
Abstract
Background
Fleas, considered to be the main transmission vectors of Bartonella, are highly prevalent and show great diversity. To date, no investigations have focused on Bartonella vectors in Southeast China. The aim of this study was to investigate the epidemiological and molecular characteristics of Bartonella in fleas in Southeast China.
Methods
From 2016 to 2022, flea samples (n = 1119) were collected from 863 rodent individuals in seven inland and coastal cities in Southeast China. Flea species, region, gender, host species and habitat were recorded. The DNA samples from each individual flea were screened by real-time PCR for the Bartonella ssrA gene. All positive samples were confirmed by PCR based on the presence of the gltA gene and sequenced. The factors associated with Bartonella infection were analyzed by the Chi-square test and Fisher's exact test. ANOVA and the t-test were used to compare Bartonella DNA load.
Results
Bartonella DNA was detected in 26.2% (293/1119) of the flea samples, including in 27.1% (284/1047) of Xenopsylla cheopis samples, 13.2% (5/38) of Monopsyllus anisus samples, 8.3% (2/24) of Leptopsylla segnis samples and 20.0% (2/10) of other fleas (Nosopsyllus nicanus, Ctenocephalides felis, Stivalius klossi bispiniformis and Neopsylla dispar fukienensis). There was a significant difference in the prevalence of Bartonella among flea species, sex, hosts, regions and habitats. Five species of Bartonella fleas were identified based on sequencing and phylogenetic analyses targeting the gltA gene: B. tribocorum, B. queenslandensis, B. elizabethae, B. rochalimae and B. coopersplainsensis.
Conclusions
There is a high prevalence and diversity of Bartonella infection in the seven species of fleas collected in Southeast China. The detection of zoonotic Bartonella species in this study, including B. tribocorum, B. elizabethae and B. rochalimae, raises public health concerns.
Graphical Abstract

Similar content being viewed by others
Background
Bartonella is a group of Gram-negative, fastidious, facultative, intracellular parasitic aerobic bacilli belonging to the class Proteobacteria, order Rhizobacteria, family Bartonellaceae and genus Bartonella that parasitize the erythrocytes and vascular endothelial cells of hosts and infect humans or other mammalian hosts through blood-sucking arthropods [1]. At least 40 species of Bartonella and its subspecies are currently recognized, of which at least 15 are human pathogens [2]. The clinical manifestations of Bartonella infection in humans range from mild to life-threatening and can be acute or chronic. Known symptoms of Bartonella in humans include endocarditis, myocarditis, fever and neurological disorders, intraocular retinitis, meningitis, splenomegaly and lymph node enlargement [3,4,5,6,7]. This constellation of nonspecific and variable symptoms make Bartonella infection difficult to diagnose clinically (Additional file 1: Table S1).
Rodents are natural hosts for approximately 20 species of Bartonella [8], and Bartonella has been detected in almost 100 rodent species worldwide. Importantly, a number of human pathogenic Bartonella species, such as B. elizabethae, B. grahamii, B. vinsonii subsp. arupensis and B. washoensis, are carried by rodents [9].
Bartonella is mainly transmitted horizontally [10], with arthropods acquiring Bartonella when blood feeding on an infected host with the subsequent transfer Bartonella to another host. Sand flies, body lice and cat fleas are involved in the transmission of B. bacilliformis, B. quintana and B. henselae, respectively [1]. Fleas are considered to be the primary vectors of Bartonella transmission among rodents, and a variety of fleas have been shown to be infected by zoonotic Bartonella species such as B. henselae, B. clarridgeiae, B. quintana, B. grahamii and B. elizabethae [11,12,13,14,15]. Fleas have been shown to play an important role in the transmission and acquisition of Bartonella species in rodents, and Bartonella DNA has been detected in fleas on rodents [16], providing evidence that fleas are vectors for the transmission of Bartonella among rodents.
Currently, 28 species of rodents belonging to seven families and 14 genera and 27 species of fleas belonging to six families and 18 genera have been identified in Fujian Province (China) [17, 18]. Previous systematic investigations conducted on 10 species of Bartonella host rodents harboring Bartonella in Southeast China identified Bartonella species in rodents, including B. tribocorum, B. grahamii, B. rattimassiliensis, B. queenslandensis, B. elizabethae, B. phoceensis, B. coopersplainsensis, B. japonica and B. rochalimae [19]. To date, however, no investigations have been conducted on Bartonella vectors. In the present study, we analyzed the epidemiological and molecular characteristics of Bartonella in fleas in Southeast China by investigating Bartonella infection in several areas of this region. Our aim was to assess the public health risk of the host-vector relationship between rodents and fleas on the transmission of Bartonella in the natural habitats of Southeast China.
