
Abstract
Background
A frequent complication of Fontan operations is unilateral diaphragmatic paresis, which leads to hemodynamic deterioration of the Fontan circulation. A potential new therapeutic option is the unilateral diaphragmatic pacemaker. In this study, we investigated the most effective stimulation location for a potential fully implantable system in a porcine model.
Methods
Five pigs (20.8 ± 0.95 kg) underwent implantation of a customized cuff electrode placed around the right phrenic nerve. A bipolar myocardial pacing electrode was sutured adjacent to the motor point and peripherally at the costophrenic angle (peripheral diaphragmatic muscle). The electrodes were stimulated 30 times per minute with a pulse duration of 200 µs and a stimulation time of 300 ms. Current intensity was the only variable changed during the experiment.
Results
Effective stimulation occurred at 0.26 ± 0.024 mA at the phrenic nerve and 7 ± 1.22 mA at the motor point, a significant difference in amperage (p = 0.005). Even with a maximum stimulation of 10 mA at the peripheral diaphragm muscle, however, no effective stimulation was observed.
Conclusion
The phrenic nerve seems to be the best location for direct stimulation by a unilateral thoracic diaphragm pacemaker in terms of the required amperage level in a porcine model.
Similar content being viewed by others

Introduction
The diaphragmatic pacemaker is a therapeutic option for the treatment of diaphragmatic paresis and used for bilateral paresis or dysfunction. This device is indicated for patients with high-level spinal cord injuries or central hypoventilation syndrome [1]. Two versions of the pacemaker are in use, one that provides direct bilateral stimulation of the phrenic nerve [2, 3] and one that provides bilateral stimulation of the abdominal motor point [4]. Both systems stimulate the diaphragm via external stimulation without triggering. A fully implantable pacemaker for unilateral diaphragmatic paresis that stimulates the paretic side by triggering the healthy side is not currently available, however. Unilateral diaphragmatic paresis is a known complication with Fontan surgery [5,6,7,8,9] and has an important influence on outcomes [10, 11].
The Fontan pathway is a multi-stage treatment performed on children with univentricular circulation. With the connection of the superior and inferior venae cavae to the pulmonary arteries, the deoxygenated blood flows passively into the pulmonary arteries without a sub-pulmonary ventricle [12]. Negative intrathoracic pressure is an important factor supporting passive pulmonary blood flow and is generated within the closed chest by diaphragmatic excursion. The importance of spontaneous breathing in patients with Fontan circulation was first demonstrated by Redington and Penny. Their report that intrathoracic negative pressure generated in the lungs during inspiration increases antegrade blood flow in the pulmonary arteries [13, 14] was later confirmed in MRI studies [15, 16].
Usually, symptomatic Fontan patients with diaphragmatic palsy are treated with diaphragmatic plication, which unfortunately does not yield hemodynamic improvement in the Fontan circulation [17]. Diaphragmatic plication thus does not seem to be an optimal treatment, and its effect on improving pulmonary blood flow has not been consistently demonstrated, especially in patients with univentricular heart and diaphragmatic palsy.
We are developing a triggered unilateral diaphragmatic pacing system for use as a pacemaker, especially in children after Fontan surgery. Our goal is to improve the reduced pulmonary blood flow in the paretic side by diaphragmatic pacing.
For such a pacemaker, an accelerometer may be utilized as a potential device for triggering [18]. Based on our animal model [19], in the current experiment, we are going to identify which stimulation location is the best target for a unilateral diaphragm pacemaker in our porcine model. For this purpose we compared the peripheral diaphragm muscle, phrenic nerve, and nerve entry point into the diaphragm (also called the motor point [20]) in terms of the amperage required to generate sufficient stimulation. Additionally, we performed the experiment with a transected phrenic nerve to exclude the influence of spontaneous breathing.
Although several stimulus thresholds have already been determined in humans [1, 21, 22] and mongrel dogs [23, 24] an evaluation in the porcine model does not exist yet. Although the porcine model represents one of the most frequently used large animal models in Europe at present [25]. Hence, it seems to be of major interest to implement such an examination in order to demonstrate the transferability of our porcine model’s data to human patients.
