
Abstract
Background
Specific detection of the type and severity of plant abiotic stresses helps prevent yield loss by considering timely actions. This study introduces a novel method to detect the type and severity of stress in cucumber plants under salinity and drought conditions. Various features, i.e., morphological (image textural features), physiological/biochemical (relative water content, chlorophyll, catalase activity, anthocyanins, phenol content, and proline), as well as miRNA characteristics (the concentration of miRNA-156a, miRNA-166i, miRNA-399g, and miRNA-477b) were extracted from plant leaves, and machine learning methods were used to predict the type and severity of stress by having these features. Support vector machine (SVM) with parameters optimized by genetic algorithm (GA) and particle swarm optimization (PSO) was used for machine learning.
Results
The coefficient of determination of predicting the stress type and severity in plants under both stresses was 0.61, 0.82, and 0.99 using morphological, physiological/biochemical, and miRNA characteristics, respectively. This reveals machine learning methods optimized by metaheuristic optimization techniques can provide specific detection of salt and drought stresses in cucumber plants based on miRNA characteristics. Among the study miRNAs, miRNA-477b and miRNA-399g had the highest and lowest contribution to salt and drought stresses, respectively.
Conclusions
Comapred to conventional plant traits, miRNAs are more reliable features for providing us with valuable information about plant abiotic diseases at early stages. Using an electrochemical miRNA biosensor similar to one used in this work to measure the miRNA concentration in plant leaves and using a machine learning algorithm such as SVM enable farmers to detect the salt and drought stress at early stages in cucumber plants with very high accuracy.
Similar content being viewed by others


Background
Although the morphological, physiological, and biochemical characteristics of plants are significantly affected by biotic and abiotic stresses, their variations are not specific to the stress [2]. For example, different sources of stress can result in similar changes in plant height and root and shoot weights as plant morphological features. For a comprehensive understanding of plant response at the molecular and cellular level in stress-induced studies, investigating the traits that vary almost specific to the stress is favorable. Today, early detection of plant biotic and abiotic stresses has been studied by examining the expression of miRNAs in many crop plants [57].
As small non-coding RNA molecules in living organisms, miRNAs are 17 to 23 nucleotides in length and have significant effects on gene expression [25]. The targets of plant miRNAs are mainly signal enzymes and proteins involved in physiological and biochemical processes such as plant growth, regulation of plant hormones, and signal transduction [43, 75]. miRNAs not only involve plant abiotic and biotic stress responses [44], they have crucial roles in all fields of plant physiology, such as plant hormonal regulation [16] and developmental processes [8]. Light, temperature, nutrient deficiency and toxicity, drought, salinity, and carbon dioxide are common sources of plant abiotic stresses [23].
Plant stress-involved miRNA expression is mainly both spatial- and temporal-specific [42]. Identifying plant stress-responsive miRNAs helps increase our knowledge about their role in the improvement of the plant stress tolerance mechanism [86]. In fact, to understand stress tolerance in plants, it is essential to investigate miRNA-mediated gene regulatory networks that control biological processes, such as responses to the environment [22]. The roles of miRNA-target gene structures in regulating plant stress responses have been extensively reported during the last decade [21]. Studies indicate that conserved miRNA families such as miR156, miR159, miR160, miR167, miR172, miR319, miR393, miR395, miR398, and miR399 respond remarkably toward abiotic stress [86]. Recent studies have reported that some of these miRNAs exert an up(down-regulation behavior toward several plant stresses, while some miRNAs are influenced by only one stress or at least only one of their roles has been characterized [2, 60].
Salt and drought stresses are among the undesirable abiotic stresses that can influence crop production, and their severe conditions affect the physiological and biochemical properties of a large number of plants significantly [13, 50]. Plant response toward drought is studied considering physiological properties, e.g., relative water content and antioxidative enzymatic responses [11]. Salt stress influences plant growth and development through osmotic stress and ion toxicity [12]. Crops adapt to salinity by inducing changes during transcription and translational levels [81].
Cucumber (Cucumis sativus L.) is strongly sensitive to drought and salinity, particularly at the early stages of growth [85]. Severe salinity and drought significantly affect plant growth, photosynthesis, biochemistry, and texture of fruits in cucumbers [58]. Various types of protein synthesis and gene transcription occur during the plant growth and development of cucumbers, which are affected by abiotic stresses. For instance, the biosynthesis and the drought resistance change greatly by the unconventional expression of CsCER1 in cucumber plants [77]. CsDCLs, CsAGOs, and CsRDRs generally respond to abiotic stresses in cucumbers [20]. Investigating the effects of drought and salt stress on molecular regulation during seed germination and seedling growth has resulted in providing deep sequencing data of miRNA expression in cucumber plants. Du et al. [18] used proteomics and transcriptomics analysis to investigate the plant response toward salinity and drought during the post-germinative development in cucumber plants. They reported that the study stresses caused differential expression of 36 miRNAs and 768 proteins compared with the control, of which four miRNAs had similar patterns by both stresses: miR156a, miR166i, miR399g, and miR477b. Therefore, monitoring these miRNAs during exposure to drought and salinity stresses is capable of providing us with useful information about the physiology of cucumbers.
Several techniques, including microarrays [82], northern blotting [74], and polymerase chain reaction (PCR) [48], have been extensively used to investigate the influence of stress on plant miRNA functions. Nonetheless, these techniques suffer from various limitations, e.g., an unfavorable detection limit, a small linear range, and low sensitivity [52, 73]. Instead, sensors that have a living biological receptor and are called biosensors have been developed as a reliable analytical technique for the accurate and sensitive detection of miRNA concentration [33]. Constructing low-cost and portable electrochemical biosensors for the specific and sensitive detection of miRNAs that are involved in the salt and drought stresses of cucumber seems to be useful for studying the response of the plant at the molecular level. Today, hundreds of electrochemical biosensors with acceptable specificity and wide linear range have been introduced to measure the concentration of miRNAs in attomolar and femtomolar levels [17, 71]).
