
Abstract
The Brown-capped Rosy-Finch is an endangered high alpine specialist that breeds on cliffs in the Rocky Mountains of North America. We know little about the parasites that infect these birds but filling this knowledge gap is important for understanding their population decline. The aim of this study was to survey haemosporidian parasite infections in Brown-capped Rosy-Finches. We sampled 104 Brown-capped Rosy-Finches during their breeding season at six sampling sites spanning the Colorado Rocky Mountains where they are nearly endemic. We used nested PCR methods to screen birds for parasite infections, and Sanger sequencing data were used to identify parasite lineages. Four of the sampling sites had birds with infections. Females were more often infected than males (prevalence = 15.6% and 9.7%, respectively). We observed twice as many infected birds sampled in July compared to birds sampled in June (prevalence = 12.5% and 6.25%, respectively). The percent of infected birds by sampling site ranged from 0 to over 20%. In total we identified eight different genetic lineages of haemosporidian parasites infecting Brown-capped Rosy-Finches—seven were in the genus Leucocytozoon, and one was in the genus Haemoproteus. Network analysis clearly separates Haemoproteus from Leucocytozoon, with Leucocytozoon lineages comprising two major clusters. Based on reports made to the MalAvi database, all lineages in Cluster 1 have the same reported insect vector, Simulium silvestre. We report that Brown-capped Rosy-Finches experience infections with haemosporidian parasites and that the percentage of infected birds differed markedly between sampling sites. We hypothesize that vector ecology and associated variation in climate at sampling sites explain our observations.

Similar content being viewed by others
Background
Host natural history and ecology play an essential role in shaping their parasite communities. Blood parasites transmitted by vectors include members of the order Haemosporida and, in birds, can involve the genera Plasmodium, Leucocytozoon, and Haemoproteus [1]. These parasites have complex life cycles and spend a portion of their life history infecting vertebrate host blood cells [1]. Haemosporidian parasites are widespread, diverse, and can influence host fitness [2,3,4,5]. For some avian hosts, such as long-distance migrants, exposure to blood-feeding insects and their associated haemosporidian parasites can occur at both wintering and breeding grounds [6, 7]. Conversely, for birds in temperate regions that are either non-migratory or are short-distance migrants, parasite transmission likely occurs during the breeding season, when insect vectors are active. Chronic infections can be retained across years [5], but for some haemosporidian parasites, infections are more commonly detected during the host breeding season [7, 8].
Brown-capped Rosy-Finches (Leucosticte australis) are Fringillidae songbirds and short-distance migrants that overwinter at lower elevations in the Southern Rocky Mountains where they are endemic [9]. During the summer, Brown-capped Rosy-Finches occur in extreme environments—they are among the highest altitudinal breeders in North America and nest on cliffs above treeline [9]. The Brown-capped Rosy-Finch is considered an endangered species whose population has declined by 95% since 1970 [10]. While climate-mediated habitat changes are suspected to be the primary cause of Brown-capped Rosy-Finch population decline, explicit investigations of other factors that might influence their conservation are nearly absent [11].
We know very little about the parasites that infect Brown-capped Rosy-Finches and the factors that shape their distributions and infection prevalence. To our knowledge only a single previous 1970 study investigated infections with vector-mediated blood parasites [12]. Given this existing knowledge gap, and the potential conservation implications, we sampled Brown-capped Rosy-Finches from throughout the Colorado Rocky Mountains for haemosporidian parasites. Sampling took place during the breeding season (June–July). Here we (i) report observed haemosporidian parasite prevalence by host sex, month, and site, (ii) identify the parasite lineages found, and (iii) report the occurrence of these lineages in other avian hosts as well as insect vectors using data from the MalAvi database [13].