Methods
Ethical aspects
This study was approved by the Ethics Committee of Fujian Center for Disease Control and Prevention (No: FJCDCNT1811-2015). All rodents were treated in accordance with the Guidelines of Regulations for the Administration of Laboratory Animals of the People's Republic of China.
Sample collection and identification
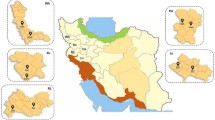
Rodents were captured in seven inland and coastal cities in Southeast China, namely Zhangzhou City, Quanzhou City, Sanming City, Longyan City, Ningde City, Fuzhou City and Putian City, and one to three fleas were collected from the body surface of each captured animal. Rodents were captured in live-capture traps baited with corn. Live traps were placed every night at each surveillance point for three consecutive nights at locations where rodent activities were detected, and retrieved the following morning.
Following capture, rodents were anesthetized with ether, and fleas were collected from the body surface of the rodents and from the cloth bags in which the rodents were held. Chinese monographs were used to identify the species of trapped rodents according to body shape, tail, coat color and other morphological characteristics [20]. The fleas were identified to species under the stereomicroscope by observing the distribution of setae and spines and the morphology of important structures such as eyes and genitalia by stereomicroscope, as well as by literature references [21]. We then individually recorded flea species, region, sex, host species and habitat. The fleas were morphologically classified and counted for registration and were stored in 75% alcohol at -20 °C until examination. After fleas had been collected, all rodents were used for surveillance in other programs.
Molecular analyses
Following published guidelines [22], before DNA extraction, each individual flea was immersed in 75% ethanol for 5–10 min, followed by two to three immersions in phosphate-buffered saline (PBS). The flea samples were then immersed in the lysate for 2 h and ground to a powder. DNA was extracted using a bacterial genomic DNA extraction kit (Tianlong Science & Technology, Xi'an, China) according to the manufacturer's instructions and stored at − 20 °C. DNA was extracted in order to identify Bartonella species using a real-time PCR (qPCR) assay targeting a transfer-mRNA gene (ssrA) [23]. The primers ssrA-F (5′-GCTATGGTAATAAATGGACAATGAAATAA-3′) and ssrA-R (5′-GCTTCTGTTGCTAGGTG-3′) and the FAM-labeled probe (FAM-ACCCCGCTTAAA CCTGCG-BHQ1) were used to amplify a 301-bp fragment of the ssrA gene. qPCR amplification was performed in a 20-μl reaction mixture containing 10 μl of Premix Ex Taq (Probe qPCR; Takara, Shiga, Japan), 0.4 μl each of 10 μM forward and reverse primers, 0.2 μl of 10 μM probe, 3 μl of DNA template and double-distilled water. The qPCR conditions were: 95 °C for 5 min; then 50 cycles of 95 °C for 15 s and 60 °C for 45 s. Samples with Ct (cycle threshold) values ≤ 35 were considered to be positive for Bartonella DNA. Positive samples were then subjected to conventional PCR to amplify the 379-bp gltA gene fragment [24] using the primers BhCS781.p (5′-GGGGACCAGCTCATGGT GG-3′) and BhCS1137.n (5′-AATGCAAAAAGAACAATAAACA-3′)[24]. The conventional PCR analysis was carried out in a total reaction volume of 25 μl containing 3 μl of template DNA, 1 μl each of 10 μM forward and reverse primers, 12.5 µl Premix Taq™ (Premix Taq Version 2.0 plus dye; Takara) and 7.5 µl double-distilled water. The amplification procedure was: 95 °C for 5 min; followed by 35 cycles of 95 °C for 30 s, 58 °C for 30 s and 72 °C for 30 s; with a final cycle at 72 °C for 5 min. The PCR products were separated by electrophoresis in a 1.5% agarose gel. During all PCR amplifications, distilled water was used as the negative control and positive DNA samples obtained from previous rodent surveys [19] were used as positive controls.
