
Abstract
Background
Theileria equi causes equine piroplasmosis, an economically significant disease that affects horses and other equids worldwide. Based on 18S ribosomal RNA (18S rRNA sequences), T. equi can be classified into five genotypes: A, B, C, D, and E. These genotypes have implications for disease management and control. However, no conventional polymerase chain reaction (PCR) assays are available to differentiate the genotypes of T. equi. To overcome this limitation, we developed and evaluated PCR assays specific for the detection of each T. equi genotype.
Methods
A pair of forward and reverse primers, specifically targeting the 18S rRNA sequence of each genotype, was designed. The genotype-specific PCR assays were evaluated for their specificity using plasmids containing inserts of the 18S rRNA sequence of each genotype. Subsequently, the assays were tested on 270 T. equi-positive equine blood DNA samples (92 from donkeys in Sri Lanka and 178 from horses in Paraguay). 18S rRNA sequences derived from the PCR amplicons were analyzed phylogenetically.
Results
Each genotype-specific PCR assay accurately targeted the intended genotype, and did not produce any amplicons when 18S rRNA from other T. equi genotypes or genomic DNA of Babesia caballi or uninfected horse blood was used as the template. Previous studies employing PCR sequencing methods identified T. equi genotypes C and D in the Sri Lankan samples, and genotypes A and C in the Paraguayan samples. In contrast, our PCR assay demonstrated exceptional sensitivity by detecting four genotypes (A, C, D, and E) in the Sri Lankan samples and all five genotypes in the Paraguayan samples. All the Sri Lankan samples and 93.3% of the Paraguayan samples tested positive for at least one genotype, further emphasizing the sensitivity of our assays. The PCR assays also had the ability to detect co-infections, where multiple genotypes in various combinations were detected in 90.2% and 22.5% of the Sri Lankan and Paraguayan samples, respectively. Furthermore, the sequences obtained from PCR amplicons clustered in the respective phylogenetic clades for each genotype, validating the specificity of our genotype-specific PCR assays.
Conclusions
The genotype-specific PCR assays developed in the present study are reliable tools for the differential detection of T. equi genotypes.
Graphical abstract

Similar content being viewed by others

Background
Equine piroplasmosis is a disease caused by Theileria equi and Babesia caballi in horses and other equids, including mules, donkeys, and zebras, and has a significant economic impact on the equine industry [1,2,3]. Both parasites have a complex life cycle that involves both equine hosts and tick vectors. In the equine host, T. equi and B. caballi reproduce asexually within infected erythrocytes, causing massive hemolysis, which leads to severe anemia and other clinical symptoms, such as fever, jaundice, hemoglobinuria, icterus, weight loss, and sometimes death [2]. In general, T. equi causes a more severe form of equine piroplasmosis as compared to B. caballi. Furthermore, although B. caballi-infected animals naturally clear the infection within 4 years, T. equi-infected animals become lifelong carriers, from which the infection can be spread to other equines via tick vectors [4,5,6,7]. Consequently, T. equi-infected carrier animals impede the international equine trade because importing countries, particularly those classified as free from equine piroplasmosis, impose stringent regulations on equine imports [6, 8]. Hence, the control of T. equi infection is crucial for an economically sustainable equine industry.
The currently available methods to control T. equi infection are heavily influenced by the genetic diversity of the parasite. On the basis of 18S ribosomal RNA gene (18S rRNA) sequences, T. equi is classified into five genotypes, named A, B, C, D, and E [9,10,11]; these can affect the diagnostic results, clinical outcome of infection, and therapeutic efficacy [12,13,14]. The World Organization for Animal Health recommends competitive enzyme-linked immunosorbent assays (cELISAs) and indirect fluorescent antibody tests [15,16,17,18] for the serodiagnosis of T. equi infection. cELISA is widely used because of its availability as a kit, predetermined cut-off value that makes the interpretation of test results simple, and suitability for analysis of a large number of samples. The cELISA for the serodiagnosis of T. equi infection has been developed based on the equi merozoite antigen 1 (EMA-1) of T. equi genotype A [16, 17]. However, a new Theileria species, Theileria haneyi, which belongs to genotype C of T. equi, lacks the ema-1 gene [10]. Therefore, it is unlikely that the cELISA can detect antibodies in animals infected with the genotype C. The absence of ema-1 in genotype C also implies that polymerase chain reaction (PCR) assays based on this gene are not suitable for detecting T. equi infection. Genotypic diversity of T. equi may also influence the outcome of infection, as previous studies have shown that genotype A is more commonly associated with clinical piroplasmosis than the other genotypes [19, 20]. Moreover, the genetic diversity of T. equi may be a determinant in drug-induced clearance of the parasite from infected horses, as suggested by the results of a previous study which demonstrated that imidocarb dipropionate eradicates T. equi genotype A but not T. haneyi [12, 13]. These diagnostic, clinical, and therapeutic implications highlight the importance of detecting the genotype for managing T. equi infection and facilitating the safe international transportation of equines.
