
Abstract
Background
Fat storage is required for the life cycle of many organisms. The primary fat depot for most vertebrates is white adipose tissue. However, in primitive vertebrates (e.g., agnathan group and elasmobranchs), the liver is usually responsible for fat storage. Among the vertebrates, amphibians have a unique status, as their larvae live in the water and exhibit some primitive traits that are similar to fish. Although it has been recognized that adult frogs use their abdominal white adipose tissue as a primary fat depot, how tadpoles store their fat is still inconclusive. The metabolic traits and physiological functions of primitive fat depots may have wide-ranging implications on the pathology of abnormal lipid deposition in mammals and the evolution of fat storage.
Results
Rana omeimontis tadpoles used their liver as the primary fat depot. In sufficiently fed tadpoles at stage 30–31, the hepatosomatic index (HSI) reached up to 7%, and triglycerides (TG) accounted for 15% of liver weight. Their liver resembled white adipose tissue in histological morphology, characterized by polygonal hepatocytes filled with fat. Their liver metabolic composition was unique, characterized by the dominance of maltotriose, arachidonic acid and dipeptides in soluble carbohydrates, free fatty acids and amino acids. Hepatic fat was the major metabolic fuel of fasted R. omeimontis tadpoles, which had similar reserve mobilization and allocation patterns as mammals. From a developmental perspective, hepatic fat was important to fuel late metamorphic climax. Interestingly, starvation induced accelerated metamorphosis in tadpoles with high HSI (4.96 ± 0.21%). However, this phenomenon was not observed in tadpoles with low HSI (2.71 ± 0.16%), even though they had similar initial body weight and developmental stage. Hepatic fat abundance was the most prominent difference between the two groups.
Conclusion
To the best of our knowledge, this is the first report that liver can be the primary fat depot in vertebrates with higher evolutionary status than bony fish. The unique hepatic histological and metabolic traits likely either guard their liver against lipotoxicity or make their hepatocytes adapt to fat accumulation. This fatty liver could be a primitive counterpart of mammalian white adipose tissue (WAT). In addition, our study showed that the hepatic reserves of tadpoles, especially TG content, may provide body condition signals to modulate metamorphosis.
Similar content being viewed by others

Background
Fat storage is required for the life cycle of many organisms. Primitive vertebrates (e.g., the agnathan group), elasmobranchs (e.g., sharks) and some bony fish (e.g., cod, coley and flatfish) synthesize and store most of their body fat in their liver [1,2,3,4,5,6]. In other bony fish, amphibians, reptiles, birds and mammals, even though liver is still responsible for lipogenesis, white adipose tissue (WAT) appears and replaces liver to become their primary fat depot [2, 7,8,9,10]. Liver can still be a secondary fat depot in fish, amphibians and reptiles [8, 9, 11,12,13], but hepatic lipid deposition in mammals and humans may be associated with increased risk of liver injuries [14, 15]. Accordingly, fat deposition in the liver is likely a primitive trait of vertebrate evolution [2]. From this point of view, the metabolic traits and physiological functions of the primitive fatty liver may have wide-ranging implications on the pathology of abnormal lipid deposition in humans and shed light on the driving force behind evolution of fat depot [15].
Although it has been recognized that the fat body, the major abdominal WAT of amphibians, is the primary fat depot in adult frogs [8, 9, 16, 17], little attention has been paid to the fat storage of tadpoles. Some previous studies indicated that the fat body could also be the major fat depot in tadpoles (e.g., Lithobates catesbeianus and R. curtipes) [9, 18,19,20,21,22]. They observed that it can provide energy to tadpoles during starvation, and it is also necessary for sustaining energy production at the late metamorphic climax of tadpoles when apoptotic tissues have been absorbed [23]. A recent study even suggested that the fat body might play critical roles in determining whether metabolic reserves meet the nutrient requirements of metamorphic climax of tadpoles [24]. The fat level of metamorphosizing tadpoles is associated with the survival strategies of froglets and the survival rate and reproductive capacity of adults [25, 26]. Despite the significance of fat body during tadpole metamorphosis, some frog species, e.g., Microhyla fissipes, R. chaochiaoensis, R. omeimontis and Bufo melanostictus, are devoid of fat-accumulating fat bodies across their larval stages (our observation and [18]). Surprisingly, in our preliminary studies, we found that R. omeimontis tadpoles store fat mainly in their livers, whose histological structure resembles that of adipose tissue if food is available. Although hepatic fat accumulation has also been observed in overwintering frogs [27,28,29], their liver is the major organ for triglyceride synthesis but a secondary site for triglyceride deposition [9]. To the best of our knowledge, the fatty liver of R. omeimontis tadpoles is the first observation that liver can be a primary fat depot in amphibians, or indeed in any animals with higher evolutionary status than bony fish. Importantly, these tadpoles likely experience a transition of the fat depot from liver to the fat body after metamorphosis. Such a phenomenon makes them unique models for investigating the regulation mechanisms of fat deposition.
Currently, we know little about the metabolism and physiology of amphibians’ use of liver as the primary fat depot. The most basic knowledge is the metabolic composition of their fatty liver. In humans and other mammals, hepatic lipid deposition is associated with increased risk of hepatic injury and steatohepatitis due to lipotoxicity [30,31,32]. It has been recognized that the lipid types that accumulate in liver, rather than their total amount, are tightly associated with the occurrence of lipotoxicity [33, 34]. For example, the accumulation of saturated free fatty acid (FFA) and diacylglycerol likely increases the probability of lipotoxicity, while a high ratio of monounsaturated FFAs to saturated FFAs is a sign of a low probability of lipotoxicity [33, 34]. Since hepatic fat deposition is a natural biological process in R. omeimontis tadpoles, these tadpoles are likely avoiding liver lipotoxicity. Uncovering the metabolic profile of their liver may be valuable in verifying and extending our knowledge of liver injuries caused by hepatic steatosis. Another basic question is whether and how metabolism differs between R. omeimontis tadpoles and animals with extrahepatic fat depots, as well as between R. omeimontis tadpoles and other animals with only a hepatic fat depot. Since liver is the common depot of glycogen, amino acids and lipids of R. omeimontis tadpoles, it would be interesting to know how their hepatic reserves are metabolized during starvation. The order of reserve mobilization during starvation is an important metabolic trait of animals. It varies between species, with potential implications for the physiological significance of metabolic reserves and their storage organs [35]. In elasmobranchs, amino acids and carbohydrates are preferentially mobilized over lipids, partly due to the role of hepatic fat in providing hydrostatic lift [36]. However, hepatic fat is preferentially consumed over glycogen and tail proteins in Atlantic cod (Gadus morhua) [37]. The differential usage of metabolic fuels between organs is also an important aspect of catabolism in animals. For example, glucose and ketone bodies synthesized in the liver are consumed by numerous organs except the liver in fasted mammals, while FFAs released from WAT are mainly used in liver and muscle for energy production. These metabolic traits in naturally fatty livers may be meaningful in understanding the metabolic disorder in human fatty liver disease. It is also important to know whether hepatic fat has physiological functions similar to those of the fat body, especially with respect to fueling and regulating metamorphosis. Answering these questions may deepen our understanding of amphibian physiology and provide insight into the functional evolution of fat depot.