Methods
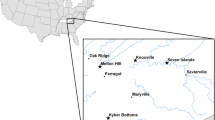
During June and July 2018, we captured 104 Brown-capped Rosy-Finches from nine locations across six Colorado mountain ranges (Fig. 1, Table 1). We captured birds using Potter traps baited with black-oil sunflower seeds. We then banded each bird with a USGS metal band and collected blood from the brachial vein using a heparinized capillary tube and a 0.20-gauge needle prior to release. Blood samples were preserved in Queen’s lysis buffer. Capture sites were all above treeline and ranged in elevation from 3513 m (FLTO) up to 4148 m (RAMP). We conducted salt extractions to isolate and purify DNA from blood preserved in lysis buffer [14]. To screen birds for infection we used nested PCR reactions to amplify a region of the cytb gene, and all birds were screened at least twice [15]. Nested PCRs were conducted in 25-μl reactions. To screen birds for Plasmodium and Haemoproteus, we used the initial primers HAEMNF and HAEMNR2 with nested primers HAEMF and HAEMR2 [16]. We screened birds for Leucocytozoon lineages using initial primers DW2 and DW4, followed by nested primers LeucoF and LeucoR [17]. All PCRs included both positive and negative controls. We checked amplifications using gel electrophoresis with SYBR safe stain and fluorescence occurring at the expected length of the amplified product. We then submitted positive testing PCR samples for Sanger sequencing by QuintaraBio (Hayward, CA) to confirm infections and identify parasite lineages. We note that none of the birds were screened for infection using microscopy to examine blood smears.

Sampling locations for Brown-capped Rosy-Finches (Leucosticte australis) are summarized by triangles with acronyms shown
We edited haemosporidian cytb sequences using Geneious prime v2020.1.2 [18]. First, we carried out a de novo assembly with the highest possible sensitivity for all sequences with forward and reverse primers for each bird in our study. We aligned the resulting sequence data to the MalAvi database [13] using the BLAST algorithm and identified parasite lineages. Birds were also screened for the presence of coinfections, which involved either infections with lineages in different genera (e.g. an infection with both Leucocytozoon and Haemoproteus parasites) or infections within the same genus (e.g. two different lineages in the genus Leucocytozoon). Some coinfections could be identified from Sanger sequencing data that resulted from the different sets of primers used. Other coinfections were identified based on the presence of mixed peaks that could be visualized in chromatograms. A mixed peak means a sequence might have two possible base pairs at the associated site. These coinfections were identified to lineage by duplicating the sequence and then assigning one of the possible base pairs to each sequence and re-aligning them to the MalAvi database. Importantly, all observed within-genus coinfections were lineages that only differed by a single base pair in their cytb sequences.
Prevalence is the proportion of infected individuals within a sampling population. We calculated prevalence partitioned by sampling site, sex, and month of sampling and estimated associated standard errors using R v4.1.2 [19]. To compare the genetic similarity of our observed haemosporidian lineages, and their distributions by sampling site, we conducted a network analysis using the program PopArt [20]. We applied the default settings for the Median-Joining method to calculate the network [21]. Importantly, while Median-Joining networks are not rooted, and therefore cannot show evolutionary directionality [22], they can be used to visualize reticulation events for closely related lineages [21].
Results
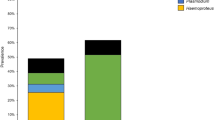
Twelve of the 104 Brown-capped Rosy-Finches that we screened for haemosporidian parasites were infected (prevalence = 11.5%, Additional file 1: Dataset S1). We identified 10 birds with Leucocytozoon infections (prevalence = 9.6%) and five birds with Haemoproteus infections (prevalence = 4.8%). We reported information on lineages we found infecting Brown-capped Rosy-Finches to the MalAvi database [13]. We did not identify any lineages that had not previously been described; therefore, we did not upload these sequences to GenBank. No birds had detectable infections with Plasmodium parasites. Coinfections involving both Leucocytozoon and Haemoproteus lineages were detected in three birds (prevalence = 2.9%), and two birds had coinfections with two Leucocytozoon lineages (prevalence = 1.9%). We observed more infections in female Brown-capped Rosy-Finches than males (prevalence = 15.6%, 5/32 infected; 9.7%, 7/72 infected, respectively), and a higher proportion of birds sampled in July were infected compared to birds sampled in June (prevalence = 12.5%, 11/88 infected; 6.25%, 1/16 infected, respectively) (Fig. 2). We did not observe infected birds at two of our sampling locations (RAMP and SACR). In contrast, over 15% of birds at NESU and FLTO were infected (Fig. 2).