Currently, PCR sequencing and real-time PCR assays are employed to determine T. equi genotypes [21,22,23,24,25]. However, these methods are not without limitations. In the PCR sequencing approach, the 18S rRNA is amplified from T. equi-positive samples, sequenced, and phylogenetically analyzed to detect the genotype of the parasite. However, in co-infected animals (i.e., those simultaneously infected with more than one T. equi genotype), this method is likely to detect the dominant genotype, potentially leaving the minor genotypes undetected [26, 27]. Real-time PCR assays can provide quick and quantitative results, but they require specialized equipment and expertise [22]. Additionally, the real-time PCR assays available for T. equi genotyping have not been validated for wide use in different geographic regions [22, 23]. Because the real-time PCR assays amplify only short fragments of 18S rRNA, validating the findings by sequencing analysis is challenging [22, 23].
On the contrary, conventional PCR assays are more accessible and cost-effective, and can be performed in resource-limited laboratories. Furthermore, the findings can be verified through sequencing analysis [28, 29]. However, conventional genotype-specific PCR assays have not yet been developed for T. equi. Therefore, in the present study, we developed 18S rRNA-based PCR assays for the genotype-specific detection of T. equi, and evaluated them using previously identified T. equi-positive equine DNA samples from Sri Lanka and Paraguay.
Methods
Primer design for genotype-specific PCR assays
Long sequences [~ 1600 base pairs (bp)] of 18S rRNA representing each T. equi genotype and those of B. caballi and equine hosts were retrieved from GenBank, and aligned using Multalin online software (http://multalin.toulouse.inra.fr/multalin/multalin.html) [30]. On the basis of the alignment, a pair of forward and reverse primers specific to each T. equi genotype was designed (Fig. 1a). Moreover, we assessed whether the PCR findings were verifiable through subsequent sequencing analysis of the PCR amplicons. Briefly, the 18S rRNA fragments targeted by the PCR assays specific to genotypes A, B, C, D, and E of T. equi were trimmed at both the 5ʹ- and 3ʹ- ends, and aligned. The resulting 687-bp alignment was then analyzed for nucleotide polymorphisms [31].

Development of genotype-specific polymerase chain reaction (PCR) assays for Theileria equi. a Primer design for T. equi genotype-specific PCR assays. 18S ribosomal RNA (18S rRNA) gene sequences of T. equi were aligned, and a set of forward and reverse primers specific to each genotype was designed. The specific binding regions (highlighted in gray) of the forward and reverse primers on the 18S rRNA sequences representing genotypes A (JX177673), B (EU642507), C (JQ390047), D (AB515312), and E (KF559357) are shown. The dashes represent gaps. b Nucleotide polymorphisms among the 18S rRNA sequences targeted by the genotype-specific PCR assays. The 18S rRNA sequences targeted by the PCR assays specific to T. equi genotypes A, B, C, D, and E were trimmed and aligned, and the resultant 687-base pair (bp) alignment was analyzed for nucleotide polymorphisms. The dots and dashes represent identical nucleotides and gaps, respectively, to those in the genotype A sequence. Conserved nucleotide polymorphisms in each genotype are shown
Plasmids containing 18S rRNA representing each T. equi genotype
Five plasmids, respectively containing an insert of 18S rRNA sequence representing each T. equi genotype, were prepared and used for specificity testing of the developed PCR assays. A long 18S rRNA fragment (1591 bp) of genotype A was amplified from DNA extracted from an in vitro culture (T. equi USDA strain) [32], using the forward primer Nbab_1F and reverse primer TB-rev [33, 34], as described previously [24]. The 18S rRNA of genotype B was amplified from a DNA template synthesized based on a Sudanese sequence (GenBank accession number AB515312). The resulting PCR amplicons were purified by using a QIAquick Gel Extraction Kit (Qiagen, Hilden, Germany), and then cloned into vector pCR™ 2.1-TOPO (Invitrogen, Carlsbad, CA). The plasmids were purified using a QIAprep Spin Miniprep Kit (Qiagen), and then sequenced. The sequencing analysis confirmed that the 18S rRNA inserts in the plasmids represented the respective genotypes. Plasmids with inserts of 18S rRNA that belong to genotypes C and D were obtained from a donkey survey in Sri Lanka [24], while a plasmid containing 18S rRNA of genotype E was prepared from a horse survey in Mongolia (unpublished data). In the aforementioned studies, the 18S rRNA sequences of genotypes C, D, and E were amplified using the primers Nbab_1F and TB-rev, and then cloned, following the same methodology as in the present investigation.