To answer the questions mentioned above, we first observed fat deposition in the R. omeimontis tadpoles. Then, we investigated the metabolic profiles and the metabolic pattern of hepatic reserves during starvation by metabolomics. Last, we investigated the role of hepatic lipids during tadpole metamorphosis.
Materials and methods
Animal sampling and culture
R. omeimontis is distributed in plain, hill and mountain areas (altitude 250–2100 m. a. s. l.) in Sichuan, Gansu, Chongqing, Hunan, Guizhou and Hubei provinces in China [38]. Their breeding seasons are from the end of August to October ([38] and our observation). Their tadpoles live across winter and metamorphosize in the next May to July [38]. In this study, ten egg clutches of R. omeimontis were collected in October at Anzihe Natural Reserve (103.459885° E, 30.744614° N, 701 m) in Sichuan Province, China. These clutches (ranging from 400 to more than 1000 eggs) were placed in aquatic containers (42 × 30 × 10 cm, water depth = 5 cm) and hatched at 20 ± 0.5 °C (water temperature, L:D = 12 h:12 h). Each clutch usually occupied one container, but a large clutch might be divided into several containers. After their egg yolks were entirely absorbed, larvae were fed a solution of boiled chicken egg yolk once a day for 2 days. Then, tadpoles were fed spirulina powder (China National Salt Industry Corporation, Table 1) once a day, and water was replaced daily. Tadpoles cultured in the same container were defined as a population. These populations ranged from 400 to 1000 individuals. The developmental stages of tadpoles were identified according to the Gosner stage [39].
Rana omeimontis tadpoles are voracious feeders, and their growth rate is positively correlated with the amount of spirulina powder provided daily. Field observation indicated that at least 5–6 months (e.g., from October to May) are required for completing their metamorphosis [38]. If enough food is provided (residual spirulina powder could be observed until next feeding), these tadpoles can complete their aquatic life history within 2 months at 20 ± 0.5 °C (our experience). Tadpole populations kept in our laboratory were normally fed with approximately 0.7 g spirulina powder daily (ground in 10 ml water, an insufficient feeding program) (Fig. 1). Such a feeding program could support the nutrient requirement of tadpole populations with 400–600 individuals to reach their metamorphic climax within 5 months. Increased food availability could improve the liver weight/volume and hepatic fat content of tadpoles. When necessary, tadpoles might be provided with enough food (sufficiently feeding program) to obtain tadpoles with better body condition and more fat deposition. According to our experience, a sufficient feeding program lasting for 7 days was enough to significantly increase the liver weight/volume of insufficiently fed tadpoles and produce typical fatty liver.

Schematic diagram of the experimental design
Experimental design
Experiment 1 aimed to reveal the mobilization order of hepatic reserves. Two normally fed tadpole populations (with most individuals reaching stage 29–30) were transferred into a sufficient feeding program. After 7 days, individuals at stage 30–31 were collected (391.5 ± 19.5 mg, mean ± SE). These tadpoles were divided randomly into two groups and fasted at 20 °C (typical temperature in spring) and 8 °C (typical water temperature in winter), respectively. Then, time series sampling (0, 2, 5 and 10 days for 20 °C; 0, 2, 8 and 30 days for 8 °C) was conducted to obtain tadpoles with reducing nutrient stores, and 6–13 individuals were sacrificed from each group at each time point.
Experiment 2 aimed to characterize the role of hepatic fat in metamorphosis. Tadpole populations in Experiment 1 were fed sufficiently until metamorphosis, and individuals were collected at stage 36, 41, 43 and 44 sequentially.
Experiment 3 aimed to reveal the influence of hepatic fat on starvation-induced metamorphosis. Stage 30–31 tadpoles with high hepatic fat accumulation were obtained following the method in experiment 1. Stage 30–31 tadpoles with low hepatic fat accumulation were collected from populations under a normal feeding program. Then, both tadpole groups were starved at 20 and 8 °C, respectively. Then, time series sampling was conducted, and 6–13 individuals were sacrificed for each group at each time point.
Micro-computed tomography (micro-CT)
After anesthetization by MS-222, tadpoles were fixed in 4% paraformaldehyde for more than 24 h and stained in I2 & KI water solutions (respectively, 1% & 2%, w/v) for 12 h. A CT scan was conducted on a Quantum GX Micro CT (PerkinElmer) with the following parameters: scanning current, 70 eV; 10 μM; field of view: 36 × 36 mm for acquisition, 25 × 25 mm for reconstruction; scan duration, 15 min.
Fat tissue-specific staining
Anesthetized tadpoles were dissected on the abdomen and immersed in 10% (w/v) dithizone-ethanol solution diluted 500-fold with water (with 2% (v/v) ammonia for solubilization) for more than 30 min. Tissues rich in fat were stained dark green, while other tissues were stained orange to red [40].
Histological sectioning
Anesthetized tadpoles were dissected for collecting liver. These samples were stored in 4% paraformaldehyde until histological analyses. After dehydration in a graded series of ethanol and transparency by xylene, livers were embedded in paraffin and sectioned in serial transverse sections (7 μm thick). Hematoxylin and eosin (H & E) staining, periodic acid-Schiff (PAS) staining and red oil (RO) staining were conducted to show general histological characteristics, neutral lipid content, and glycogen content, respectively.
Thin-layer chromatography (TLC)
Anesthetized tadpoles were weighed and dissected to collect liver, tail and the rest of the carcass. Weighted tissues were transferred into 2 ml EP tubes with 1 volume Milli-Q water, 2 volumes chloroform and 4 volumes methanol. Then, the tissues were sufficiently homogenized and incubated at 60 °C for 1 h. After centrifugation at 12,000 g for 10 min, 150 μL supernatant was transferred into a clean tube with 50 μL chloroform and 50 μL Milli-Q water, followed by 15 s of vortexing. The mixture was centrifuged at 10,000 g for 10 min to collect the chloroform phase. This solution was dried in a vacuum dryer and dissolved in 30 μL chloroform:methanol (2:1, v/v). Now, the total lipid sample was ready to be analyzed on a silica gel chromatography plate (GF254 100 × 200 mm, AR). After developing with hexane:diethyl ether:acetic acid (60:20:1, v/v), the plate loaded with lipid sample was dried by an air blower and fully sprayed with 10% phosphomolybdic acid in ethanol. Coloration was conducted in an oven at 105 °C for 5 min, and the image of the plate was captured with the Gel Logic 200 Imaging System (Kodak).
Determination of hepatic glycogen and triglycerides
Anesthetized tadpoles were weighed and dissected to collect the liver. After adding 500 μL 0.1 M PBS (pH 7.4), tissues were homogenized by a tissue grinder, followed by centrifugation at 1000 g for 15 min (4 °C). The supernatant was ready for triglyceride determination. The remaining supernatant (300 μL) was mixed with 600 μL 10% trichloroacetic acid (TCA, w/v). After centrifugation at 1000 g for 15 min, 300 μL of supernatant was transferred into a new tube with 1200 μL of 95% ethanol and allowed to rest overnight. After centrifugation at 1000 g for 15 min, the precipitate was dissolved in 50 μL of water to form the glycogen solution.
