
Abstract
Ovarian cancer (OC) is a heterogeneous malignancy with various etiology, histopathology, and biological feature. Despite accumulating understanding of OC in the post-genomic era, the preclinical knowledge still undergoes limited translation from bench to beside, and the prognosis of ovarian cancer has remained dismal over the past 30 years. Henceforth, reliable preclinical model systems are warranted to bridge the gap between laboratory experiments and clinical practice. In this review, we discuss the status quo of ovarian cancer preclinical models which includes conventional cell line models, patient-derived xenografts (PDXs), patient-derived organoids (PDOs), patient-derived explants (PDEs), and genetically engineered mouse models (GEMMs). Each model has its own strengths and drawbacks. We focus on the potentials and challenges of using these valuable tools, either alone or in combination, to interrogate critical issues with OC.
Similar content being viewed by others
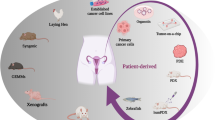

Background
Ovarian cancer incorporates multiple malignancies with a variety of etiology, histopathology, and biological feature, including those derived from epithelial, germ cell, sex cord stromal, and metastatic lesions [1]. On a global scale, OC causes 152,000 deaths among 239,000 new cases annually, making OC the second leading cause of cancer-related death in women [2]. Most OC patients were diagnosed at an advanced stage with extensive peritoneal metastasis, yet symptoms are rather vague [3, 4]. The dismal prognosis, limited treatment options, and high recurrence rate followed by stout resistance together render OC the most fatal gynecologic tumor, and the treatment of OC an ongoing challenge.
The current standard of care for OC is primary debulking surgery followed by platinum-based chemotherapy [5, 6]. Neoadjuvant chemotherapy might be considered for non-operable patients who are intolerant of surgery or deemed unlikely to reach a cytoreduction to no gross residual disease (R0) [7]. The advent of poly ADP-ribose polymerase (PARP) inhibitors, to a certain extent, revolutionized the treatment of patients with BRCA1/2 mutations. This is based on a theory named “synergetic lethality”, which was first described in 1946 [8]. Theoretically, dual inactivation of BRCA and PARP results in replication catastrophe, leading to the inevitable death of cancer cells. However, when it comes to treatment in a clinical setting, PARPi can lose its magic once resistance occurs, not to mention that the predominant beneficiary group carrying BRCA1/2 mutation only accounts for 15% of OC patients.
Personalized medicine allowed advances in the exploitation of alternative strategies to tackle the aforementioned bottlenecks, yet the biological insufficiency of OC pre-clinical models to fully recapitulate the complexity of OC, to a certain extent, restraints the understanding and resolving of this heterogeneous disease [9]. A solid pre-clinical model should mirror the morphological and biological characteristics of the corresponding tumor of origin to the full extent. Historically, ever since the establishment of HELA cells, 2D cell line models have long been the mainstay of experimental cancer research [10]. Immortalized cancer cells cultured in artificial FBS-based medium boosted the exploration of cancer biology, somehow, they failed to bridge the gap between laboratory experiments and clinical trials. Abortions in phase III trials were mainly due to inadequate efficacy, which underlines the discrepancy between conventional cell lines and individual patients [11,12,13]. In response to this intractable situation, generations of pre-clinical models have been developed, among which patient-derived xenografts, organoids, tumor explants, and genetically engineered mouse models emerged as complementary tools in cancer research and drug screening (Fig. 1). In this review, we discuss the current status of OC models, and the strengths and drawbacks of each platform, casting light on the potentials and challenges of using these tools to unravel the biological fingerprints and vulnerabilities of OC.

Schematic representation of the most commonly used OC preclinical models. Both advantages and disadvantages are summarized for each model
Conventional cell lines
Conventional cell line models, ever since the establishment of the HELA cell line, had long been acknowledged to profoundly facilitate the understanding of tumor initiation, progression, metastasis, and drug discovery [10]. Well-established cell line models recapitulate the hallmarks of parental tumors in a way that can mimic the molecular aberrations and vital biological events. As regards to OC, there have been approximately 100 cell lines publicly available, most of which, however, lack proper annotations at histological, cellular, and molecular levels [14]. Based on transcriptomic data, Barnes BM and colleagues clustered 44 OC cell lines into 5 transcriptionally different groups, highly corresponding to the 5 major histological subtypes of OC [15]. It is worthwhile to mention that a bonafide list of reliable cell line models for distinct human pathology is needed to guide type-specific OC research [16, 17]. Currently, compiling works have been devoted to the establishment of OC cell line models with defined pathology and well-characterized phenotypic and genomic profiles. The team of Kreuzinger C innovatively established reliable high-grade serous OC (HGSOC) cell lines from clinical OC samples, while providing a detailed background in clinical parameters, chemosensitivity status, and molecular alterations. And in a further step, a total of 34 established cell lines were used to decode potential therapeutic targets specific for HGSOC [18, 19]. In addition, cell lines for ovarian clear cell carcinoma (CCC) [20], low-grade serous ovarian carcinoma (LGSOC) [21], endometrioid adenocarcinoma (EMC) [22], and other pathology were cultured and introduced by researchers worldwide.
Notably, the OC cell line toolbox is further expanded by the development of drug-resistant cells and syngeneic mouse cell lines as well. A fundamental pain point in OC treatment is the stout resistance to chemotherapies, which calls for in vitro drug-resistant models to help understand the underlying mechanism. Methodologically speaking, one option is to derive resistant models from clinically relevant samples [23, 24], another is to establish isogenic drug-resistant cell lines, usually through stepwise sequential exposure to increased concentration of drugs [25]. For example, the carboplatin-resistant A2780 cells [26], taxol-resistant OC3/TAX300 cells [27]. Interestingly, comparisons between isogenic drug-resistant cell lines and their background counterpart may serve as the lens to look into the trivia during the development of OC drug resistance [28, 29]. As for syngeneic murine cell lines, ID8 is the most widely used and also the most extensively characterized model for epithelial OC. The ability of ID8 cells to form sporadic lesions in the peritoneal cavity of immune-competent mice closely mimics the biological events in stage III and/or IV OC in patients [30]. Henceforth, syngeneic murine cell lines are still representable and the most convenient models to use for the study in the organism with full immunity and comprehensive stroma components.
OC cell line model, as a fundamental tool, remains to be the cornerstone in cancer models for its easy manipulation, low cost, and short doubling time [31]. The US National Cancer Institute (NCI) 60 human tumor cell lines representing 9 distinct tumor types (including ovarian cancer) were assembled in the late 1980s under a disease-oriented concept, aiming at identifying the anti-tumor effect of compounds in particular tumor types [32, 33]. Moving beyond its original intention, the NCI 60 panel, combined with high-throughput technology, served as a pipeline in the field of drug screening [34]. Among the 60 extensively characterized cell lines, IGR-OV1, OVCAR-3, OVCAR-4, OVCAR-5, OVCAR-8, and NCI/ADR-RES are considered to be the 6 most representative OC cell lines to undergo numerous validations of promising small molecule compound [35]. Other large-scale datasets including The Cancer Cell Line Encyclopedia (CCLE) [36], the Cancer Genome Project (CGP) [37], and Cancer Therapeutic Response Portal (CTRP) facilitate the in-depth study of drug response across cell lines.
Established cell lines are and will continue to be of paramount significance in every aspect of OC research. It is now beyond doubt that cell lines facilitated the study of cellular dependency in high-throughput pharmacological interrogation and genetic screening. Bulk of cell line studies on the genome-wide level revealed the genetic alterations related to drug sensitivity and resistance. Papp et al. integrated genomic, epigenomic, and expression analyses to shed light on the molecular abnormalities in an ovarian cancer cell line panel comprised of 45 OC cell lines and revealed unique molecular dependencies of several targeted therapies [38]. Garnett MJ et al. conducted systematic pharmacogenomic profiling in a pan-cancer cohort including 17 OC cell lines to provide an extensive view of the genomics underlying drug sensitivity. Using cell line models, large-scale loss-of-function screens such as siRNA, shRNA, and CRISPR-Cas9 library yielded new insights on identifying “driver” mutations among the “passenger” ones [39,40,41,42]. Mengwasser KE and colleagues conducted shRNA and CRISPR screening with DNA-repair-based libraries on 2 pairs of BRCA2 isogenic cell lines, including ovarian PEO1 B2MUT cells, revealing FEN1 and APEX2 as BRCA2 synthetic lethal targets, which could possibly be utilized in future treatment of BRCA-deficient populations [43]. In addition, the examples of utilizing OC cell line models to predict drug efficacy and explore underlying molecular mechanisms in OC initiation, progression, metastasis, and response to clinical and novel therapies are far too numerous to be detailed. Experimental technique evolving cell line models are well-developed and still undergoing a continuous boom. In vivo tumorigenicity of some cell lines endowed them with the utility to form xenografts for the study of tumor initiation and metastasis as well as in vivo drug response in animal experiments [44, 45].
Though historically served as the main force in cancer research models, conventional cell lines are continuously questioned in their fidelity and clinical relevance [46,47,48,49,50]. Serial analysis of gene expression (SAGE) database identified distinct gene clusters overexpressed in cell lines and solid tumors respectively, which might give rise to the deviations of cell lines in respect of chemotherapy resistance [47]. Comparison between OC cell lines and clinical samples revealed that cell line models failed to capture clinical MDR gene expression patterns with their high selectivity in the expression of genes associated with MDR [48]. A research comprised of 41 OC cell lines indicated that selective pressure against BRCA1/2 mutations during adaption to 2D culture might contribute to a lower incidence of BRCA1/2 deleterious mutations than the population-level incidence [51]. Another comparative study looked at the CNVs, mutations, and mRNA expression profiles between commonly used OC cell lines and HGSOC samples and revealed pronounced gaps in molecular profiles. Startlingly, the most popular OC cell lines such as SK-OV-3, A2780, OVCAR-3, CAOV3, and IGROV1 showed low correlation value with individual tumors while rarely applied cell lines such as KURAMOCHI, OVSAHO stood out as “good cell lines” [50]. However, being a representative cell line does not necessarily mean that it can successfully and efficiently form xenografts in vivo, again restraining the utility of cell line models [44, 52].