Supervised and unsupervised machine learning methods have been developed extensively in the field of plant science to model plant behavior during planting, cultivation, maintenance, harvesting, and post-harvest [19, 28, 32]. The machine is capable of learning the complex multivariate relationships between inputs and output using the training data [64]. The application of machine learning for computational analysis of the role of miRNAs toward plant biotic and abiotic stresses is growing since we are faced with big data and large datasets in these problems. Recently, Meher et al. [51] and Pradhan et al. [62] offered software applications for predicting various abiotic stress-specific miRNAs using features derived from miRNA sequences. They reported that the introduced tools could be efficiently utilized for large-scale prediction of abiotic stress-specific miRNAs using only sequence information.
In case of obtaining a dataset, including morphological, physiological, biochemical, as well as miRNA concentrations of cucumber plants under salt and drought stresses, there is an important question: which of these features should be trained to machine learning techniques for the reliable detection of type and severity of the stress? Therefore, in this study, salt and drought stresses at different levels are applied to young cucumber plants individually and simultaneously, and various traits of the plants were measured during the experiments to create a dataset useful for training the machine. Then, support vector machine (SVM), optimized by metaheuristic optimization algorithms, i.e., genetic algorithm (GA) and particle swarm optimization (PSO), was utilized for the prediction of the stress type and severity using the dataset. Hence, the objectives of this study are: (a) to find the performance of each morphological, physiological, and biochemical trait, as well as miRNA concentrations for the detection of salt and drought stresses in cucumber plants, and (b) to determine a reliable machine learning algorithm optimized by sophisticated evolutionary and swarm-based methods for the specific detection of stress.
Material and methods
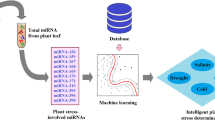
Figure 1 shows the flowchart of the present study. Morphological features, including energy, entropy, and homogeneity of images captured from plant leaves, were extracted using an image processing unit. Laboratory experiments were conducted to measure relative water content, chlorophyll, catalase activity, anthocyanins, phenol content, and proline in plant leaves. An electrochemical miRNA biosensor, introduced by Hakimian and Ghourchian [26], was used to measure miRNAs in the plant samples. In this scheme, a thiolated oligonucleotide probe is immobilized on the surface of a gold working electrode. The probe is then located in the sample to conduct probe-target hybridization. As electroactive labels, positively-charged polyethyleneimine-silver nanoparticles are then absorbed onto the hybridization product, which is negatively charged. The anodic peak current obtained due to the oxidation of silver nanoparticles will be proportional to the target miRNA concentration. Then, extracted features (as machine inputs) were trained to SVM, the parameters of which were optimized by GA and PSO. The machine outputs were the type and severity of the stress.

Flowchart of the present study
Plant material and experimental design
Cucumber seeds were purchased from a local store. After surface sterilizing with NaClO and rinsing with distilled water thoroughly, the seeds were located in plug trays containing vermiculite: peat: perlite (3: 2: 1). After 3 weeks, seedlings were transferred to 5 L pots with a similar growth medium of plug trays. The light intensity of the growth room was 120 μmolm−2 s−1 with a relative humidity of 70 ± 5%. Tap water was supplied daily for the pots to provide moisture content near the field capacity.
After two months, treatments were applied: although daily tap water supply was continued for the control plants (W0), drought-treated plants were faced with withholding water at 80% (W1), 60% (W2), and 40% (W3) of field capacity to investigate various levels of stress severity. Salinity-treated plants were irrigated with water containing various concentrations of NaCl, i.e., 0 (control, S0), 20 mM (S1), 40 mM (S2), and 60 mM (S3). Therefore, plants were treated with salt and drought simultaneously, each at four levels, and since the experiments were performed with three replications, 4 × 4 × 3 = 48 pots were used in this study. Plant features were measured five times at 3 day intervals beginning with applying the stress by transferring three leave, randomly selected from each pot, to the laboratory for further analysis.
Morpho-physiological and biochemical measurements
Morphological characteristics
Digital image processing was used to extract the morphological features of the leaves. A CCD digital camera and a 200-LEDs lighting array to increase light uniformity were used for image acquisition. Leaves were extracted from the background in the captured images through Canny edge detection [6]. After converting the color images to grayscale images, the gray-level cooccurrence matrix (GLCM) was utilized to measure the spatial dependence of gray-level values [7, 31]. An arbitrary element (i,j) in GLCM demonstrates the number of times that the pixel with a gray-level value i is located adjacent to a pixel with a gray-level value j. Recent studies have revealed that three image textural features, i.e., entropy (Eq. 1), energy (Eq. 2), and local homogeneity (Eq. 3), are useful for identifying plant health status [68]
where p(i,j) is the (i,j)-th element of the GLCM. The code for image processing was written in the MATLAB R2018b programming environment.
Physiological and biochemical characteristics
The relative water content (RWC) of the plant leaves was calculated according to Goñi et al. [24] by having the fresh weight, turgid weight, and dry weight. The chlorophyll index (CI) in the leaves was measured using a spectral chlorophyll meter (CM1000 Spectrum Technologies, USA). Catalase (EC. 1.11.1.6) activity (CA), Anthocyanin content (AC), phenol content (PhC), and proline content (PC) in the leaf samples were measured according to Havir and McHale [29], Mancinelli [47], Singleton and Rossi [67], and Bates et al. [10], respectively.
