
Abstract
Background
The tick Haemaphysalis longicornis exhibits two separate reproductive populations: bisexual and parthenogenetic, which have diploid and triploid karyotypes, respectively. The parthenogenetic population can undergo engorgement without copulation and produce viable female-only offspring with a longer incubation period than the bisexual population. Three enzymes, cathepsin B, cathepsin D and acid phosphatase, were found to be involved in vitellin degradation during the embryonic development of bisexual H. longicornis. However, the expression and activity profiles of these enzymes during the embryonic development of parthenogenetic ticks remain unknown. In the present study, the transcriptional expression profile, enzyme activity and roles in embryogenesis of the three enzymes during the embryonic development of parthenogenetic H. longicornis were investigated.
Methods
Quantitative real-time polymerase chain reaction (qPCR) and fluorescence detection were used to analyze the dynamic changes in the three enzymes during embryogenesis. The roles of the three enzymes during embryogenesis were also explored using RNA interference (RNAi).
Results
The three enzymes were all expressed during embryonic development in parthenogenetic H. longicornis. The expression of cathepsin B was highest on day 15, whereas that of cathepsin D was highest on day 3 and the peak of acid phosphatase expression occurred on day 9. The activity of cathepsin B was highest on day 3 and lowest on day 5, then gradually increased and remained stable. Cathepsin D activity was highest on day 1 and showed a gradually decreasing trend, whereas acid phosphatase showed the opposite trend and reached a peak on day 23. RNA interference experiments in engorged female ticks revealed that there was no significant difference in the number of eggs laid, but the hatching rate of the eggs was significantly decreased.
Conclusion
The three enzymes all play important roles in embryonic development of H. longicornis, but the expression patterns and changes in the activity of the enzymes in the bisexual and parthenogenetic populations are different. The results will help a better understanding of the similarities and differences underlying embryonic development in the bisexual and parthenogenetic populations and contribute to the future exploration of the development of the parthenogenetic population of H. longicornis.

Similar content being viewed by others

Background
Ticks are obligate blood-sucking ectoparasites and over 900 tick species have been reported around the world [1]. Ticks are the world’s second most common vector of disease after mosquitoes, exhibiting a wide range of hosts, including mammals, birds, reptiles and amphibians [2]. They can transmit a variety of pathogens, including viruses, bacteria, rickettsiae, helminths and protozoans [3]. In recent years, ticks have been shown to cause severe fever with thrombocytopenia syndrome [4, 5], human granulocytic anaplasmosis [6] and African swine fever [7], which affect human health and cause significant losses to the livestock industry.
Haemaphysalis longicornis is a three-host tick that is widely distributed in Australia, New Zealand, Korea, Japan and China [8]. Recently, it was found in Hunterdon County, New Jersey, USA for the first time [9]. It is an essential vector of zoonotic agents and can transmit severe fever with thrombocytopenia syndrome virus and Anaplasma, Babesia, Borrelia, Ehrlichia and Rickettsia bacteria [10]. Haemaphysalis longicornis exhibits two reproductive populations: bisexual and obligate parthenogenetic populations [11]. Comparison of the morphological characteristics of the two populations revealed that parthenogenetic individuals (except for engorged females) are significantly greater in weight than bisexual individuals, the genital apron of parthenogenetic females is wider than that of bisexual females and the Haller’s organ of parthenogenetic individuals has no seam hole structure [12, 13]. In addition, bisexual and parthenogenetic H. longicornis are diploid and triploid, respectively. Parthenogenetic H. longicornis can complete their life-cycle without fertilization, without any males being observed throughout the life-cycle and they show strict reproductive isolation from the bisexual population [11, 12]. Because of this unique mode of reproduction, the feeding period of parthenogenetic females is significantly shorter than that of bisexual females, possibly due to the mating behavior of the bisexual population. However, parthenogenetic individuals show a slightly longer development cycle than bisexual individuals, including nymphal premolting, preoviposition, oviposition and egg incubation periods. Although the egg weight of the parthenogenetic population is greater than that of the bisexual population, the hatching rate is lower [11, 12].
Embryonic development is an important physiological process in reproduction and development. Many proteolytic enzymes accumulate in oocytes together with vitellin and regulate vitellin degradation to provide nutrients during embryonic development [14]. This physiological process has been observed in many insects, such as Bombyx mori [15,16,17], Rhodnius prolixus [18, 19], Periplaneta americana [20], Blattella germanica [21], Culex quinquefasciatus [22], Musca domestica [23], Anticarsia gemmatalis [24], Dipetalogaster maxima [25], Spodoptera exigua [26] and Helicoverpa armigera [27]. However, related studies have only been reported in several tick species, including Ornithodoros moubata [28], Rhipicephalus microplus [14, 29,30,31,32,33] and bisexual H. longicornis [34]. Our previous study has proven that three enzymes, cathepsin B, cathepsin D and acid phosphatase, are involved in vitellin degradation during the embryonic development of bisexual H. longicornis [34]. However, the expression and activity profiles of these enzymes during the embryonic development of parthenogenetic ticks remain unknown. In the present study, the transcriptional expression profiles, enzyme activity and roles of the three enzymes during the embryonic development of parthenogenetic H. longicornis were investigated. The results will contribute to a better understanding of the similarities and differences underlying embryonic development between the bisexual and parthenogenetic populations and to the future exploration of the development of the parthenogenetic population of H. longicornis.