Material and methods
Experiments were conducted in accordance with the German Animal Welfare Act and its subsequent statutory acts, which are in accordance with the Council of Europe Convention ETS 123. The competent state agency, the State Office for Nature, Environment and Consumer Protection North Rhine-Westphalia, approved the study (permit: 81–02.04.2020.A392). Our study complied with the Animal Research: Reporting of In Vivo Experiments guidelines 2.0.
Animal preparation and instrumentation
Five pigs were examined and had a mean weight of 20.8 ± 0.95 kg. Pigs were supplied in-house by the Agriculture Faculty, Rheinische Friedrich-Wilhelms-University Bonn, Königswinter-Vinxel, Germany, were of conventional microbiologic status, and had an acclimatization period of 3 days at our facility.
Premedication consisted of intramuscularly applied ketamine (20 mg/kg; WDT, Garbsen, Germany) in combination with azaperone (2 mg/kg; Richter Pharma, Wels, Austria) and atropine (0.02 mg/kg; B. Braun, Melsungen, Germany). After adequate sedation was achieved, venous access was implemented with a 1.1 mm outer diameter Jelco® catheter (Smith Medical, Grasbrunn, Germany) in an ear vein. Anesthesia induction consisted of piritramide (0.5 mg/kg; Hameln Pharma, Hameln, Germany) and propofol (10 mg/kg; CP Pharma, Burgdorf, Germany). The administration of muscle relaxants was explicitly avoided in this experiment. We secured the airway via endotracheal intubation using a straight size 4 Miller blade with a 4.5 mm internal diameter curved, microcuffed endotracheal tube (Avanos, Hamburg, Germany). For invasive ventilation, we used a Servo-i (Maquet, Rastatt, Germany) with synchronized intermittent mandatory ventilation with a frequency of around 15 breaths per minute, an inspiratory pressure of 15 cmH2O, and a positive end-expiratory pressure of 5 cmH2O. The animals were ventilated on room air (FiO2 = 0.21). We used a flow trigger mode set to > 1 l/min. Anesthesia was maintained with continuous intravenous infusion of propofol (1–5 mg/kg/h) via a Perfusor® Space (B. Braun, Melsungen, Germany) and continuous intravenous infusion of piritramide (0.2–0.5 mg/kg), supplemented by occasional single doses of ketamine (5–10 mg/kg/h) or midazolam (0.5 mg/kg; B. Braun, Melsungen, Germany). We monitored the depth of total intravenous anesthesia with the Narcotrend system via a Compact M monitor for intraoperative use (Narcotrend, Hannover, Germany) using needle electrodes (Neuroline Twisted Pair Subdermal, 12 × 0.4 mm, Ambu, Ballerup, Denmark) placed at the standard positions according to the manufacturer’s instructions. During the procedure, the animals were in electroencephalographic Kugler stage D0 [26], which is equivalent to general anesthesia.
We monitored the pigs using an Infinity C500 monitor (Dräger, Lübeck, Germany) for electrocardiogram, pulse oximetry, and invasive blood pressure. For capnometry, we used a Datex-Ohmeda S/5 (Datex-Ohmeda, Duisburg, Germany). We placed an arterial line (2.7 French leadercath, Vygon, Aachen, Germany) in the right femoral artery for continuous blood pressure monitoring and a three-lumen central line (5.5 French, Teleflex, Fellbach, Germany) in the right femoral vein. In addition, we placed a 10 Charrière transurethral catheter (Asid Bonz, Herrenberg, Germany) to monitor urine output. Temperature was measured with a 9 French rectal probe (Smiths Medical, Grasbrunn, Germany). Maintenance fluids were infused at a rate of 50–70 mL/kg/h using Ionosteril 1/1 (Fresenius Kabi, Bad Homburg, Germany). At the end of the experiment, animals were euthanized using T61® (tetracaine/mebezonium/embutramide) (Intervet, München, Germany) at a dose of 0.5 mL/kg.
