1):7–17.
Damon BM, Buck AKW, Ding Z. Diffusion-tensor MRI based skeletal muscle Fiber tracking. Imaging Med. 2011;3(6):675–87.
DeBrosse C, Nanga RPR, Wilson N, D’Aquilla K, Elliott M, Hariharan H, et al. Muscle oxidative phosphorylation quantitation using creatine chemical exchange saturation transfer (CrCEST) MRI in mitochondrial disorders. JCI insight. 2016;1(18):e88207.
Acknowledgements
Not applicable.
Funding
Open Access funding enabled and organized by Projekt DEAL. No funding was obtained to support this study.
Author information
Authors and Affiliations
Contributions
The article was conceptualized by KM, AF, and RJ. All authors, namely KM, AF, RJ, and KM, were involved in drafting and revising the manuscript, and have given their approval for the final version to be published. Each author takes responsibility for all aspects of the work and agrees to be accountable for it.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
Kevin Maliszewski, and Ross Julian declare no conflicts of interest. Andri Feldmann is a minority shareholder in the company Fortiori Designs LLC. Kevin K. McCully is the president of InfraredRx, Inc.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Maliszewski, K., Feldmann, A., McCully, K.K. et al. A systematic review of the relationship between muscle oxygen dynamics and energy rich phosphates. Can NIRS help?. BMC Sports Sci Med Rehabil 16, 25 (2024). https://doi.org/10.1186/s13102-024-00809-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13102-024-00809-5