Specificity testing
Each PCR assay was evaluated for its ability to differentially detect the target genotype from the remaining genotypes of T. equi and DNA of B. caballi and noninfected horse blood. Briefly, plasmid DNA of each genotype, B. caballi DNA from an in vitro culture (USDA strain), and blood DNA from an uninfected horse were analyzed in each of the genotype-specific PCR assays. Each PCR assay was performed in a 10-μl reaction mixture, which included 1 µl of 0.01 pg/µl plasmid DNA, 5 μl of 2 × PCR buffer (KOD FX Neo; Toyobo, Osaka, Japan), 1 μl of 2 mM deoxynucleoside triphosphates (Toyobo), 0.5 μl of 10 μM forward and reverse primers (Table 1), 0.1 μl of 1 U/μl KOD FX Neo DNA polymerase (Toyobo), and 1.9 μl of double-distilled water. The PCR reaction mixture underwent an initial denaturation at 95 °C for 5 min, followed by 45 cycles, each including a denaturing step at 95 °C for 30 s, an annealing step at the appropriate temperature (see Table 1) for 1 min, and an extension step at 72 °C for 1 min. Following a final elongation step at 72 °C for 7 min, the PCR products were separated on 1.5% agarose gels, stained with MIDORI Green Xtra (Nippon Genetics, Tokyo, Japan), and then illuminated under ultraviolet light. PCR amplicons of the expected band size (Table 1) were considered to be positive for the respective genotype of T. equi. The resulting PCR products were cloned, and two clones per genotype were sequenced to confirm that each PCR assay had amplified the targeted fragment of 18S rRNA.
Validation of the PCR assays
A total of 270 T. equi-positive blood DNA samples from apparently healthy equines, including 92 donkeys from Sri Lanka [24] and 178 horses from Paraguay [35], were used to validate the newly developed genotype-specific PCR assays. Details regarding the gender, age, and breeds of the Paraguayan horses have been provided in previous studies [24, 35]. However, no any additional information is available for the free-roaming Sri Lankan donkeys. The DNA samples from blood samples of Sri Lankan donkeys and Paraguayan horses were extracted using the QIAamp DNA Blood Mini Kit (Qiagen) and DNAzol reagent (Thermo Fisher Scientific, Waltham, MA), respectively, according to the manufacturers’ instructions. Microscopic examination of thin blood smears indicated that 64 of the Sri Lankan donkeys were positive for T. equi [24]. Blood smears were not prepared from the surveyed horses in Paraguay [35]. Phylogenetic analyses were performed in previous studies by using 18S rRNA sequences prepared from a selected number of DNA samples, and detected genotypes C and D in the Sri Lankan donkeys [24] and genotypes A and C in the Paraguayan horses [35]. In the present study, all of the 270 samples were screened in each genotype-specific PCR assay. Randomly selected amplicons from Sri Lankan and Paraguayan samples were cloned for each PCR assay, and two clones per amplicon were sequenced.
Phylogenetic analysis
The Sri Lankan and Paraguayan 18S rRNA sequences derived from the amplicons from each T. equi genotype-specific PCR assay, along with those obtained from GenBank, were aligned using MAFFT online software (https://mafft.cbrc.jp/alignment/server/) [36]. The alignment was analyzed using MEGA X software [37] to predict the best nucleotide substitution model based on the lowest Akaike information criterion value. A maximum-likelihood phylogenetic tree was then constructed with MEGA, employing the Tamura-Nei substitution model [38].
