
Abstract
Background
Bluetongue is a non-contagious viral disease that affects both domestic and wild ruminants. It is transmitted primarily by small hematophagous Diptera belonging to the genus Culicoides (Diptera: Ceratopogonidae). The current study represents the first molecular investigation into the potential role of Culicoides imicola, Culicoides paolae, Culicoides newsteadi, Culicoides spp., and Culicoides circumscriptus as bluetongue virus (BTV) vectors in Morocco. Additionally, the study aimed to evaluate the vectorial activity of midges during the survey seasons.
Methods
Parous females of these species were captured from several regions of Morocco (6 out of 12) from 2018 to 2021 using Onderstepoort Veterinary Institute (OVI) traps. A total of 2003 parous female specimens were grouped into 55 batches. The midge body of each batch was dissected into three regions (head, thorax, and abdomen), and these regions were analyzed separately using reverse transcription quantitative polymerase chain reaction (RT-qPCR).
Results
BTV RNA was detected in 45 out of the 55 batches tested, indicating a positivity rate of 81.8%. The RT-qPCR-positive pools of the studied Culicoides species exhibited high levels of BTV positivity in each body part (head, thorax, and abdomen), confirming the successful replication of the virus within midge bodies. The BTV circulation was substantial across all three survey seasons (spring, summer, and autumn). High infection rates, calculated using the minimum infection rate (MIR) and maximum likelihood estimation (MLE), were observed during the collection seasons, particularly in autumn and spring, and for all investigated Culicoides species, most notably for C. imicola and C. newsteadi. These increased infection rates underscore the significant risk of Culicoides transmitting the BTV in Morocco.
Conclusions
The detection of BTV positivity in Culicoides spp. (lacking wing spots that allow their differentiation according to morphological identification keys) suggested that other Culicoides species are competent for BTV transmission in Morocco. The study results indicated, for the first time at the molecular level, that C. imicola and C. newsteadi are the primary potential vectors of BTV in Morocco and that C. paolae and C. circumscriptus are strongly implicated in the propagation of bluetongue at the national level.
Graphical Abstract

Similar content being viewed by others


Background
Bluetongue disease is a non-contagious viral disease affecting domestic and wild ruminants, and it is subject to mandatory notification to the World Organization for Animal Health (WOAH). It is primarily transmitted through Culicoides bites during a blood meal, resulting in significant economic losses [1]. Bluetongue virus (BTV) belongs to the Orbivirus genus of the Reoviridae family, characterized by a double-stranded RNA genome with 10 segments coding for various proteins. Currently, at least 36 different BTV serotypes are circulating worldwide [2,3,4].
Bluetongue first emerged in Morocco in 1956 and reappeared in 2004, triggering a severe epizootic. Since then, the disease has become enzootic, with several outbreaks reported annually throughout the country involving various serotypes [5,6,7,8,9]. Previous entomological investigations in Morocco have identified 56 Culicoides species, including at least eight species that are the principal or potential vectors of animal viruses [10,11,12,13,14,15,16,17].
A series of events occurs in female midges following ingestion of the virus during their blood meal. These include the liberation of the virus, followed by the infection of the midgut cells, proliferation within these cells, escape from the midgut, spreading into extraintestinal tissues, infection of the salivary glands located between the head and thorax, and finally, transmission of the virus to the subsequent host through the salivary glands. The viral particles must break through various barriers to infection within the abdomen, thorax, and head to successfully disseminate throughout the insect body. These obstacles influence the virus titer and its prevalence during infection, impacting pathogen transmission rates during feeding events. Moreover, the insect's susceptibility or refractoriness to a virus is related to extrinsic factors such as gut microbiota, as well as heritable genetic features like antiviral immune defenses in the insect. These factors are crucial for understanding the vector competence of a midge [18,19,20,21,22,23].
Detection, quantification, and isolation of the virus in field-collected Culicoides midges, analysis of infection levels under natural conditions, and laboratory infection trials are methods for assessing the vectorial competence of Culicoides. The species vectors of BTV have been determined accordingly [24,25,26]. Approximately 30 Culicoides species have been associated with BTV transmission, among which C. imicola is recognized as the major vector in Africa [27, 28]. Due to challenges in conducting studies on Culicoides vector competency, natural infection rates are generally considered a reliable approximation of the vector role [26].