Methods
Tick collection and rearing
Parthenogenetic H. longicornis ticks were collected by flag dragging from Cangxi County (31°37ʹ–32°10″N, 105°43ʹ–106°28″E) in Sichuan Province, China. They were maintained for several generations in our laboratory and it was verified that no males were observed throughout the entire life-cycle to ensure that the ticks were obligate parthenogenetic. These ticks were reared on the ears of rabbits in the laboratory as described previously [35]. After finishing a blood meal, the engorged ticks were placed in clean, plastic Petri plates for oviposition. The plates were maintained under standard environmental chamber conditions (26 ± 1 °C, 75 ± 5% RH and a 8:16 h L:D photoperiod). The eggs were collected and separated after 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21 and 23 days and snap frozen using liquid nitrogen before being stored at − 80 °C for further use. All eggs used in this research originated from female ticks of the same batch.
Primer design and sequence analysis
Primers for quantitative analysis were designed using sequence information from the National Center for Biotechnology Information (NCBI). Because of the paucity of sequences from parthenogenetic H. longicornis, the sequences of several ticks were aligned and compared to search for conservative sequences upon which to design corresponding primers. For cathepsin B, sequences from H. longicornis (GenBank: AB255051), Ixodes ricinus (GenBank: EF428206) and Ixodes scapularis (GenBank: XM_002435418) were used. For cathepsin D, sequences from H. longicornis (GenBank: EU019715), I. ricinus (GenBank: HQ615697) and R. microplus (GenBank: FJ655904) were used. For acid phosphatase, sequences from H. longicornis (GenBank: HM150759), I. scapularis (GenBank: XM_002410276) and Anoplophora glabripennis (GenBank: XM_018710438) were used. The primers for the β-actin gene were designed using the sequence from H. longicornis (GenBank: AY254898). All primers were designed with Primer Premier 5 (Premier Biosoft) and had lengths of 18–22 bp and were free of dimers and hairpins (Table 1). PCR amplification was performed in a total volume of 10 μl, which included 5 μl of 2× Taq PCR StarMix (GenStar BioSolutions), 4 μl of H2O, 0.6 μl of cDNA template and 0.2 μl of each 10 μM primer (Table 1). The PCR conditions were as follows: initial denaturation at 94 °C for 3 min, followed by 35 cycles of 94 °C for 30 s, 60 °C for 30 s and 72 °C for 30 s, with a final extension step at 72 °C for 10 min. The PCR assays were performed in a ProFlex™ 3 × 32-well PCR System (Applied Biosystems). The PCR products were checked on a 1% agarose gel and positive amplicons were purified using an AxyPrepTM DNA Gel Extraction Kit (Axygen). The purified products were inserted into the pEASY®-T1 Simple Cloning Vector (TransGen), which was then transformed into Escherichia coli DH5α cells (Invitrogen). The transformed cells were submitted to Sangon (Sangon Biotech) and the resulting sequences were analyzed by BLASTn searches in the NCBI database to ensure that the primers were correct.
qPCR analysis
Total RNA was extracted from eggs in different developmental stages (80 mg each) using an AxyPrep™ Multisource Total RNA Miniprep Kit (Axygen). Then, total RNA from each sample was reverse transcribed to cDNA using TransScript® One-Step gDNA Removal and cDNA Synthesis SuperMix (TransGen Biotech) following the manufacturer’s protocol. qPCR was performed in an Mx3005P system (Agilent Technologies) using TransStart® Top Green qPCR SuperMix (TransGen Biotech) following the manufacturer’s instructions. The reaction system was as follows: 10 μl of 2× TransStart® Top Green qPCR SuperMix, 7.8 μl of H2O, 1 μl of cDNA template, 0.4 μl of Passive Reference Dye (TransGen Biotech) and 0.4 μl of each primer at 10 μM (Table 1). The parameters of the machine were set as described previously [34]. The results were normalized to β-actin and analyses of gene expression were performed using the 2−ΔΔCq method [36].
Enzyme activity assays
Total protein was extracted from eggs at different developmental stages (1.0 g each) by using 50 mM sodium acetate buffer (pH 5.0). Then, the homogenates were centrifuged at 12,000 × rpm for 10 min at 4 °C to obtain the supernatants. The bicinchoninic acid (BCA) method was used to adjust the protein concentration for consistency [37]. Cathepsin B and Cathepsin D Activity Fluorometric Assay Kits (Biovision, Milpitas, USA) were used to identify the activity of the enzymes following the manufacturer’s instructions. Cathepsin B can cleave the synthetic substrate RR-amino-4-trifluoromethyl coumarin (RR-AFC) to release free AFC, which can be easily quantified using a fluorometer or fluorescence plate reader at Ex/Em = 400/505 nm. Cathepsin D acts on the synthetic substrate GKPILFFRLK(Dnp)-D-R-NH2-MCA to release fluorescence, which can be easily quantified using a fluorometer or fluorescence plate reader at Ex/Em = 328/460 nm. An Acid Phosphatase Activity Colorimetric Assay Kit (Biovision) was used to identify the activity of acid phosphatase. The phosphatase substrate p-nitrophenyl phosphate (pNPP) was dephosphorylated by acid phosphatase so that it turned yellow and could then be detected at λmax of 405 nm.