The GPO-PAP method was used for triglyceride determination. The method followed the manufacturer’s instructions for the triglyceride assay kit (Nanjing Jiancheng Bioengineering Institute, China). The reaction system included 2.5 μL of supernatant and 250 μL of reagent mix (Tris-HCl, 100 mM; lipase ≥3000 U/L; ATP, 0.5 mM; glycerol kinase, ≥ 1000 U/L; glycerol 3-phosphate oxidase, ≥ 5000 U/L; peroxidase, ≥ 1000 U/L; 4-aminoprotepyrine, 1.4 mM; p-chlorophenol, 3 mM). After incubation at 37 °C for 10 min, the optical density at 510 nm of the mixtures was determined. Glycerol solution (2.26 mM) was used to generate a standard curve.
The reaction system of glycogen measurement included 5 μL of glycogen solution, 85 μL of water and 400 μL of 0.2% anthrone in 80% sulfuric acid. After incubation at 95 °C for 15 min and cooling at room temperature, the optical density at 626 nm of the mixtures was determined. Soluble starch (2 mg/ml) was used to generate a standard curve.
Metabolomic profiling
After grinding in liquid nitrogen, 100 mg tissue powder was transferred into 1.5 ml Eppendorf tubes with 1 ml methanol:acetonitrile:water = 2:2:1 (v/v), followed by ultrasonication for 30 min × 2 and incubation at − 20 °C for 1 h. After centrifugation at 12,000 for 15 min (4 °C), the supernatants were transferred into new tubes and freeze-dried. Samples were dissolved in 100 μL acetonitrile:water (1:1, v/v) for analysis.
Extracted supernatants were analyzed by LC (1290 Infinity LC, Agilent) coupled with quadrupole-time-of-flight mass spectrometry (Triple TOF 5600+, AB SCIEX). The column was equilibrated with 95% (v/v) solvent A consisting of 25 mM ammonium acetate and 25 mM ammonium hydroxide in Milli-Q water, and 2 μL of each sample was separated by a C18 HILIC column (ACQUITY UPLC HSS T3 1.8 μm, 2.1 mm × 100 mm, Waters) at 25 °C. Separation was performed with 40–95% solvent B (acetonitrile) at 0.3 ml/min as follows: 0–0.5 min, 95% B; 0.5–7 min, decreasing B from 95 to 65%; 7–8 min, decreasing B from 65 to 40%; 8–9 min, 40% B; 9–9.1 min, increasing B from 40 to 95%; 9.1–12 min, 95% B.
Metabolite data were obtained in both positive and negative ion modes with the following settings: ion source gas 1, 60; ion source gas 2, 60; curtain gas: 30; source temperature: 600 °C; ion spray voltage floating, ± 5500 V; TOF MS scan m/z range: 60–1200 Da; product ion scan m/z range: 25–1200 Da; TOF MS scan accumulation time, 0.15 s/spectrum; product ion scan accumulation time, 0.03 s/spectrum. The MS/MS spectra were acquired by information-dependent acquisition with high sensitivity as follows: declustering potential, ± 60 V; collision energy, 30 eV.
Metabolite data were processed using XCMS software (http://metlin.scripps.edu/download/) and Microsoft Excel (Microsoft, Redmond, WA, USA). Data on impurity peaks from column bleeds were excluded. Metabolites were identified by a combination of molecular weight comparison (molecular ion peak) and MS/MS spectrum comparison to a standard library. The relative abundances/concentrations of metabolites are presented as the ion intensities of their molecular ion peaks (Additional file 1).
Statistical analysis
Hepatosomatic index (HSI) was calculated as the ratio of liver weight to body weight. As the body weight of tadpoles may be variable due to digesta excretion, the hepato-tail index (HTI; the ratio of liver weight to tail weight) was also used to evaluate the relative size of the liver.
Basic statistical analyses were conducted using IBM SPSS v21.0 (IBM, Armonk, NY, USA). Changes in metabolite levels between control and 10-d-fasted tadpoles were analyzed using the independent-sample T test. Since most analyses revealed variation trends in whole groups of metabolites, rather than variations in individual metabolites, no FDR corrections were performed. Differences in body indexes and hepatic reserves between tadpoles with different starvation durations were analyzed using one-way ANOVA, followed by the Student–Newman–Keuls post hoc test. Principal component analysis (PCA) of metabolomes was conducted using Simca-P + 11 (Umetrics AB, Umea, Sweden), with the scaling-type parameter set as ‘Par’. Graphs were created using GraphPad Prism 5 or ggplot2, an R package [41, 42]. Numeric data used in graphs are presented in Additional files 1 and 2.
Results
Liver is the primary fat depot in R. omeimontis tadpoles
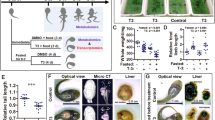
R. omeimontis tadpoles have no obvious fat body in their larval stages. Instead, a large liver (typically presenting an oily yellow color, Fig. 2a-b) can be detected in the abdominal cavity of tadpoles after Gosner stage 25. For example, the livers of tadpoles at stage 30–31 can account for 7.24 ± 0.23% of the total weight (including the massive intestinal contents) if food is sufficiently provided. After staining with dithizone, these large livers presented a dark green color (Fig. 2c), suggesting the presence of abundant neutral lipids. The histological patterns of their livers resembled the WAT, with enlarged hepatocytes filled by neutral lipid, which resulted in marginalized cell nuclei and glycogen (Fig. 2d-f). According to the results of dithizone staining (Fig. 2c) and TLC analysis (Fig. 2g), the liver is the primary fat depot of R. omeimontis tadpoles.

Hepatic fat deposition phenomenon in R. omeimontis tadpoles. a Typical morphology of a stage 31 R. omeimontis tadpole and its liver. b Micro-CT imaging of a stage 31 R. omeimontis tadpole. c Fat-specific stain by dithizone. Tissues with abundant neutral lipids are stained green. The liver is the only organ stained green. d-f Histological morphology of liver of R. omeimontis tadpoles: D, H&E staining; E, PAS staining; F, RO staining. g Distribution of triglycerides in R. omeimontis tadpoles (by TLC)
Metabolite profile of R. omeimontis tadpoles
A total of 450 metabolites were identified in R. omeimontis liver, and the most abundant metabolites (accounting for > 1% of the total abundance of identified metabolites) were mainly oligosaccharides, FFAs, phospholipids, nucleosides, and amino acids (Fig. 3a). Maltotriose was the dominant carbohydrate in the liver, accounting for 60% of the total carbohydrate (Fig. 3b). Maltose, stachyose and cellobiose also each had a higher abundance than glucose. Fifteen FFAs were identified, and unsaturated ones, including arachidonic acid (20:4Δ5,8,11,14), oleic acid (18:1Δ9), linoleic acid (18:2Δ9,12), γ-linolenic acid (18:3Δ6,9,12) and palmitoleic acid (16:1Δ9), accounted for more than 93% of the total FFAs in the liver (Fig. 3c). Amino acids were stored as both free (n = 18) and dipeptides (n = 135) in the liver (Additional file 1), accounting for 35.7 and 64.3% of their sum, respectively (Fig. 3d).

Metabolic profile of liver of R. omeimontis tadpoles (Stage 30–31). a Metabolites with an abundance higher than 1% of all identified metabolites. b Profile of identified soluble carbohydrates. c Profile of identified FFAs. d Profile of identified amino acids. e Proportion of free amino acids and dipeptides in the total amino acid pool. Each column in these two figures represents the mean ± SE of 7 samples
A total of 281 metabolites were identified in the tails. The metabolome of the tail was characterized by a high abundance of carnosine, inosine and creatine (Additional file 3: Figure S1 A), which are related to muscle function. In tails, maltotriose was still the dominant carbohydrate, with much higher abundance than that of glucose (Additional file 3: Figure S1 B). Unlike the liver, the tail had comparable total amounts of saturated and unsaturated FFAs. Stearic acid and palmitic acid were the two most dominant FFAs (Additional file 3: Figure S1 C). Amino acids in tails were mainly in the free form (84.4%) (Additional file 3: Figure S1 D).