Proportion of infected birds partitioned by sex A, sampling site B, and sampling month C with associated standard errors. The number of infected birds in proportion to total birds sampled for each category is shown above error bars
We identified eight different haemosporidian lineages that aligned 100% with lineages previously reported to the MalAvi database. Seven belonged to the genus Leucocytozoon, and a single lineage was in the genus Haemoproteus. All lineages beginning with an “L” are associated with Leucocytozoon. Similarly, “H” is used to distinguish the single Haemoproteus lineage (Table 2, Fig. 2). Our Median-Joining network analysis shows the Haemoproteus lineage H_PASILI01 is clearly diverged from all Leucocytozoon lineages (Fig. 3). Reticulation events based on cytb data show that Leucocytozoon lineages appear to cluster into two groups that we refer to as Clusters 1 and 2 (Fig. 3). Based on data reported to the MalAvi database (retrieved September 2022, summarized in Table 2), all lineages in Cluster 1 have previously been reported in Colorado and are vectored by the same black fly species, Simulium silvestre. None of the lineages in Cluster 2 have a previously reported vector but all have been described infecting birds in Western North America.

Network plot shows haemosporidian lineages by sampling site (indicated by color). Each hatch mark indicates a single base pair difference between lineages, and black circles show missing lineages. The Haemoproteus lineage “H_PASILI01” is observably divergent from Leucocytozoon lineages (designated with an “L”). Leucocytozoon lineages form two major clusters. All lineages in Cluster 1 have the same insect vector, Simulium silvestre. This plot was generated using the program PopArt [20]
Discussion
The extent to which parasite infections might influence Brown-capped Rosy-Finch survival and reproductive success is unknown. However, we report that these high elevation specialists do indeed experience infections with haemosporidian parasites. Based on molecular identification we found that 11.5% (12/104) of our sampling population was infected. Given variation in the success of primers and the potential for birds to harbor very low infection loads [23, 24], our reported prevalence data should be considered minimum estimates. We did not statistically compare prevalence based on sex, sampling month, and site because of the small sample size.
We did not observe infections with Plasmodium parasites, likely because insect vectors associated with Plasmodium are more common at lower elevations as has been suggested in other studies [25, 26]. Plasmodium parasites are known to be transmitted by mosquitoes (Diptera: Culicidae) typically in the genera Culex, Culiseta, and Aedes [1]. Surveys including a common vector for avian Plasmodium parasites, Culex pipiens, have reported that the vector does not occur above 1700 m in elevation along the Colorado Front Range [27, 28]. In contrast, black flies (Diptera: Simuliidae) are the known vector for Leucocytozoon parasites. We might expect these infections to be common at high elevations, where Brown-capped Rosy-Finches occur, given that vector species such as Simulium silvestre are known to breed in small fast-flowing streams that are common in the alpine area [29]. In songbirds, Haemoproteus infections are reportedly vectored by biting midges of the genus Culicoides [1, 30]. There is a lack of literature on the diversity and distribution of Culicoides in Colorado, but, in general, these insects are known to breed in moist areas with high levels of organic material such as ponds [31]. Of the sites where we report Haemoproteus infections, all were in close proximity to bodies of water.
We observed higher infection prevalence in female Brown-capped Rosy-Finches compared to males. Although the underlying cause of this disparity could be our sampling size, further research on Brown-capped Rosy-Finch breeding behavior and physiology might provide insight. Infections were more common in July than in June. While this might be due to sampling sites that were visited in June having inherently fewer infected birds (SAWA, RAMP, SAJU, and SACR), this difference could also be due to infections manifesting later in the summer because of both host physiology and vector phenology. Conversely, prevalence differences that we report by site could also be influenced by the timing of when birds were sampled. Testing samples collected from birds at the same field sites spanning multiple time points would resolve this knowledge gap. Additionally, climate varies across the Colorado Rocky Mountains, and this can influence the presence of vectors. For example, based on high-resolution climate mapping from the PRISM climate group [32], between 1981–2010, RAMP, a site with prevalence = 0%, exhibited an average annual rainfall < 90 cm. We also note that NESU, our most northerly site and the second lowest in elevation, had the highest prevalence of 22%. Both precipitation and elevation potentially influence Brown-capped Rosy-Finch exposure to vectors and subsequent haemosporidian infections.