Results
Development of genotype-specific PCR assays for T. equi
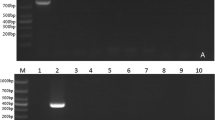
Five 18S rRNA-based PCR assays were developed in the present study for the differential detection of T. equi genotypes A, B, C, D, and E. Our in silico analysis showed that the 18S rRNA fragment amplified in each PCR assay is characterized by unique nucleotide polymorphisms, indicating that the results could be confirmed through subsequent sequencing analysis of PCR amplicons (Fig. 1b). The specificities of the genotype-specific PCR assays were evaluated using plasmids with inserts of 18S rRNA representing each T. equi genotype. We found that each PCR assay exclusively detected the target genotype, without any amplification observed when 18S rRNA of other genotypes, B. caballi DNA, or noninfected horse DNA were tested, thereby validating the specificity of the genotype-specific PCR assays (Fig. 2).

Specificity of the genotype-specific PCR assays for Theileria equi. Each genotype-specific PCR assay was evaluated for its ability to differentially detect the target genotype from the remaining genotypes and DNA of Babesia caballi and noninfected horse blood. The genotype-specific PCR assays were highly specific in detecting their target genotypes. A–E Plasmid DNA with inserts of 18S rRNA from T. equi genotypes A–E, respectively; BC DNA of B. caballi; H DNA of noninfected horse blood; N negative control (no template); M 100-bp DNA ladder; for other abbreviations, see Fig. 1
Field evaluation of genotype-specific PCR assays using T. equi-positive equine DNA samples
Our genotype-specific PCR assays were used to screen a total of 270 T. equi-positive DNA samples prepared from Sri Lankan donkeys (92 samples) and Paraguayan horses (178 samples). The surveyed donkeys in Sri Lanka were infected with four genotypes of T. equi, including A, C, D, and E (Table 2). The most common genotype was D, detected in 88 (95.7%) samples, followed by genotypes A, C, and E, which were detected in 75 (81.5%), 40 (43.5%), and 6 (6.5%) samples, respectively. All 92 T. equi-positive donkey samples were positive for at least one genotype (Additional file 1: Table S1). Co-infections were common: 83 (90.2%) donkeys were infected with two, three, or four genotypes (Table 3). Co-infection with genotypes A and D was the most frequently observed combination.
The genotype-specific PCR assays detected all five genotypes in Paraguayan horses. Genotype C was most prevalent, detected in 114 (64.0%) samples, followed by genotypes A, D, E, and B, which were detected in 76 (42.7%), 10 (5.6%), 8 (4.5%), and five (2.8%) samples, respectively. Of 178 T. equi-positive horses, 166 (93.3%) were positive for at least one genotype (Additional file 2: Table S2). Of the surveyed horses, 40 (22.5%) had co-infections with two or three T. equi genotypes (Table 3); co-infection with genotypes A and C was the most common combination.
To validate the PCR results, we sequenced four randomly selected amplicons from each PCR assay, except for genotype E from Sri Lanka and genotype B from Paraguay, for which three amplicons per genotype were sequenced. The resultant 34 18S rRNA sequences, including 15 Sri Lankan sequences representing genotypes A, C, D, and E, and 19 Paraguayan sequences representing all five genotypes, were registered with GenBank (accession nos. LC775884–LC775917) and used in the construction of a phylogenetic tree. All of the newly determined sequences from the respective PCR assays clustered in clades according to their genotype, together with known reference sequences (Fig. 3).

Phylogenetic analysis of Theileria equi 18S rRNA. The 18S rRNA sequences amplified in the genotype-specific PCR assays, together with reference sequences representing each genotype, were used to construct a maximum-likelihood phylogeny with 1000 bootstrap replicates. The newly determined sequences from donkeys from Sri Lanka (in red) and horses from Paraguay (in blue) from the amplicons from each genotype-specific PCR assay clustered in the corresponding clades. For abbreviations, see Fig. 1
Discussion
In the present study, we successfully developed a set of five specific PCR assays capable of differentiating the genotypes of T. equi. These newly developed PCR assays exhibited high specificity toward their target genotypes, as evaluated using 18S rRNA templates representing each of the genotypes. Subsequently, we validated the assays using T. equi-positive DNA samples prepared from donkeys from Sri Lanka and horses from Paraguay, focusing on their sensitivity, specificity, and ability to detect multiple genotypes in co-infected equines. We also investigated whether the findings were verifiable by subjecting the sequences of the PCR amplicons to phylogenetic analysis.