Metabolism of hepatic reserves during starvation
The liver of R. omeimontis tadpoles dwindled in size and weight with the prolongation of fasting, accompanied by color changes from yellow to red and then to brown (Fig. 4a-b). At 20 °C, livers showed rapid weight reduction after fasting, with significant decreases in hepatic protein, glycogen and triglyceride levels (Fig. 4c-e). Glycogen and triglycerides also decreased in content relative to liver weight, suggesting that they were disposable energy reserves. Although glycogen and triglycerides were consumed simultaneously, hepatic glycogen was exhausted within 5 days, while hepatic triglyceride reserves could sustain catabolism more than 10 days. At 8 °C, tadpoles consumed their hepatic resources much more slowly than their 20 °C counterparts (Fig. 4). Glycogen was preferentially consumed as fuel over triglycerides, whose level was unchanged even after 30 days of fasting. After 10 days of fasting at 20 °C, R. omeimontis tadpoles consumed most of their hepatic triglycerides. The morphology of their liver changed dramatically (Fig. 5a), to a smaller size and darker color. The liver shifted from an adipose-like histological pattern to that of common hepatocytes, and the diameter of hepatocytes was much smaller due to the disappearance of huge lipid drops (Fig. 5a).

Mobilization of hepatic reserves in fasted R. omeimontis tadpoles. a Morphological variation of the liver of fasted tadpoles; arrows indicated gall bladders. Change of liver weight (b), total protein (c), glycogen (d) and triglycerides (e) during starvation. Each value represents the mean ± standard error (n = 6–13). Different letters indicate significant differences between values (p < 0.05), as shown by the Student–Newman–Keuls post hoc test after one-way ANOVA

Comparison between fed and 10-d-fasted R. omeimontis tadpoles. a Comparison of the histological morphology of the liver; arrows indicated two typical gall bladders. b-c Scatter plots comparing the metabolic profiles of liver (b) and tail (c) as found by PCA analysis
To reveal how hepatic reserves were consumed by tadpoles at 20 °C, liver and tail metabolomes were compared between fed and 10-d-fasted tadpoles. According to the results of PCA, the first principal component (PC1, accounting for 58.8% of the total variance) well divided fed and fasted tadpoles into two groups (Fig. 5b-c). R. omeimontis tadpoles consumed most of their major hepatic FFAs after 10 days of starvation (Fig. 6a), with 85, 74, 72, 93, and 93% reductions in arachidonic acid, oleic acid, linoleic acid, linolenic acid and palmitoleic acid, respectively. In the tail, despite palmitic acid, palmitoleic acid, linolenic acid and heptadecanoic acid decreasing, oleic acid and arachidonic acid, the 2 most abundant FFAs in the liver, increased after starvation (Fig. 6b). Increased levels of carnitine, acetyl-carnitine and stearoyl-carnitine were observed after starvation in both liver and tail (Fig. 6c-d), suggesting robust lipid consumption during starvation of tadpoles. Although hepatic glycogen was exhausted after 5 days of starvation, tadpoles fasted for 10 days maintained their hepatic soluble carbohydrates and glycolic intermediates (mainly sugar phosphates) at concentrations 25–50% of those in fed tadpole livers (Fig. 7a-b). In tails, the concentrations of oligosaccharides (maltotriose, stachyose and maltopentaose), disaccharides (maltose, cellobiose, mannobiose and sucrose) and monose (glucose) were decreased, unchanged and increased, respectively (Fig. 7c). The glycolic intermediates in the tail were maintained at comparable levels to those in fed tadpoles even after 10 days of starvation (Fig. 7d). Eight of 18 free amino acids and 70/135 dipeptides decreased significantly (not shown) after 10 days of starvation, resulting in an overall downtrend of total amino acids (16 of 23 decreased, Fig. 8a). In the tail, however, starvation caused an overall uptrend of amino acids in both free and dipeptide forms (Fig. 8b), and the concentrations of glutamine, alanine, serine and threonine showed significant increases (p < 0.05). In the aerobic stage, the levels of malate and succinate, two metabolites in the TCA cycle, were decreased significantly in the liver but not in the tail (p < 0.05) after 10 days of starvation (Additional file 3: Figure S2), implying a decreased and unchanged consumption rate of metabolic fuels in liver and tail, respectively.

Variation of metabolites involved in lipid metabolism after 10 days of starvation. a-b Variation of free amino acids in liver a and tail b. c-d Variation of intermediates in lipid metabolism in liver c and tail d. Each column represents mean ± SE (n = 5–7), ***: p < 0.001, **: p < 0.01, *: p < 0.05

Variation of metabolites involved in carbohydrate metabolism after 10 days of starvation. a-b Variation of free carbohydrates (a) and intermediates in glycolytic pathways (b) in liver. c-d Variation of free carbohydrates (c) and intermediates in glycolytic pathways (D) in the tail. Each column represents a mean ± SE (n = 5–7), ***: p < 0.001, **: p < 0.01, *: p < 0.05

Variation of amino acids after 10 days of starvation. a Variation trends of amino acids in liver. b Variation trends of amino acids in the tail. Each column represents the percentage change of an amino acid after 10 days of starvation (n = 5–7). ***: p < 0.001, **: p < 0.01, *: p < 0.05
Variation of hepatic fat in metamorphosis
The liver size, hepatocyte diameter and hepatic fat content did not decrease immediately after the onset of metamorphic climax (after stage 41), and the histological morphology was unchanged from stage 36 to stage 43 (Fig. 9). When the tail of the froglet was absorbed to a stub (stage 44), rapid mobilization of hepatic fat was observed, characterized by a reduction of liver size and hepatocyte diameter, clearance of hepatic fat and higher degree of hepatic vascularization (Fig. 9).

Morphological and histological changes of liver during metamorphosis. Black arrows indicate blood sinus
Starvation accelerated the appearance of metamorphic traits in sufficiently fed tadpoles at stage 30–31, but not in insufficiently fed ones, even though they had comparable initial body and tail weights (Fig. 10a-j). These metamorphic traits included a dramatic reduction of body size and tail weight and an evacuated and shortened intestine (Fig. 10a-j & Additional file 3: Figure S3). Some individuals also showed tail apoptosis and accelerated development of the hind leg (Additional file 3: Figure S3). Sufficiently fed tadpoles had larger initial liver weight and higher HSI and HTI than the insufficiently fed ones. The contents of hepatic protein, triglycerides and glycogen in sufficiently fed tadpoles were 1.53-fold (p < 0.001), 2.43-fold (p < 0.001) and 2.83-fold (p = 0.028), respectively, higher than those in sufficiently fed tadpoles, while the relative contents of hepatic protein, triglycerides and glycogen in sufficiently fed tadpoles were 0.72-fold (p < 0.01), 1.44-fold (p = 0.039) and 1.10-fold (p = 0.72) of those in insufficiently fed tadpoles.