RNAi
Primers for RNAi were designed in the same way as the qPCR primers. The TAA TAC GAC TCA CTA TAG G (T7) promoter sequence was added to the 5ʹ end of the primers (Table 2). Primer sequences for the control gene green fluorescent protein (GFP) from Tetraselmis subcordiformis (GenBank: KJ668651) were also used. The PCR products were gel purified to synthesize RNA by using the T7 RiboMAXTM Express RNAi System (Promega, Madison, USA) according to the manufacturer’s protocol. Double-stranded RNA (dsRNA) injection was performed as described previously [38]. Engorged female ticks were microinjected with 4 μg of the enzyme-targeted dsRNAs in a volume of 1 μl. Thirty ticks were used for each group. The engorged ticks were placed in clean, plastic Petri plates for oviposition. The plates were maintained under standard environmental chamber conditions (26 ± 1 °C, 75 ± 5% RH and a 8:16 h L:D photoperiod). The eggs were collected daily until the ticks no longer laid eggs and the number of eggs in each group was counted. In the cathepsin B group, eggs were collected at 11, 13, 15 and 17 days to confirm gene-specific silencing by qPCR [39]. A portion of the eggs were used to calculate the hatching rate. The same procedure was followed for the cathepsin D (3, 5, 7 and 9 days), acid phosphatase (9, 11, 13 and 15 days) and control (3, 5, 7, 9, 11, 13, 15 and 17 days) groups.
Statistical analysis
All experiments were repeated three times and the data were analyzed using ANOVA with SPSS 12.0 software (IBM). The t-test statistical analysis was performed using the SAS JMP statistical program 13.2 (SAS Institute Inc). The differences were considered statistically significant when P ≤ 0.05. Experimental values were obtained from three independent assays and expressed as the mean ± standard error.
Results
Cloning of the cathepsin B, cathepsin D and acid phosphatase genes for qPCR
Total RNA was extracted from eggs collected at 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21 and 23 days and then converted into cDNA. A portion of these three genes were amplified and the PCR results showed that the sizes of amplicons for the cathepsin B, cathepsin D and acid phosphatase genes were 133 bp, 88 bp and 180 bp, respectively (Fig. 1a–c). The deduced amino acid sequences of the three enzymes showed 100% similarity with those of bisexual H. longicornis.

qPCR amplification products analysis of cathepsin B (a), cathepsin D (b) and acid phosphatase (c) of the parthenogenetic H. longicornis
Expression and activity of cathepsin B during embryogenesis
To illustrate the functions of cathepsin B during embryogenesis, the transcriptional expression profile of cathepsin B was monitored via qPCR. The results indicated that cathepsin B was expressed throughout embryonic development (Fig. 2a). The expression of cathepsin B was low before day 11, then rapidly rose to its highest level on day 15 (F(11, 24) = 129.77, P ≤ 0.0001). However, the expression of cathepsin B decreased to a low level after day 15.

Dynamic changes of cathepsin B during embryonic development of the parthenogenetic H. longicornis. a Gene transcripts of cathepsin B. b Activity of cathepsin B. The levels of gene mRNA expression were normalised against the mRNA of β-actin and the error bars represent the mean ± SE values and the letters (a, b, c, d) labels represent significant differences (P ≤ 0.0001)
The analysis in which the fluorescent substrate was used to detect the activity of cathepsin B showed that its activity was high throughout embryonic development (Fig. 2b). On day 3, its activity reached the highest level (F(11, 24) = 14.98, P ≤ 0.0001), but it decreased to the lowest level on day 5 and increased to a stable level thereafter.
Expression and activity of cathepsin D during embryogenesis
The results showed that the expression of cathepsin D was detectable in the eggs from different stages of embryonic development (Fig. 3a). The transcriptional level was very low on the first day but increased rapidly to the highest level on day 3 (F(11, 24) = 33.59, P ≤ 0.0001). Thereafter, the expression level gradually declined. The activity of cathepsin D was highest on day 1 (F(11, 24) = 21.15, P ≤ 0.0001) and decreased on days 3 and 5 (Fig. 3b). Thereafter, its activity was relatively stable until day 21, but it fell sharply on day 23.