From searching the MalAvi database, both lineages L_CB1 and L_CATUST14 appear to be host generalists, infecting multiple avian families (Table 2) [13]. L_CB1 is widespread throughout North America whereas reports of L_CATUST14 are more restricted to Western North America. Other Leucocytozoon lineages also appear more restricted in range, with two lineages, L_COLBF03 and L_COLBF04, having only previously been reported to the MalAvi database infecting the insect vector Simulium silvestre in the Colorado Rocky Mountains. However, infections with these two lineages have been observed in 12-day-old Black-capped Chickadees (Poecile atricapillus) and Mountain Chickadee (Poecile gambeli) nestlings in Boulder County, Colorado [33]. All lineages in Cluster 1 have been associated with the same insect vector, Simulium silvestre, in a previous study conducted in the Colorado Rocky Mountains [34].
Results from our Median-Joining network analysis, combined with previous reports made to the Malavi database, show that four of the closely related lineages that infect Brown-capped Rosy-Finches (L_CB1, L_COLBF03, L_COLBF04, and L_COLBF25, i.e. Cluster 1), are all vectored by the same black fly species: Simulium silvestre. The distribution and abundance of this vector might therefore be an effective predictor of infections with multiple Leucocytozoon lineages in Brown-capped Rosy-Finches. However, it is possible that these same lineages are transmitted by additional vectors that have not yet been described. Vectors for all other haemosporidian lineages we observed infecting Brown-capped Rosy-Finches have not yet been reported to the MalAvi database.
Conclusions
Here we reported on a survey of Brown-capped Rosy-Finch haemosporidian parasites (Genera: Plasmodium, Haemoproteus, and Leucocytozoon). To our knowledge, no previous literature has identified Leucocytozoon or Haemoproteus infections in this high alpine specialist, and we documented both genera at relatively low prevalence. Half of the haemosporidian lineages we documented (4/8) were reported in the same insect vector, Simulium silvestre, in a previous study [34]. We did not examine blood smears using microscopy methods, which prevents us from being able to quantify infection intensities and verify that the reported haemosporidian parasites are completing their life cycles in Brown-capped Rosy-Finches. However, our report provides a baseline understanding of the blood parasite infections that this endangered high elevation specialist harbors. With ongoing climate warming we might expect the haemosporidian communities infecting Brown-capped Rosy-Finches to shift over time, given potential movement of vector ranges.
Availability of data and materials
Blood samples for this work are available at the Denver Museum of Nature and Science. See Additional file 1: Table S1 for Brown-capped Rosy-Finch infection status data. Haemosporidian lineages identified from this work are described on the MalAvi database.
References
Valkiunas G. Avian malaria parasites and other haemosporidia. Boca Raton, Florida: CRC Press; 2005.
Knowles SCL, Palinauskas V, Sheldon BC. Chronic malaria infections increase family inequalities and reduce parental fitness: experimental evidence from a wild bird population. J Evol Biol. 2010;23:557–69. https://doi.org/10.1111/j.1420-9101.2009.01920.x.
Clark NJ, Clegg SM, Lima MR. A review of global diversity in avian haemosporidians (Plasmodium and Haemoproteus: Haemosporida): new insights from molecular data. Int J Parasitol. 2014;44:329–38. https://doi.org/10.1016/j.ijpara.2014.01.004.
Romano A, Nodari R, Bandi C, Caprioli M, Costanzo A, Ambrosini R, et al. Haemosporidian parasites depress breeding success and plumage coloration in female barn swallows Hirundo rustica. J Avian Biol. 2019. https://doi.org/10.1111/jav.01889.
Asghar M, Hasselquist D, Hansson B, Zehtindjiev P, Westerdahl H, Bensch S. Hidden costs of infection: chronic malaria accelerates telomere degradation and senescence in wild birds. Science. 2015;347:436–8. https://doi.org/10.1126/science.1261121.
Ricklefs RE, Medeiros M, Ellis VA, Svensson-Coelho M, Blake JG, Loiselle BA, et al. Avian migration and the distribution of malaria parasites in new world passerine birds. J Biogeogr. 2017;44:1113–23. https://doi.org/10.1111/jbi.12928.
Hellgren O, Wood MJ, Waldenström J, Hasselquist D, Ottosson U, Stervander M, et al. Circannual variation in blood parasitism in a sub-Saharan migrant passerine bird, the garden warbler. J Evol Biol. 2013;26:1047–59. https://doi.org/10.1111/jeb.12129.
Ham-Dueñas JG, Chapa-Vargas L, Stracey CM, Huber-Sannwald E. Haemosporidian prevalence and parasitaemia in the Black-throated sparrow (Amphispiza bilineata) in central-Mexican dryland habitats. Parasitol Res. 2017;116:2527–37. https://doi.org/10.1007/s00436-017-5562-3.