Different responses of sufficiently and insufficiently fed tadpoles to starvation. a Variation of body indexes during starvation at 20 °C. b Variation of body indexes during starvation at 8 °C. Each value represents the mean ± standard error (n = 6-13). Different letters indicate significant differences between values (p < 0.05), as shown by the Student–Newman–Keuls post hoc test after one-way ANOVA. c Comparison of the absolute and relative content of hepatic reserves between sufficiently and insufficiently fed tadpoles. Each column represents mean ± SE (n = 5-7), ***: p < 0.001, **: p < 0.01, *: p < 0.05
Discussion
Using liver as the primary fat depot has not been previously reported for tetrapods. One of the most well-known cases of hepatic fat deposition in tetrapods is ‘foie gras’, the fatty liver of domestic Palmipedes, i.e., ducks and geese. In geese overfed with a carbohydrate-rich diet, the liver weight may increase more than 10-fold, or up to 10% of body weight [43], and their hepatic fat can be comparable to their abdominal fat in content [13]. Even so, the liver of geese should not be as significant as their adipose tissues in fat storage because their artificial fatty liver resulted from compulsive lipogenesis due to excessive carbohydrate intake and was impelled by the limited capacity of fat exportation from liver to peripheral WAT [13]. Normally fed geese store most of their fat in WAT rather than liver [13]. Similarly, although hepatic fat storage is commonly observed in some adult amphibians and reptiles, their liver probably functions more in lipid metabolism (i.e., synthesis, degradation) than in storage [9]. Like hagfish, sharks and cods, R. omeimontis tadpoles (and some other frog species) were devoid of fat-accumulated adipose tissues across their larval stage (Fig. 2a & c & g), and thus their liver should be the indispensable fat depot. The hepatic fat deposition phenomenon observed in R. omeimontis tadpoles was not an artifact, as was evidenced in individuals caught in the field. Accordingly, to the best of our knowledge, this was the first observation that liver can be the primary fat depot in tetrapods.
Potential protective mechanisms against hepatic injury in R. omeimontis tadpoles
Rana omeimontis tadpoles store body fat mainly in their liver (Fig. 1). The histological morphology of their fatty liver is characterized by inflated polyhedral hepatocytes filled by lipids. It differs from the histology of pathological fatty liver in two ways. First, hepatic fat in pathological fatty liver presents as round droplets, which usually inflate and sphericize hepatocytes, while hepatic fat in R. omeimontis tadpoles is shaped into polygons by the hepatocytes. Second, the lipid droplets in pathological fatty liver vary greatly in size, resulting in obvious morphological asymmetry among hepatocytes. This is consistent with previous studies showing a similar histological morphology of Atlantic cod liver [44]. It seems that naturally fatty livers can better manage their hepatic fat than pathological livers, which could be a potential strategy to avoid liver injuries caused by fat deposition. To some extent, the histology of R. omeimontis tadpoles resembles WAT in cell morphology. This makes sense, as fat deposition in the liver is likely a primitive trait in vertebrate evolution [2]. This finding also implied that WAT and primitive fatty liver may share similar molecular strategies for cellular fat management. It is interesting to speculate that the liver has lost the capacity to manage massive cellular lipids during the evolution of vertebrates. In future studies, comparative lipidomics and proteomics should be conducted to investigate the differences among lipid droplets from naturally fatty liver, from pathological fatty liver, and from WAT.
In the progression of nonalcoholic steatosis hepatitis in humans, lipotoxicity can induce initial hepatic injuries, which are then amplified by inflammation [14]. However, hepatic lipid accumulation is not inevitably accompanied by lipotoxicity, whose occurrence depends on the efficiency of incorporating FFA into triglycerides and the composition of intermediates of lipid metabolism in liver [33, 45]. Previous studies have indicated that the ratio of monounsaturated FFA to saturated FFA determines whether liver cells are damaged by the flux of FFA in mammals, and a higher ratio indicates a lower risk of hepatic stress [45, 46]. R. omeimontis tadpoles kept low proportions of saturated FFAs (1.98% for palmate and 1.28% for stearate), but they exhibited much higher proportions of monounsaturated ones (19.5% for oleate and 12.99% for palmitate) in their liver FFA pool (Fig. 3c). Such a metabolic trait is expected to be associated with a low risk of lipotoxicity. Abundant free arachidonic acid is another prominent feature of the hepatic FFA pool of R. omeimontis tadpoles (Fig. 3c). This FFA has been suggested as a protective metabolite against lipotoxicity induced by saturated FFAs through its diversion of saturated FFAs into triglycerides [47,48,49]. As the precursor of both proinflammatory and anti-inflammatory eicosanoids [50], arachidonic acid may also protect the liver from damage by regulating inflammation. Overall, the FFA composition in R. omeimontis tadpole liver is beneficial to avoid lipotoxicity and inflammation. Interestingly, neither a high abundance of arachidonic acid nor a high ratio of monounsaturated FFAs to saturated FFAs was observed in the tail metabolome of these tadpoles (Additional file 3: Figure S1 C), suggesting that these metabolic traits are specific to the liver. We speculated that two approaches might be involved in maintaining this unique hepatic FFA profile. One possible approach is effective FFA desaturation in the liver. So far, many studies have indicated that the activity of stearoyl-CoA desaturase-1, the enzyme that converts saturated FFAs to unsaturated FFAs, is associated with the susceptibility of hepatocytes to lipoapoptosis induced by saturated FFAs [30, 51]. It will be meaningful to determine the expression level of stearoyl-CoA desaturase-1, as well as its upstream regulators, in hepatocytes of R. omeimontis tadpoles. Another possible approach is the selective transport of FFAs to peripheral organs [52]. In contrast to the situation in the liver, the proportions of saturated FFAs in the tail are much higher than those of monounsaturated FFAs (Additional file 3: Figure S1 C). Considering that tail FFAs also come from the liver, the huge difference in FFA profile between liver and tail imply selective export of saturated FFAs from the liver to other organs. Such a mechanism, if confirmed in the liver of R. omeimontis tadpoles, is meaningful in the treatment of pathological fatty liver.
Metabolic traits of fatty liver of R. omeimontis tadpoles
There are several prominent differences in the metabolic profile between R. omeimontis tadpoles and other invertebrates. The first is an unusually high abundance of maltotriose but a low abundance of glucose (the common blood sugar of vertebrates) in the liver and tail of R. omeimontis tadpoles (Fig. 3b). We measured the metabolic composition of spirulina and excluded the tadpole diet as the source of maltotriose (Supplementary data). We speculate that maltotriose may also exist in the circulation of R. omeimontis tadpoles. Previous studies have indicated low blood sugar in amphibians, which usually have blood sugar levels below 25 mg/dl [53]. The values in other vertebrates, including lamprey, elasmobranchs, bony fish, birds and mammals, range between 45 and 200 mg/dl [54]. If disaccharides or trisaccharides indeed accounted for a considerable proportion of blood sugars in amphibians, the total carbohydrate amount would be comparable to that in other vertebrates. This is because the methods used to measure the concentration of blood sugar in these studies were either specific to glucose or indiscriminate to reducing sugars. Therefore, both methods could underestimate the mass of disaccharides and trisaccharides in blood. It would be interesting to know why R. omeimontis tadpoles use maltotriose as their major soluble sugar in liver and whether it is associated with their hepatic fat deposition phenomenon. One plausible explanation is that the hydrophilic space in their hepatocytes is scarce, and maltotriose is more efficient than glucose (at the same concentration) at transporting carbons. The second metabolic difference between R. omeimontis tadpoles and other vertebrates is the existence of numerous and abundant dipeptides in liver (Fig. 3d). Although some dipeptides, such as carnosine and anserine, are common in vertebrate tissues such as muscle and brain [55, 56], these metabolites have not been reported as the major storage form of soluble amino acids in vertebrates. In liver of R. omeimontis tadpoles, some amino acids, including isoleucine, alanine, serine and glycine, are only stored in the form of dipeptides. This metabolic trait is seemingly specific to liver, as the tail has a much lower proportion of dipeptides and fewer dipeptide types. The physiological significance of dipeptides in liver of R. omeimontis tadpoles is still unknown. We may again speculate that their prevalence is related to the scarce hydrophilic space in hepatocytes.