The endemic status of bluetongue in Morocco underscores the importance of identifying Culicoides species vectors of the disease to implement targeted management strategies. Consequently, assessing the infection levels of these species is imperative for a deeper understanding of the transmission dynamics of the disease.
The current study marks the first molecular investigation in Morocco to investigate the potential role of Culicoides imicola, Culicoides paolae, Culicoides newsteadi, Culicoides spp., and Culicoides circumscriptus, collected during entomologic investigations conducted between 2018 and 2021 in different Moroccan regions. The study aimed to assess their involvement in BTV transmission, explore vectorial activity seasons, and investigate their epidemiological contribution and vectorial competence. This was achieved through the detection of virus dissemination in the bodies of field-caught parous females of these species, and the examination of infection rates using two estimates, the minimum infection rate (MIR) and the maximum likelihood estimation (MLE).
Methods
Culicoides field collection
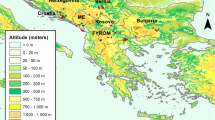
Parous females of C. imicola, C. paolae, C. newsteadi, Culicoides spp., and C. circumscriptus were collected using Onderstepoort Veterinary Institute (OVI) light traps across different regions in Morocco (6 out of 12) from 2018 to 2021 (Fig. 1). Species were identified using relevant morphological keys [29, 30]. The designation Culicoides spp. is assigned to those lacking wing spots, aiding differentiation according to identification keys. The midges were collected in a solution specifically prepared in the laboratory, composed of ethylenediaminetetraacetic acid (EDTA), sodium citrate, ammonium sulfate, and sterile water. This solution was designed to preserve any potential viral RNA that could be present in Culicoides at ambient temperature during the night of capture, from sunset to sunrise. Subsequently, the Culicoides were transferred to 70% ethanol and stored at −20 °C.

Regions of origin for Culicoides species investigated in the Kingdom of Morocco from 2018 to 2021
Detection of BTV infection in Culicoides midges
A total of 2003 specimens were grouped into 55 pools, each containing no more than 45 midges from the same zone and catch time.
The midges were dissected and separated into three parts: head, thorax, and abdomen. Each body region was collected in a separate subpool. Thus, every pool was subdivided into three subpools.
Viral RNA was isolated from the supernatant obtained by grinding the insect pools in phosphate-buffered saline (PBS) using the MagMAX™ Viral RNA Isolation Kit (Applied Biosystems) according to the manufacturer's instructions.
Reverse transcription quantitative polymerase chain reaction (RT-qPCR) assays were performed using the LSI VetMAX™ BTV NS3 All Genotypes Kit (Applied Biosystems) in accordance with the manufacturer's instructions. The kit includes two sets of primers/TaqMan probes, one labeled FAM™–NFQ (non-fluorescent quencher), encoding segment 10 of BTV RNA, and the other labeled VIC™–TAMRA™, for the internal positive control.
Reverse transcription and amplification were performed on an Applied Biosystems 7500 Fast Real-Time PCR system with the following thermal cycler program: 45 °C for 10 min, 95 °C for 10 min, followed by 40 cycles of denaturation (95 °C for 15 s) and annealing/extension (60 °C for 45 s). The cycle threshold (Ct) value was defined once the fluorescent signal breached a threshold fluorescence line. An insect pool with a Ct lower than 40 on the FAM-NFQ detector was considered BTV-positive.
Data processing and statistical analysis
QGIS 3.30.1 software was used to construct a map showing the regions of origin of the studied Culicoides species in Morocco.
The data were analyzed using R software (version 4.2.3). Infection rates for Culicoides were calculated using the PooledInfRate package, with a confidence interval (CI) of 95% and a scale of 100. MLE estimate infection rates based on probabilistic models following a binomial distribution. The MIR defines the minimum number of infected individuals per 100 midges tested. Pearson’s Chi-square and Fisher’s exact tests were used to compare Culicoides pool positivity rates by species and season. The simultaneous application of both tests strengthened the validity of the results. Differences were considered statistically significant at P < 0.05 and a 95% CI.
Results
RT-qPCR results for Culicoides batches
The individual analysis of the three body regions of the specimens within each pool revealed similar results. The Ct values of the head, thorax, and abdomen subpools were similar for each origin pool (Table 1). Consequently, RT-qPCR results are presented based on the pools of origin.