Bartonella ssrA sequences were sent to Sangon Biotech Company (Sangon Biotech, Shanghai, China) for gene synthesis to construct plasmid DNA. In addition, the Bartonella DNA load was calculated for each positive flea sample using a standard curve generated from a tenfold dilution (2log10-6log10 copies/μl) of plasmid DNA encoding a 300-bp B. henselae ssrA gene fragment.
DNA sequencing and phylogenetic analysis
Positive amplification products were subsequently sent to Sangon Biotech Company (Sangon Biotech) for sequencing.
The gltA sequences were compared with the sequences of the type strains of the validated Bartonella species in the GenBank database using NCBI BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi). After alignment of the gltA sequences by ClustalW, phylogenetic trees were created using the neighbor-joining method in MEGA 11.0 software. The best-fit nucleotide substitution model for the phylogenetic analysis was estimated based on the Bayesian information criterion (BIC) calculated using MEGA 11 software [25].
Statistical analysis
The Chi-square test (χ2) and Fisher's exact test were used to evaluate the correlations between flea species, region, gender, host species, habitat and Bartonella infection. P < 0.05 was considered to indicate statistical significance. Analysis of variance (ANOVA) and the t-test were used to compare Bartonella loads.
All statistical analyses were performed using SPSS version 23.0 statistical software (SPSS IBM Corp, Armonk, NY, USA).
Results
Flea collection and morphological identification
A total of 1119 fleas were collected in seven cities during this survey, and seven species of fleas were identified (Table 1): Xenopsylla cheopis (n = 1047), Monopsyllus anisus (n = 38), Leptopsylla segnis (n = 24 ), Ctenocephalides felis (n = 6 ), Nosopsyllus nicanus (n = 1 ), Neopsylla dispar fukienensis (n = 1) and Stivalius klossi bispiniformis (n = 2). Among these, X. cheopis was the dominant flea species collected from the rats captured Southeast China, accounting for 93.6% (1047/1119) of the total fleas. A total of 308 fleas were from Zhangzhou city, 191 fleas were from Quanzhou city, 127 fleas were from Sanming city, 60 fleas were from Longyan city, 127 fleas were from Ningde city, 34 fleas were from Fuzhou city and 272 fleas were from Putian city.
Detection and quantification of Bartonella spp. DNA
Bartonella-ssrA DNA was detected in 26.2% (293/1119, 95% confidence interval [CI] 23.6–28.8%) of the tested fleas from Southeast China (Table 2). Among the fleas found, 27.1% (284/1047) of the X. cheopis, 13.2% (5/38) of the M. anisus, 8.3% (2/24) of the L. segnis and 20.0% (2/10) of the ‘other’ fleas (N. nicanus, C. felis, S. klossi bispiniformis and N. fukienensis) were positive for Bartonella, with X. cheopis having the highest prevalence of infection and L. segnis the lowest. There was a significant difference in the prevalence of Bartonella among the different flea species (χ2 = 9.48, df = 3, P = 0.024). The infection rate of female fleas (28.9%, 217/750) was greater than that of male fleas (20.6%, 76/369), and there was a significant difference in the prevalence of infection between the sex (χ2 = 8.89, df = 1, P = 0.003).
In this study, seven rodent species, namely Rattus norvegicus, Rattus flavipectus, Rattus losea, Niviventer coninga, Bandicota indica, Mus musculus, and Niviventer fulvescens, and one mammal species, Suncus murinus, were captured. When fleas from N. coninga, B. indica and N. fulvescens were not taken into account, the prevalence of fleas ranged from 16.7% to 29.8% (note: host species was not recorded in four fleas; Table 2). There was a significant difference in the prevalence among different hosts (χ2 = 18.948, df = 7, P = 0.008). Two or more fleas were captured from 245 hosts, with 14.5% (33/245) of these infected with Bartonella; 30.6% (75/245) were infected by only one flea and 55.9% (137/245) were not infected.