Though elasmobranchs, Atlantic cod and R. omeimontis tadpoles share the same fat storage strategy, the utilization orders of their fat reserves are quite different. In elasmobranchs, their hepatic fat is neither a prior metabolic fuel nor a prior substrate for ketogenesis to amino acids [57]. In Atlantic cod, fat is only a metabolic fuel at the beginning of a fasting period, a phenomenon that is also observed in bony fish with extrahepatic fat depots [7, 37, 44, 58]. In R. omeimontis tadpoles, their liver glycogen and fat are mobilized simultaneously at 20 °C, but glycogen is exhausted more rapidly than fat (Fig. 4). After 10 days of starvation, the decrements in hepatic protein and amino acids were less severe than that of triglycerides or FFAs (Figs. 4, 6 and 8). These results suggested a sequential utilization of hepatic reserves in R. omeimontis tadpoles: glycogen > lipids > protein/amino acids. This order is in accordance with those in fasted frogs [59,60,61,62] and mammals [63, 64]. Their carbohydrates are mainly used at the beginning of a long-term fasting period, and then they depend on their lipids and protein for energy production. For R. omeimontis tadpoles, temperature could influence the utilization of hepatic reserves. At 8 °C, glycogen was preferentially consumed over fat. Such a metabolic change might result from cold-induced suppression of aerobic metabolism, which has been reported in overwintering amphibians. This is because they can suppress their aerobic metabolism to cope with cold temperatures [29, 65]. As oxidation of FFAs is an obligative aerobic process, fat mobilization is likely more effectively inhibited. The huge gap in consumption rates (more than 3-fold for all three hepatic reserves, Fig. 4) between tadpoles fasted at 20 and 8 °C also implied the existence of metabolic suppression. It should be noted that a decrease in glycogen (to less than 4% of liver weight) could not explain the reduction of liver weight (more than 30% of liver weight) in R. omeimontis tadpoles fasted at 8 °C (Fig. 4). According to our observation, the reduction in blood volume, induced by starvation and evidenced by the accumulation of biliverdin (a product of heme catabolism, Fig. 4a), should make a great contribution.
How reserves are allocated among tissues is also an important aspect of metabolism. In animals with extrahepatic depots, FFAs released from these depots are mainly used by liver and muscle during starvation. This was also the case in R. omeimontis tadpoles, as the increment of carnitine and acyl-carnitine concentrations suggested enhanced lipid catabolism in both liver and muscle after starvation (Fig. 6c-d). Unlike these metabolic intermediates, fatty acids were part of lipid storage in liver of R. omeimontis tadpoles, which can be expected to reduce in liver (Fig. 6a). Despite the dramatic decrement in hepatic FFAs detected after 10 days of starvation, the levels of major tail FFAs were maintained (Fig. 6b). This was because the tail was a major consuming organ, which could import fatty acids from the blood circulation. In the tail, the concentrations of arachidonic acid and oleic acid, the two FFAs with the highest abundance in the liver, increased significantly after 10 days of starvation. Palmitic acid, cis-9-palmitoleic acid, linolenic acid and heptadecanoic acid, which also had relatively high abundance in the liver, decreased significantly after 10 days of starvation. Such a variation pattern supported the encouraged transport of hepatic FFAs to the tail and the increased utilization of FFAs in the tail. In fasted mammals, glycogenolysis and gluconeogenesis based on glycerol and amino acids were activated in their liver to maintain the levels of blood sugar. The maintenance of a high amount of hepatic soluble sugars, even 5 days after glycogen exhaustion, suggesting that gluconeogenesis was activated in the liver of fasted R. omeimontis tadpoles (Fig. 7a). Therefore, carbohydrates were unlikely metabolic fuels in the liver of fasted tadpoles. As a major consuming organ, the tail was likely a destination of the carbohydrate from the liver, as its levels of glycolytic intermediates remained unchanged even 5 days after glycogen exhaustion (Fig. 7d). In addition, the variation pattern of tail soluble carbohydrates, which could be explained by the encouraged hydrolysis of oligosaccharides to monoses (Fig. 7c), could also suggest the consumption of carbohydrates in the tail. Considering that liver is the primary site for gluconeogenesis and that amino acids are major substrates for gluconeogenesis, gluconeogenesis in liver likely accounted for the decrement of hepatic amino acids in fasted R. omeimontis tadpoles (Fig. 8a). In contrast to the liver, the tail had an overall increment in amino acid concentrations. We thought these variations likely resulted from protein degradation, a common metabolic response to long-term starvation in vertebrates [35]. It was interesting that the concentrations of alanine and glutamine, two major nontoxic interorgan ammonia carriers [66, 67], had the most prominent increments (Fig. 8b), implying catabolism of amino acids in the tail. Taken together, these data suggest that lipids and possibly amino acids were used as metabolic fuel in the liver of fasted R. omeimontis tadpoles, while lipids, carbohydrates and amino acids were all used as metabolic fuel in their tail. In combination with the variation trends of metabolites in the TCA cycle, these results suggest that the metabolic flux throughout aerobic pathways was likely decreased and maintained in the liver and tail, respectively, after 10 days of starvation. It seemed that the tail had the priority of maintaining energy production to the liver during starvation. This is reasonable, as a large proportion of energy produced in liver is used for anabolism, which is likely suppressed in starvation, while locomotion is always required for animals in foraging and escaping.
Overall, R. omeimontis tadpoles have some unique features in their liver metabolome, and these features likely correspond to their liver-based fat deposition. R. omeimontis tadpoles share similar metabolic strategies with amphibians and mammals that have extrahepatic fat depots. This implies that the fatty liver of R. omeimontis tadpoles may be a primitive counterpart of mammal WAT.