Intervention set-up
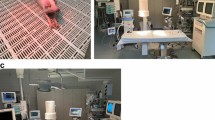
After initial supine positioning, the pigs were prepared and instrumented as described above. A median sternotomy was performed, the pleura opened, and the right diaphragm exposed, along with the phrenic nerve in continuation. The electrodes were then implanted using 5–0 polypropylene sutures (Ethicon, Norderstedt, Germany). A custom-made bipolar cuff electrode built by the Fraunhofer Institute for Biomedical Engineering (Sulzbach, Germany) was placed around the right phrenic nerve (Fig. 1A). A bipolar myocardial pacing electrode, TME (Osypka, Rheinfeld, Germany), was implanted adjacent to the motor point (Fig. 1B) and peripherally on the diaphragmatic muscle at the costo-diaphragmatic angle. The motor point was identified visually. The electrodes were sutured on each side of the nerve’s entry point into the diaphragm. The two poles of the electrode each were implanted at a distance of 1 cm. After the instrumentation the animals were allowed to recover for a period of 30 to 45 min. In this recovery period a bolus (10 ml/ kg bodyweight) of Ionosteril 1/1 (Fresenius Kabi, Bad Homburg, Germany) was administered closely observing the central venous pressure (target: 5 to 10 mmHg). Ventilation was adjusted in order to provide a balanced acid–base state. The sternal retractor was removed, and the chest was left open. The incision was covered by a moisted swap.

A Cuff electrode placement around the phrenic nerve (●). B Bipolar TME electrode implantation at the motor point (▲); → Indicates the phrenic nerve. C Cuff electrode
The electrodes were then stimulated 30 times per minute with a pulse duration of 200 µs and a stimulation time of 300 ms. Only current intensity was varied. The stimulation was carried out using a FE180 (ADInstruments, Sydney, Australia) and ISIS Neurostimulator (inomed, Emmendingen, Germany). The stimulation unit was placed on a separate mount extracorporeally. For each amperage, stimulation was performed for at least for 5 min.
Stimulation was conducted with 0.1, 0.2, 0.3, and 0.5 mA at the phrenic nerve, with 0.1, 1, 3, 5, and 10 mA at the motor point, and with 5 and 10 mA at the peripheral diaphragm muscle. The recording of lower stimulation currents at the peripheral diaphragm muscle was not performed, as no effective stimulation could be achieved. The same series of stimulations was carried out after the phrenic nerve had been transected approximately 2 cm above the cuff electrode.
Measurement
All measurements were taken at baseline without phrenic nerve stimulation with the intact phrenic nerve and at the different stimulation levels still with the phrenic nerve being intact. Thereafter, the phrenic nerve was transected, and the same varying stimulation levels were applied. Diaphragmatic movement was recorded by fluoroscopy (Ziehm Vista, Ziehm, Nuremberg, Germany) and evaluated with a tangent drawn at the highest point of the right hemi-diaphragm. At this point, diaphragmatic movement was measured at a right angle in millimeters. Arterial blood gas analyses were conducted in a RapidPoint® 500 analyzer (Siemens Healthineers, Erlangen, Germany). Ventilation parameters were measured using Servo-i (Maquet, Rastatt, Germany), and baseline ventilation settings were an inspiratory pressure of 15 cmH2O, a positive end-expiratory pressure of 5 cmH2O, and a ventilation rate of 15/min.
Statistics
Data are presented as mean ± SEM. The statistical significance of changes from baseline values within each parameter was tested with ANOVA for repeated measures. Differences between the various stimulation locations were analyzed by one-way ANOVA comparing the three stimulation locations.
Diaphragmatic excursion was defined as complete downward motion of the entire diaphragmatic plane. The threshold was detected at the three locations, and threshold mean values from all animals at the individual locations were compared for identifying the best location for diaphragmatic stimulation. Normal distribution was tested by the Shapiro–Wilk test. For nonparametric values, the Kruskal–Wallis test was used. Differences between the intact and the transected states were compared with independent t-tests. Statistical significance was accepted at p ≤ 0.05 after pairwise testing. Statistical analyses were performed with IBM SPSS Statistics Version 28.