The genotype-specific PCR assays were highly sensitive in detecting the T. equi genotypes from field samples, as shown by the fact that all of the T. equi-positive samples from Sri Lanka and 93.3% of those from Paraguay were positive for at least one genotype. However, a small number of field samples from Paraguay (12/178) tested negative in all five genotype-specific PCR assays. The field DNA samples used in the present study had tested positive in a diagnostic PCR assay using a pair of primers that had been designed based on regions in 18S rRNA that are highly conserved among all T. equi genotypes [39]. Therefore, it is possible that the negative samples might have been co-infected with multiple genotypes, each with very low parasitemia, leading to negative results in the genotype-specific PCR assays, while the combined 18S rRNA templates from each genotype could have been sufficient to generate positive results in the diagnostic PCR assay [35].
In the PCR sequencing approach, only a subset of positive samples is subjected to sequencing analysis because of cost and time limitations. As a result, any additional genotypes present in untested samples may remain undetected. By contrast, genotype-specific PCR assays offer a convenient alternative for analyzing a large number of positive samples, and potentially detect all of the genotypes that are present [28, 29]. Previous studies using the PCR sequencing approach identified genotypes C and D in the samples from Sri Lanka and genotypes A and C in those from Paraguay [24, 35]. Notably, our genotype-specific PCR assays not only confirmed the presence of these genotypes, but also detected additional genotypes A and E in the samples from Sri Lanka and B, D, and E in the samples from Paraguay, which have not been previously reported in these countries [24, 35]. The detection of genotype A as the second most common genotype in both Sri Lanka and Paraguay is worrying because this genotype is more commonly associated with clinical piroplasmosis than the others [19, 20]. Similarly, the high rates of detection of genotype C suggest that the currently available cELISA may not be suitable for the serodiagnosis of T. equi infection in Sri Lanka and Paraguay [10].
Our PCR assays successfully detected co-infections with multiple genotypes of T. equi in both Sri Lankan and Paraguayan DNA samples. The common occurrence of co-infections might have implications for the control of T. equi. For example, a previous study found that repeated treatment with imidocarb dipropionate clears the parasites from horses singly infected with genotype A, but not from horses co-infected with genotype A and T. haneyi (genotype C) [13]. The ability of the genotype-specific PCR assays to detect co-infections, as demonstrated in the present study, overcomes the limitation of the PCR sequencing approach, which tends to detect only the dominant genotype in co-infected samples. Collectively, our findings demonstrate that these genotype-specific PCR assays are a potential alternative to the PCR sequencing approach for T. equi genotyping.
Our genotype-specific PCR assays require validation using a larger number of T. equi-positive samples from diverse geographic locations. We found that the 18S rRNA fragments targeted by the genotype-specific PCR assays contain unique nucleotide polymorphisms that can be readily used to verify the PCR results. Furthermore, the 18S rRNA sequences obtained from randomly selected amplicons from each of the genotype-specific PCR assays clustered in their respective phylogenetic clades. These findings not only confirm the specificity of our PCR assays but also suggest that the genotype-specific PCR assays can be validated easily.
The genotype-specific PCR assays developed in the present study have the potential to facilitate further research into the various implications of T. equi genotypes, such as for taxonomy, virulence, immunological cross-reactivity, infection persistence, diagnosis, and transmission. A recent study that compared the morphology and whole genome data of genotypes A and C concluded that genotype C represents a novel species, named T. haneyi [10], and suggested that several cryptic Theileria species have been collectively classified as T. equi. Similar comparisons of morphology and genome data among all T. equi genotypes are essential to uncover potential novel Theileria species [18]. Moreover, the potential for a specific genotype to be a risk factor for clinical piroplasmosis in infected horses remains uncertain. Genotype A appears to be most associated with clinical cases [19, 20], but to confirm this assumption, it would be necessary to comparatively evaluate the virulence of each genotype in experimental infections. Furthermore, not all genotype A infections in horses progress to clinical piroplasmosis [19, 20]. One possibility is that immunity to other genotypes may protect against the disease caused by genotype A, similar to observations with other Theileria species [40]. To explore this hypothesis, horses could be experimentally infected with each T. equi genotype, and then challenged with genotype A. However, if immunity to T. equi is genotype-specific, it would be important to investigate if the persistence of infection arises from co-infections. Monitoring horses infected with single and multiple genotypes for infection persistence could be a simple way to explore co-infections as a reason for infection persistence. Developing novel diagnostic assays capable of detecting all T. equi genotypes is vital to overcoming the limitations of current diagnostic methods [41]. However, progress in developing such tests has been slow because of the lack of research materials representing some genotypes. The implications of the genotype for the transmission of T. equi also remain uninvestigated. Several tick species belonging to the genera Amblyomma, Dermacentor, Hyalomma, and Rhipicephalus transmit T. equi [7, 42], but no studies have investigated whether the tick species that transmit T. equi are genotype specific. The initial step towards investigating the aforementioned research topics is identifying each genotype in infected hosts. We firmly believe that the genotype-specific PCR assays newly established here, which can be used even in resource-limited laboratories, will prove to be a valuable tool to expedite research focusing on the implications of T. equi genotypes.