Role of hepatic fat in metamorphic climax
Although hepatic fat of R. omeimontis tadpoles can be mobilized immediately during starvation (Fig. 4e), the onset of metamorphic climax, a nonfeeding period, was not accompanied by mobilization of hepatic fat (Fig. 9). This finding suggested that metabolic reserves other than hepatic fat supported energy production in metamorphosing R. omeimontis tadpoles. After the tail was absorbed to a stub, a dramatic decrease in hepatic fat was observed in these tadpoles, suggesting that apoptotic tissues were used as metabolic substrates in early metamorphic climax. Our results also indicated the existence of a metabolic shift in metamorphic climax, which is consistent with observations in the tadpole of the bullfrog (Lithobates catesbeianus), whose fat body lipid is also mobilized in late metamorphic climax [23]. Our results suggest that the liver or hepatic fat is functionally equal to the fat body or fat body lipid, respectively, in fueling metamorphosis. Importantly, the metabolic shift in metamorphic climax could reconcile the controversy over whether fat reserves are required for energy fuel in metamorphosing tadpoles. Although lipid reserves have been suggested to be reduced after metamorphosis [20], several studies have indicated that tadpoles with low lipid reserves can also complete metamorphosis [26, 68]. According to our results based on R. omeimontis tadpoles, fat reserves might not be necessary for the major stage of metamorphic climax, as organ remodeling was mainly completed when the tail of R. omeimontis tadpoles was absorbed to a stub. However, fat reserves should be important in supporting the energy metabolism in these froglets [23], as their physiological functions might be incompetent to ensure a predation success rate. Additionally, mobilization of fat reserve in late metamorphic climax might relieve the dependency on amino acids for catabolism and thus facilitate more amino acids flowing into anabolism. These speculations were supported by observations in salamanders, in which low lipid levels during metamorphosis were associated with a low survival rate of metamorphosed animals [25].
Environmental stress can accelerate the metamorphosis of tadpoles by inducing the release of corticosteroids [69,70,71]. Food deprivation is a common environmental stress in the wild, and its influence on the progression of tadpole metamorphosis is associated with developmental stage and external conditions [23, 72, 73]. It has been widely reported that food deprivation likely accelerates development in pro-metamorphic tadpoles in a condition-dependent manner [74,75,76,77,78]. Crespi and Denver [76] speculated that fat stores in tadpoles should have a role in modulating the strength of food deprivation-induced corticosterone signals and that tadpoles with abundant fat stores likely respond weakly to food deprivation. Our results also suggest a role for energy reserves in modulating accelerated metamorphosis induced by food deprivation. However, nutrient storage was positively correlated with the inducibility of accelerated metamorphosis in fasted R. omeimontis tadpoles (Fig. 10). We speculated that energy reserves could be a body condition signal that is independent of the corticosterone cascade in modulating metamorphosis. This is supported by observations in Xenopus laevis tadpoles, in which increased leptin expression in the fat body just before the onset of metamorphosis serves as a body condition signal to coordinate the feeding activity and the onset of metamorphic climax [24]. In our opinion, stress signals and body condition signals are both indispensable in mediating accelerated metamorphosis induced by food deprivation. This speculation may provide an explanation for the absence of accelerated metamorphosis in pro-metamorphic tadpoles suffering food deprivation [72, 73]. In our study, despite all three energy reserves (triglyceride, glycogen and protein) being higher in the livers of sufficiently fed tadpoles than in those of insufficiently fed ones, triglycerides were most likely responsible for this suppositional body condition signal in regulating metamorphosis for several reasons. First, triglycerides are the major hepatic energy reserve in R. omeimontis tadpoles, and they have a role in fueling metamorphic climax. Second, the liver triglyceride and glycogen contents in sufficiently fed tadpoles were both twice the values in insufficiently fed tadpoles, but the content of liver glycogen in sufficiently fed tadpoles varied greatly, exceeding a 90-fold difference between individuals. Lastly, liver triglycerides were the only reserve that had higher relative content in sufficiently fed tadpoles than insufficiently fed ones. Further studies are required to verify and characterize this suppositional body condition signal in R. omeimontis tadpoles, and it will be interesting to test whether leptin can still be a signal messenger of fat stores in the liver.
Conclusion
In this study, we reported a hepatic fat deposition phenomenon in R. omeimontis tadpoles. To our knowledge, this was the first observation that liver can be the primary fat depot in animals with a higher evolutionary status than bony fish. We found that this naturally fatty liver had special histological morphology and metabolic compositions, which likely either guard the liver against injuries or make hepatocytes of R. omeimontis tadpoles adapt to fat accumulation. During starvation, R. omeimontis tadpoles shared similar metabolic traits with frogs and mammals that have extrahepatic fat depots. This finding suggested that their fatty liver could be a primitive counterpart of mammal WAT. In addition, we found that the fatty liver of R. omeimontis tadpoles had roles in fueling and even modulating metamorphosis.
Availability of data and materials
The dataset supporting the conclusions of this article is included as additional file.
Abbreviations
- FFA:
-
Free fatty acid
- H & E:
-
Hematoxylin and eosin
- HSI:
-
Hepatosomatic index
- HTI:
-
Hepato-tail index
- PAS:
-
Periodic acid-schiff
- PC:
-
Principal component
- PCA:
-
Principal component analysis
- RO:
-
Red oil
- TG:
-
Triglyceride
- TLC:
-
Thin-layer chromatography
- WAT:
-
White adipose tissue
References
Sheridan MA. Lipid dynamics in fish: aspects of absorption, transportation, deposition and mobilization. Comp Biochem Physiol B. 1988;90:679–90.
Gesta S, Tseng YH, Kahn CR. Developmental origin of fat: tracking obesity to its source. Cell. 2007;131:242–56.
Patzner RA, Adam H. Changes in weight of the liver and the relationship to reproduction in the hagfish Myxine glutinosa (Cyclostomata). J Mar Biol Assoc U K. 1981;61:461–4.
Patzner RA. Cyclical changes in weight and fat content of the liver and their relationship to reproduction in the hagfish Eptatretus burgeri (Cyclostomata). Acta Zool. 1980;61:157–60.
Kott E. Liver and muscle composition of mature lampreys. Can J Zool. 1971;49:801–5.
Shin YC. Some observations on the fine structure of lamprey liver as revealed by electron microscopy. Okajimas Folia Anat Jpn. 1977;54:25–59.
Weil C, Lefe’vre F, Bugeon JM. Characteristics and metabolism of different adipose tissues in fish. Rev Fish Biol Fisheries. 2013;23:157–73.
Sheridan MA. Regulation of lipid metabolism in poikilothermic vertebrates. Comp Biochem Physiol B. 1994;107:495–508.
Fitzpatrick LC. Life history patterns of storage and utilization of lipids for energy in amphibians. Amer Zool. 1976;16:725–32.
Derickson WK. Lipid storage and utilization in reptiles. Am Zool. 1976;16:711–23.
Rose FL. Seasonal changes in lipid levels of the salamander Amphiuma means. Copeia. 1967;1967:662–6.
Liu L, Zhao X, Wang Q, Sun X, Xia L, Yang B, Zhang Y, Montgomery S, Meng H, Geng T, Gong D. Prosteatotic and protective components in a unique model of fatty liver: gut microbiota and suppressed complement system. Sci Rep. 2016;6:31763.
Hermier D, Guy G, Guillaumin S, Davail S, André J-M, Hoo-Paris R. Differential channelling of liver lipids in relation to susceptibility to hepatic steatosis in two species of ducks. Comp Biochem Physiol B: Biochem Mol Biol. 2003;135:663–75.
Tilg H, Moschen AR. Evolution of inflammation in nonalcoholic fatty liver disease: the multiple parallel hits hypothesis. Hepatology. 2010;52:1836–46.
Lau JKC, Zhang X, Yu J. Animal models of non-alcoholic fatty liver diseases and its associated liver cancer. In: Yu J, editor. Obesity, Fatty Liver and Liver Cancer. Singapore: Springer Singapore; 2018. p. 139–47.
Bush FM. Effects of light and temperature on the gross composition of the toad, Bufo fowleri. J Exp Zool. 1963;153:1–13.