There was a significant difference in the prevalence of Bartonella in the different regions (χ2 = 75.23, df = 6, P < 0.001), with the highest incidence (33.3%, 20/60) occurring in Longyan City and the lowest prevalence (8.8%, 3/34) occurring in Fuzhou City (Table 2). In terms of geographical location of the seven cities investigated, Ningde City, Fuzhou City, Putian City, Zhangzhou City and Quanzhou City are located in the coastal area, and Sanming City and Longyan City are located in the inland area. The prevalence of Bartonella in the coastal cities was 26.4% (246/932) and that in the inland cities was 25.1% (47/187); the difference in prevalence among these two different geographic locations was not statistically significant (χ2 = 0.13, df = 1, P > 0.05). With the exception of four fleas from unrecorded habitats, 22.1% (202/915) of the fleas collected in wildernesses/farmlands were infected with Bartonella, and 45% of fleas (90/200) collected in residential areas were infected (Table 2). Fleas from residential areas had a significantly greater prevalence of Bartonella infection than did those collected in fields/farmland (χ2 = 44.62, df = 1, P < 0.001).
The prevalence of Bartonella in flea samples showed seasonal variation (Fig. 1), increasing from 16.4% in April to 26.7% in June, then decreasing to 16.8% in July, followed by an increase to a peak infection of 39.4% from August to October. The difference in Bartonella prevalence was significantly different between the different months (χ2 = 32.08, df = 6, P < 0.001).

Monthly prevalence of Bartonella in fleas in southeast China. Filled circles represent the prevalence of Bartonella
A standard curve was established using plasmid DNA from the B. henselae ssrA gene fragment with r2 = 0.996, a slope of − 3.62, and a y-intercept of 40.42 (Fig. 2). The Bartonella loads of the positive fleas ranged from 1.35 to 8.29 log10 copies/μl (mean ± standard deviation [SD] 2.78 ± 1.14). Flea Bartonella loads were statistically significantly different among the different regions (F = 2.178, P = 0.045), with the highest flea bacterial loads occurring in Longyan city (mean ± SD, 3.19 ± 1.13) and the lowest occurring in Putian city (mean ± SD, 2.34 ± 1.14) (Fig. 3c). The fecal bacterial load in fleas caught in wildernesses (mean ± SD, 2.99 ± 1.20) was significantly higher than that in fleas caught in residential areas (mean ± SD, 2.70 ± 1.11) (t = − 2.010, P = 0.045) (Fig. 3e). Changes in flea bacterial loads over time showed a trend similar to that of prevalence and were significantly different (F = 3.148, P = 0.005) (Fig. 3d). Differences in flea Bartonella loads among flea species (F = 1.108, P = 0.346) (Fig. 3a), sex (t = 0.553, P = 0.581) (Fig. 3b) and host species (F = 1.977, P = 0.098) (Fig. 3f) were not statistically significant.

Standard curve based on Bartonella henselae ssrA gene fragment plasmid DNA. Tenfold serial dilutions of the plasmid vector DNA were performed (2log10-6log10 copies/μl), and real-time PCR analyses were repeated three times for each dilution concentration. The slope and intercept of the regression curve are shown. Ct Cycle threshold; lg, log

Boxplot of Bartonella loads in positive samples from fleas of different species (a), sex (b), locations (c), time points (d), habitats (e) and hosts (f). Boxes represent IQRs, and vertical lines represent the distribution of maximum and minimum values. The values on the y-axis are expressed as log DNA copies/μl
Sequence comparison and phylogenetic analysis
In total, 114 gltA sequences were analyzed via BLAST, and the phylogenetic analysis included sequences of 18 Bartonella genotypes, six Bartonella strains previously isolated from rodents in Southeast China and 26 representative flea samples from the present study. Brucella was also included as an outgroup (Fig. 4). The phylogenetic tree showed that the Bartonella-positive samples could be divided into five different branches. A total of 35.1% (40/114) of the gltA sequences belonged to B. tribocorum, which is the dominant genotype in Southeast China and is in the same branch as KT324580 in Thailand and MW771088 in Fujian, with 100% similarity. Seven sequences were detected as B. queenslandensis, with 95.5%-100% similarity to KT324558 from Thailand and MW771064 from Fujian. Twenty-three sequences of B. elizabethae were 99.1–100% homologous to JX158352 and GU056192 from Thailand and Taiwan, as well as to MW771077 and MW771078 from Fujian. Twenty-seven B. rochalimae sequences showed 100% similarity with those of MG027988 from the USA and MW771100 from Fujian. Nine B. coopersplainsensis sequences showed 94.3–98.5% similarity with HQ444160 from Australia and MW771106 from Fujian. Although the previously investigated rodents were not the hosts of the present flea samples, their Bartonella spp. were analyzed against the present samples, and the similarity reached 96.1–100%. Interestingly, of two or more fleas from the same host, four pairs were infected with the same Bartonella species: B. tribocorum, B. rochalimae and B. elizabethae.