Conclusions
The development and evaluation of genotype-specific PCR assays for T. equi represent a significant step toward enhancing our understanding of the genotypic diversity and distribution of this important pathogen. These assays offer a sensitive, specific, and efficient approach, suitable even for resource-constrained laboratories, for the differential detection of T. equi genotypes. Genotype-specific PCR assays have important implications for disease surveillance, control strategies, and further research in the field of equine piroplasmosis.
Availability of data and materials
All relevant data generated during the present study are included in this manuscript.
Abbreviations
- cELISA:
-
Competitive enzyme-linked immunosorbent assay
- EMA-1:
-
Equi merozoite antigen-1
- 18S rRNA:
-
18S ribosomal RNA
References
Knowles DP, Kappmeyer LS, Stiller D, Hennager SG, Perryman LE. Antibody to a recombinant merozoite protein epitope identifies horses infected with Babesia equi. J Clin Microbiol. 1992;30:3122–6.
Ueti MW, Palmer GH, Kappmeyer LS, Statdfield M, Scoles GA, Knowles DP. Ability of the vector tick Boophilus microplus to acquire and transmit Babesia equi following feeding on chronically infected horses with low level parasitaemia. J Clin Microbiol. 2005;43:3755–9.
Uilenberg G. Babesia—a historical overview. Vet Parasitol. 2006;138:3–10.
Heim A, Passos LM, Ribeiro MF, Costa-Junior LM, Bastos CV, Cabral DD, et al. Detection and molecular characterization of Babesia caballi and Theileria equi isolates from endemic areas of Brazil. Parasitol Res. 2007;102:63–8.
Zobba R, Ardu M, Niccolini S, Chessa B, Manna L, Cocco R, et al. Clinical and laboratory findings in equine piroplasmosis. J Equine Vet Sci. 2008;28:301–8.
Rothschild CM. Equine piroplasmosis. J Equine Vet Sci. 2013;33:497–508.
Wise LN, Pelzel-McCluskey AM, Mealey RH, Knowles DP. Equine piroplasmosis. Vet Clin North Am Equine Pract. 2014;30:677–93.
Friedhoff KT, Tenter AM, Müller I. Haemoparasites of equines: impact on international trade of horses. Rev Sci Tech. 1990;9:1187–94.
Bhoora R, Franssen L, Oosthuizen MC, Guthrie AJ, Zweygarth E, Penzhorn BL, et al. Sequence heterogeneity in the 18S rRNA gene within Theileria equi and Babesia caballi from horses in South Africa. Vet Parasitol. 2009;159:112–20.
Knowles DP, Kappmeyer LS, Haney D, Herndon DR, Fry LM, Munro JB, et al. Discovery of a novel species, Theileria haneyi n. sp., infective to equids, highlights exceptional genomic diversity within the genus Theileria: implications for apicomplexan parasite surveillance. Int J Parasitol. 2018;48:679–90.
Tirosh-Levy S, Gottlieb Y, Fry LM, Knowles DP, Steinman A. 20 years of equine piroplasmosis research: global distribution, molecular diagnosis, and phylogeny. Pathogens. 2020;9:926.
Ueti MW, Mealey RH, Kappmeyer LS, White SN, Kumpula-McWhirter N, Pelzel AM, et al. Reemergence of the apicomplexan Theileria equi in the United States: elimination of persistent infection and transmission risk. PLoS ONE. 2012;7:44713.
Sears K, Knowles D, Dinkel K, Mshelia PW, Onzere C, Silva M, et al. Imidocarb dipropionate lacks efficacy against Theileria haneyi and fails to consistently clear Theileria equi in horses co-infected with T. haneyi. Pathogens. 2020;9:1035.