Among the 55 Culicoides batches screened for BTV, 45 tested positive (81.8%). BTV was detected in 15 of 16 batches of C. imicola (93.7%), 12 of 17 of C. paolae (70.6%), 9 of 10 of C. newsteadi (90%), six of eight of Culicoides spp. (75%), and three of four batches tested of C. circumscriptus (75%) (Fig. 2).

Batches of Culicoides species tested for bluetongue virus by RT-qPCR
The statistical analysis showed no significant differences among batches of different species (Chi-square test, χ 2 = 3.797, df = 4, P = 0.434; Fisherʼs exact test, P = 0.374).
The viral loads of positive pools were significant for all tested species, with Ct < 30. Ct values for positive pools of C. imicola ranged between 19.6 and 29.5, C. paolae between 24.7 and 29, C. newsteadi between 18.4 and 29.8, Culicoides spp. between 22 and 25.8, and C. circumscriptus between 21.1 and 26.6. The highest virus loads were observed in the head, thorax, and abdomen of C. imicola and C. newsteadi (Ct < 20) (Table 1).
RT-qPCR results for Culicoides pools according to season
The different batches comprised specimens collected during three seasons (spring, summer, and autumn). The analysis resulted in nine Culicoides-positive pools out of 10 collected in spring (90%), 25 out of 32 in summer (78.1%), and 11 out of 13 in autumn (84.6%).
In spring, all tested batches of C. imicola were positive (2/2), as were seven out of eight of C. newsteadi. For the summer pools, four of five C. imicola, 12 of 17 C. paolae, three of four C. circumscriptus, and all Culicoides spp. pools tested (6/6) were positive. In the autumn population, all pools of C. imicola (9) and C. newsteadi (2) were positive, while both batches of Culicoides spp. tested were negative (Fig. 3).

The batches of Culicoides species screened for bluetongue virus by RT-qPCR per season
The statistical analysis revealed no significant differences among collection seasons (Chi-square test, χ 2 = 0.812, df = 2, P = 0.666; Fisherʼs exact test, P = 0.891).
The infection rates of bluetongue virus in Culicoides species (MIR and MLE)
The MIR and MLE are key indicators of Culicoides dynamics in BTV transmission. Assuming that a PCR-positive batch includes at least one infected female, the average MIR for Culicoides was 2.2% (95% CI: 1.6–2.9%). However, MLE directly estimates the proportion of infected insects in the pool, giving an average value of 4.9% (95% CI: 3.3–6.4%). Both MIR and MLE were calculated to understand the distribution of bluetongue within the dominant Culicoides species in Morocco and across the seasons covered by the survey.
Culicoides imicola, C. paolae, C. newsteadi, Culicoides spp., and C. circumscriptus showed comparable MIRs, ranging from 1.9 to 2.8% (Table 2). The highest MLE rates were found in C. imicola (7.5%) and C. newsteadi (5.8%), while the other species also showed significant MLE values between 3.5 and 4.6% (Fig. 4A).

Variations in the infection rates of bluetongue virus (MIR and MLE) in Culicoides midges tested across species (A) and seasons (B). BTV bluetongue virus; MIR minimum infection rate; MLE maximum likelihood estimation
The infection rates of Culicoides for three seasons of capture, spring, summer, and autumn, were similarly high (Table 3), ranging from 2.2 to 2.4% for MIR and 4.4 to 5.8% for MLE. The highest rates were reached in autumn and spring (Fig. 4B).
Discussion
The parous females of C. imicola, C. paolae, C. newsteadi, Culicoides spp., and C. circumscriptus collected across different regions of Morocco (6 out of 12 Moroccan regions) were screened for BTV RNA using RT-qPCR. The analysis revealed BTV positivity in 45 of the 55 batches tested. The detection of the virus in the head, thorax (containing salivary glands), and abdomen with narrow Ct values (Ct < 30) for each species indicated BTV replication in the body of these species and their potential implication as BTV vectors [25, 31]. An understanding of the barriers that control the dissemination of BTV within the Culicoides body is essential for studying their competence and susceptibility to infection.