Results
At baseline, mean values were as follows: pH, 7.44 ± 0.02; respiratory minute volume, 2.76 ± 0.18 l/min; pO2, 71.8 ± 2.01 mmHg; end-tidal CO2, 38.2 ± 0.97 mmHg; breathing rate, 15/min; and diaphragmatic excursion, 4.2 ± 0.3 mm.
Stimulation
Phrenic nerve
Compared with the above baseline values, direct stimulation at a threshold of 0.2 mA resulted in significantly increased respiratory minute volume to 4.73 ± 0.62 L/min (p = 0.019) and breathing rate to 25.93 ± 2.73/min (p = 0.05). At 0.3 mA, compared with baseline, there were significant changes in end-tidal CO2 (to 30.4 ± 0.75 mmHg; p = 0.016) and diaphragm excursion (to 9.3 ± 1 mm; p = 0.003). Change from baseline pO2 was not seen until application of 0.5 mA (to 85.64 ± 3.24 mmHg; p = 0.039), and no change in pH was detected at any level (Figs. 2 and 3).

A Respiratory minute volume (L/min), B Breathing rate (per min) and C Diaphragm excursion in relation to stimulation of the phrenic nevre (n = 5; ns: not siginificant; *p<0.05, **p<0.01, ***p<0.001)

A pO2, B End-tidal CO2, and C pH in relation to stimulation of the phrenice nerve (n =5; ns:not significant; *p <0.05, **p <0.01, ***p <0.001)
After the right phrenic nerve was transected, compared with baseline, there were significant changes in respiratory minute volume (4.86 ± 0.66 L/min; p = 0.024) and breathing rate (27.07 ± 2.08/min; p = 0.02) at 0.2 mA. At 0.3 mA, changes from baseline were seen for end-tidal CO2 (31.2 ± 1.28 mmHg; p = 0.003), diaphragm excursion (7.7 ± 0.95 mm; p = 0.032), and pO2 (84.88 ± 3.53 mmHg; p = 0.042). Values for pH remained unchanged (Figs. 2 and 3).
We found no significant difference in mean values between intact and transected phrenic nerves (Figs. 2 and 3).
Motor point
Motor point stimulation with the phrenic nerve intact differed significantly from baseline at 5 mA for respiratory minute volume (3.9 ± 0.30 L/min; p = 0.017), breathing rate (23.5 ± 3.2/min; p = 0.009), and diaphragm excursion (6.3 ± 1 mm; p = 0.01). Significant changes from baseline were observed for end-tidal CO2 (33.6 ± 1.81 mmHg; p = 0.046) and pO2 (84.3 ± 3.48 mmHg; p = 0.045) with 10 mA. No significant difference could be seen for pH as a function of the different amperage levels (Figs. 4 and 5).

A Respiratory minute volume (L/min), B Breathing rate (per min) and C Diaphragm excursion in relation to stimulation of the motor point (n= 5; ns: not significant; *p<0.05, **p<0.01, ***p<0.001)

A pO2, B End-tidal CO2 and C pH in relation to stimulation of the motor point (n= 5; ns: not significant; *p<0.05, **p<0.01, ***p<0.001)
After transection of the phrenic nerve, stimulation of the motor point yielded no difference from baseline for end-tidal CO2, pO2, pH, or diaphragm excursion. For breathing rate, a significant change was noted at 3 mA (to 17.46 ± 0.76/min; p = 0.026), as was also the case for respiratory minute volume (to 3.5 ± 0.13 L/min; p = 0.044) at 5 mA.
Mean values between intact and transected nerves during stimulation of the motor point differed no significant difference could be observed between the two groups (Figs. 4 and 5).
