
Abstract
Phlebotomine sand flies are proven or suspected vectors of several pathogens of importance, including leishmaniasis, bartonellosis and sand fly fevers. Although sand flies have a worldwide distribution, there has been limited research published on sand flies and sand fly-borne pathogens throughout the Greater Mekong Sub-region (GMS). This review followed the PRISMA guidelines to determine the biodiversity and presence of phlebotomine sand flies and their associated pathogens in the GMS, specifically Cambodia, Thailand, the Lao People’s Democratic Republic (Laos), Malaysia and Vietnam. A total of 1472 records were identified by searching electronic databases, scanning reference lists of articles and consulting experts in the field. After screening of title and abstracts, 178 records remained and were further screened for original data (n = 34), not having regional data (n = 14), duplication of data (n = 4), records not available (n = 4) and no language translation available (n = 2). A total of 120 studies were then included for full review, with 41 studies on sand fly-related disease in humans, 33 studies on sand fly-related disease in animals and 54 entomological studies focused on sand flies (5 papers contained data on > 1 category), with a majority of the overall data from Thailand. There were relatively few studies on each country, with the exception of Thailand, and the studies applied different methods to investigate sand flies and sand fly-borne diseases, impacting the ability to conduct meaningful meta-analysis. The findings suggest that leishmaniasis in humans and the presence of sand fly vectors have been reported across several GMS countries over the past 100 years, with local transmission in humans confirmed in Thailand and Vietnam. Additionally, local Mundinia species are likely transmitted by biting midges. Findings from this study provide a framework for future investigations to determine the geographic distribution and risk profiles of leishmaniasis and other associated sand fly-borne disease throughout the GMS. It is recommended that researchers expand surveillance efforts across the GMS, with an emphasis placed on entomological surveys, syndromic and asymptomatic monitoring in both humans and animals and molecular characterization of sand flies and sand fly-borne pathogens, particularly in the understudied countries of Cambodia, Vietnam and Laos.
Graphical abstract

Similar content being viewed by others


Background
Over the last century, there has been limited research published discussing sand flies (Diptera: Psychodidae, Phlebotominae) and sand fly-borne pathogens in the Greater Mekong Sub-region (GMS). Phlebotomine sand flies are medically-important biting insects, with more than 800 identified species, genera or subgenera, found in both temperate and tropical regions of the world [1]. They have a wide host range and are proven or suspected vectors of several pathogens of importance for both civilian and military personnel, including leishmaniasis, bartonellosis and sand fly fevers.
Leishmaniases are a group of diseases caused by the protozoan parasites belonging to the genus Leishmania. More than 20 Leishmania species are known to infect humans and are transmitted by the bite of infected female phlebotomine sand flies and some genera of biting midges [2,3,4,5]. There are three main types of leishmaniasis: (i) visceral leishmaniasis (VL), often known as “kala–azar” and the most serious form of the disease; (ii) cutaneous leishmaniasis (CL), the most common; and (iii) mucocutaneous leishmaniasis [4]. Leishmaniasis alone infects more than 12 million people globally, with 0.9–1.6 million new cases and 20–30,000 deaths each year [6]. In 2017, 94% of global VL cases were reported from seven countries: Brazil, Ethiopia, India, Kenya, Somalia, South Sudan and Sudan [7].
Bartonella is a genus of hemotropic, Gram-negative, aerobic bacilli bacteria with several species implicated in causing human and animal bartonellosis [8]. Infections can be widespread in domestic and wild animals, as the pathogens are capable of being transmitted by a variety of blood-feeding arthropods including sand flies, fleas, mites and ticks [8]. Bartonella baciliformis, also known as Oroyo Fever or Carrion’s disease in humans, is presumably transmitted by Lutzomyia spp. sand flies, particularly by L. verrucarum [9, 10], but limited information is available about the role of sand flies in transmission of other Bartonella species.
Diseases caused by sand fly-borne viruses remain neglected when compared to parasites such as leishmaniasis. Sand fly fevers, caused by Phlebovirus (Phenuiviridae), have a global distribution with four main serotypes: sand fly fever Sicilian virus (SFSV), sand fly fever Cyprus virus (SFCV), sand fly fever Naples virus (SFNV) and Toscana virus (TOSV) [11]. A more recently emerging virus from the Rhabdoviridae family, Chandipura virus (CHPV), has been recorded in the Indian subcontinent, Sri Lanka and Africa [12,13,14,15]. This virus causes encephalitis and is associated with a high fatality rate; there have been several outbreaks in the Indian subcontinent [16,17,18,19]. Like Bartonella infections, the overall disease burden and distribution of these sand fly-borne viruses remain poorly characterized in the GMS.
Historically, Southeast Asia has been considered to be leishmaniasis-free, with minimal emphasis placed on sand fly-borne disease and prevention. Recent detections in Thailand of autochthonous cases of a novel Leishmania pathogen in healthy individuals [20, 21], paired with detections of Leishmania DNA in vectors collected near human cases [22], have resulted in a renewed interest in sand fly research in the GMS. The purpose of this study was to compile and document what is known about sand flies and the diseases they carry in the GMS and provide recommendations for future research priorities in the region.
The objective of the review was: (i) to determine the biodiversity of phlebotomine sand flies and (ii) to determine the presence of the associated pathogens they carry in the GMS, specifically Cambodia, Thailand, the Lao People’s Democratic Republic (Laos), Malaysia and Vietnam.
Methods
Search strategy and eligibility criteria
This review follows Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines [23] and was performed between 1 March 2019 and 30 September 2019. All data were extracted by two independent researchers, with discrepancies resolved by consensus. The inclusion criterion was any study that reported primary data on the collection of sand flies and the presence of sand fly-borne illness (e.g. inclusive of detections in vector, humans and animals) in the GMS. Grey literature and unpublished data were included when identified, and no restrictions were set based on the language of the report or dates published. The exclusion criteria included any reviews or studies which report secondary data or did not contain primary data on sand fly collections or detections of sand fly-borne diseases.
Data sources
Studies were identified by searching electronic databases, scanning reference lists of articles and through consulting with experts in the field. No limits were applied for language, with translation services and software applied to studies published in languages other than English. The search was conducted in PubMed, EMBASE, Web of Science, LILACS, Global Health and the Cochrane Database of Systematic Reviews. The term Lao was added to make sure all references to Laos or Lao PDR were captured. The following search terms were applied to all databases: “Phlebotomine sand*” AND (Cambodia OR Thailand OR Lao OR Malaysia OR Vietnam OR Mekong); “Sand fl*” AND (Cambodia OR Thailand OR Lao OR Malaysia OR Vietnam OR Mekong); Sandfl* AND (Cambodia OR Thailand OR Lao OR Malaysia OR Vietnam OR Mekong); Leishmania AND (Cambodia OR Thailand OR Lao OR Malaysia OR Vietnam OR Mekong); bartonellosis AND (Cambodia OR Thailand OR Lao OR Malaysia OR Vietnam OR Mekong); “sand* fevers” AND (Cambodia OR Thailand OR Lao OR Malaysia OR Vietnam OR Mekong); “Rift Valley Fever” AND (Cambodia OR Thailand OR Lao OR Malaysia OR Vietnam OR Mekong); Phlebovirus AND (Cambodia OR Thailand OR Lao OR Malaysia OR Vietnam OR Mekong); Sergentomyia AND (Cambodia OR Thailand OR Lao OR Malaysia OR Vietnam OR Mekong); Phlebotomus AND (Cambodia OR Thailand OR Lao OR Malaysia OR Vietnam OR Mekong).
Study selection
For each search, titles and abstracts were imported into Mendeley reference manager software (Elsevier, Mendeley Ltd., Amsterdam, The Netherlands), duplicates were removed and titles and abstracts were screened. Full texts of potentially relevant records were retrieved and assessed for eligibility, utilizing interlibrary loan and attempting to contact authors directly when necessary to increase the total number of reviewed full text. Reference lists of all potentially eligible articles and reviews were also searched (Additional file 1: Table S1).