miRNA concentrations
Total RNA was isolated from 0.05 g of the leaves, according to Yamaguchi et al. [79]. A three-electrode electrochemical biosensor was used to measure the stress-involved miRNA concentrations in the isolated total RNA samples. Table 1 shows a list of the target miRNAs investigated in this study and their sequence. The working, reference, and auxiliary electrodes used in the biosensor were Au, Ag/AgCl, and Pt, respectively. Oligonucleotide miRNAs were purchased in the form of thiolated capture probes. The electrochemical biosensor was prepared according to Hakimian and Ghourchian [26]. Briefly, 25 mL of 9 mM AgNO3 and 5 mL of polyethyleneimine (PEI) were stirred during an increase in temperature to prepare PEI-Ag nanoparticles. 8 mL of 1 mM thiolated probe and 8 mL of phosphate-buffered saline (PBS) were mixed and poured onto the working electrode. Then, the working electrode was immersed in a solution of 1 M PBS buffer for 30 min, and then, 2 μL of the sample was mixed with 2 mL PBS and poured onto the electrode surface. 6 mL of positively charged PEI-Ag nanoparticles were poured onto the surface of the working electrode, and the adsorption process was performed on a negatively charged probe/sample compound. The electrodes were then placed in total RNA extracts of the leaves. To measure the electrochemical data of the biosensor, cyclic voltammetry curves were obtained by scanning in the range of 0.5 to − 0.5 V and a sweeping rate of 0.2 V/s.
Machine learning method
Statistical regression models generally cannot find complex relationships between input and output variables in high-dimension problems. However, machine learning techniques can learn the relationships in datasets with hundreds of model inputs and outputs. A dataset was constructed to train machine learning methods with the experimental data gathered at different stress levels. Since 48 pots were used to investigate the effects of stresses and the data were gathered five times, the number of samples in the dataset was 240. Moreover, the total number of features was 13, including three morphological, six physiological/biochemical, and four miRNA features. SVM was used in this study to predict the type and severity of plant stress by having the values of experimental data as inputs. Two essential variables significantly influence the SVM performance: kernel type and kernel parameter (γ) [49]. Linear, polynomial, Gaussian, and sigmoid kernels are conventional kernel types in SVM. The value of γ was optimized using sophisticated metaheuristic optimization methods. As evolutionary-based and swarm intelligence-based methods, respectively, GA and PSO were utilized to optimize the kernel parameter of the SVM machine learning method to obtain a reliable machine for predicting plant stress having its characteristics.
GA involves a set of random chromosomes that describe possible solutions for the optimization problem. During generations, chromosomes with better values of fitness function can survive and crossover to create new offspring in the subsequent generations that are likely closer to the solution. The fitness function in this study is minimizing the error of predicting the plant stress using SVM. The maximum number of iterations, population size, and crossover percentage of GA were considered 500, 100, and 0.5, respectively. As another optimization technique, PSO includes a set of randomly-defined particles that move toward the solution in each iteration. The moving force of the particles in the search space is provided by a velocity vector. The particles close to the best positions (points with higher values of fitness function) will have a slow velocity, while the others will reach the best positions with higher velocities. After a number of iterations, all the particles approach the optimal point. The maximum number of iterations, population size, initial inertia weight, and cognitive acceleration in PSO were considered 100, 200, 1, and 1, respectively. The procedure of optimizing the SVM model using GA and PSO was similar to the one we used in our previous work on using optimization methods to tune the kernel parameter of SVM to obtain a reliable decision-making unit for a durable electrochemical nitrate biosensor [5].
Cooperative game theory is a reliable feature selection method in multivariate problems which was utilized in this work to determine the importance of morphological and physiological/biochemical features, as well as miRNA concentration, in the prediction of the plant stress response. There are variables with various importances in predicting the model outputs in datasets obtained from nature-based systems [69]. The cooperative game theory evaluates the amount of shared information (i.e., power or importance) by the model inputs using the Banzhaf power index [4, 70]).
A code written in the MATLAB R2018b programming environment was provided in this study to implement the machine learning and optimization algorithms. Five-fold cross-validation was used for investigating the performance of the algorithms based on the mean squared error (MSE) (Eq. 4) and coefficient of determination (R2) (Eq. 5)
where xo is the severity of the applied stress to the plant, xp is the predicted value of the severity using machine learning, and n is the number of samples.
Results and discussion
Previous studies have revealed the remarkable influence of salinity and drought stress on the morphological and physiological characteristics of cucumber plants. However, specific detection of the stress can be challenging since plant response to stress can be complex and results in delayed response to the source of the stress. According to the objectives of this study, machine learning is used along with some miRNAs up- or down-regulated in cucumber plants for the specific detection of the source of stress shortly after applying the salinity and drought treatments. The characteristics are divided into three groups to investigate their individual effects on the prediction of the source and severity of the stress in cucumber plants: morphological, physiological/biochemical, and miRNA concentration characteristics. A dataset was created to implement the machine learning algorithms. The raw data collected during the experiment is brought in Additional file 1: Table S1 in the Additional file information.
Performance of morphological characteristics in predicting plant stresses
Table 2 shows the performance of morphological variables in the prediction of plant stresses investigated in this study. As can be seen in the table, the morphological variables could not predict the type and severity of the plant abiotic stresses because of the low R2 and high MSE values. The highest prediction performance of the SVM model when both stresses were applied to the plants was 0.59, which was achieved for an SVM model with a Gaussian kernel. The morphological variables used in this study were image textural features examined by probability-density functions on GLCM. These features were extracted from the leaf images captured by an image acquisition system and transferred to a computer for further analysis. Although GA and PSO, as the metaheuristic optimization methods, could influence the performance of the machine in predicting the plant stresses, this influence was not sufficient to consider variables extracted from image processing useful for the prediction of plant abiotic stresses in cucumber plants. The highest prediction performance of the SVM model optimized with metaheuristic methods when both stresses were applied to the plants was 0.61, which was achieved for the SVM-GA model with a Gaussian kernel.