As the genomic fingerprints, even druggable targets might be lost over extensive passages in vitro, the utilization of this conventional model is fairly discouraged [48]. Notably, given the random nature of the incidence of genomic drift, it’s possible for identical cell lines to differ even among laboratories. Clearly, OC cell lines commonly used in published studies do not stand for the full complexity of OC. To summarize, conventional OC cell line models, by their very nature, cannot afford the rapid expansion of precision medicine alone.
Patient-derived xenografts
In 1953, Toolan HW made the first systematic report on human tumors growing in cortisone-treated rats. Thereby, the PDX model began to take shape [53]. Although basic concepts and methodologies began to emerge, it was not until the introduction of immunodeficient nude mice that PDX had its first major breakthrough [54]. Bosma, G.C. introduced CB17-SCID mice deficient in immune functions mediated by T and B lymphocytes [55], followed by NOD-SCID mice (Jackson Lab) with abrogated NK cells [56] and NOD-SCID IL2rg null (NSG) mice with no functional T, B and NK cells [57]. Mouse strains mentioned above improved the success rate of transplantation, optimized the methodology, and provided researchers with more options [55,56,57,58]. Specific recipient strain can therefore be applied depending on the specific research purpose. The very first report on successful transplantation and passaging of OC tissue into nude mice could date back to 1977 when Davy M described a poorly differentiated ovarian adenocarcinoma PDX and confirmed its correspondence with the donor patient in drug response [59].
Up till now, compiling works have been devoted to the establishment, characterization and utilization of OC PDX (Table 1). To establish a PDX, fresh OC tissues obtained from surgery, biopsy, and ascites fluid were directly transplanted into immunodeficient mice orthotopically, subcutaneously, or intraperitoneally after simple initial processing. The inoculum can be tissue mass or cell suspension, either alone or coated with upholders such as Matrigel or human fibroblasts [74]. Orthotopic engraftment recapitulates the favorable anatomic microenvironment of the original tumor [75, 76]. Intraperitoneal dissemination and ascites, thereafter, can be formed based on the intrinsic metastasis mechanism of OC [77]. Subcutaneous engraftment, by contrast, is the most common choice for the convenience of observation and monitoring of tumor volume using a caliper [74]. Intraperitoneal transplantation often results in metastatic lesions in the peritoneum, omentum, pelvic and abdominal organs [64]. Albeit better resemblance in terms of biological behaviors, both orthotopic and intraperitoneal engraftment require non-invasive imaging techniques which are relatively time- and resource-consuming. A complementary evaluation method adopted by Joyce F Liu et al. is to monitor surrogate biomarkers such as CA125 and human LINE-1 [64]. Glaser G et al. came up with a physical-exam-based scoring method which was reported to share a high correlation with tumor weight at necropsy [78]. Additionally, the subrenal capsule transplantation offered another option based on a previous report that this highly vascularized site yielded a high take rate [66, 71, 79], however, this was not observed in several other studies [80].
The success rates of OC PDX implantation ranged from 25% [68] to more than 95% [71] according to previous reports. The possible confounding factors are as follows. 1) Pathological type, stage, and grade of the parental tumor. In common sense, samples with more aggressive pathological features and more advanced stages are better able to be engrafted than indolent tumors [81]. 2) The quality of the tissue grafts. Tumor tissues are required to be rapidly transferred from the operating room to the experimental center and a proper medium is needed during the transfer process to ensure viability. Moreover, the segmentation of the specimen, tumor/necrosis percentage of inoculum also play a decisive role. 3) Type of the tissue implanted. The material can be solid tissue chunks or dissociated cell suspension. Metastatic lesions are reported to have a higher take rate than the primary lesion [82]. 4) The strain of recipient mice, which has been described previously. 5) The implantation site. Dobbin et al. evaluated the tumorigenicity of OC in subcutaneous (SQ), mammary fat pat (MFP), intraperitoneal (IP), subrenal capsule (SRC) site and the engraftment rates are 85.3, 63.64, 22.2, and 8.3% respectively [80]. Further systematic comparative studies focusing on improving OC engraftment rate are warranted.
As a heterogeneous disease, the research of OC requires preclinical models that are personalized to solve individual issues, and OC cell lines, as discussed previously, are clearly not up to it. Recently, the NCI decided to take up with PDXs for drug screening as a substitution for the NCI-60 cell line panel, considering that PDX better mimics the human tumor [46]. In fact, PDX models are competent to capture therapeutic candidates that are missing in cell line screening [83]. Numerous studies have demonstrated that the histological structure and genomic signatures (including the mutation profile, CNV, MSI, and STR) are faithfully preserved in murine models and remain stable even after several sequential passages between recipient mice [74]. Liu Y. et al. examined the molecular fidelity of OC PDX versus primary ovarian tumors at the mRNA level and devised a bioinformatic pipeline to separate the murine component transcriptome confounders [84]. Consequently, individual PDX can serve as an avatar model for donor patients, retaining the biological uniqueness to undergo laboratory and clinical interrogation. By expanding PDX through mouse-to-mouse passages, a large cohort of tumor-bearing mice testable can therefore be assembled to receive multiple trials and determine the best regimen for OC. Provided that an OC PDX bank comprising a variety of patients, it can simulate clinical trials based on molecular pathological and molecular characteristics of a real OC patient population. Such concept of “xeno-trials” motivated researchers and organizations to set up international collaborations, for example, the EurOPDX and PDXNET consortium [74, 85]. By integrating worldwide PDX panels together with corresponding clinical data, genomic information, and drug responsiveness, authoritative resources can be achieved to facilitate precision medicine [58].
The advantages of the OC PDX model have evoked promises to tackle crucial issues in OC research. Via serial passaging, the original tumor tissue can be expanded on host mice and harvested to undergo various manipulations. Tissue bio-banked together with tumor-bearing mice, which is a living bio-bank itself, constitute a resourceful OC library [86]. The highlight of PDX has always been to validate the efficacy of various regimens. Parmar K. et al. assessed the activity of prexasertib and its combination with olaparib in 14 clinically characterized PDXs [87]. Cornelison R. et al. utilized PDX with or without chemotherapy to examine the potential of targeting ribosomal machinery in OC [88]. In general, compared with traditional cell lines and cell line-derived xenografts (CDX), OC PDX serves as a better and more convincing pre-clinical model in testing drug response of chemo- and targeted therapy. Consequently, the application of PDX is gaining momentum and has gradually become mainstream in OC research. George E et al. introduced a BRCA-deficient OC PDX platform to test novel targeted therapies [65]. Moving beyond the laboratory field, Colon-Otero G. et al. made the first attempt at PDX co-clinical trial in OC [89]. Unfortunately, this study only attempted to establish PDX with biopsy samples from tested patients, leaving a gap in the drug response of the corresponding tumor-bearing xeno-patients. Pioneering works incorporating PDX cohorts in clinical trials have been done in breast cancer, lung cancer, soft tissue sarcoma et al. [90,91,92,93,94] and we expect further exploration in OC to come. Apart from pre-clinical drug testing, PDX is a suitable tool for biomarker identification and validation. One example comes from clear cell ovarian carcinoma PDX to address the mechanism of carboplatin resistance and the study identified APOBEC3B as a new biomarker for intervention [95]. Palmer AC et al. analyzed the genetic profile of OC PDX models exposed to 21 monotherapies and reported that nearly 90% of models responded to at least one biomarker-guided therapy [96].
Nevertheless, there are certain inherent weaknesses of PDX models that cannot be ignored in OC research. Immunodeficient recipient strains lack key immune components. On the one hand, it avoids host versus graft rejection and enables successful engraftment. On the other hand, it severely restrains the utility of PDX in the exploitation of immune-related therapies, which is a key topic in OC research [74]. In order to solve this issue, humanized mice in which human immune components are introduced into immunocompromised mice have been developed. The immune reconstruction procedure may utilize human peripheral blood, tumor-infiltrating lymphocytes (TILs), or CD34+ human hematopoietic stem cells [97]. Transplantation of different human immune materials renders mice with different immune competencies. In addition, previous reports have confirmed that tumor stroma, though preserved in tumor fragments, is rapidly replaced by mouse counterparts even in the first generation [98]. The taking over of murine stroma altered the transcriptomic signature of the original tumor [99]. A study by the group of Blomme A has previously demonstrated that murine stroma adopts a metabolic phenotype similar to human [100], still, the discrepancies in drug response may occur to an unknown degree. Additionally, the adaptation of cancer cells to the murine microenvironment throughout passages might give priority to certain subpopulations, namely clonal selection [101, 102]. OC is known to be a type of cancer with high heterogeneity, which can be undermined by the enrichment of dominant clones in murine models. The subclone constitution is constantly altered by selection pressure. Therefore, early passages ensure better consistency and predictive value when questing biology and molecular clues [99]. Besides, PDX models are rather hard to handle on a genetic level, genetical intervention could be implemented via managing cell suspension during the passaging process. Last but not least, in view of both known and unknown contributing factors leading to successful engraftment, any PDX archives certainly do not represent the full spectrum of OC.
Patient-derived organoids
In 2009, Hans Clevers and colleagues released a landmark study which documented the initiation of crypt-villus organoids by single sorted Lgr5+ stem cells [103]. This ground-breaking milestone kick-started a brand-new area in stem cell research as well as in 3D culture technology, followed by mountainous study devoted to the development, optimization, and broader utilization of organoid models [104,105,106]. Naturally, organoid was thought to hold great potential to prompt preclinical OC research based on the fact that the delicate 3D structure, to a large extent, carries the phenotypic and genomic characteristics of original tumors, and in the meanwhile preserves the inter- and intra-patient heterogeneity which are major topics of interest in the study of OC [107]. By mechanical or enzymatic digestion, the tumor tissue is embedded in the in vitro 3D matrix, whereby a homoeostatic environment is mimicked by adding growth factors and small molecule inhibitors cocktail. The artificial niche environment allows stem cells to self-organize into 3D structures, self-renew, and multi-differentiate to keep viability and integrity. These features endowed the model with the merits of biological stability through extensive passaging [108]. So far, organoid models of gastrointestinal cancers [109], glioblastoma [110], colorectal cancer [111], pancreatic cancer [112], prostate cancer [113] have been built, setting paradigms for their respective cancer research fields. With regard to OC, Hill et al. innovatively reported a short-term cultured OC platform to identify targetable DNA damage repair defects in parent tumors [114]. The first major breakthrough in expandable OC organoid came from Kopper O’s group. In this pioneering study, an OC organoid platform was built, which enables long-term expansion and genetic manipulation. And the organoid models of various OC subtypes were demonstrated to share high similarity with the corresponding tumor at histological and genomic levels [115]. Subsequent works mainly focused on the fidelity and utility of this cutting-edge technology in the OC research field (Table 2).