We developed data extraction sheets, pilot tested it on randomly selected included studies and refined accordingly (Additional file 2: Table S2). Data extracted for studies with entomology collections included: (i) species name of sand fly collected; (ii) density of sand flies collected; (iii) location collected; (iv) number of days of collection; (v) date collected; (vi) method of collection; (vii) sample size of sites; and (viii) any sand fly-borne pathogens detected. Data extracted for pathogen studies included: (i) type of sand fly pathogen detected; (ii) study design; (iii) target of study (e.g. sand fly/human/both); (iv) sample size; (v) location; (vi) date; and (vii) method used for detection of pathogens (Additional file 3: Table S3, Additional file 4: Table S4, Additional file 5: Table S5). One review author extracted data and a second author checked the extracted data. The primary outcomes were: (i) reported endemic species of sand flies; and (ii) the presence of sand fly-borne pathogens in the GMS.
The approach to synthesis included aggregating data at the study level and producing a quantitative and qualitative synthesis, with tables reporting all extracted data made using Excel (Microsoft Corp., Redmond, WA, USA). The qualitative synthesis attempted to aggregate information about each country, species of sand flies identified and detected pathogens. No statistical tests or meta-analysis were performed, with findings from this study providing primarily descriptive data to identify gaps in the literature. Disagreements were resolved by discussion between the two review authors; a third author resolved disagreements between the two review authors.
Risk of bias
The commonly used guidelines for risk of bias in scientific studies (such as Effective Practice and Organization of Care [24] and Grading of Recommendations Assessment, Development and Evaluation [GRADE] [25]) are often focused on clinical trials in humans, and are not as useful when trying to determine bias in entomological laboratory studies. Therefore, any cluster randomized trial studies utilized the Cochrane risk of bias, but for all other studies we used a modified version of the GRADE guidelines [26] which rank a study as either high quality (further research is very unlikely to change our confidence in the estimate) to moderate quality (further research is likely to have an important impact on our confidence in the estimate and may change the estimate), low quality (further research is very likely to have an important impact on our confidence in the estimate and is likely to change the estimate) and very low quality (we are very uncertain about the estimate).
Results
Search results
Search results, including inclusion and exclusion numbers, are illustrated in Fig. 1. Initially, 1472 records were identified through database searches, with 10 additional records identified through items that were shared internally and following the checking of references of included studies. After the screening of titles and abstracts, the remaining 178 records were assessed and reviewed in full, following which 58 articles were excluded. The reasons for exclusion were: (i) not having original data (n = 34); (ii) not having regional data (n = 14); (iii) duplication of data (n = 4); (iv) records not available (n = 4); and (v) no language translation available (n = 2). A total of 120 studies were then included in the review.

PRISMA diagram for a systematic review on Phlebotomine sand flies (Diptera: Psychodidae) and sand fly-borne pathogens in the Greater Mekong Sub-region
Study characteristics
Included studies were published between 1914 and 2019 (Fig. 2), with most studies (58%) being published since 2010. Twelve studies were written in French, one in Thai, one in Norwegian and the remaining 106 in English. The majority of studies focused on Thailand (65%), followed by Malaysia (17%), Vietnam (11%), Laos (5%) and Cambodia (2%) (Fig. 3). All included studies were related to one or more of the following three core topics (5 studies included more > 1 core topic):
-
Sand fly-borne disease in humans (41 studies) [20,21,22, 27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64]
-
Sand fly-borne disease in animals (33 studies) [22, 38, 47, 65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93]
-
Entomology studies on sand flies (54 studies) [22, 37, 38, 56, 94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111,112,113,114,115,116,117,118,119,120,121,122,123,124,125,126,127,128,129,130,131,132,133,134,135,136,137,138,139,140,141,142,143]

The number of sand fly and sand fly-borne disease articles published by year

The Number of sand fly and sand fly-borne disease articles published by country
Sand fly-borne disease in humans
References with human data were primarily case studies (n = 23) and cross-sectional surveys (n = 12), with the remaining seven studies being a mix of laboratory, retrospective and prospective studies (Additional file 1: Table S1). There were seven species of Bartonella and nine species of Leishmania reported (Table 1). The majority of included references were from Thailand (74%), with no pathogen studies reported in Cambodia or Vietnam. Older references reported unspecified sand fly-borne disease (e.g. “kala-azar,” Phlebotomous fever) but did not identify the actual pathogen causing the disease.
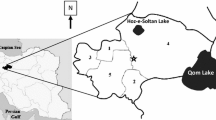
The first reports of leishmaniasis from Thailand were published by European physicians sent to combat plague in 1918 who noted the presence of both L. donovani and L. tropica, with the latter being found in more than 2% of patient ulcerations [53]. Despite this, recent papers in Thailand consider leishmaniasis to be an imported disease before 1999 [52]. From 1999 onwards, 22 laboratory-confirmed cases were reported (19 of which have been identified with molecular methods) [58], with Leishmania martiniquensis being the most common (15/22 cases) and the others associated with Leishmania infantum and L. orientalis [58]. The species Leishmania siamensis reported in earlier publications have been re-identified as L. martiniquensis using more comprehensive molecular methods, and since L. siamensis has never been formally described, the name is invalid and should not be used [21, 58]. Leishmania martiniquensis and L. orientalis were grouped together in Mundinia, a new subgenus recently introduced in 2016 [144, 145]. Members of this subgenus are presumably transmitted not only by sand flies but also by biting midges [2, 146]. Autochthonous symptomatic CL and VL cases have been reported sporadically in five southern, four northern, two central and one eastern province of Thailand [52, 55], and several additional asymptomatic cases among immunocompromised patients with human immunodeficiency virus/acquired immunodeficiency syndrome (HIV/AIDS) have occurred in nine provinces in southern Thailand [54] (Fig. 4).

Map of sand fly-borne pathogens in humans
Evidence for leishmaniasis transmission in other included GMS countries is lacking, with the majority of remaining references focusing on Malaysian immigrants [39, 44, 48, 50, 56]. A large cross-sectional study found high levels of Leishmania antibodies detected by RIDASCREEN® antibody enzyme-linked immunosorbent assay (ELISA) test kit in migrant workers from Myanmar (44%), Vietnam (26%) and Indonesia (26%) [56]. Two case reports of “kala–azar” were reported from Vietnam in 1931 and 1950 [49, 51]. However, no other studies on leishmaniasis were found from Vietnam, Laos or Myanmar throughout this review.
As previously mentioned, only B. bacilliformis has been linked to human transmission by sand flies and, as expected, there have been no reports of this bacterium in the GMS. However, other bartonelloses have been reported in the GMS, and many questions still remain regarding the maintenance and transmission of these pathogens. In 2000, Bartonella henselae was reported in 6.7% of healthy Thai blood donors, demonstrating the likelihood of frequent exposure across ages ranging from 20 to 70 years and in both sexes [41]. Subsequently, three novel Bartonella tamiae species were identified in 2002–2003 within a large prospective fever surveillance study in Chaing Rai and Khon Kaen, Thailand [34]. Findings indicated 7.7% positivity for Bartonella (B. elizabethae, B. rattimassiliensi and B. tribocorum, B. henselae, B. vinsonii and B. tamiae) by PCR among one study [31] and 12% positivity (B. henselae, B. quintana, B. elizabethae, B. vinsonii subsp. vinsonii) by immunofluorescence assay (IFA) in another study [28]. These results suggested that Bartonella is more common in rural Thailand than previously thought, although further evidence from an undifferentiated febrile illness study completed in 2017 in Nan Province only found 1% positivity by IFA. Additionally, a retrospective and subsequent prospective study in Laos, along the Thai border, found 7% positivity by IFA in patients admitted with definite or possible endocarditis [61]. These results, along with additional reports of Bartonella infections over various time points [30, 36, 62], suggest ongoing domestic transmission in humans within Thailand and likely Laos. Evidence for Bartonella transmission in the other GMS countries included in this review is limited, with only one case report of B. henselae and another case report of an unspecific Bartonella species reported from Malaysia [32, 40].
Sand fly-borne disease reported in animals
Animal studies have only recently been reported, starting in 2001, with the majority of articles included in this review focusing on human or entomological investigations dating back almost 100 years. Animal studies in GMS are sparse and focused on Leishmania, predominantly reported from Thailand. In Thailand, L. martiniquensis was isolated in rodents [22], L. dovovani complex was isolated from both dogs and cats [73, 92] and Leishmania spp. was isolated in dogs, cats and buffalo [92]. Leishmania donovani complex (L. infantum) was isolated in a singular dog in Vietnam [147] (Table 2).