Culicoides females ingested approximately 100 TCID50 (50% tissue culture infectious dose; an average of 106 TCID50/ml of virus) during a blood meal of typically 10–4 ml. After feeding, the viral concentration decreased following inactivation of part of the viral particles in the intestine and excretion via the anus. Subsequently, the viral load increased, reaching a plateau of around 5–6 log10 TCID50 a week later, corresponding to a viral multiplication of 103 to 104 per midge. The intestinal wall represents the first barrier—the mesenteron infection barrier (MIB)—crossed by the virus. Only the particles remaining in the intestinal lumen can infect intestinal cells. To enter the hemocoel, the virions infecting the intestinal cells must face a second barrier, the mesenteron escape barrier (MEB). Once inside the hemocoel, these viral particles contaminate secondary organs. Before reaching the salivary glands, particles infect adipocytes, representing the third barrier, the dissemination barrier (DB), which can destroy part of the virus. At higher concentrations, viral particles can successfully cross the fourth barrier, the salivary gland infection barrier (SGIB), to penetrate the salivary glands. However, for an insect to infect a new susceptible host, virus particles must also overcome the fifth and final barrier, the salivary gland escape barrier (SGEB), which depends on genetic factors [32, 33]. The interplay between the virus and the insect’s innate immune system, expressed at the barriers, plays a crucial role in the outcome of virus infection and vector survival. The primary antiviral defense mechanism involves the RNA interference (RNAi) pathway, reacting to the detection of the virus-derived double-stranded RNA to suppress its replication. Other innate immune pathways such as Toll, IMD (immune deficiency), and JAK/STAT (Janus kinase/signal transducer and activator of transcription), as well as the autophagy pathway, are also implicated in regulating immune responses in Culicoides. Nonetheless, the midges display refractory and permissive lineages, underlining the heritability of traits linked to their vectorial competence. This indicates that Culicoides exhibit inter-individual variability in vector competence, relating to their transcriptomes and the genomic characteristics of refractory and permissive lineages [21, 22, 34,35,36,37]. Accordingly, the mechanisms of virus transmission by Culicoides can differ according to the virus in question and Culicoides species. That is why certain Culicoides species may not express some of these barriers, as observed in Culicoides variipennis, which does not appear to exhibit SGIB or SGEB for BTV [23, 38]. Indeed, detecting the BTV in the head and thorax after it has crossed several barriers could indicate its successful dissemination within the Culicoides body [20, 23].
The calculation of MIR and MLE is essential for assessing the transmission risk of vector-borne diseases. MIR estimates the minimum number of infected vector species among a given sample of vectors, while the MLE accurately estimates the infection prevalence in the vector population. The interpretation of infection rates, especially for low abundance species with small batch sizes, can lead to abnormally high rates. Calculating infection rates using both indices, with MLE considering the size of the tested pools, helps overcome this bias [26]. These infection rates can be employed to evaluate the vectorial competence of Culicoides [24]. The high infection rates (≥ 1‰) recorded in field populations in the current study suggest a significant risk of BTV transmission in Morocco [39].
The findings revealed active BTV circulation during the survey seasons (spring, summer, and autumn), with increased proportions of positivity. Although the statistical difference in pool positivity between seasons and among species was not statistically significant, it suggests a similar potential for different species to transmit the virus across the three seasons. Nevertheless, the highest MIR and MLE infection rates were observed in autumn and spring. These collection seasons are associated with favorable circumstances for vector activity, thereby increasing the opportunities for BTV transmission [40, 41].
The roles of C. imicola and C. newsteadi as BTV vectors have previously been identified in the literature [25, 42,43,44,45,46]. Their abundance, along with their highest positivity and infectious rates, confirms, for the first time, their role as a primary potential vector of BTV in Morocco.
Culicoides paolae was initially implicated in BTV transmission in Sardinia in 2017, and C. circumscriptus has recently been found BTV-positive in Turkey and Sardinia [42, 47]. Both species were frequently collected in the current study, and their tested pools exhibited strong positivity for BTV. Higher MIR and MLE values were associated with these Culicoides species, indicating their greater susceptibility to BTV. As a result, this study was the first to highlight the implications of C. paolae and C. circumscriptus as potential vectors of BTV at the national level.
Meanwhile, numerous specimens of Culicoides spp. lacking wing spots and posing identification challenges were captured en masse in this survey. Wing spot patterns are commonly used to identify Culicoides species. However, Culicoides can also be identified based on features other than wing spots. These include microscopic details of the antennae, maxillary palps, and genitalia, as well as morphological characteristics such as body and wing size, the coloration of the dorsum of the thorax, type and number of antennal sensilla, and leg color patterns. This approach is time-consuming and impractical for a large number of individual midges. To address this limitation, molecular methods can be used for the rapid identification of Culicoides species [13, 29, 48,49,50,51]. Given the frequency of these Culicoides spp. in Morocco, RT-qPCR analysis was carried out on batches of these midges to detect BTV and assess their possible contribution to its spread in the country. The pools exhibited impressive infection rates, highlighting the potential role of these midges in BTV epidemiology. This emphasizes the need for more detailed molecular analysis to identify them, as other Culicoides species could be competent to transmit BTV in Morocco [48, 52,53,54].