Bhoora R, Quan M, Matjila PT, Zweygarth E, Guthrie AJ, Collins NE. Sequence heterogeneity in the equi merozoite antigen gene (ema-1) of Theileria equi and development of an ema-1-specific TaqMan MGBTM assay for the detection of T. equi. Vet Parasitol. 2010;172:33–45.
Knowles DP, Perryman LE, Kappmeyer LS, Hennager SG. Detection of equine antibody to Babesia equi merozoite proteins by a monoclonal antibody-based competitive inhibition enzyme-linked immunosorbent assay. J Clin Microbiol. 1991;29:2056–8.
Kappmeyer LS, Perryman LE, Hines SA, Baszler TV, Katz JB, Hennager SG, et al. Detection of equine antibodies to Babesia caballi by recombinant B. caballi rhoptry-associated protein 1 in a competitive-inhibition enzyme-linked immunosorbent assay. J Clin Microbiol. 1999;37:2285–90.
Madden PA, Holbrook AA. Equine piroplasmosis: indirect fluorescent antibody test for Babesia caballi. Am J Vet Res. 1968;29:117–23.
World Organization for Animal Halth (WOAH)—terrestrial manual. 2021. https://www.woah.org/en/disease/equine-piroplasmosis/. Accessed 27 May 2023.
Manna G, Cersini A, Nardini R, Bartolom’e Del Pino LE, Antognetti V, Zini M, et al. Genetic diversity of Theileria equi and Babesia caballi infecting horses of central-southern Italy and preliminary results of its correlation with clinical and serological status. Ticks Tick Borne Dis. 2018;9:1212–20.
Tirosh-Levy S, Mazuz ML, Savitsky I, Pinkas D, Gottlieb Y, Steinman A. A serological and molecular prevalence of Babesia caballi in apparently healthy horses in Israel. Pathogens. 2021;10:445.
Kim CM, Blanco LB, Alhassan A, Iseki H, Yokoyama N, Xuan X, et al. Diagnostic real-time PCR assay for the quantitative detection of Theileria equi from equine blood samples. Vet Parasitol. 2008;151:158–63.
Coultous RM, McDonald M, Raftery AG, Shiels BR, Sutton DGM, Weir W. Analysis of Theileria equi diversity in The Gambia using a novel genotyping method. Transbound Emerg Dis. 2020;67:1213–21.
Bhoora RV, Collins NE, Schnittger L, Troskie C, Marumo R, Labuschagne K, et al. Molecular genotyping and epidemiology of equine piroplasmids in South Africa. Ticks Tick Borne Dis. 2020;11:101358.
Ahedor B, Kothalawala H, Kanagaratnam R, Vimalakumar SC, Otgonsuren D, Tuvshintulga B, et al. First detection of Theileria equi in free-roaming donkeys (Equus africanus asinus) in Sri Lanka. Infect Genet Evol. 2022;99:105244.
Chen K, Hu Z, Yang G, Guo W, Qi T, Liu D, et al. Development of a duplex real-time PCR assay for simultaneous detection and differentiation of Theileria equi and Babesia caballi. Transbound Emerg Dis. 2022;69:1338–49.
Chandler DP, Fredrickson JK, Brockman FJ. Effect of PCR template concentration on the composition and distribution of total community 16S rDNA clone libraries. Mol Ecol. 1997;6:475–82.
Vestheim H, Jarman SN. Blocking primers to enhance PCR amplification of rare sequences in mixed samples: a case study on prey DNA in Antarctic krill stomachs. Front Zool. 2008;5:12.
Yokoyama N, Ueno A, Mizuno D, Kuboki N, Khukhuu A, Igarashi I, et al. Genotypic diversity of Theileria orientalis detected from cattle grazing in Kumamoto and Okinawa prefectures of Japan. J Vet Med Sci. 2011;73:305–12.
Liyanagunawardena N, Sivakumar T, Kothalawala H, Silva SS, Battsetseg B, Lan DT, et al. Type-specific PCR assays for Babesia bovis msa-1 genotypes in Asia: revisiting the genetic diversity in Sri Lanka, Mongolia, and Vietnam. Infect Genet Evol. 2016;37:64–9.
Corpet F. Multiple sequence alignment with hierarchical clustering. Nucleic Acids Res. 1988;16:10881–90.