Entomology studies on sand flies
Entomology studies on sand flies were largely broken up between identification keys (n = 26) and cross-sectional studies (n = 31). There were also two studies focused on laboratory methods from Thailand [98, 119] and two reports with unpublished primary data from Vietnam [104] and Laos [106]. The database of publications aggregated in this study indicate that most entomological activities regarding sand flies are focused on diversity rather than population control or insecticide resistance. However, one study in Thailand [111] did successfully establish a colony of wild-caught Phlebotomus stantoni and Phlebotomus major major, permitting further research on bionomic, vector competence and insecticide susceptibility status.
The sand flies found in the GMS are conservatively represented by three Old World genera: Phlebotomus, Sergentomyia, and Chinius [1]. Excluding taxonomic synonyms, 17 Phlebotomus species, including one unidentifiable species, 40 Sergentomyia species, including one unidentifiable species, and three Chinius species have been reported from the region (Table 3). Several studies documenting sand fly species diversity suggest that Sergentomyia gemmea is predominant in southern Thailand [103, 130]. However, recent molecular evidence raises concerns of misclassification for some of these S. gemmea specimens [136].
Species richness per country largely paralleled the number of entomology studies conducted (Thailand > Malaysia > Vietnam > Laos > Cambodia). Thailand and Malaysia recorded 34 and 32 species, respectively, compared to the number of species reported from Vietnam (n = 18), Cambodia (n = 10) and Laos (n = 8). Phlebotomus stantoni was the only species recorded in all countries, but presumably this is an artifact of the limited surveillance conducted in the region. For example, Sergentomyia barraudi was collected in every country but Malaysia. Similarly, Sergentomyia iyengari was reported everywhere except Cambodia; Laos was the only country yet to report Sergentomyia perturbans and Sergentomyia silvatica.
At the time of writing this report, Malaysia and Thailand were the only countries in the region that have reported Leishmania screening in their sand fly populations. Leishmania martiniquensis DNA has been detected in three Sergentomyia species from Thailand: S. barraudi, S. gemmea and S. iyengari [122]. Among these species, human DNA was been detected in the blood meal of S. gemmea and S. iyengari, suggesting that these species also feed upon humans. However, recent molecular evidence raises concerns of misclassification for some of these S. gemmea specimens [136]. In Malaysia, none of the sand flies collected in one study from 10 districts in Peninsular Malaysia tested positive for Leishmania DNA [94]. While the presence of both phlebotomine sand flies and the sand fly-borne pathogens have been reported in some Mekong countries, information remains sparse due to the absence of strong surveillance systems and limited laboratory resources for analysis.
Discussion
An increasing number of autochthonous VL cases have been reported in Thailand since 1999 [58], which in turn may have led to a disproportionate number of studies occurring in Thailand. The results of this review also suggest that bartonellosis does not appear to be a medically important sand fly-associated disease in the GMS region. However, the lack of knowledge on the hosts, vectors and transmission pathways of other Bartonella spp. in this region warrants further investigation.
The first reports of endemic human leishmaniasis in GMS occurring in Thailand came in 1918 [53], with additional reports in Vietnam as early as 1931 [51]. Despite these reports, leishmaniasis was considered to be an imported disease in Thailand until 1999 [52, 58]. Leishmaniasis in non-human animals was first reported in 2001, with most studies focused on rodents, dogs, cats and buffalos in Thailand. Older references reported “Kala Azar” or “Phlebotomous fever” but they did not specifically identify the pathogen. These findings suggest that leishmaniasis has been detected in several GMS countries over the past 100 years, with local transmission confirmed in Thailand and Vietnam. These findings pose a larger question about the burden and risk of leishmaniasis in the GMS and whether it is possibly re-emerging over the past decade, or if it has remained endemic and has simply been neglected from a research and surveillance perspective?
Entomology studies throughout Southeast Asia have detected leishmaniasis in both humans and animals in Southeast Asia, with the sand fly vectors of the disease identified regionally over the past 100 years. A concerted effort is needed to determine species richness, syndromic surveillance and serological monitoring in GMS countries other than Thailand, to fill in major gaps regarding sand fly-borne diseases in Southeast Asia [148, 149]. The absence of data regarding sand fly-borne diseases in humans may also limit targeted vector surveillance efforts; for example, a lack of leishmaniasis cases recorded in humans may result in entomology studies not being prioritized over more pressing disease vectors such as Anopheles and Aedes mosquitoes. There may be unreported sand fly species capable of vectoring Leishmania and Bartonella pathogens throughout the region that warrant updated investigations that utilize various modern molecular methods for species identification and pathogen detection. However, caution should be taken when using the detection of the pathogen’s DNA to suggest vectors are transmitting the pathogens to animals as the presence of pathogen DNA in an arthropod’s body does not provide evidence on the viability of the parasite or its ability to infect the host [146].
An important aspect of sand fly biology is their anthropophilic behavior. Most of the entomological studies reviewed were carried out using CDC light traps as a sole collection device. Studies in Thailand and Malaysia have included human landing collection as an additional collection method to better understand host preference of sand flies [38, 96, 106, 123]; similar studies are needed in Laos, Cambodia and Vietnam. A more recent effort by Siripattanapipong et al. [126] determined sand flies’ blood meal preferences using DNA sequencing technology, with the authors reporting the detection of human DNA in P. stantoni, S. barraudi, and S. iyengari blood meals. Further research is needed to see if host availability or certain abiotic factors affect blood meal preference in sand flies and how this impacts disease transmission risk in GMS.
Zoonotic disease transmissions can be exacerbated by large populations of freely roaming animals, increased pet ownership (e.g. cats, dogs) and the presence of agricultural animals living in close proximity to humans, as seen in many regions in Southeast Asia [147]. Throughout this sand fly review, there were very few manuscripts that addressed sand fly pathogens in animals, with most research coming from Thailand and focusing on leishmaniasis. Sand fly-associated pathogens, such as Phleboviruses (Salehabad virus and sand fly fevers) and CHPV occur throughout Asia and are addressed in many research papers examined during this review; however the roles animals play as reservoirs is not clear [150]. Research into sand fly-associated pathogens in animals throughout GMS could lead to an understanding of the transmission cycles and enable better integrated vector management techniques.
In the window of time between conducting this systematic review and publishing this manuscript, eight additional papers have been published related to sand flies and leishmaniasis in the GMS. One study on the prevalence and associated risk factors of Leishmania infection among immunocompetent hosts in Thailand focused on an outbreak investigation of an index case of CL caused by L. martiniquensis in 2015 and found that 7.1% of 392 participants were positive for Leishmania infection [151]. One participant tested positive with L. martiniquensis, while the rest were positive with either L. orientalis or Leishmania spp. The most statistically significant risk factor was having domestic animals in the housing area, indicating a sevenfold greater risk of infection. At the index patient’s school, seven S. gemma and one P. stantoni sand fly samples and one black rat were positive for Leishmania spp. However, the occurrence of these species in humans and animals, as well as in the sand fly species identified, have all been previously reported in other studies in Thailand. The additional seven papers included one report on the detection of Bartonella in Vietnamese rodents [152], one on Vietnamese sand fly collections [153] and five on Thai sand fly collections [154,155,156,157,158]. In these seven papers, three new species of sand fly species were reported, including Sergentomyia rudnicki [154] (2 samples) and Phlebotomus papatasi [157] (16 samples) in Thailand, and Sergentomyia khawi [153] (27 samples) in Vietnam. While not substantially changing the findings of this body of work, these additional studies reiterate the need to expand sand fly entomological surveillance and conduct research targeting leishmaniasis to areas outside of Thailand in the GMS region.
There were several limitations to this study, many of which are common in systematic reviews. Each country, with the exception of Thailand, had relatively few studies and applied different methods to investigate sand flies and sand fly-borne diseases, impacting our ability to conduct a meaningful meta-analysis. Another limitation was that many of the studies occurred several decades ago, when molecular methods had not been developed, leading to potential risks of vector misclassification and underestimation of disease burden. Furthermore, land use and deforestation in the GMS has drastically changed the environments where many of these vector studies originally occurred over 50 years ago, warranting a re-examination of sand fly-borne diseases and vector ecology for the greater region.