Several criteria are required to confirm that an arthropod serves as an arbovirus vector. Firstly, it is necessary to assess the abundance and wide distribution of the suspected arthropod. Secondly, it is imperative to demonstrate that this arthropod has fed on a susceptible host. Furthermore, the virus must be isolated from naturally infected arthropods. It is also important to prove the ability of the arthropod to become infected by feeding either on a viremic host or on a laboratory-infected blood meal. Ultimately, the ability of the infected arthropod to transmit the virus to a susceptible host during hematophagy needs to be confirmed [55,56,57]. While BTV infection levels were high and the virus was well-spread in all Culicoides body regions, confirming the competence of the vector requires further validation through virus isolation and experimental infection studies [25].
BTV was detected in small ruminants in the same study regions during the same survey period [7], providing evidence of the active circulation of the virus and the impact of these Culicoides species in bluetongue outbreaks in Morocco. The diversity of BTV-vector Culicoides species in the Moroccan fauna poses a potential threat to livestock herds, reinforcing the importance of implementing effective management strategies and vector control programs.
Conclusions
The current study provided the first molecular evidence illustrating the involvement of C. imicola, C. paolae, C. newsteadi, Culicoides spp., and C. circumscriptus as potential vectors for BTV propagation at the national level, along with insights into the vector activity seasons of these insects. BTV detection in parous females of field species, after the virus has crossed multiple body barriers, and the analysis of infection rates (MIR and MLE) for these midge populations served to assess the vectorial competence of Culicoides. Indeed, the results confirmed the implication of the studied species (C. imicola, C. paolae, C. newsteadi, Culicoides spp., and C. circumscriptus) as potential BTV vectors in bluetongue epidemics in Morocco, with active virus circulation during spring, summer, and autumn.
Availability of data and materials
All data generated or analyzed during this study are included in this published article.
Abbreviations
- BTV:
-
Bluetongue virus
- OVI:
-
Onderstepoort Veterinary Institute
- RT-PCR:
-
Reverse transcription polymerase chain reaction
- MLE:
-
Maximum likelihood estimation
- MIR:
-
Minimum infection rate
- WOAH:
-
World Organization for Animal Health
- EDTA:
-
Ethylenediaminetetraacetic acid
- PBS:
-
Phosphate-buffered saline
- RT-qPCR:
-
Reverse transcription quantitative polymerase chain reaction
- Ct:
-
Cycle threshold
- CI:
-
Confidence interval
- TCID50:
-
50% Tissue culture infectious dose
- MIB:
-
Mesenteron infection barrier
- MEB:
-
Mesenteron escape barrier
- DB:
-
Dissemination barrier
- SGIB:
-
Salivary gland infection barrier
- SGEB:
-
Salivary gland escape barrier
- RNAi:
-
RNA interference
- IMD:
-
Immune deficiency
- JAK/STAT:
-
Janus kinase/signal transducer and activator of transcription
References
Rojas JM, Rodríguez-Martín D, Martín V, Sevilla N. Diagnosing bluetongue virus in domestic ruminants: current perspectives. Vet Med Res Rep. 2019;10:17–27.
Attoui H, Mohd Jaafar F, Monsion B, Klonjkowski B, Reid E, Fay PC, et al. Increased clinical signs and mortality in IFNAR(−/−) mice immunised with the bluetongue virus outer-capsid proteins VP2 or VP5, after challenge with an attenuated heterologous serotype. Pathogens. 2023;12:602.
Zientara PS, Gonzalez G. Actualités sur deux maladies virales vectorielles zoonotiques en Europe: West Nile et Usutu - Réémergence du sérotype 8 du virus de la fièvre catarrhale ovine (FCO). Bull Acad Vet Fr. 2023;176:1–5.
Subhadra S, Sreenivasulu D, Pattnaik R, Panda BK, Kumar S. Bluetongue virus: past, present, and future scope. J Infect Dev Ctries. 2023;17:147–56.
Placidi L. La “Blue-tongue” au Maroc. Bull Acad Vet Fr. 1957;110:79–84.