When compared with monolayer cell culture, organoid captures a more diverse repertoire that incorporates early-stage neoplastic lesions, contributing to the spectrum of cancer and precancerous models. Considerable discrepancies were reported between conventional cell lines and 3D culture systems. The underlying possibilities for this phenomenon are not only manifold but involute. Lack of cell-cell and cell-matrix interaction can be partly blamed while the diffusion rate of nutrients and metabolic waste might differ greatly in cell aggregates as well [125]. The structural difference in the culturing system brings genomic unconformity and the consequences are far more than that. Loessner D et al. reported a higher survival rate in spheroid-grown OC cells compared to their monolayer counterparts when exposed to paclitaxel treatment, which is the current first-line therapy for OC [126]. One pioneering work conducted by Jabs J’s group in exploiting OC organoid responses to clinically relevant drugs under respective genome alteration backgrounds highlighted the culture system dependency concerning cytostatic drug effect and pharmacogenomic associations [127]. With the aid of DeathPro, an originally designed automated workflow, cell death, and growth arrest rates were evaluated, and it was surprising to find drug effects clustered according to culture type to the same extent as intrinsic tumor heterogeneity [127]. Hopefully, the organoid model is endowed with the ability to uncover key information concealed by monolayer culture, demonstrating more potential and caveats in personalized drug development and biomarker validation. Apart from that, the successful establishment of cell lines is often unpredictable [72]. Generating organoid culture from normal and cancer specimens saves the trouble of getting rid of fibroblast contamination compared with cell line establishment [112].
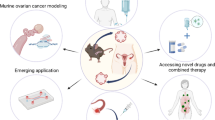
With PDX remains to be the gold standard in the in vivo settings, PDO overcomes several major concerns with regard to PDX. When it comes to preclinical models for OC, one has to consider the expansion rate, and whether that rate provides a suitable time window for decisions to be made under the clinical conditions of OC patients. The rapid expansion capabilities of organoids set up a compatible timeline to keep pace with clinical decision-making. This clinically meaningful time window makes room for real-time co-clinical trials to guide clinical treatment (Fig. 2). Besides, unlike PDX which can only culture malignant tissues, PDO can closely mimic the normal physiology and anatomy of normal tissues by supplementation of appropriate growth factor to preserve the stem cell niche. Similarly, pre-cancerous lesions and tumor types and stages that are underrepresented in PDX modeling can be replenished by PDO [128]. For example, Kessler M et al. established stable human fallopian tube organoids and the generation of mutant organoids was further expected for the study of tumor evolution [129]. Moreover, normal tissue organoids can be used to evaluate the non-specific cytotoxicity of drug candidates, ensuring treatment safety. Bi J et al. created paired tumor and adjacent normal tissue organoids from the same patients and described minimal cell killing of several chemotherapeutic drugs at a tumor-killing dose [118]. Besides, compared with PDX which is unsuitable for high-throughput drug screening, PDO can be exploited as a complementary screening platform at a relatively low cost [130, 131]. Overall, PDO reconciled the pros and cons of traditional cell lines as in vitro models and PDX as in vivo models and bridges the gaps between laboratories and clinical trials.

Schematic representation of co-clinical trial approach with patient-derived OC preclinical models. PDX and/or PDO models are derived from individual OC patients enrolled in a trial. Clinical data from treated patients, in vitro drug response from PDO models, and in vivo drug response from PDX models are collected and integrated with the corresponding sample to undergo comprehensive functional and genetical analysis. The clinical relevant models can therefore be exploited to facilitate the study of drug resistance mechanisms, biomarker development, and drug combination strategy, and to guide future clinical trials. PDX, patient-derived xenograft; PDO, patient-derived organoid; PDXO, PDX-derived organoid; PDOX, PDO-derived xenograft
In terms of applications of PDO, so far, attempts have been made on drug-screening procedures in various cancer types and systematic methodology has been recently reviewed in detail [132]. For OC, Hill SJ et al. devised a short-term HGSOC PDO platform to functionally profile DNA repair defects and further predict the therapeutic effects of drugs targeting DNA damage response (DDR) [114]. Nanki Y et al. performed drug sensitivity and resistance testing of 23 FDA-approved compounds on HGSOC PDOs and proved high concordance of the models [119]. Except for distinguishing responders and non-responders in the laboratory and preclinical drug screening, PDO can be used to capture spatially and chronologically different lesions from one exact patient [133, 134]. Considering the high recurrence rate of OC, patients might undergo multiple invasive medical interventions successively with tissues available to chronologically develop organoid culture. Ideally, the chronological model series could be used to inform clinical decisions and help to monitor the development of drug resistance in a real-time manner. Metastatic lesions of OC can be harvested and built into organoid models concurrently to verify drug response under a heterogenous background [135]. de Witte CJ et al. examined intra-patient heterogeneity and were surprised to find that organoids derived from a patient at different sites varied in monotherapy response of 31% drugs [120].
Moreover, the OC organoid model is expected to be instrumental in immunotherapy research based on its highly manipulatable nature. With immune components incorporated into the organoid culture, the co-culture system poses promising bases for immuno-oncology investigations and facilitates individualized immunotherapy screening [136, 137]. Neal et al. adopted an air-liquid interface (ALI) system to embed tumor cells with T, B, NK cells, and macrophages. The robustness of the model was proved by the accurate preservation of the parental tumor T cell receptor (TCR) spectrum [137]. Disappointingly, Immunotherapies represented by immune checkpoint blockade (ICB) agents demonstrated minimal efficacy in OC which requires a better understanding of the contributions of immune components of OC. Wan C et al. generated organoids from 12 HGSOC patients and co-cultured the models with a full component of intra-tumoral immune cells to study the mechanisms of ICBs. The application of ICBs led to decreased T cell and NK cell exhaustion and BRD1 was identified as a possible immune therapy target [138]. Other tumor microenvironment (TME) elements such as cancer-associated fibroblasts (CAF) and human blood vessel components recapitulate other aspects of the tumor microenvironment in vitro [139, 140]. For example, human umbilical vascular endothelial cells were added into OC organoid system to better mimic the early organogenesis of the human fallopian tube [141]. Additionally, the role of cancer-associated mesothelial (CAM) cells in OC was studied in a co-culture system and was proved to promote tumor chemoresistance [142].
Furthermore, organoid models are inherently amenable to genetic manipulation. Genetic aberrations can be easily introduced into culture systems by transfection and viral infection. This feature is favorable for cancer modeling and oncogene validation [143, 144]. In the field of OC research, Zhang S et al. modeled the initiation of HGSOC in mouse fallopian tube epithelial (FTE) organoids by lentivirus transduction and/or CRISPR/Cas9 mutagenesis [145]. More recently, exponential interest has been fostered in PDO co-clinical trials. Rational combining PDO, as well as PDX in clinical trials, facilitates information integration, biomarker development, and pharmacodynamic monitoring [104]. A clinical observational trial has been launched to investigate potential therapeutic for OC (NCT04555473).
Despite the great promise of PDO in OC research, translational challenges still remain. First, there are noticeable differences in the procedure of organoid establishment and passaging among different researchers. A standardized ingredients and procedure of OC organoid culture and drug response assessment criterion are further warranted. Besides, the assay methods of this 3D culture system are fairly limited and the protocols are more sophisticated and require a relatively high workload. It is gratifying that efforts have been made by experts to confront these challenges [146]. We could hold enthusiasm and expectation to explore more possibilities with OC PDO.
Patient-derived explants
Another patient-derived model taken into consideration is the patient-derived explants (PDE), which refers to the short-term ex vivo culture of freshly obtained, surgically resected human tissue, either in chunks or cut into slices [147]. Although the methodology of PDE and PDE-related drug testing has been around for a long time, unfortunately, it has not translated into a wide range of applications and thus has not become mainstream in cancer research, particularly in OC. Limited reports have been made on the preclinical application of this model. Hence we will briefly introduce the PDE model and its application in OC research.
Technologically, the thickness of explants might affect the viability during culture and drug response considering the efficiency of nutrition diffusion and metabolites transportation. Manual slicing with surgical equipment used to be the most common method in tissue preparation yet has now been gradually replaced by mechanized methods such as the vibratome [148], which can uniform the thickness of slices. According to the report of Parajuli N et al., a slice thickness of 160 μm is optimal for tissue handling and viability [149]. There is no universal formula for PDE culture conditions, tissue-specific supplements are thought to improve the viability of different types of PDEs [150, 151]. Explants could be placed on support media such as gelatin sponges and pore membranes or could be cultured in a free-floating manner [147]. OC tissue could be cultured for about 5 days on gelatin sponges in RPMI1640 media with serum added [151]. Abreu S et al. reported OC free-floating PDE culture which reached the longevity of 30 days in DMEM media with serum addition [152]. Both studies lack supplementary additives to extend the lifetime of OC PDE models. Another technical issue is the endpoint analysis of PDE, which is critical for the evaluation of drug response. The most frequently used strategy is to measure cell viability with MTT assay after enzymatic digestion of the explant. Immunohistochemistry analysis of protein markers of cell proliferation and death is another commonly adopted method. Recently, multiple-immunofluorescence was adopted by researchers, offering innovative analytical approaches and better interpretation for PDE assays [147].
Though less adopted in OC research, the PDE model has its own unique advantages compared with PDX and PDO models. Firstly, PDE can be rapidly generated post-surgery without much technological and financial burden [153]. Besides, PDE retained the cell-cell interaction and cell polarity in a much more proper way. The existence of resident immune cells was validated by immunohistochemistry on OC PDE, showing evidence of the perseverance of tumor-infiltrating CD4+ and CD8+ T cells, macrophages and B cells [152]. Though more intuitively correlating drug response with patient pathology, the longevity of PDE severely restricted its practical applications. However, we hold the belief that by rationally combing this short-term culture with other models in OC research, PDE can assist in OC precision medicine.