Johnson RE, Hendricks P, Pattie DL, Hunter KB. Brown-capped Rosy-Finch (Leucosticte australis). In: Billerman SM, Keeney BK, Rodewald PG, Schulenberg TS, editors. Birds of the world. Cornell lab of ornithology; 2020. Available from: https://birdsoftheworld.org/bow/species/bcrfin/1.0/introduction.
Rosenberg KV, Blancher PJ, Stanton JC, Panjabi AO. Use of North American breeding bird survey data in avian conservation assessments. Condor Ornithol Appl. 2017;119:594–606.
DeSaix MG, George TL, Seglund AE, Spellman GM, Zavaleta ES, Ruegg KC. Forecasting climate change response in an alpine specialist songbird reveals the importance of considering novel climate. Divers Distrib. 2022;28:2239–54. https://doi.org/10.1111/ddi.13628.
Stabler RM, Kitzmiller NJ. Hematozoa from colorado birds. III. Passeriformes. J Parasitol 1970; 56:12. Available from: https://www.jstor.org/stable/3277443?origin=crossref.
Bensch S, Hellgren O, Pérez-Tris J. MalAvi: a public database of malaria parasites and related haemosporidians in avian hosts based on mito cytochrome b lineages. Mol Ecol Resour. 2009;9:1353–8. https://doi.org/10.1111/j.1755-0998.2009.02692.x.
Miller SA, Dykes DD, Polesky HF. A simple salting out procedure for extracting DNA from human nucleated cells. Nucl Acids Res. 1988;16:1215–1215. https://doi.org/10.1093/nar/16.3.1215.
Hellgren O, Waldenström J, Bensch S. A new PCR assay for simultaneous studies of Leucocytozoon, Plasmodium, and Haemoproteus from avian blood. J Parasitol. 2004;90:797–802. https://doi.org/10.1645/GE-184R1.
Waldenström J, Bensch S, Hasselquist D, Östman Ö. A new nested polymerase chain reaction method very efficient in detecting plasmodium and haemoproteus infections from avian blood. J Parasitol. 2004;90:191–4. https://doi.org/10.1645/GE-3221RN.
Sehgal RNM, Hull AC, Anderson NL, Valkiūnas G, Markovets MJ, Kawamura S, et al. Evidence for cryptic speciation of Leucocytozoon spp. (Haemosporida, Leucocytozoidae) in diurnal raptors. J Parasitol. 2006;92:375–9. https://doi.org/10.1645/GE-656R.1.
Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, Sturrock S, et al. Geneious basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics. 2012;28:1647–9. https://doi.org/10.1093/bioinformatics/bts199.
R Core Team. R: a language and environment for statistical computing. 2013; Vienna: R Foundation for Statistical Computing.
Leigh JW, Bryant D. Popart: full-feature software for haplotype network construction. Methods Ecol Evol. 2015;6:1110–6. https://doi.org/10.1111/2041-210X.12410.
Bandelt HJ, Forster P, Rohl A. Median-joining networks for inferring intraspecific phylogenies. Molecul Biol Evol. 1999;16:37–48. https://doi.org/10.1093/oxfordjournals.molbev.a026036.
Kong S, Sánchez-Pacheco SJ, Murphy RW. On the use of median-joining networks in evolutionary biology. Cladistics. 2016;32:691–9. https://doi.org/10.1111/cla.12147.
Valkiūnas G, Bensch S, Iezhova TA, Križanauskienė A, Hellgren O, Bolshakov CV. Nested cytochrome B polymerase chain reaction diagnostics underestimate mixed infections of avian blood haemosporidian parasites: microscopy is still essential. J Parasitol. 2006;92:418–22. https://doi.org/10.1645/GE-3547RN.1.
Bernotienė R, Palinauskas V, Iezhova T, Murauskaitė D, Valkiūnas G. Avian haemosporidian parasites (Haemosporida): a comparative analysis of different polymerase chain reaction assays in detection of mixed infections. Experiment Parasitol. 2016; 163: 31–7. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0014489416300091.
LaPointe DA, Atkinson CT, Samuel MD. Ecology and conservation biology of avian malaria: ecology of avian malaria. Ann N Y Acad Sci. 2012;1249:211–26. https://doi.org/10.1111/j.1749-6632.2011.06431.x.