Conclusions
Findings from this study may help future investigations on the epidemiology of leishmaniases to determine the geographic distribution and risk profiles of leishmaniasis and other associated sand fly-borne disease throughout the GMS. Updated entomology keys are also needed within the GMS as the last regional sand fly key was published in the 1930s. It is recommended that researchers expand surveillance efforts across the GMS, with an emphasis placed on entomological surveys, syndromic and asymptomatic monitoring in both humans and animals and molecular characterization of sand flies and sand fly-borne pathogens, particularly in the understudied countries of Cambodia, Vietnam and Laos.
Availability of data and materials
All data generated or analyzed during this study are included in this published article and its Supplementary Information files.
Change history
17 November 2022
A Correction to this paper has been published: https://doi.org/10.1186/s13071-022-05550-x
Abbreviations
- CL:
-
Cutaneous leishmaniasis
- ELISA:
-
Enzyme-linked immunosorbent assay
- GMS:
-
Greater Mekong Sub-region
- HIV/AIDS:
-
Human immunodeficiency virus/acquired immunodeficiency syndrome
- IFA:
-
Immunofluorescent assay
- PRISMA:
-
Preferred Reporting Items for Systematic Reviews and Meta-Analyses
- VL:
-
Visceral leishmaniasis
References
Akhoundi M, Kuhls K, Cannet A, Votýpka J, Marty P, Delaunay P, et al. A Historical overview of the classification, evolution, and dispersion of Leishmania parasites and sandflies. PLoS Negl Trop Dis. 2016;10:e0004349.
Dougall AM, Alexander B, Holt DC, Harris T, Sultan AH, Bates PA, et al. Evidence incriminating midges (Diptera: Ceratopogonidae) as potential vectors of Leishmania in Australia. Int J Parasitol. 2011;41:571–9.
Chanmol W, Jariyapan N, Somboon P, Bates MD, Bates PA. Axenic amastigote cultivation and in vitro development of Leishmania orientalis. Parasitol Res. 2019;118:1885–97.
WHO. Leishmaniasis: situation and trends. 2019. https://www.who.int/gho/neglected_diseases/leishmaniasis/en/. Accessed 19 Sept 2019
Becvar T, Vojtkova B, Siriyasatien P, Votypka J, Modry D, Jahn P, et al. Experimental transmission of Leishmania (Mundinia) parasites by biting midges (Diptera: Ceratopogonidae). PLoS Pathog. 2021;17:e1009654.
WHO. Leishmaniasis. 2017. https://www.paho.org/hq/dmdocuments/2017/2017-cha-leishmaniasis-factsheet-work.pdf. Accessed 3 Sept 2019.
WHO. Leishmaniasis: epidemiological situation. 2019. https://www.who.int/leishmaniasis/burden/en/. Accessed 10 Sept 2019.
Jiyipong T, Jittapalapong S, Morand S, Rolain JM. Bartonella species in small mammals and their potential vectors in Asia. Asian Pac J Trop Biomed. 2014;4:757–67.
Noguchi H, Shannon RC, Tilden EB, Tyler JR. Etiology of oroya fever. XIV. The insect vectors of carrion’s disease. J Exp Med. 1929;49:993–1008.
Battisti JM, Lawyer PG, Minnick MF. Colonization of Lutzomyia verrucarum and Lutzomyia longipalpis sand flies (Diptera: Psychodidae) by Bartonella bacilliformis, the etiologic agent of Carrión’s disease. PLoS Negl Trop Dis. 2015;9:e0004128.
Tavana AM. Sandfly fever in the world. Ann Trop Med Public Healh. 2015;8:83–7.
Bhatt PN, Rodrigues FM. Chandipura: a new Arbovirus isolated in India from patients with febrile illness. Indian J Med Res. 1967;55:1295–305.
Peiris JSM, Dittus WPJ, Ratnayake CB. Seroepidemiology of dengue and other arboviruses in a natural population of toque macaques (Macaca sinica) at Polonnaruwa Sri Lanka. J Med Primatol. 1993;22:24–245.
Ba Y, Trouillet J, Thonnon J, Fontenille D. Phlebotomus of Senegal: survey of the fauna in the region of Kedougou. Isolation of arbovirus. Bull Soc Pathol Exot. 1999;92:131–5. (In French).
Traoré-Lamizana M, Fontenille D, Diallo M, Bâ Y, Zeller HG, Mondo M, et al. Arbovirus surveillance from 1990 to 1995 in the Barkedji area (Ferlo) of Senegal, a possible natural focus of Rift Valley fever virus. J Med Entomol. 2001;38:480–92.
Rao BL, Basu A, Wairagkar NS, Gore MM, Arankalle VA, Thakare JP, et al. A large outbreak of acute encephalitis with high fatality rate in children in Andhra Pradesh, India, in 2003, associated with Chandipura virus. Lancet. 2004;364:869–74.
Chadha MS, Arankalle VA, Jadi RS, Joshi MV, Thakare JP, Mahadev PVM, et al. An outbreak of Chandipura virus encephalitis in the eastern districts of Gujarat State. India Am J Trop Med Hyg. 2005;73:566–70.
Gurav YK, Tandale BV, Jadi RS, Gunjikar RS, Tikute SS, Jamgaonkar AV, et al. Chandipura virus encephalitis outbreak among children in Nagpur division, Maharashtra, 2007. Indian J Med Res. 2010;132:395–9.
Dwibedi B, Sabat J, Hazra RK, Kumar A, Dinesh DS, Kar SK. Chandipura virus infection causing encephalitis in a tribal population of Odisha in eastern India. Natl Med J India. 2015;28:185–7.
Osatakul S, Mungthin M, Siripattanapipong S, Hitakarun A, Kositnitikul R, Naaglor T, et al. Recurrences of visceral leishmaniasis caused by Leishmania siamensis after treatment with amphotericin B in a seronegative child. Am J Trop Med Hyg. 2014;90:40–2.
Pothirat T, Tantiworawit A, Chaiwarith R, Jariyapan N, Wannasan A, Siriyasatien P, et al. First isolation of Leishmania from Northern Thailand: case report, identification as Leishmania martiniquensis and phylogenetic position within the Leishmania enriettii complex. PLoS Negl Trop Dis. 2014;8:e3339.
Chusri S, Thammapalo S, Chusri S, Thammapalo S, Silpapojakul K, Siriyasatien P. Animal reservoirs and potential vectors of Leishmania siamensis in southern Thailand. Southeast Asian J Trop Med Public Health. 2014;45:13–9.
Liberati A, Altman DG, Tetzlaff J, Mulrow C, Gøtzsche PC, Ioannidis JPA, et al. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: explanation and elaboration. J Clin Epidemiol. 2009;339:2700.
Cochrane Effective Practice and Organisation of Care. EPOC Resources for review authors. 2016. https://epoc.cochrane.org/resources/epoc-resources-review-authors. Accessed 10 Sept 2019.
Grading of Recommendations Assessment, Development and Evaluation (GRADE). Development and Evaluation Working Group. 2019. http://www.gradeworkinggroup.org/. Accessed 4 Sept 2019.
Atkins D, Best D, Briss PA, Eccles M, Falck-Ytter Y, Flottorp S, et al. Grading quality of evidence and strength of recommendations. BMJ. 2004;328:1490.
Kattipathanapong P, Akaraphanth R, Krudsood S, Riganti M, Viriyavejakul P. The first reported case of autochthonous cutaneous leishmaniasis in Thailand. Southeast Asian J Trop Med Public Health. 2012;43:17–20.
Bhengsri S, Baggett HC, Peruski LF, Morway C, Bai Y, Fisk TL, et al. Bartonella seroprevalence in rural Thailand. Southeast Asian J Trop Med Public Health. 2011;42:687–92.
Pandey N, Siripattanapipong S, Leelayoova S, Manomat J, Mungthin M, Tan-Ariya P, et al. Detection of Leishmania DNA in saliva among patients with HIV/AIDS in Trang Province, southern Thailand. Acta Trop Netherlands. 2018;185:294–300.
Pachirat O, Kosoy M, Bai Y, Prathani S, Puapairoj A, Zeidner N, et al. The first reported case of Bartonella endocarditis in Thailand. Infect Dis Rep Italy. 2011;3:e9.