Genetically engineered mouse model
The first transgenic onco-mice generated by pronuclear injection of oncogene DNA were reported in the 1980s, initiating a novel field of genetically engineered mouse models (GEMMs) created by accurate manipulation of specific gene expression [154]. So far, the application of this sophisticated mouse model includes but is not limited to various types of cancers, helping to figure out the multistage in tumor initiation and progression, validate candidate genes and assess therapeutic efficacy [155]. Though tumor xenografts remain the most extensively applied mouse models in preclinical research, GEMMs circumvent several issues with xenografts that were discussed in the previous chapter [156]. In GEMMs, tumor cells are generated de novo in the context of a native milieu and within a whole organism. This way, the crucial tumor properties and modulators, such as immune cells and stromal elements are preserved and tumor cells can therefore co-evolve with the surrounding microenvironment [155].
Despite the success of GEMMs in other tumor types, OC GEMMs remain less than satisfactory. The putative reason is complex, yet can mainly be attributed to the paucity of prior knowledge of the origin and genetic basis of OC [157]. Above all, there are still controversies regarding the precursor of OC. On the genetic level, mutations in TP53 predominated the mutational spectrum of OC, other extensively reported alterations include RB1, EGFR, PTEN, PI3K/AKT, BRCA1/2, and KRAS et al. [158]. But how and to what extent perturbation of these genes contributes to the oncogenesis and progression of OC remains enigmatic. Efforts have been made to explore the consequence of the ablation or overexpression of genes of interest in OC GEMMs, which has been reviewed in Table 3.
Another factor that confounds the path of developing OC GEMM might be the lack of validated ovary-specific promoters to facilitate tissue or cell-specific functioning of the gene-editing system. In 2003, Connolly et al. reported the first transgenic mouse with poorly differentiated ovarian carcinoma which was developed by induced expression of the transforming region of SV40 driven by the Müllerian inhibitory substance type II receptor gene promoter (MISRII) [175]. SV40 Tag could functionally inactivate the tumor suppressor gene RB and P53, leading to malignant transformation of epithelial cells [176]. Using this method, approximately 50% of cases successfully developed into bilateral OC with peritoneal metastasis and ascites, which shared high similarity with clinical OC patients and thus were of high clinical relevance. In addition, the malignant ascites of the model were further utilized to establish a cell line model, which exhibited the key properties of epithelial OC. Notably, as a major defect in this transgenic model, the infertility of the female mice precluded the stabilization and expansion of this transgenic line [175]. In order to solve this issue, Connolly’s group generated an affected male founder, TgMISIIR-Tag-DR26, the female offspring of which would develop bilateral ovarian tumors with varying latency and similar histological characteristics of HGSOC. By backcrossing the TgMISIIR-TAg transgenic line, the authors obtained murine ovarian carcinoma (MOVCAR) cell lines from the malignant ascites of tumor-bearing C57BL/6 TgMISIIR-TAg transgenic mice [177]. Next, the advent of the Cre-loxP system helped to explore more possibilities in OC GEMMs development [157]. Regarding the mechanism of this mammalian gene-editing technology, Cre recombinase discovered from bacteriophage P1 recognizes a 34 base pair specific sequences called loxP site and meditates exact recombination between two loxP sites that flank the target gene. Apart from gene excision, the preset location and orientation of loxP sites can also mediate gene translocation and inversion [178]. Later, strategies were explored including AdCre injection into the ovarian bursa and Amhr2, Pax8, and Ovgp1-mediated Cre expression in Müllerian-derived epithelia and rendered various OC phenotypes under the manipulation of different suspicious OC driver genes [179].
The most extensive application of GEMMs lies in the study of cancer initiation and progression. A quadruple combination of perturbations including Pten, Trp53, Rb1, and/or Cdh1 was adopted by Shi M et al. using Amhr2cre/+ mice, developing invasive OC with extensive peritoneal metastasis by targeting ovarian surface epithelium [160]. The cell-of-origin of HGSOC remains controversial during the past decades. GEMMs are suitable models to decipher this critical question [180,181,182]. Flesken-Nikitin A et al. identified the hilum region of the mouse ovary as a previously undefined stem cell niche of the OSE and were susceptible to malignant transformation into epithelial OC. Hilum cells showed preferential transformation after conditional deletion of Trp53 and Rb1 using the Ad-Cre/LoxP system [183]. By introducing genetic abnormalities of combined RB family inactivation and Tp53 mutation in Pax8+ FTE and Lgr5+ OSE or OSE-derived organoids, Zhang S et al. confirmed that HGSOC may originate from both FTE and OSE and the biological behavior of tumor might vary between different tumor of origins [184].
Collectively, OC GEMMs still face major challenges both in development and application. This kind of model may have a relative disadvantage in mirroring the heterogeneity of OC, but as critically emphasized, GEMM bears an irreplaceable value in the study of carcinogenesis and the cross-talk between immune and stromal components and cancer cells.
Conclusions
The past few years have witnessed accumulating knowledge concerning the tumorigenesis, progression, and evolution of ovarian cancer, which can be largely attributed to the development of a plethora of faithful preclinical OC models. It is now clear that OC is hallmarked by a high degree of inter- and intra-patient heterogeneity. By phenocopying the original tumor and/or expanding vital patient-derived tissues, tumor experimental models allow in-depth preclinical assessment of drug candidates and identification of tumor biomarkers at an individual level. The development and optimization of OC models are still actively ongoing, with the overall goal of better management and even the cure of OC.
Given the inherent strengths and drawbacks of each model, it is imperative to be aware that one single model alone is definitely not competent to cover all OC research. Wise selection and rational combination of OC models are instrumental in solving the pain points in respect of OC. GEMMs are naturally suitable models for studying the cell of origin of OC and can be complemented by the newly developed OC organoid model. For the study of tumor invasion and metastasis, GEMMs and orthotopic PDX models can be utilized to mimic the biological process, otherwise, cell line and organoid models could be used to exploit the underlying mechanism. As for capturing and deciphering the heterogeneity and clonal evolution of OC, the highly individualized PDX, PDO, and PDE models stood out as edged tools. Ulteriorly, the patient-derived personalized models could be applied to drug development and repurposing, serving as “avatar models” for individual patients and further facilitating patient stratification, drug response assessment, and biomarker development in a clinical setting.
To conclude, research in the post-genomic era yielded brand new insights into the biological and genetic fingerprints of OC. Henceforth, robust tumor models are required to validate the insights and distinguish those of value and therefore targetable. Conventional cell lines, PDXs, PDOs, PDEs, and GEMMs are all historically indispensable models for OC research. Especially, to realize the full translation from bench to beside, the new generation of patient-derived models will undoubtedly grow to be the mainstream in precision medicine. Future OC research should flexibly adopt suitable experimental models for various applications.
Availability of data and materials
Not applicable.
Abbreviations
- OC:
-
Ovarian cancer
- PDX:
-
Patient-derived xenograft
- PDO:
-
Patient-derived organoid
- PDE:
-
Patient-derived explant
- GEMMs:
-
Genetically engineered mouse models
- PARP:
-
Poly ADP-ribose polymerase
- PARPi:
-
PARP inhibitor
- BRCA:
-
Breast cancer susceptibility gene
- FBS:
-
Fetal bovine serum
- HGSOC:
-
High-grade serous ovarian carcinoma
- CCC:
-
Clear cell carcinoma
- LGSOC:
-
Low-grade serous ovarian carcinoma
- EMC:
-
Endometrioid adenocarcinoma
- NCI:
-
The National Cancer Institute
- CCLE:
-
The Cancer Cell Line Encyclopedia
- CGP:
-
The Cancer Genome Project
- CTRP:
-
The Cancer Therapeutic Response Portal
- CRISPR:
-
Clustered regularly interspersed short palindromic repeats
- SAGE:
-
Serial analysis of gene expression
- MDR:
-
Multiple drug resistance
- NSG:
-
NOD-SCID IL2rg null
- OTP:
-
Orthotopic
- SQ:
-
Subcutaneous
- IP:
-
Intraperitoneal
- SRC:
-
Subrenal capsule
- MFP:
-
Mammary fat pat
- CNV:
-
Copy number variation
- MSI:
-
Microsatellite instability
- STR:
-
Short tandem repeat
- CDX:
-
Cell line-derived xenograft
- TIL:
-
Tumor-infiltrating lymphocyte
- DDR:
-
DNA damage response
- FDA:
-
Food and drug administration (USA)
- ALI:
-
Air-liquid interface
- TCR:
-
T cell receptor
- ICB:
-
Immune checkpoint blockade
- TME:
-
Tumor microenviorment
- CAF:
-
Cancer-associated fibroblasts
- CAM:
-
Cancer-associated mesothelial
- FTE:
-
Fallopian tube epithelial
- MISRII:
-
Müllerian inhibitory substance type II receptor gene
References
Karnezis AN, Cho KR, Gilks CB, Pearce CL, Huntsman DG. The disparate origins of ovarian cancers: pathogenesis and prevention strategies. Nat Rev Cancer. 2017;17(1):65–74.
Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424.
National Cancer Institute. Cancer stat facts: ovarian cancer. Available from: https://seer.cancer.gov/statfacts/html/ovary.html.
Naora H, Montell DJ. Ovarian cancer metastasis: integrating insights from disparate model organisms. Nat Rev Cancer. 2005;5(5):355–66.
Lheureux S, Gourley C, Vergote I, Oza AM. Epithelial ovarian cancer. Lancet. 2019;393(10177):1240–53.
Raja FA, Chopra N, Ledermann JA. Optimal first-line treatment in ovarian cancer. Ann Oncol. 2012;23(Suppl 10):x118–27.
Kuroki L, Guntupalli SR. Treatment of epithelial ovarian cancer. BMJ. 2020;371:m3773.
Dobzhansky T. Genetics of natural populations; recombination and variability in populations of Drosophila pseudoobscura. Genetics. 1946;31(3):269–90.
Johnson JI, Decker S, Zaharevitz D, Rubinstein LV, Venditti JM, Schepartz S, et al. Relationships between drug activity in NCI preclinical in vitro and in vivo models and early clinical trials. Br J Cancer. 2001;84(10):1424–31.
Scherer WF, Syverton JT, Gey GO. Studies on the propagation in vitro of poliomyelitis viruses. IV. Viral multiplication in a stable strain of human malignant epithelial cells (strain HeLa) derived from an epidermoid carcinoma of the cervix. J Exp Med. 1953;97(5):695–710.