Dynamic changes of cathepsin D during embryonic development of parthenogenetic H. longicornis. a Gene transcripts of cathepsin D. b Activity of cathepsin D. The levels of gene mRNA expression were normalised against the mRNA of β-actin and the error bars represent the mean ± SE values and the letters (a, b, c, d) labels represent significant differences (P ≤ 0.0001)
Expression and activity of acid phosphatase during embryogenesis
On the first five days, the acid phosphatase gene showed very little expression (Fig. 4a). After day 5, its expression increased and was highest on day 9 (F(11, 24) = 50.14, P ≤ 0.0001). Thereafter, its expression dropped to a low level, where it remained until the end of embryogenesis. The activity trend of acid phosphatase showed a gradual increase until reaching the highest level on day 23 (Fig. 4b, F(11, 24) = 145.08, P ≤ 0.0001).

Dynamic changes of acid phosphatase during embryonic development of parthenogenetic H. longicornis. a Genes transcripts of acid phosphatase. b Activity of acid phosphatase. The levels of gene mRNA expression were normalised against the mRNA of β-actin and the error bars represent the mean ± SE values and the letters (a, b, c, d, e, f) labels represent significant differences (P ≤ 0.0001)
Gene cloning and assessment of gene silencing
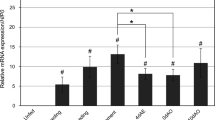
A portion of these three genes were amplified and the amplified fragment sizes of cathepsin B, cathepsin D and acid phosphatase were 430 bp, 497 bp and 539 bp, respectively (Fig. 5a–c). After microinjection, we collected the eggs of the experimental and control groups at different developmental stages to detect the expression of the enzyme-encoding genes by qPCR. The dsRNA-mediated knockdown of the transcripts of cathepsin B (F(3, 11) = 11.13, P = 0.0032), cathepsin D (F(3, 11) = 7.57, P = 0.0101) and acid phosphatase (F(3, 11) = 126.43, P ≤ 0.0001)resulted in a decrease in their relative levels compared to GFP (Fig. 6a–c). These results verified that the RNAi knockdown of enzyme-encoding genes for engorged female ticks was effective.

RNAi PCR amplification products analysis of cathepsin B (a), cathepsin D (b) and acid phosphatase (c) of parthenogenetic H. longicornis

Assessment of genes silencing efficiency of cathepsin B (a), cathepsin D (b) and acid phosphatase (c) of parthenogenetic H. longicornis. The levels of gene mRNA expression were normalised against the mRNA of β-actin and the error bars represent the mean ± SE values; asterisks indicate represent significant differences (P ≤ 0.05)
Effect of RNAi targeting enzyme-encoding genes on oviposition and hatching rates
The engorged ticks of the experimental and control groups were maintained in the incubator to lay eggs. The oviposition of the ticks and the hatching rate of the eggs were recorded. The weights of the eggs laid by the ticks in the cathepsin B, cathepsin D, acid phosphatase and GFP groups were 140 mg, 140 mg, 130 mg and 130 mg, respectively (Fig. 7a). There was no significant difference between the groups (F(3, 8) = 0.129, P = 0.940). However, the hatching rates of these groups were 49.33%, 52.17%, 66.40% and 84.87%, respectively (Fig. 7b). A significant difference was observed between the experimental and control groups (F(3, 8) = 22.127, P ≤ 0.0001).

DsRNA-mediated knockdown of transcripts for cathepsin B (CB), cathepsin D (CD) and acid phosphatase (AP) affect oviposition (a) and eggs’ hatchability (b) of parthenogenetic H. longicornis. The error bars represent the mean ± SE values and the letter (a, b, c) labels represent significant differences (P ≤ 0.0001)
Discussion
The main biochemical event in the embryonic development of arthropods is the utilization of yolk components [31]. As the major yolk protein, vitellin is the largest source of energy and nutrients during embryonic development and plays an irreplaceable role. Vitellin is still present in the larvae and is degraded in an orderly manner under the regulation of hydrolases [40]. The hydrolases stored in structures such as the modified lysosomes referred to as yolk granules are inactive proenzymes and they are activated mainly by developmentally controlled acidification [41]. These hydrolases can be classified into four categories: cysteine proteases (cathepsin B), serine proteases, aspartic proteases (cathepsin D) and acid phosphatases.
Cathepsin B is a cysteine protease found in the cytolysosomes that plays a vital role in the degradation of yolk during embryogenesis. In our study, the expression of cathepsin B was low on days 1–11 and rapidly increased to reach the highest level on day 15, then decreased sharply in the late development stage. However, in bisexual H. longicornis, cathepsin B showed a rapid rising trend on days 1–5, followed by a slow decline on later days [34]. The expression differences between the two populations might be due to the longer incubation of parthenogenetic eggs [12]. In other species, cathepsin B presents a high expression level at different developmental stages [42, 43]; therefore, in parthenogenetic H. longicornis, cathepsin B is more likely to be highly expressed in late embryonic development. The activity of cathepsin B fluctuated and was maintained at a high level throughout embryonic development in the parthenogenetic population, but in the bisexual population, it gradually decreased. Combined with the results obtained for cathepsin B expression, it can be concluded that cathepsin B played a vital role in late embryonic development in the parthenogenetic population, while in the bisexual population, cathepsin B tended to play a role in the early stage.