Figure 2 shows that based on the results of cooperative game theory, entropy and homogeneity shared the highest and lowest amount of information among morphological variables in the prediction of plant stress, respectively. The leaves of the control plants that grew under optimal conditions were healthy and colorful, with high levels of entropy. Treated plants with salinity and drought had lower complexity in surface structure, and therefore, the entropy of their images decreased. Over time, the leaves of the control plants became darker green in color, and the energy of their images decreased [68]. In contrast, the yellowish appearance in the leaves of the treatment plants resulted in an increase in the energy levels in their images. Moreover, the local homogeneity of the images belonging to control plants decreased as they became colorful during growth, with different shades of green. Nonetheless, the treated plants, due to their uniform color, exerted higher homogeneity values in their images. Although the treated plants showed different morphological variables than the control plants, these differences were unreliable for specific detection of the type and severity of stresses using image processing (Table 2). Other morphological traits, e.g., plant height, shoot weight, and root weight, although generally influenced by increasing the severity of stress, this influence is not specific to the type of stress, as discussed in previous investigations [2, 35, 66].

The amount of information shared by morphological variables in predicting the plant stress
Performance of physiological/biochemical characteristics in predicting plant stresses
Table 3 shows the performance of combined physiological and biochemical variables in the prediction of plant stresses investigated in this study. According to the table, a machine with inputs containing physiological and biochemical variables could perform better than one with morphological variables in specific detection of stress. In this situation, the highest prediction performance of the single SVM model when both stresses were applied to the plants was 0.79, which was achieved for a model with a linear kernel. Metaheuristic optimization methods increased this performance up to 0.82, obtained for the SVM-PSO model. Generally, SVM-PSO provided better results than SVM-GA during the modeling process.
The results of cooperative game theory revealed that CA (33%) and RWC (6%) shared the highest and lowest amount of information among physiological and biochemical variables in predicting plant abiotic stresses, respectively (Fig. 3). Other variables had scores between these two values. RWC is an essential indicator for investigating plant water status and dehydration tolerance [41]. A remarkable decrease in the RWC of cucumber plant leaves might be due to the unavailability of water in the plant soil/root system [36, 59]. However, the results of this study show RWC had the lowest score in the prediction of stress. The possible reason is that various sources of stress have similar effects on RWC, and the prediction of the type and severity of the stress is challenging using only RWC as the model input.

The amount of information shared by physiological/biochemical variables in predicting the plant stress
During the experiments, the CA values in treatment plants were remarkably greater than those of control plants. [30] have reported similar results on the increased enzyme activity due to abiotic stresses. The complex enzymatic antioxidant system in plants affects oxygen species production during their exposure to abiotic stresses [27]. Similar trends were observed for AC, PhC, and PC, so these values were higher in the plants faced with stress compared to the control. Previous studies have shown positive correlations between abiotic tolerance and changes in antioxidant activity [15]. In fact, when plants are under stress conditions, antioxidative enzyme activities protect them against stress as a defense mechanism [63]. Studies have shown that younger plants express better protection against stress by increasing the antioxidant enzymatic activity [54].
CI, which is an essential plant physiological characteristic, decreased at severe stress levels. CI is positively correlated with photosynthetic activity [45, 72], and its decrement under abiotic stresses is considered an indicator of oxidative stress [53]. CI exerted an increasing trend in the control plant leaves as they became darker green in color over time, while CI in the treatment plants decreased, which was indicated by the yellowish appearance due to water deficiency.
Performance of miRNA concentration characteristics in predicting plant stresses
As can be seen in Tables 2, 3, the prediction performance of machines trained by plant morphological and physiological/biochemical characteristics is not suitable for actual field situations since a reliable machine with high R2 and low MSE values is required by the farmers and agricultural specialists to determine the type and severity of stress correctly. Several studies have shown that the expression of miRNA156a, miRNA166i, miRNA399g, and miRNA477b changes in a tissue-specific behavior when cucumber plants are under abiotic stresses [18]. To encounter the problem during the prediction of stress, we tried to develop an intelligent model in which machine learning methods link the leaf miRNA concentration (or, in other words, expression) measured by an electrochemical biosensor to the stress. In this situation, portable electrochemical biosensors introduced in the literature that are capable of measuring the concentration of miRNAs at fM levels can be helpful for the accurate prediction of plant stresses. Moreover, features selection methods that determine the importance of each input in the prediction of model outputs can determine the contribution, i.e., effectiveness, of miRNAs to the plant stresses. This means that these methods provide us with valuable information about the miRNAs that should be measured by the biosensor to detect plant stress at early stages.
Table 4 shows the performance of miRNA concentrations in the prediction of plant stresses investigated in this study. Based on the table, miRNA concentrations were able to predict the type and severity of the stress with acceptable performance. The table also shows the type of optimization method to optimize the parameters of the SVM model has significant effects on the model performance. The most promising results (R2 = 0.99 and MSE = 0.23) were obtained by SVM-GA for the prediction of the type and severity of stress when the plants are faced with both salinity and drought conditions simultaneously.
As expected, the miRNA concentrations altered toward the stresses. If not, the machine was unable to predict the stress with reliable performance. Some of the miRNAs were induced toward the stress conditions, while others were inhibited. From the perspective of machine learning approaches, it does not matter which miRNA is up-regulated and which one is down-regulated. What matters is that their response to the stress is specific, and the machine can discover this specificity. Similarly, various miRNAs are reported to act specifically toward metabolic activities in plants and animals [1, 80, 83]. Moreover, although a search in the literature shows that investigations on miRNA concentration using biosensors mainly belong to studies on humans [38, 55], studies on using biosensors for plant miRNA concentration have been growing rapidly since 2015s [3, 14, 40, 56].
The main hypothesis that its correctness was proved in this study was that the concentration of some miRNAs extracted from cucumber plant leaves using a biosensor combined with the results of machine learning methods might be a specific marker of plant stress. However, the role of all miRNAs is not similar in the prediction task. As shown in Fig. 4, miRNA-477b concentration exerted the greatest contribution to the correct prediction of salt and drought stresses using machine learning. Therefore, among the miRNAs that are involved in plant abiotic stress, miRNA-477b concentration had the highest correlation with stress severity in cucumber plants. After that, miRNA-156a and miRNA-166i had rather similar importance in the prediction of stress, and miRNA-399 g had the lowest contribution.