Harrison RK. Phase II and phase III failures: 2013-2015. Nat Rev Drug Discov. 2016;15(12):817–8.
Alteri E, Guizzaro L. Be open about drug failures to speed up research. Nature. 2018;563(7731):317–9.
DiMasi JA, Reichert JM, Feldman L, Malins A. Clinical approval success rates for investigational cancer drugs. Clin Pharmacol Ther. 2013;94(3):329–35.
Beaufort CM, Helmijr JC, Piskorz AM, Hoogstraat M, Ruigrok-Ritstier K, Besselink N, et al. Ovarian cancer cell line panel (OCCP): clinical importance of in vitro morphological subtypes. Plos One. 2014;9(9):e103988.
Barnes BM, Nelson L, Tighe A, Burghel GJ, Lin IH, Desai S, et al. Distinct transcriptional programs stratify ovarian cancer cell lines into the five major histological subtypes. Genome Med. 2021;13(1):140.
Anglesio MS, Wiegand KC, Melnyk N, Chow C, Salamanca C, Prentice LM, et al. Type-specific cell line models for type-specific ovarian cancer research. Plos One. 2013;8(9):e72162.
Jacob F, Nixdorf S, Hacker NF, Heinzelmann-Schwarz VA. Reliable in vitro studies require appropriate ovarian cancer cell lines. J Ovarian Res. 2014;7:60.
Kreuzinger C, Gamperl M, Wolf A, Heinze G, Geroldinger A, Lambrechts D, et al. Molecular characterization of 7 new established cell lines from high grade serous ovarian cancer. Cancer Lett. 2015;362(2):218–28.
Kreuzinger C, von der Decken I, Wolf A, Gamperl M, Koller J, Karacs J, et al. Patient-derived cell line models revealed therapeutic targets and molecular mechanisms underlying disease progression of high grade serous ovarian cancer. Cancer Lett. 2019;459:1–12.
Jiang W, Ye S, Xiang L, Yang W, He T, Pei X, et al. Establishment and molecular characterization of a human ovarian clear cell carcinoma cell line (FDOV1). J Ovarian Res. 2018;11(1):58.
De Thaye E, Van de Vijver K, Van der Meulen J, Taminau J, Wagemans G, Denys H, et al. Establishment and characterization of a cell line and patient-derived xenograft (PDX) from peritoneal metastasis of low-grade serous ovarian carcinoma. Sci Rep. 2020;10(1):6688.
Yamada T, Kanda T, Mori H, Shimokawa K, Kagawa M, Shibayama Y. Establishment and characterization of a cell line (NOMH-1) originating from a human endometrioid adenocarcinoma of the ovary. J Ovarian Res. 2013;6(1):8.
Akahane T, Hirasawa A, Imoto I, Okubo A, Itoh M, Nanki Y, et al. Establishment and characterization of a new malignant peritoneal mesothelioma cell line, KOG-1, from the ascitic fluid of a patient with pemetrexed chemotherapy resistance. Hum Cell. 2020;33(1):272–82.
Teng PN, Bateman NW, Wang G, Litzi T, Blanton BE, Hood BL, et al. Establishment and characterization of a platinum- and paclitaxel-resistant high grade serous ovarian carcinoma cell line. Hum Cell. 2017;30(3):226–36.
Behrens BC, Hamilton TC, Masuda H, Grotzinger KR, Whang-Peng J, Louie KG, et al. Characterization of a cis-diamminedichloroplatinum (II)-resistant human ovarian cancer cell line and its use in evaluation of platinum analogues. Cancer Res. 1987;47(2):414–8.
Viscarra T, Buchegger K, Jofre I, Riquelme I, Zanella L, Abanto M, et al. Functional and transcriptomic characterization of carboplatin-resistant A2780 ovarian cancer cell line. Biol Res. 2019;52(1):13.
Zhang J, Zhao J, Zhang W, Liu G, Yin D, Li J, et al. Establishment of paclitaxel-resistant cell line and the underlying mechanism on drug resistance. Int J Gynecol Cancer. 2012;22(9):1450–6.
Cicchillitti L, Di Michele M, Urbani A, Ferlini C, Donat MB, Scambia G, et al. Comparative proteomic analysis of paclitaxel sensitive A2780 epithelial ovarian cancer cell line and its resistant counterpart A2780TC1 by 2D-DIGE: the role of ERp57. J Proteome Res. 2009;8(4):1902–12.
Golan Berman H, Chauhan P, Shalev S, Hassanain H, Parnas A, Adar S. Genomic characterization of cisplatin response uncovers priming of cisplatin-induced genes in a resistant cell line. Int J Mol Sci. 2021;22(11):5814.
Roby KF, Taylor CC, Sweetwood JP, Cheng Y, Pace JL, Tawfik O, et al. Development of a syngeneic mouse model for events related to ovarian cancer. Carcinogenesis. 2000;21(4):585–91.
Li S, Zhang Z, Han L. Molecular treasures of Cancer cell lines. Trends Mol Med. 2019;25(8):657–9.
Shoemaker RH. The NCI60 human tumour cell line anticancer drug screen. Nat Rev Cancer. 2006;6(10):813–23.
Ledford H. US cancer institute to overhaul tumour cell lines. Nature. 2016;530(7591):391.
Abaan OD, Polley EC, Davis SR, Zhu YJ, Bilke S, Walker RL, et al. The exomes of the NCI-60 panel: a genomic resource for cancer biology and systems pharmacology. Cancer Res. 2013;73(14):4372–82.
List of NCI-60 Human Tumor Cell Lines Available from: https://dtp.cancer.gov/discovery_development/nci-60/cell_list.htm.
Barretina J, Caponigro G, Stransky N, Venkatesan K, Margolin AA, Kim S, et al. The Cancer cell line encyclopedia enables predictive modelling of anticancer drug sensitivity. Nature. 2012;483(7391):603–7.
Garnett MJ, Edelman EJ, Heidorn SJ, Greenman CD, Dastur A, Lau KW, et al. Systematic identification of genomic markers of drug sensitivity in cancer cells. Nature. 2012;483(7391):570–5.
Papp E, Hallberg D, Konecny GE, Bruhm DC, Adleff V, Noë M, et al. Integrated genomic, Epigenomic, and expression analyses of ovarian Cancer cell lines. Cell Rep. 2018;25(9):2617–33.
Marcotte R, Brown KR, Suarez F, Sayad A, Karamboulas K, Krzyzanowski PM, et al. Essential gene profiles in breast, pancreatic, and ovarian cancer cells. Cancer Discov. 2012;2(2):172–89.
Huesken D, Lange J, Mickanin C, Weiler J, Asselbergs F, Warner J, et al. Design of a genome-wide siRNA library using an artificial neural network. Nat Biotechnol. 2005;23(8):995–1001.
Koike-Yusa H, Li Y, Tan EP, Velasco-Herrera Mdel C, Yusa K. Genome-wide recessive genetic screening in mammalian cells with a lentiviral CRISPR-guide RNA library. Nat Biotechnol. 2014;32(3):267–73.
Cheung HW, Cowley GS, Weir BA, Boehm JS, Rusin S, Scott JA, et al. Systematic investigation of genetic vulnerabilities across cancer cell lines reveals lineage-specific dependencies in ovarian cancer. Proc Natl Acad Sci U S A. 2011;108(30):12372–7.
Mengwasser KE, Adeyemi RO, Leng Y, Choi MY, Clairmont C, D'Andrea AD, et al. Genetic screens reveal FEN1 and APEX2 as BRCA2 synthetic lethal targets. Mol Cell. 2019;73(5):885–99.e6.
Hernandez L, Kim MK, Lyle LT, Bunch KP, House CD, Ning F, et al. Characterization of ovarian cancer cell lines as in vivo models for preclinical studies. Gynecol Oncol. 2016;142(2):332–40.
Mitra AK, Davis DA, Tomar S, Roy L, Gurler H, Xie J, et al. In vivo tumor growth of high-grade serous ovarian cancer cell lines. Gynecol Oncol. 2015;138(2):372–7.
Gillet JP, Varma S, Gottesman MM. The clinical relevance of cancer cell lines. J Natl Cancer Inst. 2013;105(7):452–8.
Stein WD, Litman T, Fojo T, Bates SE. A serial analysis of gene expression (SAGE) database analysis of chemosensitivity: comparing solid tumors with cell lines and comparing solid tumors from different tissue origins. Cancer Res. 2004;64(8):2805–16.
Gillet JP, Calcagno AM, Varma S, Marino M, Green LJ, Vora MI, et al. Redefining the relevance of established cancer cell lines to the study of mechanisms of clinical anti-cancer drug resistance. Proc Natl Acad Sci U S A. 2011;108(46):18708–13.
Sandberg R, Ernberg I. Assessment of tumor characteristic gene expression in cell lines using a tissue similarity index (TSI). Proc Natl Acad Sci U S A. 2005;102(6):2052–7.
Domcke S, Sinha R, Levine DA, Sander C, Schultz N. Evaluating cell lines as tumour models by comparison of genomic profiles. Nat Commun. 2013;4:2126.
Stordal B, Timms K, Farrelly A, Gallagher D, Busschots S, Renaud M, et al. BRCA1/2 mutation analysis in 41 ovarian cell lines reveals only one functionally deleterious BRCA1 mutation. Mol Oncol. 2013;7(3):567–79.
Elias KM, Emori MM, Papp E, MacDuffie E, Konecny GE, Velculescu VE, et al. Beyond genomics: critical evaluation of cell line utility for ovarian cancer research. Gynecol Oncol. 2015;139(1):97–103.
Toolan HW. Growth of human tumors in cortisone-treated laboratory animals: the possibility of obtaining permanently transplantable human tumors. Cancer Res. 1953;13(4–5):389–94.
Flanagan SP. ‘Nude’, a new hairless gene with pleiotropic effects in the mouse. Genet Res. 1966;8(3):295–309.
Bosma GC, Custer RP, Bosma MJ. A severe combined immunodeficiency mutation in the mouse. Nature. 1983;301(5900):527–30.
Makino S, Kunimoto K, Muraoka Y, Mizushima Y, Katagiri K, Tochino Y. Breeding of a non-obese, diabetic strain of mice. Jikken Dobutsu. 1980;29(1):1–13.