Lhor Y, Kyriaki N, Khayli M, Mm B, Fassi Fihri O, El Harrak M. Bluetongue in Morocco 2004 to 2015: an overview. J Infect Dis Epidemiol. 2017;3.
Daif S, El Berbri I, Lhor Y, Fassi FO. Serological and molecular prevalence study of bluetongue virus in small domestic ruminants in Morocco. Sci Rep. 2022;12:19448.
Drif K. Bluetongue virus (BTV) serological survey and evidence of emergent BTV-8 serotype in Morocco. J Agric Sci Technol. 2014;4:353–8.
Drif K, Venter G, El Harrak M, Fassi Fihri O, Loutfi C, Touil N, et al. The value of camels as sentinels for bluetongue virus in Morocco. Vet Ital. 2021;54:343–8.
Bourquia M. Déterminants environnementaux de la distribution des Culicoides (Diptera: Ceratopogonidae), moucherons vecteurs de virus animaux d’intérêt économique, au Maroc. 2019. https://www.theses.fr/2019NSAM0022. Accessed 22 Feb 2023.
Bourquia M, Garros C, Rakotoarivony I, Gardes L, Huber K, Boukhari I, et al. Update of the species checklist of Culicoides Latreille, 1809 biting midges (Diptera: Ceratopogonidae) of Morocco. Parasit Vectors. 2019;12:459.
Kremer M, Hommel M, Bailly-Choumara H. Troisième contribution à l’étude faunistique des Culicoides du Maroc. Ann Parasitol Hum Comp. 1971;46:661–70.
Kremer M, Rebholtz-Hirtzel C, Delecolle JC. Quatrième contribution à l’étude faunistique des Culicoides (Diptera, Ceratopogonidae) du Maroc : description de C. landauae n. sp. (Diptera, Ceratopogonidae), redescription de C. fagbibi Navai et dune forme de C. subfascipennis Kieffer. Entomol Méd Parasitol. 1975;13:205–14.
Kremer M, Delecolle JC, Bailly-Choumara H, Chaker E. Cinquième contribution à l’étude faunistique des Culicoides (Diptera, Ceratopogonidae) du Maroc : description de C. calloti nsp. Entomol Méd Parasitol. 1979;17:195–9.
Bailly-Choumara H. Deuxième contribution à l’étude des Culicoides du Maroc (Diptera, Ceratopogonidae). Entomol Méd Parasitol. 1970;4:383–91.
Callot J, Kremer M, Bailly-Choumara H. Note faunistique préliminaire sur les Culicoides (Diptera, Ceratopogonidae) du Maroc. Gynandromorphisme de C. circumscriptus parasite par un mermis. Bull Soc Pathol Exot. 1968;61:885–9.
Kettani K, Ebejer M, Ackland D, Bächli G, Barraclough D, Bartak M, et al. Catalogue of the Diptera (Insecta) of Morocco—an annotated checklist, with distributions and a bibliography. ZooKeys. 2022;1094:1–466.
Feenstra F, Drolet BS, Boonstra J, van Rijn PA. Non-structural protein NS3/NS3a is required for propagation of bluetongue virus in Culicoides sonorensis. Parasit Vectors. 2015;8:476.
Baylis M, O’Connell L, Mellor PS. Rates of bluetongue virus transmission between Culicoides sonorensis and sheep. Med Vet Entomol. 2008;22:228–37.
Koenraadt CJ, Balenghien T, Carpenter S, Ducheyne E, Elbers AR, Fife M, et al. Bluetongue, Schmallenberg - what is next? Culicoides-borne viral diseases in the 21st century. BMC Vet Res. 2014;10:77.
Mills MK, Michel K, Pfannenstiel RS, Ruder MG, Veronesi E, Nayduch D. Culicoides–virus interactions: infection barriers and possible factors underlying vector competence. Curr Opin Insect Sci. 2017;22:7–15.
Tabachnick WJ. Genetic control of oral susceptibility to infection of Culicoides variipennis with bluetongue virus. Am J Trop Med Hyg. 1991;45:666–71.
Fu H, Leake CJ, Mertens PPC, Mellor PS. The barriers to bluetongue virus infection, dissemination and transmission in the vector, Culicoides variipennis (Diptera: Ceratopogonidae). Arch Virol. 1999;144:747–61.