Phylogenetic tree based on the gltA gene of Bartonella. The phylogenetic tree was constructed using the neighbor-joining method based on the maximum composite likelihood model, and bootstrap values were calculated with 1000 replicates
The differences in flea Bartonella loads among the different regions were significantly different (P < 0.001). Bartonella coopersplainsensis-infected fleas were found to have higher bacterial loads (mean ± SD, 3.92 ± 0.57 log10 copies/μl) than other species. Among all species, B. elizabethae had the lowest load (mean ± SD, 2.13 ± 0.566 log10 copies/μl).
Discussion
Fleas are recognized as key players in the transmission of Bartonella, as they are able to carry a high diversity of Bartonella species and transmit them efficiently among rodents [26]. This efficient transmission of Bartonella is regarded as an important factor in maintaining its high prevalence in the natural environment. In China, there are relatively few investigations on ectoparasite infections caused by Bartonella. Li DM [27, 28], who detected Bartonella from the bacteria Chlamydophila felis and Leptopsylla segnis, isolated Bartonella strains from fleas and ticks. Bartonella infection in fleas has also been found in Qinghai Province, the Qinghai-Tibet Plateau and the China-Kazakhstan Border [29,30,31]. The present study emphasized the prevalent distribution of Bartonella in fleas and the related genotypes in Southeast China, with the data showing that there was a high prevalence of Bartonella in fleas in Southeast China and that multiple Bartonella genotypes could be identified.
The reported prevalence of flea Bartonella DNA detection in various countries varies, ranging from 2.2% to 40% in Egypt, the USA, France, Chile and Japan [9, 14, 32,33,34]. The overall Bartonella infection rate in fleas in the present study was 26.2% (293/1119) according to the qPCR analyses, which is higher than that reported our previous study of Bartonella infection in rodents in Southeast China (14.6–14.9%) [19, 35]. One factor for fleas possessing such a high infection rate may be their frequent feeding and ability to move from one host to another [36]. Our results also showed that B. tribocorum was the predominant genotype of Bartonella fleas in Southeast China; this species can cause causing acute fever and bacteremia in humans. Therefore, it is necessary to evaluate the epidemiological characteristics of Bartonella in fleas.
The transmission and acquisition of Bartonella are mediated by the host specificity of fleas, flea exchange between rodents and flea abundance [37]. According to our survey, X. cheopis was the dominant flea species in Southeast China and also the most prevalent flea. Our observations are similar to the results of a survey of rodents in the USA [38], which showed that the highest prevalence usually occurred among the most common species in rodent communities. These results indicate that the increase in the prevalence of Bartonella in fleas may also be related to the dominance of flea species in the population. Moreover, the density of the hosts may also increase flea transmission and infection among the hosts, as we mainly conducted surveys in villages and surrounding farmland where captured rodents were dominated by domestic rats, such as R. norvebicus, R. flavipectus and S. murinus, which were also accompanied by a high prevalence of parasitic flea infestations on their body surfaces (20.8–29.8%). Additionally, we found that residential areas have higher infection rates than wildernesses/farmlands, which undoubtedly increases the likelihood of flea contact with humans and disease transmission. In addition, animal sex has not been identified as a risk factor for Bartonella infection in rodents from Taiwan and France [39, 40]. However, in our study, we found that the incidence of Bartonella infection was significantly greater in females than in males, and this difference may be related to the parasitism and blood-sucking habits of fleas, with females sucking a greater amount of blood more frequently and for a longer period than males.
Several previous studies have shown that the prevalence of Bartonella in rodents and their ectoparasitic fleas is influenced by seasonality, peaking from the summer to fall [41,42,43]. In the present study, we found that the prevalence of Bartonella in fleas was markedly seasonal, with a clear upward trend in the prevalence of this genus from July to October. Late summer and early fall are not only periods of prevalence of Bartonella transmission but also periods of peak vector activity [44], making this period a risky time for Bartonella transmission to other species, including humans.