Ito M, Hiramatsu H, Kobayashi K, Suzue K, Kawahata M, Hioki K, et al. NOD/SCID/gamma(c)(null) mouse: an excellent recipient mouse model for engraftment of human cells. Blood. 2002;100(9):3175–82.
Okada S, Vaeteewoottacharn K, Kariya R. Application of highly immunocompromised mice for the establishment of patient-derived xenograft (PDX) models. Cells. 2019;8(8):889.
Davy M, Mossige J, Johannessen JV. Heterologous growth of human ovarian cancer. A new in vivo testing system. Acta Obstet Gynecol Scand. 1977;56(1):55–9.
Shin HY, Lee EJ, Yang W, Kim HS, Chung D, Cho H, et al. Identification of prognostic markers of gynecologic cancers utilizing patient-derived xenograft mouse models. Cancers (Basel). 2022;14(3):829.
Cybula M, Wang L, Wang L, Drumond-Bock AL, Moxley KM, Benbrook DM, et al. Patient-derived xenografts of high-grade serous ovarian cancer subtype as a powerful tool in pre-clinical research. Cancers (Basel). 2021;13(24):6288.
Chen J, Jin Y, Li S, Qiao C, Peng X, Li Y, et al. Patient-derived xenografts are a reliable preclinical model for the personalized treatment of epithelial ovarian cancer. Front Oncol. 2021;11:744256.
Cybulska P, Stewart JM, Sayad A, Virtanen C, Shaw PA, Clarke B, et al. A Genomically characterized collection of high-grade serous ovarian cancer xenografts for preclinical testing. Am J Pathol. 2018;188(5):1120–31.
Liu JF, Palakurthi S, Zeng Q, Zhou S, Ivanova E, Huang W, et al. Establishment of patient-derived tumor xenograft models of epithelial ovarian Cancer for preclinical evaluation of novel therapeutics. Clin Cancer Res. 2017;23(5):1263–73.
George E, Kim H, Krepler C, Wenz B, Makvandi M, Tanyi JL, et al. A patient-derived-xenograft platform to study BRCA-deficient ovarian cancers. JCI Insight. 2017;2(1):e89760.
Heo EJ, Cho YJ, Cho WC, Hong JE, Jeon HK, Oh DY, et al. Patient-derived xenograft models of epithelial ovarian Cancer for preclinical studies. Cancer Res Treat. 2017;49(4):915–26.
Dong R, Qiang W, Guo H, Xu X, Kim JJ, Mazar A, et al. Histologic and molecular analysis of patient derived xenografts of high-grade serous ovarian carcinoma. J Hematol Oncol. 2016;9(1):92.
Ricci F, Bizzaro F, Cesca M, Guffanti F, Ganzinelli M, Decio A, et al. Patient-derived ovarian tumor xenografts recapitulate human clinicopathology and genetic alterations. Cancer Res. 2014;74(23):6980–90.
Topp MD, Hartley L, Cook M, Heong V, Boehm E, McShane L, et al. Molecular correlates of platinum response in human high-grade serous ovarian cancer patient-derived xenografts. Mol Oncol. 2014;8(3):656–68.
Weroha SJ, Becker MA, Enderica-Gonzalez S, Harrington SC, Oberg AL, Maurer MJ, et al. Tumorgrafts as in vivo surrogates for women with ovarian cancer. Clin Cancer Res. 2014;20(5):1288–97.
Lee CH, Xue H, Sutcliffe M, Gout PW, Huntsman DG, Miller DM, et al. Establishment of subrenal capsule xenografts of primary human ovarian tumors in SCID mice: potential models. Gynecol Oncol. 2005;96(1):48–55.
Verschraegen CF, Hu W, Du Y, Mendoza J, Early J, Deavers M, et al. Establishment and characterization of cancer cell cultures and xenografts derived from primary or metastatic Mullerian cancers. Clin Cancer Res. 2003;9(2):845–52.
Massazza G, Tomasoni A, Lucchini V, Allavena P, Erba E, Colombo N, et al. Intraperitoneal and subcutaneous xenografts of human ovarian carcinoma in nude mice and their potential in experimental therapy. Int J Cancer. 1989;44(3):494–500.
Hidalgo M, Amant F, Biankin AV, Budinská E, Byrne AT, Caldas C, et al. Patient-derived xenograft models: an emerging platform for translational cancer research. Cancer Discov. 2014;4(9):998–1013.
Kim MP, Evans DB, Wang H, Abbruzzese JL, Fleming JB, Gallick GE. Generation of orthotopic and heterotopic human pancreatic cancer xenografts in immunodeficient mice. Nat Protoc. 2009;4(11):1670–80.
Hoffman RM. Patient-derived orthotopic xenografts: better mimic of metastasis than subcutaneous xenografts. Nat Rev Cancer. 2015;15(8):451–2.
Vanderhyden BC, Shaw TJ, Ethier JF. Animal models of ovarian cancer. Reprod Biol Endocrinol. 2003;1:67.
Glaser G, Weroha SJ, Becker MA, Hou X, Enderica-Gonzalez S, Harrington SC, et al. Conventional chemotherapy and oncogenic pathway targeting in ovarian carcinosarcoma using a patient-derived tumorgraft. Plos One. 2015;10(5):e0126867.
Press JZ, Kenyon JA, Xue H, Miller MA, De Luca A, Miller DM, et al. Xenografts of primary human gynecological tumors grown under the renal capsule of NOD/SCID mice show genetic stability during serial transplantation and respond to cytotoxic chemotherapy. Gynecol Oncol. 2008;110(2):256–64.
Dobbin ZC, Katre AA, Steg AD, Erickson BK, Shah MM, Alvarez RD, et al. Using heterogeneity of the patient-derived xenograft model to identify the chemoresistant population in ovarian cancer. Oncotarget. 2014;5(18):8750–64.
Eoh KJ, Chung YS, Lee SH, Park SA, Kim HJ, Yang W, et al. Comparison of clinical features and outcomes in epithelial ovarian Cancer according to Tumorigenicity in patient-derived xenograft models. Cancer Res Treat. 2018;50(3):956–63.
Némati F, Sastre-Garau X, Laurent C, Couturier J, Mariani P, Desjardins L, et al. Establishment and characterization of a panel of human uveal melanoma xenografts derived from primary and/or metastatic tumors. Clin Cancer Res. 2010;16(8):2352–62.
Gao H, Korn JM, Ferretti S, Monahan JE, Wang Y, Singh M, et al. High-throughput screening using patient-derived tumor xenografts to predict clinical trial drug response. Nat Med. 2015;21(11):1318–25.
Liu Y, Chanana P, Davila JI, Hou X, Zanfagnin V, McGehee CD, et al. Gene expression differences between matched pairs of ovarian cancer patient tumors and patient-derived xenografts. Sci Rep. 2019;9(1):6314.
Conte N, Mason JC, Halmagyi C, Neuhauser S, Mosaku A, Yordanova G, et al. PDX Finder: A portal for patient-derived tumor xenograft model discovery. Nucleic Acids Res. 2019;47(D1):D1073–d9.
Alkema NG, Tomar T, Duiker EW, Jan Meersma G, Klip H, van der Zee AG, et al. Biobanking of patient and patient-derived xenograft ovarian tumour tissue: efficient preservation with low and high fetal calf serum based methods. Sci Rep. 2015;5:14495.
Parmar K, Kochupurakkal BS, Lazaro JB, Wang ZC, Palakurthi S, Kirschmeier PT, et al. The CHK1 inhibitor Prexasertib exhibits monotherapy activity in high-grade serous ovarian Cancer models and sensitizes to PARP inhibition. Clin Cancer Res. 2019;25(20):6127–40.
Cornelison R, Dobbin ZC, Katre AA, Jeong DH, Zhang Y, Chen D, et al. Targeting RNA-polymerase I in both Chemosensitive and Chemoresistant populations in epithelial ovarian Cancer. Clin Cancer Res. 2017;23(21):6529–40.
Colon-Otero G, Weroha SJ, Foster NR, Haluska P, Hou X, Wahner-Hendrickson AE, et al. Phase 2 trial of everolimus and letrozole in relapsed estrogen receptor-positive high-grade ovarian cancers. Gynecol Oncol. 2017;146(1):64–8.
Savaikar MA, Whitehead T, Roy S, Strong L, Fettig N, Prmeau T, et al. Preclinical PERCIST and 25% of SUV (max) threshold: precision imaging of response to therapy in co-clinical (18) F-FDG PET imaging of triple-negative breast cancer patient-derived tumor xenografts. J Nucl Med. 2020;61(6):842–9.
Owonikoko TK, Zhang G, Kim HS, Stinson RM, Bechara R, Zhang C, et al. Patient-derived xenografts faithfully replicated clinical outcome in a phase II co-clinical trial of arsenic trioxide in relapsed small cell lung cancer. J Transl Med. 2016;14(1):111.
Whitley MJ, Cardona DM, Lazarides AL, Spasojevic I, Ferrer JM, Cahill J, et al. A mouse-human phase 1 co-clinical trial of a protease-activated fluorescent probe for imaging cancer. Sci Transl Med. 2016;8(320):320ra4.
Chen Z, Akbay E, Mikse O, Tupper T, Cheng K, Wang Y, et al. Co-clinical trials demonstrate superiority of crizotinib to chemotherapy in ALK-rearranged non-small cell lung cancer and predict strategies to overcome resistance. Clin Cancer Res. 2014;20(5):1204–11.
Kim HR, Kang HN, Shim HS, Kim EY, Kim J, Kim DJ, et al. Co-clinical trials demonstrate predictive biomarkers for dovitinib, an FGFR inhibitor, in lung squamous cell carcinoma. Ann Oncol. 2017;28(6):1250–9.
Serebrenik AA, Argyris PP, Jarvis MC, Brown WL, Bazzaro M, Vogel RI, et al. The DNA cytosine deaminase APOBEC3B is a molecular determinant of platinum responsiveness in clear cell ovarian cancer. Clin Cancer Res. 2020;26(13):3397–407.
Palmer AC, Plana D, Gao H, Korn JM, Yang G, Green J, et al. A proof of concept for biomarker-guided targeted therapy against ovarian cancer based on patient-derived tumor xenografts. Cancer Res. 2020;80(19):4278–87.
Morton JJ, Alzofon N, Jimeno A. The humanized mouse: emerging translational potential. Mol Carcinog. 2020;59(7):830–8.