The amount of information shared by miRNA concentrations in predicting the plant stress
miRNA-477 is associated with zeatin-O-glycosyltransferase regulation in leaf tissue. Expression of this miRNA is antagonistic to the respective gene under stress conditions. Previous studies have also reported that under mild and severe stress conditions, miRNA-477 and its target gene are both up-regulated and down-regulated, respectively. Zeation-O-glucosyltransferase is involved in cytokinin homeostasis and maintains the auxin/cytokinin ratio for plant growth regulation [61].
miRNA-156 is one of the well-studied plant miRNAs. It has essential functions in plant development, especially in leaf and flower development [84]. It targets Squamosa promoter-binding protein-like (SPL) transcription factors and, therefore, affects plant developmental timing [76]. Overexpression of this miRNA results in the fast establishment of rosette leaves and a rather delay in the flowering stage [65]. Besides, nitrogen deficiency in the environment induces the expression of miRNA-156 in plants [39].
An essential miRNA that controls leaf development in a wide variety of plants is miRNA-166 [46]. miRNA-166 mediates leaf morphogenesis by targeting the gene family that involves the class-III homeodomain leucine zipper transcription factor [34]. Overexpression of miRNA-166-resistant PHABULOSA mutants causes unusual changes in the leaf developmental stage. Furthermore, the overexpression of plant miRNA-166 results in vascular cell differentiation, followed by the production of a more vascular system with expanded xylem tissue [37].
miRNA-399 is an essential miRNA in plants that is highly induced in plant tissues under phosphate stress [9]. miRNA399 expression in transgenic seedlings results in suppressing the putative ubiquitin-conjugating enzyme transcript under high inorganic phosphate conditions [78]. Since miRNA-399 g had a negligible contribution to stress prediction compared to the other three miRNAs using machine learning algorithms, it was omitted from the dataset, and SVM-GA was used to predict the type and severity of the study stresses by having only the concentration of miRNA-156a, miRNA-166i, and miRNA-477b. The results showed that the R2 and MSE of prediction when both stresses were applied to the cucumber plants were 0.98 and 0.25, respectively. This indicates that developing a biosensor that measures the concentration of these three miRNAs is reliable for in-field stress determination.
Conclusions
An effort to introduce a novel method to detect the type and severity of two main abiotic stresses, i.e., salinity and drought, in cucumber plants is reported in this paper. Treatments were selected in levels to apply a range of mild to rather severe stress conditions. The results proved that in all stress conditions, the miRNA biosensor equipped with a machine learning algorithm optimized by metaheuristic methods, that can measure the concentration of miRNAs shown effective in the plant stress response based on previous studies, is suitable to detect stress in cucumber plants (R2 = 0.99). This reveals that compared to plant morphological and physiological features, miRNA concentrations are more reliable features for providing us with valuable information about plant abiotic diseases at early stages. Therefore, extracting the plant morphological and physiological features, which are usually time-consuming and require transferring the samples to well-equipped laboratories, to provide a reliable understanding of plant stress status will no longer be needed if a miRNA biosensor is available to measure the miRNA concentrations.
As a sensor based on a three-electrode platform, the biosensing method to measure the concentration of miRNAs is portable and can be easily used by farmers and agricultural specialists. The biosensor can be connected to a laptop in which the machine learning algorithm is running. Even internet-of-things (IoT) solutions to wirelessly transfer the electrochemical data of the biosensor to a host computer can be implemented to increase the functionality of the method described in this study for real in-field applications.
Availability of data and materials
The datasets used and analyzed during the current study are available from the corresponding author upon reasonable request.
References
Al-Rawaf HA. Circulating microRNAs and adipokines as markers of metabolic syndrome in adolescents with obesity. Clin Nutr. 2019;38(5):2231–8.
Asefpour Vakilian K. Gold nanoparticles-based biosensor can detect drought stress in tomato by ultrasensitive and specific determination of miRNAs. Plant Physiol Biochem. 2019;145:195–204.
Asefpour Vakilian K. Determination of nitrogen deficiency-related microRNAs in plants using fluorescence quenching of graphene oxide nanosheets. Mol cell probes. 2020;52: 101576.
Asefpour Vakilian K. Machine learning improves our knowledge about miRNA functions towards plant abiotic stresses. Sci Rep. 2020;10:3041.
Asefpour Vakilian K. Optimization methods can increase the durability of smart electrochemical biosensors. In 2022 8th Iranian Conference on Signal Processing and Intelligent Systems (ICSPIS). Behshahr, Iran. 2022 Dec 28 (pp. 1–5).
Asefpour Vakilian K, Massah J. An artificial neural network approach to identify fungal diseases of cucumber (Cucumis sativus L) plants using digital image processing. Arch Phytopathol Plant Prot. 2013;46(13):1580–8.
Asefpour Vakilian K, Massah J. A farmer-assistant robot for nitrogen fertilizing management of greenhouse crops. Comput Electron Agric. 2017;139:153–63.
Ayubov MS, Mirzakhmedov MH, Sripathi VR, Buriev ZT, Ubaydullaeva KA, Usmonov DE, Norboboyeva RB, Emani C, Kumpatla SP, Abdurakhmonov IY. Role of MicroRNAs and small RNAs in regulation of developmental processes and agronomic traits in Gossypium species. Genomics. 2019;111(5):1018–25.
Bari R, Datt Pant B, Stitt M, Scheible WR. PHO2, microRNA399, and PHR1 define a phosphate-signaling pathway in plants. Plant Physiol. 2006;141(3):988–99.
Bates LS, Waldren RA, Teare ID. Rapid determination of free proline for water-stress studies. Plant Soil. 1973;39:205–7.
Batool T, Ali S, Seleiman MF, Naveed NH, Ali A, Ahmed K, Abid M, Rizwan M, Shahid MR, Alotaibi M, Al-Ashkar I, Mubushar M. Plant growth promoting rhizobacteria alleviates drought stress in potato in response to suppressive oxidative stress and antioxidant enzymes activities. Sci Rep. 2020;10:16975.