Phylogenetic analysis of the Bartonella gltA gene revealed five Bartonella genotypes, namely B. tribocorum, B. queenslandensis, B. elizabethae, B. rochalimae and B. coopersplainsensis, indicating the high diversity of Bartonella in the fleas of Southeast China. Bartonella tribocorum, B. elizabethae and B. rochalimae were the major genotypes identified in this survey, and all of them are pathogenic to humans, causing endocarditis, myocarditis, fever and neurological diseases. The high diversity of Bartonella genotypes may be a result of frequent host changes in fleas and their high efficiency in transmitting Bartonella. We compared the sequences of Bartonella species previously isolated from rodents in Southeast China with those isolated in the present study; the homology was 96.2%-100%, indicating the high adaptation of Bartonella species to rodents and fleas. In addition, Bowen et al. [11] reported that 75% (21/28) of bank voles housed with wild-caught fleas for 4 weeks developed Bartonella infections, and the present study also revealed multiple groups of fleas from the same host infected with the same Bartonella genotype at the same time, suggesting that fleas may play a potential role as vectors for the transmission of Bartonella among rodents. However, it is worth noting that the PCR detection of Bartonella spp. in fleas does not necessarily mean that they actively infest the host. Consequently, the mechanism of Bartonella spp. transmission between fleas and rodents still needs to be investigated more thoroughly.
Conclusions
The present study describes the prevalence and genetic characteristics of Bartonella species in fleas in southeast China. The results showed that there was a high prevalence and diversity of Bartonella in fleas. We identified five Bartonella genotypes in fleas, of which the zoonotic B. tribocorum, B. elizabethae, and B. rochalimae will pose a threat to human health in southeast China. However, the vector capacity of fleas was not determined in this study. In future studies, the host-vector relationship of Bartonella can be further investigated via animal experiments.
Availability of data and materials
All data generated or analysed during this study are included in this published article [and its supplementary information files].
Abbreviations
- qPCR:
-
Quantitative real-time PCR
References
Jin X, Gou Y, Xin Y, Li J, Sun J, Li T, et al. Advancements in understanding the molecular and immune mechanisms of Bartonella pathogenicity. Front Microbiol. 2023;14:1196700.
Mardosaitė-Busaitienė D, Radzijevskaja J, Balčiauskas L, Bratchikov M, Jurgelevičius V, Paulauskas A. Prevalence and diversity of Bartonella species in small rodents from coastal and continental areas. Sci Rep. 2019;9:12349.
Amin O, Rostad CA, Gonzalez M, Rostad BS, Caltharp S, Quincer E, et al. Cat scratch disease: 9 years of experience at a pediatric center. Open Forum Infect Dis. 2022;9:ofac426.
Maria HKS, Gazzoli E, Drummond MR, Almeida AR, Santos LSD, Pereira RM, et al. Two-year history of lymphadenopathy and fever caused by Bartonella henselae in a child. Rev Inst Med Trop Sao Paulo. 2022;64:e15.
Nawrocki CC, Max RJ, Marzec NS, Nelson CA. Atypical manifestations of cat-scratch disease, United States, 2005–2014. Emerg Infect Dis. 2020;26:1438–46.
Niederer RL, Al-Ani HH. Bartonella Neuroretinitis. N Engl J Med. 2021;384:952.
Sato S, Shapira L, Tasher D, Maruyama S, Giladi M. Molecular epidemiology of Bartonella quintana endocarditis in patients from Israel and Eastern Africa. BMC Infect Dis. 2023;23:142.
Krügel M, Król N, Kempf VAJ, Pfeffer M, Obiegala A. Emerging rodent-associated Bartonella: a threat for human health? Parasit Vectors. 2022;15:113.
Müller A, Gutiérrez R, Seguel M, Monti G, Otth C, Bittencourt P, et al. Molecular survey of Bartonella spp. in rodents and fleas from Chile. Acta Trop. 2020;212:105672.
Tsai YL, Chang CC, Chuang ST, Chomel BB. Bartonella species and their ectoparasites: selective host adaptation or strain selection between the vector and the mammalian host? Comp Immunol Microbiol Infect Dis. 2011;34:299–314.
Bown KJ, Bennet M, Begon M. Flea-borne Bartonella grahamii and Bartonella taylorii in bank voles. Emerg Infect Dis. 2004;10:684–7.
Chomel BB, Kasten RW, Floyd-Hawkins K, Chi B, Yamamoto K, Roberts-Wilson J, et al. Experimental transmission of Bartonella henselae by the cat flea. J Clin Microbiol. 1996;34:1952–6.