Hylander BL, Punt N, Tang H, Hillman J, Vaughan M, Bshara W, et al. Origin of the vasculature supporting growth of primary patient tumor xenografts. J Transl Med. 2013;11:110.
Ben-David U, Ha G, Tseng YY, Greenwald NF, Oh C, Shih J, et al. Patient-derived xenografts undergo mouse-specific tumor evolution. Nat Genet. 2017;49(11):1567–75.
Blomme A, Van Simaeys G, Doumont G, Costanza B, Bellier J, Otaka Y, et al. Murine stroma adopts a human-like metabolic phenotype in the PDX model of colorectal cancer and liver metastases. Oncogene. 2018;37(9):1237–50.
Greaves M, Maley CC. Clonal evolution in cancer. Nature. 2012;481(7381):306–13.
Isella C, Brundu F, Bellomo SE, Galimi F, Zanella E, Porporato R, et al. Selective analysis of cancer-cell intrinsic transcriptional traits defines novel clinically relevant subtypes of colorectal cancer. Nat Commun. 2017;8:15107.
Sato T, Vries RG, Snippert HJ, van de Wetering M, Barker N, Stange DE, et al. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature. 2009;459(7244):262–5.
Yoshida GJ. Applications of patient-derived tumor xenograft models and tumor organoids. J Hematol Oncol. 2020;13(1):4.
Drost J, Clevers H. Organoids in cancer research. Nat Rev Cancer. 2018;18(7):407–18.
Grönholm M, Feodoroff M, Antignani G, Martins B, Hamdan F, Cerullo V. Patient-derived organoids for precision cancer immunotherapy. Cancer Res. 2021;81(12):3149–55.
Prasetyanti PR, Medema JP. Intra-tumor heterogeneity from a cancer stem cell perspective. Mol Cancer. 2017;16(1):41.
Rossi G, Manfrin A, Lutolf MP. Progress and potential in organoid research. Nat Rev Genet. 2018;19(11):671–87.
Lau HCH, Kranenburg O, Xiao H, Yu J. Organoid models of gastrointestinal cancers in basic and translational research. Nat Rev Gastroenterol Hepatol. 2020;17(4):203–22.
Jacob F, Salinas RD, Zhang DY, Nguyen PTT, Schnoll JG, Wong SZH, et al. A patient-derived glioblastoma organoid model and biobank recapitulates inter- and intra-tumoral heterogeneity. Cell. 2020;180(1):188–204.e22.
Fujii M, Shimokawa M, Date S, Takano A, Matano M, Nanki K, et al. A colorectal tumor organoid library demonstrates progressive loss of niche factor requirements during tumorigenesis. Cell Stem Cell. 2016;18(6):827–38.
Boj SF, Hwang CI, Baker LA, Chio II, Engle DD, Corbo V, et al. Organoid models of human and mouse ductal pancreatic cancer. Cell. 2015;160(1–2):324–38.
Gao D, Vela I, Sboner A, Iaquinta PJ, Karthaus WR, Gopalan A, et al. Organoid cultures derived from patients with advanced prostate cancer. Cell. 2014;159(1):176–87.
Hill SJ, Decker B, Roberts EA, Horowitz NS, Muto MG, Worley MJ Jr, et al. Prediction of DNA repair inhibitor response in short-term patient-derived ovarian cancer organoids. Cancer Discov. 2018;8(11):1404–21.
Kopper O, de Witte CJ, Lõhmussaar K, Valle-Inclan JE, Hami N, Kester L, et al. An organoid platform for ovarian cancer captures intra- and interpatient heterogeneity. Nat Med. 2019;25(5):838–49.
Tao M, Sun F, Wang J, Wang Y, Zhu H, Chen M, et al. Developing patient-derived organoids to predict PARP inhibitor response and explore resistance overcoming strategies in ovarian cancer. Pharmacol Res. 2022;179:106232.
Yucer N, Ahdoot R, Workman MJ, Laperle AH, Recouvreux MS, Kurowski K, et al. Human iPSC-derived fallopian tube organoids with BRCA1 mutation recapitulate early-stage carcinogenesis. Cell Rep. 2021;37(13):110146.
Bi J, Newtson AM, Zhang Y, Devor EJ, Samuelson MI, Thiel KW, et al. Successful patient-derived organoid culture of gynecologic cancers for disease modeling and drug sensitivity testing. Cancers (Basel). 2021;13(12):2901.
Nanki Y, Chiyoda T, Hirasawa A, Ookubo A, Itoh M, Ueno M, et al. Patient-derived ovarian cancer organoids capture the genomic profiles of primary tumours applicable for drug sensitivity and resistance testing. Sci Rep. 2020;10(1):12581.
de Witte CJ, Espejo Valle-Inclan J, Hami N, Lõhmussaar K, Kopper O, Vreuls CPH, et al. Patient-derived ovarian cancer organoids mimic clinical response and exhibit heterogeneous inter- and Intrapatient drug responses. Cell Rep. 2020;31(11):107762.
Chen H, Gotimer K, De Souza C, Tepper CG, Karnezis AN, Leiserowitz GS, et al. Short-term organoid culture for drug sensitivity testing of high-grade serous carcinoma. Gynecol Oncol. 2020;157(3):783–92.
Maenhoudt N, Defraye C, Boretto M, Jan Z, Heremans R, Boeckx B, et al. Developing organoids from ovarian Cancer as experimental and preclinical models. Stem Cell Reports. 2020;14(4):717–29.
Hoffmann K, Berger H, Kulbe H, Thillainadarasan S, Mollenkopf HJ, Zemojtel T, et al. Stable expansion of high-grade serous ovarian cancer organoids requires a low-Wnt environment. EMBO J. 2020;39(6):e104013.
Maru Y, Tanaka N, Itami M, Hippo Y. Efficient use of patient-derived organoids as a preclinical model for gynecologic tumors. Gynecol Oncol. 2019;154(1):189–98.
Wulftange WJ, Rose MA, Garmendia-Cedillos M, da Silva D, Poprawski JE, Srinivasachar D, et al. Spatial control of oxygen delivery to three-dimensional cultures alters cancer cell growth and gene expression. J Cell Physiol. 2019;234(11):20608–22.
Loessner D, Stok KS, Lutolf MP, Hutmacher DW, Clements JA, Rizzi SC. Bioengineered 3D platform to explore cell-ECM interactions and drug resistance of epithelial ovarian cancer cells. Biomaterials. 2010;31(32):8494–506.
Jabs J, Zickgraf FM, Park J, Wagner S, Jiang X, Jechow K, et al. Screening drug effects in patient-derived cancer cells links organoid responses to genome alterations. Mol Syst Biol. 2017;13(11):955.
Fan H, Demirci U, Chen P. Emerging organoid models: leaping forward in cancer research. J Hematol Oncol. 2019;12(1):142.
Kessler M, Hoffmann K, Brinkmann V, Thieck O, Jackisch S, Toelle B, et al. The notch and Wnt pathways regulate stemness and differentiation in human fallopian tube organoids. Nat Commun. 2015;6:8989.
LeSavage BL, Suhar RA, Broguiere N, Lutolf MP, Heilshorn SC. Next-generation cancer organoids. Nat Mater. 2022;21(2):143–59.
Bleijs M, van de Wetering M, Clevers H, Drost J. Xenograft and organoid model systems in cancer research. EMBO J. 2019;38(15):e101654.
Driehuis E, Kretzschmar K, Clevers H. Establishment of patient-derived cancer organoids for drug-screening applications. Nat Protoc. 2020;15(10):3380–409.
van de Merbel AF, van der Horst G, van der Pluijm G. Patient-derived tumour models for personalized therapeutics in urological cancers. Nat Rev Urol. 2021;18(1):33–45.
Lee SH, Hu W, Matulay JT, Silva MV, Owczarek TB, Kim K, et al. Tumor evolution and drug response in patient-derived organoid models of bladder cancer. Cell. 2018;173(2):515–28.e17.
Liu HD, Xia BR, Jin MZ, Lou G. Organoid of ovarian cancer: genomic analysis and drug screening. Clin Transl Oncol. 2020;22(8):1240–51.
Yuki K, Cheng N, Nakano M, Kuo CJ. Organoid models of tumor immunology. Trends Immunol. 2020;41(8):652–64.
Neal JT, Li X, Zhu J, Giangarra V, Grzeskowiak CL, Ju J, et al. Organoid modeling of the tumor immune microenvironment. Cell. 2018;175(7):1972–88.e16.
Wan C, Keany MP, Dong H, Al-Alem LF, Pandya UM, Lazo S, et al. Enhanced efficacy of simultaneous PD-1 and PD-L1 immune checkpoint blockade in high-grade serous ovarian cancer. Cancer Res. 2021;81(1):158–73.
Xu R, Zhou X, Wang S, Trinkle C. Tumor organoid models in precision medicine and investigating cancer-stromal interactions. Pharmacol Ther. 2021;218:107668.
Wimmer RA, Leopoldi A, Aichinger M, Wick N, Hantusch B, Novatchkova M, et al. Human blood vessel organoids as a model of diabetic vasculopathy. Nature. 2019;565(7740):505–10.
Chang YH, Chu TY, Ding DC. Human fallopian tube epithelial cells exhibit stemness features, self-renewal capacity, and Wnt-related organoid formation. J Biomed Sci. 2020;27(1):32.
Qian J, LeSavage BL, Hubka KM, Ma C, Natarajan S, Eggold JT, et al. Cancer-associated mesothelial cells promote ovarian cancer chemoresistance through paracrine osteopontin signaling. J Clin Invest. 2021;131(16):e146186.
Li X, Nadauld L, Ootani A, Corney DC, Pai RK, Gevaert O, et al. Oncogenic transformation of diverse gastrointestinal tissues in primary organoid culture. Nat Med. 2014;20(7):769–77.
Drost J, van Boxtel R, Blokzijl F, Mizutani T, Sasaki N, Sasselli V, et al. Use of CRISPR-modified human stem cell organoids to study the origin of mutational signatures in cancer. Science. 2017;358(6360):234–8.
Zhang S, Iyer S, Ran H, Dolgalev I, Gu S, Wei W, et al. Genetically defined, syngeneic organoid platform for developing combination therapies for ovarian cancer. Cancer Discov. 2021;11(2):362–83.