Kosoy M, Bai Y, Sheff K, Morway C, Baggett H, Maloney SA, et al. Identification of Bartonella infections in febrile human patients from Thailand and their potential animal reservoirs. Am J Trop Med Hyg. 2010;82:1140–5.
Lina LC, Rosalind S, Chong AW, Toha A, Shaffie B. Cat scratch disease: a diagnostic dilemma. Med J Malaysia. 2010;65:155–6.
Suankratay C, Suwanpimolkul G, Wilde H, Siriyasatien P. Autochthonous visceral leishmaniasis in a human immunodeficiency virus (HIV)-infected patient: the first in Thailand and review of the literature. Am J Trop Med Hyg. 2010;82:4–8.
Kosoy M, Morway C, Sheff KW, Bai Y, Colborn J, Chalcraft L, et al. Bartonella tamiae sp. nov., a newly recognized pathogen isolated from three human patients from Thailand. J Clin Microbiol. 2008;46:772–5.
Maharom P, Siripattanapipong S, Mungthin M, Naaglor T, Sukkawee R, Pudkorn R, et al. Visceral leishmaniasis caused by Leishmania infantum in Thailand. Southeast Asian J Trop Med Public Health. 2008;39:988–90.
Paitoonpong L, Chitsomkasem A, Chantrakooptungool S, Kanjanahareutai S, Tribuddharat C, Srifuengfung S. Bartonella henselae: first reported isolate in a human in Thailand. Southeast Asian J Trop Med Public Health. 2008;39:123–9.
Sukmee T, Siripattanapipong S, Mungthin M, Worapong J, Rangsin R, Samung Y, et al. A suspected new species of Leishmania, the causative agent of visceral leishmaniasis in a Thai patient. Int J Parasitol. 2008;6:617–22.
Kongkaew W, Siriarayaporn P, Leelayoova S, Supparatpinyo K, Areechokchai D, Duang-ngern P, et al. Autochthonous visceral leishmaniasis: a report of a second case in Thailand. Southeast Asian J Trop Med Public Health. 2007;38:8–12.
Abdul Halim A, Lokman Hakim S, Norashikin M, Yusri MY, Ahmad Dasuki AG, Chong CK, et al. Seroprevalence of leishmaniasis among Bangladeshi immigrant workers in Cameron Highlands. Malaysia Trop Biomed. 2001;18:143–4.
Tan CL, Fhun LC, Tai ELI, Gani NSA, Muhammed J, Jaafar TNT, et al. Clinical profile and visual outcome of ocular bartonellosis in Malaysia. J Trop Med. 2017;2017:7946123. https://www.hindawi.com/journals/jtm/2017/7946123/
Maruyama S, Boonmar S, Morita Y, Sakai T, Tanaka S, Yamaguchi F, et al. Seroprevalence of Bartonella henselae and Toxoplasma gondii among healthy individuals in Thailand. J Vet Med Sci. 2000;62:635–7.
Thisyakorn U, Jongwutiwes S, Vanichsetakul P, Lertsapcharoen P. Visceral leishmaniasis: the first indigenous case report in Thailand. Trans R Soc Trop Med Hyg. 1999;93:23–4.
Viriyavejakul P, Viravan C, Riganti M, Punpoowong B. Imported cutaneous leishmaniasis in Thailand. Southeast Asian J Trop Med Public Health. 1997;28:558–62.
Hamidah NH, Cheong SK, Hassan JB. A case of kala–azar diagnosed by bone marrow aspiration. Malays J Pathol. 1995;17:39–42.
Suttinont P, Thammanichanont C, Chantarakul N. Visceral leishmaniasis: a case report. Southeast Asian J Trop Med Public Health. 1987;18:103–6.
Picher O, Aspöck H. Häufigkeit und Bedeutung parasitärer Infektionen bei vietnamesischen Flüchtlingen. Wien Med Wochenschr. 1980;130:190–3. http://search.bvsalud.org/portal/resource/en/mdl-7395245
Takhampunya R, Korkusol A, Pongpichit C, Yodin K, Rungrojn A, Chanarat N, et al. Metagenomic approach to characterizing disease epidemiology in a disease-endemic environment in Northern Thailand. Front Microbiol. 2019;10:319.
Pond WL. Arthropod-borne virus antibodies in sera from residents of South-East Asia. Trans R Soc Trop Med Hyg. 1963;57:364–71.
Germain A, Marty J. Kala Azar treated by glucantime (RP2168) [Kala–azar Trait par le 2168 RP (Glucantime)]. Bulletin de la Société de Pathologie Exotique. 1950;43:240–2.
Pallister RA. A case of Kala–Azar. J Malayan Branch Br Med Assoc. 1939;2:250.
Roton M. Case of Infantile K.A. seen at Saigon.[Un cas de kala-azar Infantile Observe a Saigon.]. Bulletin de la Societe Medico-Chirurgicale de I'Indochine. 1931;9:511.
Leelayoova S, Siripattanapipong S, Manomat J, Piyaraj P, Tan-Ariya P, Bualert L, et al. Leishmaniasis in Thailand: a review of causative agents and situations. Am J Trop Med Hyg. 2017;96:534–42.
Mendelson RW. Tropical diseases observed in Siam. J Am Med Assoc. 1919;72:1199.
Manomat J, Leelayoova S, Bualert L, Tan-Ariya P, Siripattanapipong S, Mungthin M, et al. Prevalence and risk factors associated with Leishmania infection in Trang Province, Southern Thailand. PLoS Negl Trop Dis. 2017;11:e0006095.
Supsrisunjai C, Kootiratrakarn T, Puangpet P, Bunnag T, Chaowalit P, Wessagowit V. Disseminated autochthonous dermal leishmaniasis caused by Leishmania siamensis (PCM2 Trang) in a patient from Central Thailand infected with human immunodeficiency virus. Am J Trop Med Hyg. 2017;96:1160–3.
Azian MYN, Hakim SL, Khadri MS, Yusri MY, Adela JI, Noor M, et al. Leishmaniasis in peninsular Malaysia: the role of immigrant workers and the vector. Southeast Asian J Trop Med Public Health. 2016;47:607–16.
Siriyasatien P, Chusri S, Kraivichian K, Jariyapan N, Hortiwakul T, Silpapojakul K, et al. Early detection of novel Leishmania species DNA in the saliva of two HIV-infected patients. BMC Infect Dis. 2016;16:89.
Jariyapan N, Daroontum T, Jaiwong K, Chanmol W, Intakhan N, Sor-Suwan S, et al. Leishmania (Mundinia) orientalis n. sp. (Trypanosomatidae), a parasite from Thailand responsible for localised cutaneous leishmaniasis. Parasit Vectors. 2018;11:351.
Chiewchanvit S, Tovanabutra N, Jariyapan N, Bates MD, Mahanupab P, Chuamanochan M, et al. Chronic generalized fibrotic skin lesions from disseminated leishmaniasis caused by Leishmania martiniquensis in two patients from northern Thailand infected with HIV. Br J Dermatol. 2015;173:663–70.
Phumee A, Chusri S, Kraivichian K, Wititsuwannakul J, Hortiwakul T, Thavara U, et al. Multiple cutaneous nodules in an HIV-infected patient. PLoS Negl Trop Dis. 2014;8:e3291.
Rattanavong S, Fournier P-E, Chu V, Frichitthavong K, Kesone P, Mayxay M, et al. Bartonella henselae endocarditis in Laos—“the unsought will go undetected.” PLoS Negl Trop Dis. 2014;8:e3385.
Bai Y, Kosoy MY, Diaz MH, Winchell J, Baggett H, Maloney SA, et al. Bartonella vinsonii subsp. arupensis in humans Thailand. Emerg Infect Dis. 2012;18:989–91.
Bualert L, Charungkiattikul W, Thongsuksai P, Mungthin M, Siripattanapipong S, Khositnithikul R, et al. Autochthonous disseminated dermal and visceral leishmaniasis in an AIDS patient, southern Thailand, caused by Leishmania siamensis. Am J Trop Med Hyg. 2012;86:821–4.
Chusri S, Hortiwakul T, Silpapojakul K, Siriyasatien P. Consecutive cutaneous and visceral leishmaniasis manifestations involving a novel Leishmania species in two HIV patients in Thailand. Am J Trop Med Hyg. 2012;87:76–80.