Cathepsin D is the major lysosomal aspartic protease and is widely distributed in the cells, where it regulates programmed cell death, autophagy and the degradation of yolk protein [18, 25]. Our study showed that the expression of cathepsin D rapidly reached the highest level on day 3 and then gradually decreased to a low level in the later development stage. The activity of cathepsin D was also highest in the early stage (on day 1) and was maintained at a high level during the later stage, except on day 23. However, in bisexual H. longicornis, the expression and activity of cathepsin D were highest on days 11 and 13, respectively [34]. In contrast to cathepsin B, in parthenogenetic H. longicornis, cathepsin D tended to play a role in the early stage, while in the bisexual population, cathepsin D was more likely to regulate embryogenesis in the late stage. Although both cathepsin B and D were indicated to regulate embryogenesis, their dynamic changes were not the same in the parthenogenetic and bisexual populations. In our previous work, we found that the vitellin of the parthenogenetic population exhibits 9 subunits, ranging from 31 kDa to 203 kDa; however, in the bisexual population, vitellin exhibits only 8 subunits, ranging from 52 kDa to 112 kDa [44, 45]. This might be another reason for the different dynamic changes in the enzymes, in addition to the longer incubation period.
In contrast to cathepsins B and cathepsins D, acid phosphatase is a classical lysosomal enzyme that catalyzes the hydrolysis of orthophosphoric monoesters [24]. The expression of the enzyme in the parthenogenetic and bisexual populations was highest on days 9 and 11, respectively. Its activity rose in the early stage (1–5 days) and was maintained at a high level on the following days in both populations [34]. The dynamic changes in acid phosphatase in the two populations were similar, possibly because the presence of acid phosphatase facilitates the degradation of yolk protein by the other two enzymes [18,19,20, 23, 25].
After RNAi treatment, the expression of the three targeted genes in the eggs was found to be significantly suppressed and the hatching rate was significantly reduced compared with that in the GFP group. However, the oviposition of the ticks showed no significant change. Similar results were previously found in bisexual H. longicornis [46, 47]. In Radopholus similis [48], Schistosoma mansoni [49, 50], Schistosoma japonicum [51] and Schmidtea mediterranea [52], knocking down the cathepsin B gene affects the development of the eggs and the hatching rate and even slows development and results in a shorter body length. In B. mori, cathepsin B participates in programmed cell death during metamorphosis and RNAi knockdown of cathepsin B leads to the stagnation of the larval-pupal metamorphosis [53]. The role of cathepsin D in the metamorphosis of S. exigua has also been reported and RNAi knockdown of cathepsin D reduces the survival rate in the fifth-instar [26]. A similar result was found in B. mori, in which deficiency of cathepsin D affects the pupation of larvae [54]. RNAi knockdown of cathepsin D in S. mansoni affects its growth and reproduction of polypides in mice, which indicates the important role of cathepsin D in S. mansoni development [55]. Through proteomic analysis of the acidocalcisomes of Trypanosoma brucei, researchers screened acid phosphatase and the functional analysis of this enzyme through RNAi showed that it was involved in growth and development [56]. These results indicated that cathepsin B, cathepsin D and acid phosphatase are involved in embryonic development and play essential roles in growth and reproduction.
The dynamic changes and functions of these three genes in embryonic development have been studied, but the enzyme-enzyme and enzyme-vitellin interactions are still not clear. The interaction of cathepsin B and acid phosphatase has been found in P. americana and M. domestica and the hydrolysis of vitellin is increased when both enzymes are present [20, 23]. In R. prolixus, acid phosphatase can hydrolyze polyphosphate (polyP) to abolish the inhibitory effect of polyP on cathepsin D, which could facilitate the degradation of vitellin by cathepsin D [18, 19]. The same interaction of the two enzymes is found in D. maxima [25]. In future work, we will investigate the relationships between the three enzymes and identify their effects on vitellin and embryogenesis to reveal the reasons for the differences between the two reproductive mechanisms.
Conclusions
The dynamic changes in cathepsin B, cathepsin D and acid phosphatase during the embryonic development of parthenogenetic H. longicornis have been identified. The three enzymes all play an important role in embryonic development, but the expression patterns and changes in the activity of the enzymes in the bisexual and parthenogenetic populations are different. After the knockdown of the genes encoding the enzymes, oviposition was not affected, whereas the hatching rate of eggs was significantly decreased. These above results will help a better understanding of the similarities and differences in embryonic development between the bisexual and parthenogenetic populations and will contribute to the future exploration of the development of the parthenogenetic population of H. longicornis.
Availability of data and materials
The data supporting the conclusions of this article are included within the article. Raw data used or analyzed during the present study are available from the corresponding author upon reasonable request.