Maenhoudt N, Vankelecom H. Protocol for establishing organoids from human ovarian cancer biopsies. STAR Protoc. 2021;2(2):100429.
Powley IR, Patel M, Miles G, Pringle H, Howells L, Thomas A, et al. Patient-derived explants (PDEs) as a powerful preclinical platform for anti-cancer drug and biomarker discovery. Br J Cancer. 2020;122(6):735–44.
Li Y, Ding Z, Deng L, Fan G, Zhang Q, Gong H, et al. Precision vibratome for high-speed ultrathin biotissue cutting and organ-wide imaging. iScience. 2021;24(9):103016.
Parajuli N, Doppler W. Precision-cut slice cultures of tumors from MMTV-neu mice for the study of the ex vivo response to cytokines and cytotoxic drugs. In Vitro Cell Dev Biol Anim. 2009;45(8):442–50.
Naipal KA, Verkaik NS, Sánchez H, van Deurzen CH, den Bakker MA, Hoeijmakers JH, et al. Tumor slice culture system to assess drug response of primary breast cancer. BMC Cancer. 2016;16:78.
Gerlach MM, Merz F, Wichmann G, Kubick C, Wittekind C, Lordick F, et al. Slice cultures from head and neck squamous cell carcinoma: a novel test system for drug susceptibility and mechanisms of resistance. Br J Cancer. 2014;110(2):479–88.
Abreu S, Silva F, Mendes R, Mendes TF, Teixeira M, Santo VE, et al. Patient-derived ovarian cancer explants: preserved viability and histopathological features in long-term agitation-based cultures. Sci Rep. 2020;10(1):19462.
Templeton AR, Jeffery PL, Thomas PB, Perera MPJ, Ng G, Calabrese AR, et al. Patient-derived explants as a precision medicine patient-proximal testing platform informing Cancer management. Front Oncol. 2021;11:767697.
Hanahan D, Wagner EF, Palmiter RD. The origins of oncomice: a history of the first transgenic mice genetically engineered to develop cancer. Genes Dev. 2007;21(18):2258–70.
Kersten K, de Visser KE, van Miltenburg MH, Jonkers J. Genetically engineered mouse models in oncology research and cancer medicine. EMBO Mol Med. 2017;9(2):137–53.
Politi K, Pao W. How genetically engineered mouse tumor models provide insights into human cancers. J Clin Oncol. 2011;29(16):2273–81.
Howell VM. Genetically engineered mouse models for epithelial ovarian cancer: are we there yet? Semin Cell Dev Biol. 2014;27:106–17.
Lynch HT, Casey MJ, Snyder CL, Bewtra C, Lynch JF, Butts M, et al. Hereditary ovarian carcinoma: heterogeneity, molecular genetics, pathology, and management. Mol Oncol. 2009;3(2):97–137.
Teng K, Ford MJ, Harwalkar K, Li Y, Pacis AS, Farnell D, et al. Modeling high-grade serous ovarian carcinoma using a combination of in vivo fallopian tube electroporation and CRISPR-Cas9-mediated genome editing. Cancer Res. 2021;81(20):5147–60.
Shi M, Whorton AE, Sekulovski N, Paquet M, MacLean JA, Song Y, et al. Inactivation of TRP53, PTEN, RB1, and/or CDH1 in the ovarian surface epithelium induces ovarian cancer transformation and metastasis. Biol Reprod. 2020;102(5):1055–64.
Zhai Y, Wu R, Kuick R, Sessine MS, Schulman S, Green M, et al. High-grade serous carcinomas arise in the mouse oviduct via defects linked to the human disease. J Pathol. 2017;243(1):16–25.
Wu R, Zhai Y, Kuick R, Karnezis AN, Garcia P, Naseem A, et al. Impact of oviductal versus ovarian epithelial cell of origin on ovarian endometrioid carcinoma phenotype in the mouse. J Pathol. 2016;240(3):341–51.
Zhai Y, Kuick R, Tipton C, Wu R, Sessine M, Wang Z, et al. Arid1a inactivation in an Apc- and Pten-defective mouse ovarian cancer model enhances epithelial differentiation and prolongs survival. J Pathol. 2016;238(1):21–30.
Ren YA, Mullany LK, Liu Z, Herron AJ, Wong KK, Richards JS. Mutant p53 promotes epithelial ovarian Cancer by regulating tumor differentiation, metastasis, and responsiveness to steroid hormones. Cancer Res. 2016;76(8):2206–18.
van der Horst PH, van der Zee M, Heijmans-Antonissen C, Jia Y, DeMayo FJ, Lydon JP, et al. A mouse model for endometrioid ovarian cancer arising from the distal oviduct. Int J Cancer. 2014;135(5):1028–37.
Perets R, Wyant GA, Muto KW, Bijron JG, Poole BB, Chin KT, et al. Transformation of the fallopian tube secretory epithelium leads to high-grade serous ovarian cancer in Brca;Tp53;Pten models. Cancer Cell. 2013;24(6):751–65.
Szabova L, Yin C, Bupp S, Guerin TM, Schlomer JJ, Householder DB, et al. Perturbation of Rb, p53, and Brca1 or Brca2 cooperate in inducing metastatic serous epithelial ovarian cancer. Cancer Res. 2012;72(16):4141–53.
Kim J, Coffey DM, Creighton CJ, Yu Z, Hawkins SM, Matzuk MM. High-grade serous ovarian cancer arises from fallopian tube in a mouse model. Proc Natl Acad Sci U S A. 2012;109(10):3921–6.
Kinross KM, Montgomery KG, Kleinschmidt M, Waring P, Ivetac I, Tikoo A, et al. An activating Pik3ca mutation coupled with Pten loss is sufficient to initiate ovarian tumorigenesis in mice. J Clin Invest. 2012;122(2):553–7.
Mullany LK, Fan HY, Liu Z, White LD, Marshall A, Gunaratne P, et al. Molecular and functional characteristics of ovarian surface epithelial cells transformed by KrasG12D and loss of Pten in a mouse model in vivo. Oncogene. 2011;30(32):3522–36.
Xing D, Orsulic S. A mouse model for the molecular characterization of brca1-associated ovarian carcinoma. Cancer Res. 2006;66(18):8949–53.
Dinulescu DM, Ince TA, Quade BJ, Shafer SA, Crowley D, Jacks T. Role of K-ras and Pten in the development of mouse models of endometriosis and endometrioid ovarian cancer. Nat Med. 2005;11(1):63–70.
Flesken-Nikitin A, Choi KC, Eng JP, Shmidt EN, Nikitin AY. Induction of carcinogenesis by concurrent inactivation of p53 and Rb1 in the mouse ovarian surface epithelium. Cancer Res. 2003;63(13):3459–63.
Orsulic S, Li Y, Soslow RA, Vitale-Cross LA, Gutkind JS, Varmus HE. Induction of ovarian cancer by defined multiple genetic changes in a mouse model system. Cancer Cell. 2002;1(1):53–62.
Connolly DC, Bao R, Nikitin AY, Stephens KC, Poole TW, Hua X, et al. Female mice chimeric for expression of the simian virus 40 TAg under control of the MISIIR promoter develop epithelial ovarian cancer. Cancer Res. 2003;63(6):1389–97.
Ahuja D, Sáenz-Robles MT, Pipas JM. SV40 large T antigen targets multiple cellular pathways to elicit cellular transformation. Oncogene. 2005;24(52):7729–45.
Quinn BA, Xiao F, Bickel L, Martin L, Hua X, Klein-Szanto A, et al. Development of a syngeneic mouse model of epithelial ovarian cancer. J Ovarian Res. 2010;3:24.
Branda CS, Dymecki SM. Talking about a revolution: the impact of site-specific recombinases on genetic analyses in mice. Dev Cell. 2004;6(1):7–28.
Karakashev S, Zhang RG. Mouse models of epithelial ovarian cancer for preclinical studies. Zool Res. 2021;42(2):153–60.
Sutherland KD, Proost N, Brouns I, Adriaensen D, Song JY, Berns A. Cell of origin of small cell lung cancer: inactivation of Trp53 and Rb1 in distinct cell types of adult mouse lung. Cancer Cell. 2011;19(6):754–64.
Molyneux G, Geyer FC, Magnay FA, McCarthy A, Kendrick H, Natrajan R, et al. BRCA1 basal-like breast cancers originate from luminal epithelial progenitors and not from basal stem cells. Cell Stem Cell. 2010;7(3):403–17.
Van Keymeulen A, Lee MY, Ousset M, Brohée S, Rorive S, Giraddi RR, et al. Reactivation of multipotency by oncogenic PIK3CA induces breast tumour heterogeneity. Nature. 2015;525(7567):119–23.
Flesken-Nikitin A, Hwang CI, Cheng CY, Michurina TV, Enikolopov G, Nikitin AY. Ovarian surface epithelium at the junction area contains a cancer-prone stem cell niche. Nature. 2013;495(7440):241–5.
Zhang S, Dolgalev I, Zhang T, Ran H, Levine DA, Neel BG. Both fallopian tube and ovarian surface epithelium are cells-of-origin for high-grade serous ovarian carcinoma. Nat Commun. 2019;10(1):5367.
Acknowledgments
Not applicable.
Funding
Fundings of Gang Chen.
The Nature and Science Foundation of China (81874106, 82073259), and the Key R&D Program of Hubei Province (2020BCA067).
Fundings of Chaoyang Sun.
The Nature and Science Foundation of China (81974408), and The Hubei Province Science Fund for Distinguished Young Scholars (2020CFA066), and Beijing Kanghua Foundation for the Development of Traditional Chinese and Western Medicine (KH-2021-LQJJ-006).
Author information
Authors and Affiliations
Contributions
T.Q. developed the concepts; T.Q., J. F performed literature searching; T.Q., F.L. and L. Z analyzed selected papers; T.Q., J. F wrote the manuscripts; C.L. and Q.X. sorted the data; T.Q. and Y.Z. realized the figs. G.C. and C.S. supervised and coordinated the whole process. All athuors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
This study was approved by the Ethics or Institutional Review Board of Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, China.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Qin, T., Fan, J., Lu, F. et al. Harnessing preclinical models for the interrogation of ovarian cancer. J Exp Clin Cancer Res 41, 277 (2022). https://doi.org/10.1186/s13046-022-02486-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13046-022-02486-z