De Sousa R, Edouard-Fournier P, Santos-Silva M, Amaro F, Bacellar F, Raoult D. Molecular detection of Rickettsia felis, Rickettsia typhi and two genotypes closely related to Bartonella elizabethae. Am J Trop Med Hyg. 2006;75:727–31.
Rolain JM, Franc M, Davoust B, Raoult D. Molecular detection of Bartonella quintana, B. koehlerae, B. henselae, B. clarridgeiae, Rickettsia felis, and Wolbachia pipientis in cat fleas, France. Emerg Infect Dis. 2003;9:338–42.
Sasaki T, Poudel SK, Isawa H, Hayashi T, Seki N, Tomita T, et al. First molecular evidence of Bartonella quintana in Pediculus humanus capitis (Phthiraptera: Pediculidae), collected from Nepalese children. J Med Entomol. 2006;43:110–2.
Gutiérrez R, Cohen C, Flatau R, Marcos-Hadad E, Garrido M, Halle S, et al. Untangling the knots: co-infection and diversity of Bartonella from wild gerbils and their associated fleas. Mol Ecol. 2018;27:4787–807.
Zhan S. Species and regional distribution of rodents in Fujian Province. Chin J Vector Biol Control. 2002;04:317.
Zhou S, Lin D, Chen L, Li S, Wang L, Deng Y. Fleas floristic distribution in Fujian Province. Chin J Control Endemic Dis. 2013;28:172–6.
Liu H, Han T, Liu W, Xu G, Zheng K, Xiao F. Epidemiological characteristics and genetic diversity of Bartonella species in rodents from southeastern China. Zoonoses Public Health. 2022;69:224–34.
Huang WJ. Rodents of China. Shanghai: Fudan University Press; 1995.
Abbot P, Aviles AE, Eller L, Durden LA. Mixed infections, cryptic diversity, and vector-borne pathogens. Evidence from Polygenis fleas and Bartonella species. Appl Environ Microbiol. 2007;73:6045–52.
Gutiérrez R, Vayssier-Taussat M, Buffet JP, Harrus S. Guidelines for the isolation, molecular detection, and characterization of Bartonella species. Vector Borne Zoonotic Dis. 2017;17:42–50.
Diaz MH, Bai Y, Malania L, Winchell JM, Kosoy MY. Development of a novel genus-specific real-time PCR assay for detection and differentiation of Bartonella species and genotypes. J Clin Microbiol. 2012;50:1645–9.
Norman AF, Regnery R, Jameson P, Greene C, Krause DC. Differentiation of Bartonella-like isolates at the species level by PCR-restriction fragment length polymorphism in the citrate synthase gene. J Clin Microbiol. 1995;33:1797–803.
Tamura K, Stecher G, Kumar S. MEGA11. Molecular evolutionary genetics analysis version 11. Mol Biol Evolut. 2021;38:3022–7.
Brinkerhoff RJ, Kabeya H, Inoue K, Bai Y, Maruyama S. Detection of multiple Bartonella species in digestive and reproductive tissues of fleas collected from sympatric mammals. Isme J. 2010;4:955–8.
Li D, Liu Q, Yu D, Dong X. Molecular evidence of Bartonella species from fleas in Yunnan. Chin J Vector Biol Control. 2005;01:5–8.
Li D, Liu Q, Yu D, Zhang L, Dong X. Isolation and molecular identification of Bartonella from fleas and ticks. Chin J Zoonoses. 2005;21::1052-1058+1074.
Zheng Y, Wu P, Wei R, Wang Y, Zhao Z, Li Y, et al. Survey on the status of Bartonella and Yersinia Pestis infection in part of Qinghai province. Modern Prevent Med. 2014;41:1112–4+1117.
Dong L, Li Y, Yang C, Gong J, Zhu W, Huang Y, et al. Species-level microbiota of ticks and fleas from Marmota himalayana in the Qinghai-Tibet Plateau. Front Microbiol. 2023;14:1188155.
Yin X, Zhao S, Yan B, Tian Y, Ba T, Zhang J, et al. Bartonella rochalimae, B. grahamii, B. elizabethae, and Wolbachia spp. in fleas from wild rodents near the China-Kazakhstan Border. Korean J Parasitol. 2019;57:553–9.