Reis S, Melo M, Covas R, Doutrelant C, Pereira H, Lima R de, et al. Influence of land use and host species on parasite richness, prevalence and co-infection patterns. Int J Parasitol. 2021; 51:83–94. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0020751920302745.
Eisen L, Bolling BG, Blair CD, Beaty BJ, Moore CG. Mosquito species richness, composition, and abundance along habitat-climate-elevation gradients in the northern Colorado front range. J Med Entomol. 2008;45:12. https://doi.org/10.1093/jmedent/45.4.800.
Baker M. The altitudinal distribution of mosquito larvae in the Colorado front range. Trans Am Entomol Soc. 1961;87:231–46.
Adler PH, Curry DC, Wood MD. The black flies (Simuliidae) of North America. Ithaca, New York: Cornell University Press; 2004.
Bukauskaitė D, Žiegytė R, Palinauskas V, Iezhova TA, Dimitrov D, Ilgūnas M, et al. Biting midges (Culicoides, Diptera) transmit Haemoproteus parasites of owls: evidence from sporogony and molecular phylogeny. Parasites Vectors. 2015; 8:303. Available from: http://www.parasitesandvectors.com/content/8/1/303.
Mellor PS, Boorman J, Baylis M. Culicoides biting midges: their role as arbovirus vectors. Annu Rev Entomol. 2000;45:307–40. https://doi.org/10.1146/annurev.ento.45.1.307.
Daly C, Halbleib M, Smith JI, Gibson WP, Doggett MK, Taylor GH, et al. Physiographically sensitive mapping of climatological temperature and precipitation across the conterminous United States. Int J Climatol. 2008;28:2031–64. https://doi.org/10.1002/joc.1688.
Theodosopoulos A. Haemosporidian infection dynamics and immune gene variation in a population of hybridizing chickadees. Doctoral dissertation, University of Colorado at Boulder; 2022. Available from: https://scholar.colorado.edu/concern/graduate_thesis_or_dissertations/j67315255.
Murdock CC, Adler PH, Frank J, Perkins SL. Molecular analyses on host-seeking black flies (Diptera: Simuliidae) reveal a diverse assemblage of Leucocytozoon (Apicomplexa: Haemospororida) parasites in an alpine ecosystem. Parasites Vectors. 2015. Available from: http://www.parasitesandvectors.com/content/8/1/343.
Acknowledgements
We thank Joel Such and Luke George for their assistance with sampling all Brown-capped Rosy-Finches involved in this study. We thank Carolina Chagas, Paul Hendricks, and two anonymous reviewers for feedback on the manuscript. Thank you, Erik Funk, for providing helpful background information on Brown-capped Rosy-Finch natural history. We are grateful to Amanda Hund for providing training on DNA salt extractions. Thank you to Dustin Quandt for assistance with equipment that was essential for laboratoty work. Thank you, Liz Clayton Fuller, for the Brown-capped Rosy-Finch illustration that is shown in Fig. 1. Thank you, Tamara Emmenegger for the Haemoproteus and Leucocytozoon images.
Funding
Funding for this work was provided by a University of Colorado Beverly Sears Graduate Student Research Grant and by a generous donation from Jack Ferguson to the Denver Museum of Nature & Science.
Author information
Authors and Affiliations
Contributions
ANT and SAT conceived the study. GMS collected and prepared samples for the study. ANT conducted laboratory work for the study. ANT wrote the manuscript and both SAT and GMS contributed with revisions. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All birds were sampled under Federal Bird Banding Permit (#22415) and Scientific Collecting License (#18BD2631). All participants consented to this study.
Consent for publication
All authors consent to the publication of this manuscript.
Competing interests
The authors declare no competing interests related to this study.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Dataset S1.
Brown-capped Rosy-Finch capture data show individual birds by their USGS band number. For each bird, information on their sex, sampling location, the elevation of sampling (in meters), year and month of capture, infection status, and haemosporidian lineage(s) (if applicable).
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Theodosopoulos, A.N., Spellman, G.M. & Taylor, S.A. Survey of haemosporidian parasite infections in an endangered high alpine bird. Parasites Vectors 16, 67 (2023). https://doi.org/10.1186/s13071-023-05667-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-023-05667-7