Leaché AD, Oaks JR. The utility of single nucleotide polymorphism (SNP) data in phylogenetics. Annu Rev Ecol Evol Syst. 2017;48:69–84.
Avarzed A, Igarashi I, De Waal DT, Kawai S, Oomori Y, Inoue N, et al. Monoclonal antibody against Babesia equi: characterization and potential application of antigen for serodiagnosis. J Clin Microbiol. 1998;36:1835–9.
Matjila PT, Leisewitz AL, Oosthuizen MC, Jongejan F, Penzhorn BL. Detection of a Theileria species in dogs in South Africa. Vet Parasitol. 2008;157:34–40.
Oosthuizen MC, Zweygarth E, Collins NE, Troskie M, Penzhorn BL. Identification of a novel Babesia sp. from a sable antelope (Hippotragus niger Harris, 1838). J Clin Microbiol. 2008;46:2247–51.
Ahedor B, Sivakumar T, Valinotti MFR, Otgonsuren D, Yokoyama N, Acosta TJ. PCR detection of Theileria equi and Babesia caballi in apparently healthy horses in Paraguay. Vet Parasitol Reg Stud Reports. 2023;39:100835.
Katoh K, Rozewicki J, Yamada KD. MAFFT online service: multiple sequence alignment, interactive sequence choice and visualization. Brief Bioinform. 2019;20:1160–6.
Kumar S, Stecher G, Li M, Knyaz C, Tamura K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol Biol and Evol. 2018;35:1547–9.
Tamura K, Nei M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol Biol Evol. 1993;10:512–26.
Alhassan A, Pumidonming W, Okamura M, Hirata H, Battsetseg B, Fujisaki K, et al. Development of a single-round and multiplex PCR method for the simultaneous detection of Babesia caballi and Babesia equi in horse blood. Vet Parasitol. 2005;129:43–9.
Woolhouse ME, Thumbi SM, Jennings A, Chase-Topping M, Callaby R, Kiara H, et al. Co-infections determine patterns of mortality in a population exposed to parasite infection. Sci Adv. 2015;1:e1400026.
Onyiche TE, Suganuma K, Igarashi I, Yokoyama N, Xuan X, Thekisoe O. A Review on equine piroplasmosis: epidemiology, vector ecology, risk factors, host immunity, diagnosis and control. Int J Environ Res Public Health. 2019;16:1736.
Scoles GA, Ueti MW. Vector ecology of equine piroplasmosis. Annu Rev Entomol. 2015;60:561–80.
Acknowledgements
We thank the veterinary staff in Sri Lanka and Paraguay for their support in sampling and DNA extraction. We thank Ms. Hiroko Yamamoto of the National Research Center for Protozoan Diseases, Obihiro University of Agriculture and Veterinary Medicine, Japan, for technical support.
Funding
This study was supported by grants from the Japan Society for the Promotion of Science (JSPS KAKENHI nos. 22H02511 and 22KK0095) and the Open Partnership Joint Project of the JSPS Bilateral Joint Research Projects (PJSBP120229902).
Author information
Authors and Affiliations
Contributions
BA: conceptualization, methodology, formal analysis, investigation, writing—original draft. DO: investigation. AZ: investigation. AG: investigation. NMMN: investigation. MFRV: investigation. HK: investigation, project administration. NK: investigation. SSPS: investigation. HK: investigation, project administration. TJA: conceptualization, funding acquisition, project administration. TS: conceptualization, methodology, formal analysis, writing—review and editing, supervision. NY: conceptualization, writing—review and editing, funding acquisition, supervision, project administration. All of the authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The Animal Care and Use Committee of Obihiro University of Agriculture and Veterinary Medicine, Japan, approved all of the animal procedures (approval nos. 29-53 and 23-24). All of the experiments were carried out in accordance with the Fundamental Guidelines for Proper Conduct of Animal Experiments and Related Activities in Academic Research Institutions under the Ministry of Education, Culture, Sports, Science and Technology, Japan.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Table S1.
The distribution of Theileria equi genotypes among Sri Lankan donkeys investigated in this study.
Additional file 2: Table S2.
The distribution of Theileria equi genotypes among Paraguayan horses investigated in this study.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Ahedor, B., Otgonsuren, D., Zhyldyz, A. et al. Development and evaluation of specific polymerase chain reaction assays for detecting Theileria equi genotypes. Parasites Vectors 16, 435 (2023). https://doi.org/10.1186/s13071-023-06045-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-023-06045-z