Oliveira ARS, Cohnstaedt LW, Strathe E, Hernández LE, McVey DS, Piaggio J, et al. Meta-analyses of the proportion of Japanese encephalitis virus infection in vectors and vertebrate hosts. Parasit Vectors. 2017;10:418.
Foxi C, Delrio G, Falchi G, Marche MG, Satta G, Ruiu L. Role of different Culicoides vectors (Diptera: Ceratopogonidae) in bluetongue virus transmission and overwintering in Sardinia (Italy). Parasit Vectors. 2016;9:440.
Grimaud PY. Dynamique des populations de Culicoides à l’île de La Réunion, moucherons vecteurs d’orbiviroses. 2019. https://theses.hal.science/tel-02468114. Accessed 10 Jan 2023.
Meiswinkel R, Gomulski LM, Delécolle JC, Goffredo M, Gasperi G. The taxonomy of Culicoides vector complexes—unfinished business. Vet Ital. 2004;40:151–9.
Wilson AJ, Mellor PS. Bluetongue in Europe: past, present and future. Philos Trans R Soc Lond B Biol Sci. 2009;364:2669–81.
Mathieu B, Cêtre-Sossah C, Garros C, Chavernac D, Balenghien T, Carpenter S, et al. Development and validation of IIKC: an interactive identification key for Culicoides (Diptera: Ceratopogonidae) females from the Western Palaearctic region. Parasit Vectors. 2012;5:137.
Delecolle JC. Nouvelle contribution à l’étude systématique et iconographique des espèces du genre Culicoides, (Diptéra: Cératopogonidae) du Nord-Est de la France. 1985. https://search.worldcat.org/. Accessed 16 Apr 2018.
Elbers ARW, Meiswinkel R, van Weezep E, van Sloet Oldruitenborgh-Oosterbaan MM, Kooi EA. Schmallenberg virus in Culicoides spp. biting midges, the Netherlands, 2011. Emerg Infect Dis. 2013;19:106–9.
Perie P, Chermette R, Millemann Y, Zientara S. Les Culicoides, Diptères hématophages vecteurs de la fièvre catarrhale du mouton. Bull Acad Vet Fr. 2005;158:213–24.
Mellor PS. The replication of bluetongue virus in Culicoides vectors. In: Roy P, Gorman BM, editors. Bluetongue viruses. Berlin: Springer; 1990. p. 143–61.
Mills MK, Nayduch D, Michel K. Inducing RNA interference in the arbovirus vector, Culicoides Sonorensis. Insect Mol Biol. 2015;24:105–14.
Schnettler E, Ratinier M, Watson M, Shaw AE, McFarlane M, Varela M, et al. RNA interference targets arbovirus replication in Culicoides Cells. J Virol. 2013;87:2441–54.
Kingsolver MB, Huang Z, Hardy RW. Insect antiviral innate immunity: pathways, effectors, and connections. J Mol Biol. 2013;425:4921–36.
Tabachnick WJ. Genetic variation in laboratory and field populations of the vector of bluetongue virus, Culicoides variipennis (Diptera: Ceratopogonidae). J Med Entomol. 1990;27:24–30.
Mellor PS, Carpenter S, White DM. Bluetongue virus in the insect host. In: Mellor P, Baylis M, Mertens P, editors. Bluetongue. London: Elsevier; 2009. p. 295–320.
Kęsik-Maliszewska J, Larska M, Collins ÁB, Rola J. Post-epidemic distribution of schmallenberg virus in Culicoides arbovirus vectors in Poland. Viruses. 2019;11:447.
Lhor Y, Khayli M, Bouslikhane M, El Harrak M, Fassi FO. Spatial and seasonal distribution of Culicoides species in Morocco in relation to the transmission of bluetongue viruses. Br J Virol. 2015;2:88–95.
Ander M, Meiswinkel R, Chirico J. Seasonal dynamics of biting midges (Diptera: Ceratopogonidae: Culicoides), the potential vectors of bluetongue virus, in Sweden. Vet Parasitol. 2012;184:59–67.
Foxi C, Meloni G, Puggioni G, Manunta D, Rocchigiani A, Vento L, et al. Bluetongue virus detection in new Culicoides species in Sardinia. Italy Vet Rec. 2019;184:621–621.
Sghaier. New species of the genus Culicoides (Diptera Ceratopogonidae) for Tunisia, with detection of bluetongue viruses in vectors. Vet Ital. 2017;53:357–66.