Kabeya H, Inoue K, Izumi Y, Morita T, Imai S, Maruyama S. Bartonella species in wild rodents and fleas from them in Japan. J Vet Med Sci. 2011;73:1561–7.
Loftis AD, Reeves WK, Szumlas DE, Abbassy MM, Helmy IM, Moriarity JR, et al. Surveillance of Egyptian fleas for agents of public health significance: Anaplasma, Bartonella, Coxiella, Ehrlichia, Rickettsia, and Yersinia pestis. Am J Trop Med Hyg. 2006;75:41–8.
Stevenson HL, Bai Y, Kosoy MY, Montenieri JA, Lowell JL, Chu MC, et al. Detection of novel Bartonella strains and Yersinia pestis in prairie dogs and their fleas (Siphonaptera: Ceratophyllidae and Pulicidae) using multiplex polymerase chain reaction. J Med Entomol. 2003;40:329–37.
Xiao F, Lin D, Zhou S, Xu G, Deng Y. Investigation and sequence analysis on Bartonella infection in rodents in Southeast China. China Chin J Zoonoses. 2017;33:607–12.
Kosoy M, Mandel E, Green D, Marston E, Jones D, Childs J. Prospective studies of Bartonella of rodents. Part II. Diverse infections in a single rodent community. Vector Borne Zoonotic Dis. 2004;4:296–305.
Gutiérrez R, Krasnov B, Morick D, Gottlieb Y, Khokhlova IS, Harrus S. Bartonella infection in rodents and their flea ectoparasites: an overview. Vector Borne Zoonotic Dis. 2015;15:27–39.
Kosoy MY, Regnery RL, Tzianabos T, Marston EL, Jones DC, Green D, et al. Distribution, diversity, and host specificity of Bartonella in rodents from the Southeastern United States. Am J Trop Med Hyg. 1997;57:578–88.
Gundi VA, Davoust B, Khamis A, Boni M, Raoult D, La Scola B. Isolation of Bartonella rattimassiliensis sp. nov. and Bartonella phoceensis sp. Nov. from European Rattus norvegicus. J Clin Microbiol. 2004;42:3816–8.
Tsai YL, Chuang ST, Chang CC, Kass PH, Chomel BB. Bartonella species in small mammals and their ectoparasites in Taiwan. Am J Trop Med Hyg. 2010;83:917–23.
Bai Y, Kosoy MY, Ray C, Brinkerhoff RJ, Collinge SK. Temporal and spatial patterns of Bartonella infection in black-tailed prairie dogs (Cynomys ludovicianus). Microb Ecol. 2008;56:373–82.
Cevidanes A, Altet L, Chirife AD, Proboste T, Millán J. Drivers of Bartonella infection in micromammals and their fleas in a Mediterranean peri-urban area. Vet Microbiol. 2017;203:181–8.
Paziewska A, Harris PD, Zwolińska L, Bajer A, Siński E. Differences in the ecology of Bartonella infections of Apodemus flavicollis and Myodes glareolus in a boreal forest. Parasitology. 2012;139:881–93.
Krasnov BR, Morand S, Hawlena H, Khokhlova IS, Shenbrot GI. Sex-biased parasitism, seasonality and sexual size dimorphism in desert rodents. Oecologia. 2005;146:209–17.
Acknowledgements
Not applicable.
Funding
This work was supported by the National Science and Technology Major Project, Award Numbers, Grant/Award Number: 2017ZX10103008, and the Fujian Provincial Science and Technology Innovation Platform Construction Project, Award Numbers, Grant/Award Number: 2022CXA034.
Author information
Authors and Affiliations
Contributions
ZK and XF designed the study. ZS, ZZ, HT and LW prepared and provided experimental materials. NY and ZS conducted the experiments and data analysis. XF supervised the study. NY wrote the manuscript draft. All the authors reviewed and approved the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
This study has been approved by the Ethics Committee of Fujian Center for Disease Control and Prevention (No: FJCDCNT1811-2015). All rodents were treated in accordance with the Guidelines of Regulations for the Administration of Laboratory Animals of the People's Republic of China.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Table S1.
Rodent species and their ectoparasitic flea species.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zhou, S., Nian, Y., Zeng, Z. et al. Epidemiological survey and genetic diversity of Bartonella in fleas collected from rodents in Fujian Province, Southeast China. Parasites Vectors 17, 264 (2024). https://doi.org/10.1186/s13071-024-06305-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-024-06305-6