Abbreviations
- RH:
-
relative humidity
- L:D:
-
light:dark photocycle
- PCR:
-
polymerase chain reaction
- qPCR:
-
quantitative real-time PCR
- BCA:
-
bicinchoninic acid
- AFC:
-
amino-4-trifluoromethyl coumarin
- pNPP:
-
p-nitrophenyl phosphate
- MCA:
-
4-methylcoumarin-7-amide
- T7:
-
TAA TAC GAC TCA CTA TAG G
- dsRNA:
-
double-stranded RNA
- RNAi:
-
RNA interference
- polyp:
-
polyphosphate
- SE:
-
standard error
References
Zhang YK, Zhang XY, Liu JZ. Ticks (Acari: Ixodoidea) in China: geographical distribution, host diversity, and specificity. Arch Insect Biochem Physiol. 2019;11:e21544.
Sonenshine DE, Roe RM. Biology of ticks. 2nd ed. New York: Oxford University Press; 1991.
Yu ZJ, Wang H, Wang TH, Sun WY, Yang XL, Liu JZ. Tick-borne pathogens and the vector potential of ticks in China. Parasites Vectors. 2015;8:24.
Yu XJ, Liang MF, Zhang SY, Liu Y, Li JD, Sun YL, et al. Fever with thrombocytopenia associated with a novel bunyavirus in China. N Engl J Med. 2011;364:1523–32.
Feldmann H. Truly emerging—a new disease caused by a novel virus. N Engl J Med. 2011;364:1561–3.
Uminski K, Kadkhoda K, Houston BL, Lopez A, MacKenzie LJ, Lindsay R, et al. Anaplasmosis: an emerging tick-borne disease of importance in Canada. IDCases. 2018;14:e00472.
Galindo I, Alonso C. African swine fever virus: a review. Viruses. 2017;9:E103.
Zheng HY, Yu ZJ, Zhou LF, Yang XL, Liu JZ. Seasonal abundance and activity of the hard tick Haemaphysalis longicornis (Acari: Ixodidae) in North China. Exp Appl Acarol. 2012;56:133–41.
Rainey T, Occi JL, Robbins RG, Egizi A. Discovery of Haemaphysalis longicornis (Ixodida: Ixodidae) parasitizing a sheep in New Jersey, United States. J Med Entomol. 2018;55:757–9.
Beard CB, Occi J, Bonilla DL, Egizi AM, Fonseca DM, Mertins JW, et al. Multistate infestation with the exotic disease-vector tick Haemaphysalis longicornis—United States, August 2017—September 2018. MMWR Morb Mortal Wkly Rep. 2018;67:1310–3.
Chen XJ, Xu SQ, Yu ZJ, Guo LD, Yang SJ, Liu LM, et al. Multiple lines of evidence on the genetic relatedness of the parthenogenetic and bisexual Haemaphysalis longicornis (Acari: Ixodidae). Infect Genet Evol. 2014;21:308–14.
Chen Z, Yang XJ, Bu FJ, Yang XL, Liu JZ. Morphological, biological and molecular characteristics of bisexual and parthenogenetic Haemaphysali longicornis. Vet Parasitol. 2012;189:344–52.
Wang D, Hu YH, Liu JZ. Ultrastructure and development of the Haller’ s organ of parthenogenetic Haemaphysalis longicornis (Acari: Ixodidae). Acta Entomol Sinica. 2013;56:306–11.
Estrela A, Seixas A, Termignoni C. A cysteine endopeptidase from tick (Rhipicephalus (Boophilus) microplus) larvae with vitellin digestion activity. Comp Biochem Phys B. 2007;148:410–6.
Yamahama Y, Uto N, Tamotsu S, Miyata T, Yamamoto Y, Watabe S, et al. In vivo activation of pro-form Bombyx cysteine protease (BCP) in silkmoth eggs: localization of yolk proteins and BCP, and acidification of yolk granules. J Insect Physiol. 2003;49:131–40.
Cai XY, Yu J, Yu HY, Liu YW, Fang Y, Ren ZX, et al. Core promoter regulates the expression of cathepsin B gene in the fat body of Bombyx mori. Gene. 2014;542:232–9.
Wang DD, Zhang Y, Dong ZM, Guo PC, Ma SY, Guo KY, et al. Serine protease P-IIc is responsible for the digestion of yolk proteins at the late stage of silkworm embryogenesis. Insect Biochem Mol Biol. 2016;74:42–9.
Fialho E, Nakamura A, Juliano L, Masuda H, Silvaneto MA. Cathepsin D-mediated yolk protein degradation is blocked by acid phosphatase inhibitors. Arch Biochem Biophys. 2005;436:246–53.
Gomes FM, Oliveira DM, Motta LS, Ramos IB, Miranda KM, Machado EA. Inorganic polyphosphate inhibits an aspartic protease-like activity in the eggs of Rhodnius prolixus (Stahl) and impairs yolk mobilization in vitro. J Cell Physiol. 2010;222:606–11.
Oliveira DM, Ramos IB, Reis FC, Lima AP, Machado EA. Interplay between acid phosphatase and cysteine proteases in mediating vitellin degradation during early embryogenesis of Periplaneta americana. J Insect Phys. 2008;54:883–91.