KaiLing K, FuiXian K, Jaafar T, Quaza Nizamuddin HN, SunTee T. Prevalence and molecular heterogeneity of Bartonella bovis in cattle and Haemaphysalis bispinosa ticks in Peninsular Malaysia. BMC Vet Res. 2015;11:153.
Kim KS, Inoue K, Kabeya H, Sato S, Takada T, Pangjai D, et al. Prevalence and diversity of Bartonella species in wild small mammals in Asia. J Wildl Dis. 2016;52:10–21.
Klangthong K, Promsthaporn S, Leepitakrat S, Schuster AL, McCardle PW, Kosoy M, et al. The distribution and diversity of Bartonella species in rodents and their Ectoparasites across Thailand. PLoS ONE. 2015;10:e0140856.
Anh PH, Van Cuong N, Son NT, Tue NT, Kosoy M, Woolhouse MEJ, et al. Diversity of Bartonella spp. in bats Southern Vietnam. Emerg Infect Dis. 2015;21:1266–7.
Loan HK, Van CN, Takhampunya R, Klangthong K, Osikowicz L, Kiet BT, et al. Bartonella species and trombiculid mites of rats from the Mekong Delta of Vietnam. Vector Borne Zoonotic Dis. 2015;15:40–7.
Maruyama S, Sakai T, Morita Y, Tanaka S, Kabeya H, Boonmar S, et al. Prevalence of Bartonella species and 16s rRNA gene types of Bartonella henselae from domestic cats in Thailand. Am J Trop Med Hyg. 2001;65:783–7.
McKee CD, Kosoy MY, Bai Y, Osikowicz LM, Franka R, Gilbert AT, et al. Diversity and phylogenetic relationships among Bartonella strains from Thai bats. PLoS ONE. 2017;12:e0181696.
Mokhtar AS, Tay ST. Molecular detection of Rickettsia felis, Bartonella henselae, and B. clarridgeiae in fleas from domestic dogs and cats in Malaysia. Am J Trop Med Hyg. 2011;85:931–3.
Nimsuphan B, Chimnoi W, Kengradomkij C, Pangjai D, Jiyipong T, Khomkao J, et al. Detection of anti-Leishmania donovani complex antibodies of dogs and cats from Southern Thailand. KKU Vet J. 2014;24. http://vet.kku.ac.th/journal/2557_1/02-Burin.pdf
Pangjai D, Intachinda S, Maruyama S, Boonmar S, Kabeya H, Sato S, et al. Isolation and phylogenetic analysis of Bartonella species from Rusa deer (Rusa timorensis) in Thailand. Comp Immunol Microbiol Infect Dis. 2018;56:58–62.
Pangjai D, Maruyama S, Boonmar S, Kabeya H, Sato S, Nimsuphan B, et al. Prevalence of zoonotic Bartonella species among rodents and shrews in Thailand. Comp Immunol Microbiol Infect Dis. 2014;37:109–14.
Rosypal AC, Hailemariam S, Wekheye V, Huong LTT, Dubey JP, Lindsay DS, et al. Survey of dogs from Vietnam for antibodies to visceralizing Leishmania spp. J Parasitol. 2009;95:767.
Saisongkorh W, Wootta W, Sawanpanyalert P, Raoult D, Rolain JM. “Candidatus Bartonella thailandensis”: a new genotype of Bartonella identified from rodents. Vet Microbiol. 2009;139:197–201.
Assarasakorn S, Veir JK, Hawley JR, Brewer MM, Morris AK, Hill AE, et al. Prevalence of Bartonella species, hemoplasmas, and Rickettsia felis DNA in blood and fleas of cats in Bangkok. Thailand Res Vet Sci. 2012;93:1213–6.
Satranarakun P, Maruyama S, Kabeya H, Sato S, Jitapalapong S, Jitchum S, et al. Prevalence of Bartonella infection in well-cared cats in Bangkok metropolitan. Thai J Vet Med. 2016;46:555–60.
Tay ST, Mokhtar AS, Siti Nursheena MZ, Low KC. Isolation and molecular identification of Bartonellae from wild rats (Rattus species) in Malaysia. Am J Trop Med Hyg. 2014;90:1039–42.
Bai Y, Kosoy MY, Boonmar S, Sawatwong P, Sangmaneedet S, Peruski LF. Enrichment culture and molecular identification of diverse Bartonella species in stray dogs. Vet Microbiol. 2010;146:314–9.
Bai Y, Kosoy MY, Lerdthusnee K, Peruski LF, Richardson JH. Prevalence and genetic heterogeneity of Bartonella strains cultured from rodents from 17 provinces in Thailand. Am J Trop Med Hyg. 2009;81:811–6.
Bai Y, Malania L, Alvarez Castillo D, Moran D, Boonmar S, Chanlun A, et al. Global distribution of Bartonella infections in domestic bovine and characterization of Bartonella bovis strains using multi-locus sequence typing. PLoS ONE. 2013;8:e80894.
Billeter SA, Colton L, Sangmaneedet S, Suksawat F, Evans BP, Kosoy MY. Molecular detection and identification of Bartonella species in rat fleas from Northeastern Thailand. Am J Trop Med Hyg. 2013;89:462–5.
Angelakis E, Khamphoukeo K, Grice D, Newton PN, Roux V, Aplin K, et al. Molecular detection of Bartonella species in rodents from the Lao PDR. Clin Microbiol Infect. 2009;15:95–7.
Billeter SA, Sangmaneedet S, Kosakewich RC, Kosoy MY. Bartonella species in dogs and their Ectoparasites from Khon Kaen Province, Thailand. Southeast Asian J Trop Med Public Health. 2012;43:1186–92.
Brenner EC, Chomel BB, Singhasivanon O-U, Namekata DY, Kasten RW, Kass PH, et al. Bartonella infection in urban and rural dogs from the tropics: Brazil, Colombia, Sri Lanka and Vietnam. Epidemiol Infect. 2013;141:54–61.
Castle KT, Kosoy M, Lerdthusnee K, Phelan L, Bai Y, Gage KL, et al. Prevalence and diversity of Bartonella in rodents of northern Thailand: a comparison with Bartonella in rodents from southern China. Am J Trop Med Hyg. 2004;70:429–33.
Hassan UL, Dhaliwal GK, Watanabe M, Ong BL, Yakubu Y, Tay ST. Feline bartonellosis associated with some clinicopathological conditions in a veterinary hospital in Selangor Malaysia. Trop Biomed. 2017;34:174–9.
Inoue K, Maruyama S, Kabeya H, Kawanami K, Yanai K, Jitchum S, et al. Prevalence of Bartonella infection in cats and dogs in a metropolitan area Thailand. Epidemiol Infect. 2009;137:1568–73.
Jiyipong T, Morand S, Jittapalapong S, Rolain J-M. Bartonella spp. infections in rodents of Cambodia, Lao PDR, and Thailand: identifying risky habitats. Vector Borne Zoonotic Dis. 2015;15:48–55.
Junsiri W, Wongnarkpet S, Chimnoi W, Kengradomkij C, Kajeerum W, Pangjai D, et al. Seroprevalence of Leishmania infection in domestic animals in Songkhla and Satun provinces, southern Thailand. Trop Biomed. 2017;34:352–62.
Kabeya H, Colborn JM, Bai Y, Lerdthusnee K, Richardson JH, Maruyama S, et al. Detection of Bartonella tamiae DNA in Ectoparasites from rodents in Thailand and their sequence similarity with bacterial cultures from Thai patients. Vector Borne Zoonotic Dis. 2010;10:429–34.
Knudsen AB, Lewis DJ, Tesh RB, Rudnick A, Jeffery J, Singh I. Phlebotomine sand flies (Diptera: Psychodidae) from a primary hill forest in West Malaysia. J Med Entomol. 1979;15:286–91.
Leger N, Depaquit J, Gay F. Chinius eunicegalatiae n. sp. (Diptera; Psychodidae), a cavernicolous sandfly from Laos. Ann Trop Med Parasitol. 2010;104:595–600.
Apiwathnasorn C, Sucharit S, Surathin K, Deesin T. Anthropophilic and zoophilic phlebotomine sand flies (Diptera, Psychodidae) from Thailand. J Am Mosq Control Assoc. 1993;9:135–7.