Quaglia M. Culicoides species responsible for the transmission of epizootic haemorrhagic disease virus (EHDV) serotype 8 in Italy. Vet Ital. 2023;59:83–9. https://doi.org/10.12834/VetIt.3347.22208.1.
Fujisawa Y, Homat T, Thepparat A, Changbunjong T, Sutummaporn K, Kornmatitsuk S, et al. DNA barcode identification and molecular detection of bluetongue virus in Culicoides biting midges (Diptera: Ceratopogonidae) from Western Thailand. Acta Trop. 2021;224:106147.
Goffredo M, Catalani M, Federici V, Portanti O, Marini V, Mancini G, et al. Vector species of Culicoides midges implicated in the 2012–2014 bluetongue epidemics in Italy. Vet Ital. 2015;51:131–8.
Yavru S, Dik B, Bulut O, Uslu U, Yapici O, Kale M, et al. New Culicoides vector species for BTV transmission in Central and Central West of Anatolia. Annu Res Rev Biol. 2018;27:1–9.
Zhang X, Phillips RA, Gerry AC. Morphological and molecular identification of Culicoides (Diptera: Ceratopogonidae) species of the Southern California desert. J Med Entomol. 2022;59:1589–600.
Miranda MÁ. Case studies of vector-borne diseases in livestock: bluetongue virus. In: Garros C, Bouyer J, Takken W, Smallegange RC, editors. Pests and vector-borne diseases in the livestock industry. Wageningen: Wageningen Academic Publishers; 2018. p. 221–71.
Harrup LE, Bellis GA, Balenghien T, Garros C. Culicoides Latreille (Diptera: Ceratopogonidae) taxonomy: current challenges and future directions. Infect Genet Evol. 2015;30:249–66.
Pagès N, Muñoz-Muñoz F, Talavera S, Sarto V, Lorca C, Núñez JI. Identification of cryptic species of Culicoides (Diptera: Ceratopogonidae) in the subgenus Culicoides and development of species-specific PCR assays based on barcode regions. Vet Parasitol. 2009;165:298–310.
Nolan DV, Carpenter S, Barber J, Mellor PS, Dallas JF, Mordue Luntz AJ, et al. Rapid diagnostic PCR assays for members of the Culicoides obsoletus and Culicoides pulicaris species complexes, implicated vectors of bluetongue virus in Europe. Vet Microbiol. 2007;124:82–94.
Shults P, Moran M, Blumenfeld AJ, Vargo EL, Cohnstaedt LW, Eyer PA. Development of microsatellite markers for population genetics of biting midges and a potential tool for species identification of Culicoides sonorensis Wirth & Jones. Parasit Vectors. 2022;15:69.
Nielsen SA, Kristensen M. Delineation of Culicoides species by morphology and barcode exemplified by three new species of the subgenus Culicoides (Diptera: Ceratopogonidae) from Scandinavia. Parasit Vectors. 2015;8:151.
Tanya VN, Greiner EC, Gibbs EPJ. Evaluation of Culicoides insignis (Diptera: Ceratopogonidae) as a vector of bluetongue virus. Vet Microbiol. 1992;32:1–14.
Standfast HA, Dyce AL. Potential vectors of arboviruses of cattle and buffalo in Australia. Aust Vet J. 1972;48:224–7.
DeFoliart GR, Grimstad PR, Watts DM. Advances in mosquito-borne arbovirus/vector research. Annu Rev Entomol. 1987;32:479–505.
Acknowledgements
The authors would like to acknowledge with gratitude the National Office of Food Safety (ONSSA) for their assistance in facilitating access to farms for fieldwork.
Funding
This research received funding from the EU Horizon 2020 project “PALE-Blu-Understanding pathogens, livestock environment, interactions involving bluetongue,” under Grant Agreement No. 727393. The funders had no role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
OFF, IEB, and SD designed and coordinated the work. SD and IEB led the actions in the field. SD pre-sorted and identified the Culicoides, determined the physiological stages of the females, conducted the laboratory analysis, analyzed and interpreted the data, prepared the maps, performed the statistical analysis, produced all the figures, and wrote the manuscript. OFF and IEB revised the manuscript. All the authors have read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Daif, S., El Berbri, I. & Fassi Fihri, O. First molecular evidence of potential Culicoides vectors implicated in bluetongue virus transmission in Morocco. Parasites Vectors 17, 71 (2024). https://doi.org/10.1186/s13071-024-06167-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-024-06167-y