Behera LM, Hembram P. Advances on plant salinity stress responses in the post-genomic era: a review. J Crop Sci Biotechnol. 2021;24:117–26.
Chandra S, Roychoudhury A. Penconazole, paclobutrazol, and triacontanol in overcoming environmental stress in plants. In: Roychoudhury A, Tripathi DK, editors. Protective chemical agents in the amelioration of plant abiotic stress: biochemical and molecular perspectives. Hoboken: John Wiley & Sons; 2020. p. 510–34.
Chen L, Fang Y, Zhou X, Zhang M, Yao R, Tian B. Magnetic DNA nanomachine for on-particle cascade amplification-based ferromagnetic resonance detection of plant MicroRNA. Anal Chem. 2023;95:5411–8.
Chutipaijit S, Cha-um S, Sompornpailin K. High contents of proline and anthocyanin increase protective response to salinity in Oryza sativa L spp indica. Aust J Crop Sci. 2011;5(10):1191–8.
Ding Y, Ding L, Xia Y, Wang F, Zhu C. Emerging roles of microRNAs in plant heavy metal tolerance and homeostasis. J Agric Food Chem. 2020;68(7):1958–65.
Dorosti N, Khatami SH, Karami N, Taheri-Anganeh M, Mahhengam N, Rajabvand N, Asadi P, Movahedpour A, Ghasemi H. MicroRNA biosensors for detection of gastrointestinal cancer. Clin Chim Acta. 2023;541: 117245.
Du C, Li H, Liu C, Fan H. Understanding of the postgerminative development response to salinity and drought stresses in cucumber seeds by integrated proteomics and transcriptomics analysis. J Proteomics. 2021;232: 104062.
Esmaili M, Aliniaeifard S, Mashal M, Asefpour Vakilian K, Ghorbanzadeh P, Azadegan B, Seif M, Didaran F. Assessment of adaptive neuro-fuzzy inference system (ANFIS) to predict production and water productivity of lettuce in response to different light intensities and CO2 concentrations. Agric Water Manag. 2021;258: 107201.
Gan D, Zhan M, Yang F, Zhang Q, Hu K, Xu W, Lu Q, Zhang L, Liang D. Expression analysis of argonaute, Dicer-like, and RNA-dependent RNA polymerase genes in cucumber (Cucumis sativus L) in response to abiotic stress. J Genet. 2017;96:235–49.
Gao Z, Ma C, Zheng C, Yao Y, Du Y. Advances in the regulation of plant salt-stress tolerance by miRNA. Mol Biol Rep. 2022;49(6):5041–55.
Ghosh S, Adhikari S, Adhikari A, Hossain Z. Contribution of plant miRNAome studies towards understanding heavy metal stress responses: current status and future perspectives. Environ Exp Bot. 2022;194: 104705.
Gong Z, Xiong L, Shi H, Yang S, Herrera-Estrella LR, Xu G, Chao DY, Li J, Wang PY, Qin F, Li J, Ding Y, Shi Y, Wang Y, Yang Y, Guo Y, Zhu JK. Plant abiotic stress response and nutrient use efficiency. Sci China Life Sci. 2020;63:635–74.
Goñi O, Quille P, O’Connell S. Ascophyllum nodosum extract biostimulants and their role in enhancing tolerance to drought stress in tomato plants. Plant Physiol Biochem. 2018;126:63–73.
Ha M, Kim VN. Regulation of microRNA biogenesis. Nat Rev Mol Cell Biol. 2014;15(8):509–24.
Hakimian F, Ghourchian H. Ultrasensitive electrochemical biosensor for detection of microRNA-155 as a breast cancer risk factor. Anal Chim Acta. 2020;1136:1–8.
Hasanuzzaman M, Hossain MA, da Silva JAT, Fujita M. Plant response and tolerance to abiotic oxidative stress: antioxidant defense is a key factor. In: Venkateswarlu B, Shanker AK, Shanker C, Maheswari M, editors. Crop stress and its management: perspectives and strategies. Dordrecht: Springer; 2012. p. 2012.
Hashemi A, Asefpour Vakilian K, Khazaei J, Massah J. An artificial neural network modeling for force control system of a robotic pruning machine. J Inf Organ Sci. 2014;38(1):35–41.
Havir EA, McHale NA. Biochemical and developmental characterization of multiple forms of catalase in tobacco leaves. Plant Physiol. 1987;84(2):450–5.
Jafari S, Hashemi Garmdareh SE, Azadegan B. Effects of drought stress on morphological, physiological, and biochemical characteristics of stock plant (Matthiola incana L). Sci Hortic. 2019;253:128–33.
Javidan SM, Banakar A, Asefpour Vakilian K, Ampatzidis Y. A feature selection method using slime mould optimization algorithm in order to diagnose plant leaf diseases. In 2022 8th Iranian Conference on Signal Processing and Intelligent Systems (ICSPIS). Behshahr, Iran. 2022 Dec 28 (pp. 1–5).
Javidan SM, Banakar A, Asefpour Vakilian K, Ampatzidis Y. Diagnosis of grape leaf diseases using automatic K-means clustering and machine learning. Smart Agric Technol. 2023;3: 100081.
Johnson BN, Mutharasan R. Biosensor-based microRNA detection: techniques, design, performance, and challenges. Analyst. 2014;139(7):1576–88.
Juarez M, Timmermans M. MiRNAs specify dorsoventral polarity during leaf development. Dev Biol. 2004;271(2):551–2.
Khan MM, Al-Mas’oudi RS, Al-Said F, Khan I. Salinity effects on growth, electrolyte leakage, chlorophyll content and lipid peroxidation in cucumber (Cucumis sativus L). In International Conference on Food and Agricultural Sciences Malaysia IACSIT Press. 2013. PP, 28-32
Khan AL, Hamayun M, Ahmad N, Waqas M, Kang SM, Kim YH, Lee IJ. Exophiala sp LHL08 reprograms Cucumis sativus to higher growth under abiotic stresses. Physiol Plant. 2011;143(4):329–43.