Lewis DJ, Killick-Kendrick R. Some phlebotomid sand-flies and other diptera of Malaysia and Sri Lanka. Trans R Soc Trop Med Hyg. 1973;67:4–5.
Lewis DJ, Lane RP. A taxonomic review of Phlebotomus (Idiophlebotomus) (Psychodidae). Syst Entomol. 1976;1:53–60.
Lewis DJ, Wharton RH. Some Malayan sandflies (Diptera: Psychodidae). Proc R Entomol Soc London. 1963;32:117–24.
Loyer M, Depaquit J, Gay F. A new cavernicolous sand fly from Cambodia: Idiophlebotomus nicolegerae n. sp. (Diptera: Psychodidae). Acta Trop. 2016;155:43–50.
Muller F, Depaquit J, Leger N. Phlebotomus (Euphlebotomus) mascomai n. sp. (Diptera-Psychodidae). Parasitol Res. 2007;101:1597–602.
Newstead R. Notes on Phlebotomus, with descriptions of new species. Bull Entomol Res. 1914;5:179.
Panthawong A, Chareonviriyaphap T, Phasuk J. Species diversity and seasonality of phlebotomine sand flies (Diptera: Psychodidae) in Satun province, Thailand. Southeast Asian J Trop Med Public Health. 2015;46:857–65.
Parrot L, Clastrier J. Note sur les phlébotomes LXV. Arch Inst Pastor Algér. 1952;2:152–70.
Phumee A, Tawatsin A, Thavara U, Pengsakul T, Thammapalo S, Depaquit J, et al. Detection of an unknown Trypanosoma DNA in a Phlebotomus stantoni (Diptera: Psychodidae) collected from Southern Thailand and records of new sand flies with reinstatement of Sergentomyia hivernus Raynal & Gaschen, 1935 (Diptera: Psychodidae). J Med Entomol. 2017;54:429–34.
Apiwathnasorn C, Samung Y, Prummongkol S, Phayakaphon A, Panasopolkul C. Cavernicolous species of phlebotomine sand flies from Kanchanaburi Province, with an updated species list for Thailand. Southeast Asian J Trop Med Public Health. 2011;42:1405–9.
Polseela R, Apiwathnasorn C, Samung Y. Seasonal variation of cave-dwelling phlebotomine sandflies (Diptera: Psychodidae) in Phra Phothisat Cave, Saraburi Province, Thailand. Southeast Asian J Trop Med Public Health. 2007;38:1011–5.
Polseela R, Apiwathnasorn C, Samung Y. Seasonal distribution of phlebotomine sand flies (Diptera: Psychodidae) in Tham Phra Phothisat temple, Saraburi province. Thailand Trop Biomed. 2011;28:366–75.
Polseela R, Vitta A, Apiwathnasorn C. Distribution of phlebotomine sand flies (Diptera: Psychodidae) in limestone caves, Khao Pathawi, Uthai Thani province, Thailand. Southeast Asian J Trop Med Public Health. 2015;46:425–33.
Polseela R, Vitta A, Nateeworanart S, Apiwathnasorn C. Distribution of cave-dwelling phlebotomine sand flies and their nocturnal and diurnal activity in Phitsanulok Province, Thailand. Southeast Asian J Trop Med Public Health. 2011;42:1395–404.
Polseela R, Apiwathnasorn C. Preliminary observations on biology of a man-and cattlebiting Phlebotomus major major and a cave dwelling Phlebotomus stantoni under laboratory conditions. Trop Biomed. 2016;33:403–8.
Polseela R, Depaquit J, Apiwathnasorn C. Description of Sergentomyia phadangensis n. sp. (Diptera, Psychodidae) of Thailand. Parasites and Vectors. 2016;9:21.
Polseela R, Jaturas N, Thanwisai A, Sing K-W, Wilson J-J. Towards monitoring the sandflies (Diptera: Psychodidae) of Thailand: DNA barcoding the sandflies of Wihan Cave, Uttaradit. Mitochondrial DNA. 2016;27:3795–801.
Raynal J, Gaschen H. Sur les phlebotomes d’Indochine II Presence Phlebotomus bailyi, var campester au Tonkin. Bulletin de la Société de Pathologie Exotique. 1934;27:564.
Raynal J. Sur les phlebotomes d’Indochine III Phlebotomus stantoni, Newstead 1914, au Tonkin Sur la synonymie Phlebotomus maynei ?, Sint 1930 Phlebotomus stantoni? Bulletin de la Société de Pathologie Exotique. 1934;27:670.
Raynal J. Methode des precipitines Appl au contenu stomacal quelques phlebotomes du Tonkin. Bulletin de la Société de Pathologie Exotique. 1936;29:56
Causey OR. Phlebotomus of Siam with a description of a new variety. Am J Hyg. 1938;28:487.
Raynal J. Geographical Distribution of the Phlebotomus of Northern Indo-China, and certain aspects of their biology. Contrib a l’etude des phlebotomes d’Indochine III Distrib Geogr des phlebotomes l’Indochine Nord Quelques Asp leur Biol. Arch Inst Pasteur d’Indochine. 1936;6:349.
Raynal J. Sur les phlébotomes d’Indochine. VI. Présence de Phlebotomus sylvestris Sinton 1924 en Nord-Annam et au Tonkin. Bulletin de la Société de Pathologie Exotique. 1935;219.
Raynal J, Gaschen H. Sur les phlebotomes d’Indochine I Presence Phlebotomus barraudi, Sinton 1929, au Tonkin. Bulletin de la Société de Pathologie Exotique. 1934;27:559.
Raynal J, Gaschen H. The phlebotomids of Indochina. X. Phlebotomus morini n. sp. [description of male from Annam]. Sur les Phlebotomes d’Indochine VI Presence Phlebotomus sylvestris, Sint 1924, en Nord au Tonkin. Bulletin de la Société de Pathologie Exotique. 1935;28:219.
Rudnick A, Garcia R, Jeffery J, Marchette NJ, MacVean DW. The phlebotomine sandflies of Malaysia (Diptera: Psychodidae). Southeast Asian J Trop Med Public Health. 1971;2:86.
Shahar MK, Hassan AA, Lee HL, Salmah MRC. Studies of phlebotomine sand fly (Diptera: Psychodidae) populations in limestone areas and caves of western Malaysia. Southeast Asian J Trop Med Public Health. 2011;42:83–93.
Apiwathnasorn C, Sucharit S, Rongsriyam Y, Leemingsawat S, Kerdpibule V, Deesin T, et al. A brief survey of phlebotomine sandflies in Thailand. Southeast Asian J Trop Med Public Health. 1989;20:429–32.
Sinton JA. Phlebotomus stantoni Newstead, 1914 and some other Siamese sandflies. Indian J Med Res. 1931;19:99.
Siripattanapipong S, Leelayoova S, Ninsaeng U, Mungthin M. Detection of DNA of Leishmania siamensis in Sergentomyia (Neophlebotomus) iyengari (Diptera: Psychodidae) and molecular identification of blood meals of sand flies in an affected area, Southern Thailand. J Med Entomol. 2018;55:1277–83.
Sor-suwan S, Jariyapan N, Mano C, Apiwathnasorn C, Sriwichai P, Samung Y, et al. Species composition and population dynamics of phlebotomine sand flies in a Leishmania infected area of Chiang Mai. Thailand Trop Biomed. 2017;34:855–62.
Sukantamala J, Sing K-W, Jaturas N, Polseela R, Wilson J-J. Unexpected diversity of sandflies (Diptera: Psychodidae) in tourist caves in Northern Thailand. Mitochondrial DNA A DNA Mapp Seq Anal. 2017;28:949–55.
Chittsamart B, Samruayphol S, Sungvorayothin S, Pothiwat R, Samung Y, Apiwathnasorn C. Phlebotomine sand flies of edible-nest swiftlet cave of Lang Ga Jiew Island, Chumphon province, Thailand. Trop Biomed. 2015;32:402–6.
Sukra K, Kanjanopas K, Amsakul S, Rittaton V, Mungthin M, Leelayoova S. A survey of sandflies in the affected areas of leishmaniasis, Southern Thailand. Parasitol Res. 2013;112:297–302.