Peripheral diaphragm muscle
Although a local contraction in the vicinity of the electrodes could be visually perceived, complete diaphragmatic contraction was not achieved. However, maximum stimulation (10 mA) of the peripheral diaphragmatic muscle with the nerve intact yielded a significant change in breathing rate (18.53 ± 3.57/min; p = 0.026) and diaphragm excursion (5.32 ± 0.50 mm; p = 0.028). After transection of the nerve, there was a significant change in breathing rate only with 10 mA (20 ± 5.9/min; p = 0.019). At 5 mA, we also observed a significant deterioration in pO2 (60.62 ± 8.13 mmHg; p = 0.027) from baseline (71.8. ± 4.5 mmHg).
Comparison of mean values at the peripheral diaphragm muscle with the intact and transected phrenic nerve showed no significant difference (Figs. 6 and 7).

A Respiratory minute volume (L/min), B Breathing rate (per min) and C Diaphragm excursion in relation to stimulation of the peripheral diaphragm muscle (n=5; ns:not significant; *p<0.05, **p<0.01, ***p<0.001)

A pO2, B End-tidal CO2 and C pH in relation to stimulation of the peripheral diaphragm muscle (n=5; ns:not significant; *p<0.05, **p<0.01, ***p<0.001)
Comparison of the three stimulation locations
The comparison between motor point and phrenic nerve stimulation showed a significant difference, with much less amperage required at the phrenic nerve to initiate diaphragmatic contraction. An additional comparison with stimulation of the peripheral diaphragm muscle was excluded, as only a local contraction was visually detectable. On average, direct phrenic nerve stimulation resulted in diaphragmatic contraction at 0.26 ± 0.02 mA, compared with the significantly higher amperage of 7 ± 1.22 mA being required at the motor point (p = 0.005).
Discussion
The current results show that for diaphragmatic contraction in the porcine model, stimulation directly at the phrenic nerve requires less amperage compared with the motor point and peripheral diaphragmatic muscle stimulation via a thoracic approach. Even with high amperages, stimulation of the peripheral diaphragm leads to inefficient contraction. Hence, direct phrenic nerve stimulation provides the most efficient pacing site, especially in the context of developing a fully implantable system to support a patient over the long term with a reduced need for battery changes. These results accord to the known studies on humans [1, 22] and mongrel dogs [24]. This suggests that our prospective device of a unilateral diaphragmatic pacemaker in porcine model is transferable to human patients in terms of the best target for stimulation.
Stimulation of intact versus transected nerves did not result in any relevant differences in amperage required. No significant change in pO2, pCO2, pH, respiratory minute volume, respiratory rate and diaphragmatic excursion was observed between the intact and transected phrenic nerve, independent of the location of stimulation. So, during phrenic nerve stimulation the effect of short-term phrenic nerve transection on all observed parameters appears to be minute. The acute loss of central nervous control therefore does not seem to have any significance for the amperage level required. In the setting of congenital cardiac surgery, clinically relevant unilateral diaphragmatic paresis usually occurs intraoperatively and can be treated promptly by diaphragmatic pacemaker implantation. In further studies, we plan to investigate how long a damaged phrenic nerve can still be effectively stimulated and how early phrenic nerve stimulation affects prevention of nerve dystrophy after nerve trauma.
In this study, the most commonly used stimulation locations were compared for the first time in the same setting [4, 24, 27]. In agreement with Bilgutay et al., we demonstrated the superiority of direct phrenic nerve stimulation compared with diaphragmatic pacing via the motor point or the even worse-performing peripheral diaphragmatic site. These data support diaphragmatic pacing via direct phrenic nerve stimulation as the most effective site, especially in light of Johnson et al.’s findings. They demonstrated increased tidal volume during diaphragmatic pacing compared with baseline at 6 weeks after the initial phrenic nerve injury [28]. Further data on the longevity of direct phrenic nerve stimulation after phrenic nerve trauma or damage are needed.
In our model, we opted for transthoracic instrumentation because of the intended use of the unilateral diaphragmatic pacemaker in children after cardiac surgery. A phrenic nerve injury would be spotted immediately after extubation, and in case of severe respiratory compromise, the implantation of a unilateral pacemaker could be performed using the existing sternotomy to minimize further surgical trauma. Additionally, direct phrenic nerve instrumentation provides the best results in terms of a low threshold required for diaphragmatic stimulation. In contrast, the abdominal approach carries the risk of further surgical trauma and allows only for electrode implantation at the motor point, resulting in higher required amperages [1, 22]. Additionally, a time-consuming mapping procedure must be performed [20]. In the current era, though, abdominal electrodes can be placed laparoscopically at the motor point, minimizing surgical trauma compared with conventional techniques [29].