Purcell JP, Quinn TM, Kunkel JG, Nordin JH. Correlation of yolk phosphatase expression with the programmed proteolysis of vitellin in Blattella germanica during embryonic development. Arch Insect Biochem Physiol. 1988;9:237–50.
Moura AS, Cardoso AF, Costa da Silva AL, Winter CE, Bijovsky AT. Two cathepsins B are responsible for the yolk protein hydrolysis in Culex quinquefasciatus. PLoS ONE. 2015;10:e0118736.
Ribolla PE, Daffre S, Bianchi AGD. Cathepsin B and acid phosphatase activities during Musca domestica embryogenesis. Insect Biochem Mol Biol. 1993;23:217–23.
Oliveira DM, Gomes FM, Carvalho DB, Ramos I, Carneiro AB, Silvaneto MA, et al. Yolk hydrolases in the eggs of Anticarsia gemmatalis hubner (Lepidoptera: Noctuidae): a role for inorganic polyphosphate towards yolk mobilization. J Insect Physiol. 2013;59:1242–9.
Leyria J, Fruttero LL, Nazar M, Canavoso LE. The role of DmCatD, a cathepsin D-like peptidase, and acid phosphatase in the process of follicular atresia in Dipetalogaster maxima (Hemiptera: Reduviidae), a vector of Chagasʼ disease. PLoS ONE. 2015;10:e0130144.
Kang T, Jin R, Zhang Y, Wang H, Lee KS, Jin BR, et al. Functional characterization of the aspartic proteinase cathepsin D in the beet armyworm (Spodoptera exigua). Gene. 2017;617:1–7.
Zhao XF, An XM, Wang JX, Dong DJ, Du XJ, Sueda S, et al. Expression of the Helicoverpa cathepsin B-like proteinase during embryonic development. Arch Insect Biochem Physiol. 2005;58:39–46.
Fagotto F. Yolk degradation in tick eggs: 1. Occurrence of a cathepsin L-like acid proteinase in yolk spheres. Arch Insect Biochem Physiol. 1990;14:217–35.
Seixas A, Santos PC, Velloso FF, Vaz Ida S Jr, Masuda A, Horn F. A Boophilus microplus vitellin-degrading cysteine endopeptidase. Parasitology. 2003;126:155–63.
Abreu LA, Valle D, Manso PP, Façanha AR, Pelajo-Machado M, Masuda H. Proteolytic activity of Boophilus microplus yolk pro-cathepsin D (BYC) is coincident with cortical acidification during embryogenesis. Insect Biochem Mol Biol. 2004;34:443–9.
Silveira AB, Castro-Santos J, Senna R, Logullo C, Fialho E. Tick vitellin is dephosphorylated by a protein tyrosine phosphatase during egg development: effect of dephosphorylation on VT proteolysis. Insect Biochem Mol Biol. 2006;36:200–9.
Nascimento-Silva MC, Leal AT, Daffre S, Juliano L, Vaz Ida S Jr, Paiva-Silva GO, et al. BYC, an atypical aspartic endopeptidase from Rhipicephalus (Boophilus) microplus eggs. Comp Biochem Phys B. 2008;149:599–607.
Oldiges DP, Parizi LF, Zimmer KR, Lorenzini DM, Seixas A, Masuda A, et al. A Rhipicephalus (Boophilus) microplus cathepsin with dual peptidase and antimicrobial activity. Int J Parasitol. 2012;42:635–45.
Zhang TT, Qiu ZX, Li Y, Wang WY, Li MM, Guo P, et al. The mRNA expression and enzymatic activity of three enzymes during embryonic development of the hard tick Haemaphysalis longicornis. Parasites Vectors. 2019;12:96.
Liu JZ, Liu ZN, Zhang Y, Yang XL, Gao ZH. Biology of Dermacentor silvarum (Acari: Ixodidae) under laboratory conditions. Exp Appl Acarol. 2005;36:131–8.
Dai CH, Gan LN, Qin WU, Zi C, Zhu GQ, Wu SL, et al. Use of fluorescence quantitative polymerase chain reaction (PCR) for the detection of Escherichia coli adhesion to pig intestinal epithelial cells. Pol J Vet Sci. 2016;19:619–25.
Huang T, Long M, Huo B. Competitive binding to cuprous ions of protein and BCA in the bicinchoninic acid protein assay. Open Biomed Eng J. 2010;4:271–8.
Gao X, Shi L, Zhou YZ, Cao J, Zhang HS, Zhou JL. Characterization of the anticoagulant protein Rhipilin-1 from the Rhipicephalus haemaphysaloides tick. J Insect Physiol. 2011;57:339–43.
Hao JW, Luo J, Chen Z, Ren QY, Guo JX, Liu XC, et al. MicroRNA-275 and its target vitellogenin-2 are crucial in ovary development and blood digestion of Haemaphysalis longicornis. Parasites Vectors. 2017;10:253.
Canal CW, Maia HM, Vaz Júnior IS, Chies JM, Farias NA, Masuda A, et al. Changing patterns of vitellin-related peptides during development of the cattle tick Boophilus microplus. Exp Appl Acarol. 1995;19:325–36.