Sumruayphol S, Chittsamart B, Polseela R, Sriwichai P, Samung Y, Apiwathnasorn C, et al. Wing geometry of Phlebotomus stantoni and Sergentomyia hodgsoni from different geographical locations in Thailand. C R Biol. 2017;340:37–46.
Theodor O. On sandflies (Phlebotomus) from Ceylon, Siam and Malay. Indian J Med Res. 1938;26:261.
Choocherd S, Phasuk J. Nocturnal activity of phlebotomine sand flies in Satun province, Thailand. Southeast Asian J Trop Med Public Health. 2018;49:383–90.
Curler GR. Records of phlebotomine sand flies (Diptera, Psychodidae, Phlebotominae) with a description of a new species of Sergentomyia Franca & Parrot from Khao Yai National Park, Thailand. Zootaxa. 2011;2806:60–8.
Depaquit J, Muller F, Leger N. Phlebotomus (Euphlebotomus) barguesae n. sp. from Thailand (Diptera–Psychodidae). Parasit Vectors. 2009;2:5.
Depaquit J, Vongphayloth K, Siriyasatien P, Polseela R, Phumee A, Loyer M, et al. On the true identity of Sergentomyia gemmea and description of a closely related species: Se. raynali n. sp. Med Vet Entomol. 2019;33:521–9.
Depaquit J, Léger N, Beales P. Chinius barbazani n. sp. de Thailande (Diptera: Psychodidae). Parasite. 2006;13:151–8.
Lewis DJ. The phlebotomine sandflies (Diptera: Psychodidae) of the oriental region. Bull Br Mus nat Hist (Ent). 1978;37:217–343.
Vu NS, Tran SH, Tran PV, Tran TC, Tran DN, Dang AD, et al. Diversity and ecology of sand flies (Diptera: Psychodidae), potential vectors of Leishmania in the Quang Ninh Province, Vietnam. J Med Entomol. 2020;57:259-65. https://doi.org/10.1093/jme/tjz129.
Gaschen H, Nguyen-Nguyen. Presence phlebotomes en Indochine Sud. Masson. 1938;31:115.
Jaturas N, Vitta A, Samung Y, Apiwathnasorn C, Polseela R. Species composition and nocturnal activity of phlebotomine sand flies (Diptera: Psychodidae) inhabiting a limestone cave in Thailand. J Vector Ecol. 2018;43:52–8.
Kanjanopas K, Siripattanapipong S, Ninsaeng U, Hitakarun A, Jitkaew S, Kaewtaphaya P, et al. Sergentomyia (Neophlebotomus) gemmea, a potential vector of Leishmania siamensis in Southern Thailand. BMC Infect Dis. 2013;13:333.
Khadri MS, Depaquit J, Bargues MD, Ferte H, Mas-coma S, Lee HL, et al. First description of the male of Phlebotomus betisi Lewis and Wharton, 1963 (Diptera: Psychodidae). Parasitol Int. 2008;57:295–9.
Sereno D. Leishmania (Mundinia) spp.: from description to emergence as new human and animal Leishmania pathogens. New Microbes New Infect. 2019;30:100540.
Espinosa OA, Serrano MG, Camargo EP, Teixeira MMG, Shaw JJ. An appraisal of the taxonomy and nomenclature of trypanosomatids presently classified as Leishmania and Endotrypanum. Parasitology. 2018;145:430–42.
Seblova V, Sadlova J, Vojtkova B, Votypka J, Carpenter S, Bates PA, et al. The biting midge Culicoides sonorensis (Diptera: Ceratopogonidae) is capable of developing late stage infections of Leishmania enriettii. PLoS Negl Trop Dis. 2015;9:e0004060.
Colella V, Nguyen VL, Tan DY, Lu N, Fang F, Zhijuan Y, et al. Zoonotic vectorborne pathogens and ectoparasites of dogs and cats in eastern and Southeast Asia. Emerg Infect Dis. 2020;26:1221–33.
Cam E, Nichols JD, Sauer JR, Hines JE. On the estimation of species richness based on the accumulation of previously unrecorded species. Ecography. 2002;25:102–8.
Azovsky AI. Species-area and species-sampling effort relationships: disentangling the effects. Ecography. 2011;34:18–30.
Endy T. Viral febrile illnesses and emerging pathogens. In: Ryan ET, Hill DR, Endy TP, Solomon T, Aronson NE, editors. Hunter’s tropical medicine and emerging infectious diseases. Amsterdam: Elsevier; 2020. p. 325–50.
Sriwongpan P, Nedsuwan S, Manomat J, Charoensakulchai S, Lacharojana K, Sankwan J, et al. Prevalence and associated risk factors of Leishmania infection among immunocompetent hosts, a communitybased study in Chiang Rai, Thailand. PLoS Negl Trop Dis. 2021;15:e0009545.
Anh LTL, Balakirev AE, Van Chau N. Investigation of multiple infections with zoonotic pathogens of rodents in Northern Vietnam. J Vector Borne Dis. 2021;58:47–53.
Vu SN, Tran HS, Tran VP, Tran CT, Tran ND, Dang DA, et al. Taxonomical insights and ecology of sandfly (Diptera, Psychodidae) species in six provinces of Northern Vietnam. Parasite. 2021;28:85.
Phuphisut O, Nitatsukprasert C, Pathawong N, Jaichapor B, Pongsiri A, Adisakwattana P, et al. Sand fly identification and screening for Leishmania spp. in six provinces of Thailand. Parasit Vectors. 2021;14:352.
Somwang P, Khositharattanakool P, Pathawong N, Pongsiri A, Davidson SA, Ponlawat A. Field evaluation of four commercial light traps, trap placement, and effect of carbon dioxide on phlebotomine sand fly collection in northern Thailand. Acta Trop. 2021;220:e105953.
Thammapalo S, Pawestri AR, Kolaeh K, Boondej P, Benarlee R, Apiwathnasorn C, et al. Distribution of phlebotomine sandflies in the cave area of Satun Province, Thailand. Trop Med Infect Dis. 2020;5:174.
Phumee A, Wacharapluesadee S, Petcharat S, Siriyasatien P. A new cluster of rhabdovirus detected in field-caught sand flies (Diptera: Psychodidae: Phlebotominae) collected from southern Thailand. Parasit Vectors. 2021;14:569.
Buatong J, Dvorak V, Thepparat A, Thongkhao K, Koyadun S, Siriyasatien P, et al. Phlebotomine sand flies in Southern Thailand: entomological survey, identification of blood meals and molecular detection of Trypanosoma spp. Insects. 2022;13:197.
Acknowledgements
Not applicable
Funding
Funding for this manuscript was partially supported by the Armed Forces Health Surveillance Division (AFHSD)—Global Emerging Infections Surveillance (GEIS) Branch ProMIS ID #P0150_20_N2_08.01, work unit number D0016. The views expressed in this publication are those of the author and do not necessarily reflect the official policy or position of the Department of the Navy, Department of Defense, nor the US Government. JF and JCH are military Service members. This work was prepared as part of their official duties. Title 17, USC, §105 provides that copyright protection under this title is not available for any work of the US Government. Title 17, USC, §101 defines a US Government work as a work prepared by a military Service member or employee of the US Government as part of that person’s official duties.
Author information
Authors and Affiliations
Contributions
JCH conceived the study and substantially revised the manuscript. JH designed the study, performed the analysis and wrote the first draft of the manuscript. DP helped perform some of the analysis of the data, supported the interpretation of the data and substantially revised the manuscript. JF supported the analysis and interpretation of data and substantially revised the manuscript. MF substantially revised the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Ethics approval was not required for this study.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1:
Table S1. Bartonellosis reported in humans.
Additional file 2
: Table S2. Bartonellosis reported in animals.
Additional file 3
: Table S3. Data extracted from the entomology papers included in this review.
Additional file 4
: Table S4. Data extracted from the animal epidemiology papers included in this review.
Additional file 5
: Table S5. Data extracted from the human epidemiology papers included in this review.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Hustedt, J., Prasetyo, D.B., Fiorenzano, J.M. et al. Phlebotomine sand flies (Diptera: Psychodidae) and sand fly-borne pathogens in the Greater Mekong Subregion: a systematic review. Parasites Vectors 15, 355 (2022). https://doi.org/10.1186/s13071-022-05464-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-022-05464-8