Although augmented ventilation after stimulation of the phrenic nerve and the motor point was achieved, only the pCO2 significantly decreased. In addition, the pO2 in these two experimental groups showed only a trend towards an increase, and pH remained stable throughout the entire experiment. These findings are in accord with previously published results [30, 31].
Limitations
This study was performed using healthy animals with normal organ function and without any previous operations. In particular,respiratory function was completely normal before the investigation. Thus, our results should be carefully interpreted, and direct extrapolation into the clinical setting is not warranted. Furthermore, the time frame in this study was brief, and no conclusions about longer term effects are possible. The experimental animals were ventilated during the entire experiment, including the period of diaphragmatic pacing, during which the level of ventilatory support was reduced to the absolute minimal level.
Conclusion
Direct stimulation of the phrenic nerve seems also to be the best choice for stimulation in a porcine model by a unilateral thoracic diaphragm pacemaker in the context of the amperage level needed.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Le Pimpec-Barthes F, et al. Diaphragm pacing: the state of the art. J Thorac Dis. 2016;8(Suppl 4):S376–86.
Glenn WW, et al. Central hypoventilation; long-term ventilatory assistance by radiofrequency electrophrenic respiration. Ann Surg. 1970;172(4):755–73.
Glenn WW, et al. Total ventilatory support in a quadriplegic patient with radiofrequency electrophrenic respiration. N Engl J Med. 1972;286(10):513–6.
DiMarco AF, et al. Phrenic nerve pacing in a tetraplegic patient via intramuscular diaphragm electrodes. Am J Respir Crit Care Med. 2002;166(12):1604–6.
Akay TH, et al. Diaphragmatic paralysis after cardiac surgery in children: incidence, prognosis and surgical management. Pediatr Surg Int. 2006;22(4):341–6.
Akbariasbagh P, et al. Risk Factors for post-Cardiac Surgery Diaphragmatic Paralysis in Children with Congenital Heart Disease. J Tehran Heart Cent. 2015;10(3):134–9.
de Leeuw M, et al. Impact of diaphragmatic paralysis after cardiothoracic surgery in children. J Thorac Cardiovasc Surg. 1999;118(3):510–7.
Joho-Arreola AL, et al. Incidence and treatment of diaphragmatic paralysis after cardiac surgery in children. Eur J Cardiothorac Surg. 2005;27(1):53–7.
Talwar S, et al. Diaphragmatic palsy after cardiac surgical procedures in patients with congenital heart. Ann Pediatr Cardiol. 2010;3(1):50–7.
Amin Z, et al. Hemidiaphragmatic paralysis increases postoperative morbidity after a modified Fontan operation. J Thorac Cardiovasc Surg. 2001;122(5):856–62.
Ovroutski S, et al. Paralysis of the phrenic nerve as a risk factor for suboptimal Fontan hemodynamics. Eur J Cardiothorac Surg. 2005;27(4):561–5.
de Leval MR, et al. Total cavopulmonary connection: a logical alternative to atriopulmonary connection for complex Fontan operations Experimental studies and early clinical experience. J Thorac Cardiovasc Surg. 1988;96(5):682–95.
Penny DJ, Redington AN. Doppler echocardiographic evaluation of pulmonary blood flow after the Fontan operation: the role of the lungs. Br Heart J. 1991;66(5):372–4.
Redington AN, Penny D, Shinebourne EA. Pulmonary blood flow after total cavopulmonary shunt. Br Heart J. 1991;65(4):213–7.
van der Woude SFS, et al. The Influence of Respiration on Blood Flow in the Fontan Circulation: Insights for Imaging-Based Clinical Evaluation of the Total Cavopulmonary Connection. Front Cardiovasc Med. 2021;8: 683849.