Kim J, Jung JH, Reyes JL, Kim YS, Kim SY, Chung KS, Kim JA, Lee M, Lee Y, Narry Kim V, Chua NH, Park CM. microRNA-directed cleavage of ATHB15 mRNA regulates vascular development in Arabidopsis inflorescence stems. Plant J. 2005;42(1):84–94.
Kim ER, Joe C, Mitchell RJ, Gu MB. Biosensors for healthcare: current and future perspectives. Trends Biotechnol. 2022;41(3):374–95.
Koroban NV, Kudryavtseva AV, Krasnov GS, Sadritdinova AF, Fedorova MS, Snezhkina AV, Bolsheva NL, Muravenko OV, Dmitriev AA, Melnikova NV. The role of microRNA in abiotic stress response in plants. Mol Biology. 2016;50:337–43.
Kumar V, Arora K. Trends in nano-inspired biosensors for plants. Mater Sci Energy Technol. 2020;3:255–73.
Lata C, Prasad M. Association of an allele-specific marker with dehydration stress tolerance in foxtail millet suggests SiDREB2 to be an important QTL. J Plant Biochem and Biotechnol. 2014;23:119–22.
Li C, Nong W, Zhao S, Lin X, Xie Y, Cheung MY, Xiao Z, Wong AYP, Chan TF, Hui JHL, Lam HM. Differential microRNA expression, microRNA arm switching, and microRNA: long non-coding RNA interaction in response to salinity stress in soybean. BMC Genomics. 2022;23(1):65.
Li M, Yu B. Recent advances in the regulation of plant miRNA biogenesis. RNA Biol. 2021;18(12):2087–96.
Luo P, Di D, Wu L, Yang J, Lu Y, Shi W. MicroRNAs are involved in regulating plant development and stress response through fine-tuning of TIR1/AFB-dependent auxin signaling. Int J Mol Scls. 2022;23(1):510.
Mafakheri A, Siosemardeh AF, Bahramnejad B, Struik PC, Sohrabi Y. Effect of drought stress on yield, proline and chlorophyll contents in three chickpea cultivars. Austr J Crop Sci. 2010;4(8):580–5.
Mallory AC, Reinhart BJ, Jones-Rhoades MW, Tang G, Zamore PD, Barton MK, Bartel DP. MicroRNA control of PHABULOSA in leaf development: importance of pairing to the microRNA 5′ region. EMBO J. 2004;23(16):3356–64.
Mancinelli AL. Interaction between light quality and light quantity in the photoregulation of anthocyanin production. Plant Physiol. 1990;92(4):1191–5.
Marabita F, de Candia P, Torri A, Tegner J, Abrignani S, Rossi RL. Normalization of circulating microRNA expression data obtained by quantitative real-time RT-PCR. Briefings Bioinf. 2016;17(2):204–12.
Massah J, Asefpour Vakilian K. An intelligent portable biosensor for fast and accurate nitrate determination using cyclic voltammetry. Biosyst Eng. 2019;177:49–58.
Massonnet C, Costes E, Rambal S, Dreyer E, Regnard JL. Stomatal regulation of photosynthesis in apple leaves: evidence for different water-use strategies between two cultivars. Ann Bot. 2007;100(6):1347–56.
Meher PK, Begam S, Sahu TK, Gupta A, Kumar A, Kumar U, Rao AR, Singh KP, Dhankher OP. ASRmiRNA: abiotic stress-responsive miRNA prediction in plants by using machine learning algorithms with pseudo K-tuple nucleotide compositional features. Int J Mol Sci. 2022;23(3):1612.
Mourya DT, Yadav PD, Mehla R, Barde PV, Yergolkar PN, Kumar SR, Thakare JP, Mishra AC. Diagnosis of Kyasanur forest disease by nested RT-PCR, real-time RT-PCR and IgM capture ELISA. J Virol Methods. 2012;186:49–54.
Naing AH, Kim CK. Abiotic stress-induced anthocyanins in plants: their role in tolerance to abiotic stresses. Physiol Plant. 2021;172(3):1711–23.
Nayyar H, Chander S. Protective effects of polyamines against oxidative stress induced by water and cold stress in chickpea. J Agron Crop Sci. 2004;190(5):355–65.
Negahdary M, Angnes L. Application of electrochemical biosensors for the detection of microRNAs (miRNAs) related to cancer. Coord Chem Rev. 2022;464: 214565.
Nehra A, Kumar A, Ahlawat S, Kumar V, Singh KP. Substrate-free untagged detection of miR393a using an ultrasensitive electrochemical biosensor. ACS Omega. 2022;7(6):5176–89.
Noman A, Aqeel M. miRNA-based heavy metal homeostasis and plant growth. Environ Sci Pollut Res. 2017;24(11):10068–82.
Ouzounidou G, Giannakoula A, Ilias I, Zamanidis P. Alleviation of drought and salinity stresses on growth, physiology, biochemistry and quality of two Cucumis sativus L cultivars by Si application. Braz J Bot. 2016;39:531–9.
Parkash V, Singh S. A review on potential plant-based water stress indicators for vegetable crops. Sustainability. 2020;12(10):3945.
Patel P, Yadav K, Ganapathi TR, Penna S. Plant miRNAome: Cross talk in abiotic stressful times. In: Rajpal VR, Sehgal D, Kumar A, Raina SN, editors. Genetic enhancement of crops for tolerance to abiotic stress: Mechanisms and approaches. Cham: Springer; 2019. p. 25–52.
Pineda Rodó A, Brugiere N, Vankova R, Malbeck J, Olson JM, Haines SC, Martin RC, Habben JE, Mok DWS, Mok MC. Over-expression of a zeatin O-glucosylation gene in maize leads to growth retardation and tasselseed formation. J Exp Bot. 2008;59(10):2673–86.