Estrela AB, Seixas A, Teixeira Vde O, Pinto AF, Termignoni C. Vitellin- and hemoglobin-digesting enzymes in Rhipicephalus (Boophilus) microplus larvae and females. Comp Biochem Phys B. 2010;157:326–35.
Palomino J, Herrera G, Torres-Fuentes J, Dettleff P, Patel A, Martínez V. Assessment of cathepsin mRNA expression and enzymatic activity during early embryonic development in the yellowtail kingfish Seriola lalandi. Anim Reprod Sci. 2017;180:23–9.
Tingaud-Sequeira A, Carnevali O, Cerdà J. Cathepsin B differential expression and enzyme processing and activity during Fundulus heteroclitus embryogenesis. Comp Biochem Phys A. 2011;158:221–8.
Yang XL, Yu ZJ, He YJ, Xu XL, Gao ZH, Wang H, et al. Purification of vitellin and dynamics of vitellogenesis in the parthenogenetic tick Haemaphysalis longicornis (Acari: Ixodidae). Exp Appl Acarol. 2015;65:377–88.
Yang XL, Yu ZJ, Gao ZH, Yang XH, Liu JZ. Morphological characteristics and developmental changes of the ovary in the tick Haemaphysalis longicornis Neumann. Med Vet Entomol. 2014;28:217–21.
Zhang TT. The role of two enzymes in vitellin degradation and embryogenesis of Haemaphysalis longicornis. PhD Thesis, Hebei Normal University, Shijiazhuang, China; 2019.
Li Y. Changes and roles of cathepsin D in the embryonic development of Haemaphysalis longicornis. MSc Thesis, Hebei Normal University, Shijiazhuang, China; 2019 .
Li Y, Wang K, Xie H, Wang DW, Xu CL, Huang X, et al. Cathepsin B cysteine proteinase is essential for the development and pathogenesis of the plant parasitic nematode Radopholus similis. Int J Biol Sci. 2015;11:1073–87.
Smooker PM, Jayaraj R, Pike RN, Spithill TW. Cathepsin B proteases of flukes: the key to facilitating parasite control? Trends Parasitol. 2010;26:506–14.
Correnti JM, Brindley PJ, Pearce EJ. Long-term suppression of cathepsin B levels by RNA interference retards schistosome growth. Mol Biochem Parasitol. 2005;143:209–15.
Li J, Xiang M, Zhang R, Xu B, Hu W. RNA interference in vivo in Schistosoma japonicum: establishing and optimization of RNAi mediated suppression of gene expression by long dsRNA in the intra-mammalian life stages of worms. Biochem Biophys Res Commun. 2018;503:1004–10.
Goupil LS, Ivry SL, Hsieh I, Suzuki BM, Craik CS, O’Donoghue AJ, et al. Cysteine and aspartyl proteases contribute to protein digestion in the gut of freshwater planaria. PLoS Negl Trop Dis. 2016;10:e0004893.
Lee KS, Kim BY, Choo YM, Yoon HJ, Kang PD, Woo SD, et al. Expression profile of cathepsin B in the fat body of Bombyx mori during metamorphosis. Comp Biochem Phys B. 2009;154:188–94.
Gui ZZ, Lee KS, Kim BY, Choi YS, Wei YD, Choo YM, et al. Functional role of aspartic proteinase cathepsin D in insect metamorphosis. BMC Dev Biol. 2006;6:49.
Morales ME, Rinaldi G, Gobert GN, Kines KJ, Tort JF, Brindley PJ. RNA interference of Schistosoma mansoni cathepsin D, the apical enzyme of the hemoglobin proteolysis cascade. Mol Biochem Parasitol. 2008;157:160–8.
Huang G, Ulrich PN, Storey M, Johnson D, Tischer J, Tovar JA, et al. Proteomic analysis of the acidocalcisome, an organelle conserved from bacteria to human cells. PLoS Pathog. 2014;10:e1004555.
Acknowledgements
The authors are especially grateful to Drs Zhi-Jun Yu and Abolfazl Masoudi for their help in English editing. We also thank Duo Wang for animal care and technical assistance.
Funding
This work was supported by the National Natural Science Foundation of China (Grant No. 31672365) and the Department of Education of Hebei Province (Grant No. CXZZBS2019086).
Author information
Authors and Affiliations
Contributions
JZL and ZXQ designed and the study and drafted the manuscript. ZXQ and YL extracted RNA, synthesized cDNA, designed primers, performed qPCR, extracted protein, detected the activity of enzymes, synthesized and injected dsRNA and analyzed data. MML, WYW and TTZ collected samples, participated in data collection and tick feeding. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This study was approved by the Animal Ethics Committee of Hebei Normal University as complying with the Animal protection law of the People’s Republic of China.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Qiu, ZX., Li, Y., Li, MM. et al. Investigation of three enzymes and their roles in the embryonic development of parthenogenetic Haemaphysalis longicornis. Parasites Vectors 13, 46 (2020). https://doi.org/10.1186/s13071-020-3916-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-020-3916-7