Gabbert DD, et al. Heart beat but not respiration is the main driving force of the systemic venous return in the Fontan circulation. Sci Rep. 2019;9(1):2034.
Hsia TY, et al. Subdiaphragmatic venous hemodynamics in patients with biventricular and Fontan circulation after diaphragm plication. J Thorac Cardiovasc Surg. 2007;134(6):1397–405 discussion 1405.
Kratz T, et al. Proof of concept of an accelerometer as a trigger for unilateral diaphragmatic pacing: a porcine model. Biomed Eng Online. 2023;22(1):55.
Kratz T, et al. A porcine model of postoperative hemi-diaphragmatic paresis to evaluate a unilateral diaphragmatic pacemaker. Sci Rep. 2023;13(1):12628.
Onders RP, et al. Mapping the phrenic nerve motor point: the key to a successful laparoscopic diaphragm pacing system in the first human series. Surgery. 2004;136(4):819–26.
Chervin RD, Guilleminault C. Diaphragm pacing: review and reassessment. Sleep. 1994;17(2):176–87.
DiMarco AF, et al. Inspiratory muscle pacing in spinal cord injury: case report and clinical commentary. J Spinal Cord Med. 2006;29(2):95–108.
Oda T, et al. Evaluation of electrical parameters for diaphragm pacing: an experimental study. J Surg Res. 1981;30(2):142–53.
Bilgutay AM, et al. Augmented ventilation by synchronous phrenic nerve stimulation. Trans Am Soc Artif Intern Organs. 1970;16:213–7.
Commission, E. 2019 Report on the Statistics on the Use of Animals for Scientific Purposes in the Member States of the European Union in 2015–2017. Belgium: European Commission Brussels; 2020.
Hess R, Kugler J. Elektroencephalographie In Klinik Und Praxis. Eine Einfuhrung. Switzerland: Schwabe & CO; 1967.
Judson JP, Glenn WW. Radio-frequency electrophrenic respiration. Long-term application to a patient with primary hypoventilation. Jama. 1968;203(12):1033–7.
Johnson V, Eiseman B. Reinforcement of ventilation with electrophrenic pacing of the paralyzed diaphragm. J Thorac Cardiovasc Surg. 1971;62(4):651–7.
Schmit BD, et al. Laparoscopic placement of electrodes for diaphragm pacing using stimulation to locate the phrenic nerve motor points. IEEE Trans Rehabil Eng. 1998;6(4):382–90.
Kaneyuki T, et al. Diaphragm pacing. Evaluation of current waveforms for effective ventilation. J Thorac Cardiovasc Surg. 1977;74(1):109–15.
Glenn WW, et al. Diaphragm pacing by radiofrequency transmission in the treatment of chronic ventilatory insufficiency. Present status J Thorac Cardiovasc Surg. 1973;66(4):505–20.
Acknowledgements
The authors thank Anne-Sophie Kronberg and Christina Oetzmann von Sochaczewski for their invaluable technical assistance. We thank Andrea Pieper and Kerstin Hagemeister for their support in organizing the animal experiments.
Funding
Open Access funding enabled and organized by Projekt DEAL. The present study was supported by the Kinderherzen Fördergemeinschaft Deutsche Kinderherzzentren e.V. (www.kinderherzen.de).
Author information
Authors and Affiliations
Contributions
TK, UH, and BB proposed the study and were responsible for the study protocol. TK analyzed the data and prepared the figures. TK, RR, and JD were responsible for the experimental devices and equipment and measurement accuracy. TK and BB designed the animal care protocol and supervised animal care during the whole study. All authors participated in planning and performing the experiments. TK and BB wrote the manuscript. All authors reviewed the manuscript and read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Kratz, T., Dauvergne, J., Ruff, R. et al. In a porcine model of implantable pacemakers for pediatric unilateral diaphragm paralysis, the phrenic nerve is the best target. J Cardiothorac Surg 19, 181 (2024). https://doi.org/10.1186/s13019-024-02707-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13019-024-02707-w