Pradhan UK, Meher PK, Naha S, Rao AR, Kumar U, Pal S, Gupta A. ASmiR: a machine learning framework for prediction of abiotic stress–specific miRNAs in plants. Funct Integr Genom. 2023;23(2):92.
Rajput VD, Singh RK, Verma KK, Sharma L, Quiroz-Figueroa FR, Meena M, Gour VS, Minkina T, Sushkova S, Mandzhieva S. Recent developments in enzymatic antioxidant defence mechanism in plants with special reference to abiotic stress. Biology. 2021;10(4):267.
Sarlaki E, Sharif Paghaleh A, Kianmehr MH, Asefpour Vakilian K. Valorization of lignite wastes into humic acids: process optimization, energy efficiency and structural features analysis. Renew Energy. 2021;163:105–22.
Schwab R, Palatnik JF, Riester M, Schommer C, Schmid M, Weigel D. Specific effects of microRNAs on the plant transcriptome. Dev Cell. 2005;8(4):517–27.
Schwarz D, Rouphael Y, Colla G, Venema JH. Grafting as a tool to improve tolerance of vegetables to abiotic stresses: Thermal stress, water stress and organic pollutants. Sci Hortic. 2010;127(2):162–71.
Singleton VL, Rossi JA. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am J Enol Vitic. 1965;16(3):144–58.
Story D, Kacira M, Kubota C, Akoglu A, An L. Lettuce calcium deficiency detection with machine vision computed plant features in controlled environments. Comput Electron Agric. 2010;74(2):238–43.
Sun X, Liu Y, Li J, Zhu J, Chen H, Liu X. Feature evaluation and selection with cooperative game theory. Pattern recognit. 2012;45(8):2992–3002.
Sun J, Zhong G, Huang K, Dong J. Banzhaf random forests: cooperative game theory based random forests with consistency. Neural Netw. 2018;106:20–9.
Tran HV, Piro B. Recent trends in application of nanomaterials for the development of electrochemical microRNA biosensors. Microchim Acta. 2021;188(4):128.
Tsai YC, Chen KC, Cheng TS, Lee C, Lin SH, Tung CW. Chlorophyll fluorescence analysis in diverse rice varieties reveals the positive correlation between the seedlings salt tolerance and photosynthetic efficiency. BMC Plant Biol. 2019;19:1–17.
Turner AP. Biosensors: sense and sensibility. Chem Soc Rev. 2013;42(8):3184–96.
Várallyay E, Burgyán J, Havelda Z. MicroRNA detection by northern blotting using locked nucleic acid probes. Nat Protoc. 2008;3(2):190–6.
Varkonyi-Gasic E, Gould N, Sandanayaka M, Sutherland P, MacDiarmid RM. Characterisation of miRNAs from apple (Malus domestica ‘Royal Gala’) vascular tissue and phloem sap. BMC Plant Biol. 2010;10:159.
Wang QL, Li ZH. The functions of microRNAs in plants. Front Biosci. 2007;12(10):3975–82.
Wang W, Zhang Y, Xu C, Ren J, Liu X, Black K, Gai X, Wang Q, Ren H. Cucumber ECERIFERUM1 (CsCER1), which influences the cuticle properties and drought tolerance of cucumber, plays a key role in VLC alkanes biosynthesis. Plant Mol Biol. 2015;87:219–33.
Xie FL, Huang SQ, Guo K, Xiang AL, Zhu YY, Nie L, Yang ZM. Computational identification of novel microRNAs and targets in Brassica napus. FEBS Lett. 2007;581(7):1464–74.
Yamaguchi K, Takahashi Y, Berberich T, Imai A, Takahashi T, Michael AJ, Kusano T. A protective role for the polyamine spermine against drought stress in Arabidopsis. Biochem Biophys Res Commun. 2007;352(2):486–90.
Yang B, Tang J, Yu Z, Khare T, Srivastav A, Datir S, Kumar V. Light stress responses and prospects for engineering light stress tolerance in crop plants. J Plant Growth Regul. 2019;38:1489–506.
Yepes-Molina L, Bárzana G, Carvajal M. Controversial regulation of gene expression and protein transduction of aquaporins under drought and salinity stress. Plants. 2020;9(12):1662.
Yin JQ, Zhao RC, Morris KV. Profiling microRNA expression with microarrays. Trends Biotechnol. 2008;26(2):70–6.
Zen K, Zhang CY. Circulating microRNAs: a novel class of biomarkers to diagnose and monitor human cancers. Med Res Rev. 2012;32(2):326–48.
Zhang BH, Pan XP, Wang QL, Cobb GP, Anderson TA. Identification and characterization of new plant microRNAs using EST analysis. Cell Res. 2005;15(5):336–60.
Zhang W, Jiang B, Li W, Song H, Yu Y, Chen J. Polyamines enhance chilling tolerance of cucumber (Cucumis sativus L) through modulating antioxidative system. Sci Hortic. 2009;122(2):200–8.
Zhang F, Yang J, Zhang N, Wu J, Si H. Roles of microRNAs in abiotic stress response and characteristics regulation of plant. Front Plant Sci. 2022;13: 919243.
Acknowledgements
Not applicable.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
PM collected data, implemented the machine learning codes, and wrote the draft. KAV designed the experiment, developed the electrochemical biosensor, and revised the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
All authors affirm consent for publication.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Table S1.
Cucumber plant characteristics in various drought and salinity stress levels. W0 to W3 and S0 to S3 indicate drought and salinity stress levels, respectively, which are described in the main manuscript.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Mohammadi, P., Asefpour Vakilian, K. Machine learning provides specific detection of salt and drought stresses in cucumber based on miRNA characteristics. Plant Methods 19, 123 (2023). https://doi.org/10.1186/s13007-023-01095-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13007-023-01095-x