Caroline MP. The evolution of mammalian adipose tissue. New York, Dordrecht, Heidelberg, London: Springer; 2012.
Gramapurohit NP, Shanbhag BA, Saidapur SK. Pattern of growth and utilization of abdominal fat bodies during larval development and metamorphosis in five south Indian anurans. Curr Sci. 1998;75:1188–93.
Jørgensen CB. Growth and reproduction. In: Feder ME, Burggren WW, editors. Environmental Physiology of the Amphibians. Chicago: The University of Chicago Press; 1992. p. 439–66.
Sheridan MA, Kao Y-H. Regulation of metamorphosis-associated changes in the lipid metabolism of selected vertebrates. Am Zool. 1998;38:350–68.
Brown G. The metabolism of Amphibia. New York: Academic Press; 1964.
Ryuzaki M, Oonuki M. Changes in lipid composition in the tail of Rana catesbeiana larvae during metamorphosis: biochemistry. Zool Sci. 1990;7:409–17.
Wright ML, Richardson SE, Bigos JM. The fat body of bullfrog (Lithobates catesbeianus) tadpoles during metamorphosis: changes in mass, histology, and melatonin content and effect of food deprivation. Comp Biochem Physiol A Mol Integr Physiol. 2011;160:498–503.
Bender MC, Hu C, Pelletier C, Denver RJ. To eat or not to eat: ontogeny of hypothalamic feeding controls and a role for leptin in modulating life-history transition in amphibian tadpoles. Proc R Soc B. 2018;285:20172784.
Scott DE, Casey ED, Donovan MF, Lynch TK. Amphibian lipid levels at metamorphosis correlate to post-metamorphic terrestrial survival. Oecologia. 2007;153:521–32.
Bouchard SS, O'Leary CJ, Wargelin LJ, Charbonnier JF, Warkentin KM, Moore I. Post-metamorphic carry-over effects of larval digestive plasticity. Funct Ecol. 2016;30:379–88.
Milone M, Caliendo MF, Rastogi RK, Chieffi G. Seasonal lipid composition in the liver, fat body and gonads of Rana esculenta. Bolletino Di Zoologia. 2009;50:227–34.
Schlaghecke R, Blüm V. Seasonal variations in liver metabolism of the green frog Rana esculenta (L.). Experientia. 1978;34:456–7.
Donohoe PH, West TG, Boutilier RG. Respiratory, metabolic, and acid-base correlates of aerobic metabolic rate reduction in overwintering frogs. Am J Physiol. 1998;274:R704.
Tiniakos DG, Vos MB, Brunt EM. Nonalcoholic fatty liver disease: pathology and pathogenesis. Annu Rev Pathol. 2010;5:145–71.
Lau JKC, Zhang X, Yu J. Animal models of non-alcoholic fatty liver disease: current perspectives and recent advances. J Pathol. 2017;241:36–44.
Nanji AA. Animal models of nonalcoholic fatty liver disease and steatohepatitis. Clin Liver Dis. 2004;8:559–74.
Alkhouri N, Dixon LJ, Feldstein AE. Lipotoxicity in nonalcoholic fatty liver disease: Not all lipids are created equal. Expert Rev Gastroenterol Hepatol. 2009;3:445–51.
Cheung O, Sanyal AJ. Abnormalities of lipid metabolism in nonalcoholic fatty liver disease. Seminars in Liver Disease. 2008;28:351–9.
McCue MD. Starvation physiology: reviewing the different strategies animals use to survive a common challenge. Comp Biochem Physiol A Mol Integr Physiol. 2010;156:1–18.
Hamlett WC. The elasmobranch liver. A model for chemical carcinogenesis studies in a ‘naturally resistant’ vertebrate. Dordrecht: Springer Netherlands; 1988.
Black D, Love RM. The sequential mobilisation and restoration of energy reserves in tissues of Atlantic cod during starvation and refeeding. J Comp Physiol B. 1986;156:469–79.
Fei L, Ye CY, Jiang JP. Colored atlas of Chinese amphibians. China: Sichuan Publishing Group, Sichuan Publishing House of Science & Technology; 2010.
Gosner KL. A simplified table for staging anuran embryos and larvae with notes on identification. Herpetologica. 1960;16:183–90.
Epple A, Marques M, Miller SB. Fast and simple identification of adipose tissues in situ with dithizon. Anat Rec. 1974;180:341–50.
R Team C. Team RDC.R: A Language And Environment For Statistical Computing. Vienna: R Foundation for Statistical Computing; 2012.
Wickham H. ggplot2: Elegant Graphics for Data Analysis. USA: Springer-Verlag New York; 2016.
Hermier D, Rousselot-Pailley D, Peresson R, Sellier N. Influence of orotic acid and estrogen on hepatic lipid storage and secretion in the goose susceptible to liver steatosis. Biochim Biophys Acta. 1994;1211:97–106.
Kjær MA, Vegusdal A, Berge GM, Galloway TF, Hillestad M, Krogdahl Å, Holm H, Ruyter B. Characterisation of lipid transport in Atlantic cod (Gadus morhua) when fasted and fed high or low fat diets. Aquaculture. 2009;288:325–36.
Li ZZ, Berk M, McIntyre TM, Feldstein AE. Hepatic lipid partitioning and liver damage in nonalcoholic fatty liver disease: role of stearoyl-CoA desaturase. J Biol Chem. 2009;284:5637–44.
Neuschwander-Tetri BA. Hepatic lipotoxicity and the pathogenesis of nonalcoholic steatohepatitis: the central role of nontriglyceride fatty acid metabolites. Hepatology. 2010;52:774–88.
Cheon HG, Cho YS. Protection of palmitic acid-mediated lipotoxicity by arachidonic acid via channeling of palmitic acid into triglycerides in C2C12. J Biomed Sci. 2014;21:13.
Cho YS, Chi HK, Kim KY, Cheon HG. Protective effects of arachidonic acid against palmitic acid-mediated lipotoxicity in HIT-T15 cells. Mol Cell Biochem. 2012;364:19–28.
Xiao YJ, Yi-Ming MU, Zhan XR, Ming LI, Zu-Qian LU, Ju-Ming LU. Via activation of calcium channel system arachidonic acid protects ECV-304 endothelial cells against lipotoxicity induced by saturated fatty acids. Chinese J Endocrinol Metab. 2005;21:269–70.
Meng H, McClendon CL, Dai Z, Li K, Zhang X, He S, Shang E, Liu Y, Lai L. Discovery of novel 15-lipoxygenase activators to shift the human arachidonic acid metabolic network toward inflammation resolution. J Med Chem. 2016;59:4202–9.
Leamy AK, Egnatchik RA, Young JD. Molecular mechanisms and the role of saturated fatty acids in the progression of non-alcoholic fatty liver disease. Prog Lipid Res. 2013;52:165–74.
Polozova A, Salem N. Role of liver and plasma lipoproteins in selective transport of n-3 fatty acids to tissues: a comparative study of 14 C-DHA and 3 H-oleic acid tracers. J Mol Neurosci. 2007;33:56–66.
Wright PA. Blood sugar studies in the bullfrog, Rana catesbiana. Endocrinology. 1959;64:551.
Umminger BL. Relation of whole blood sugar concentrations in vertebrates to standard metabolic rate. Comp Biochem Physiol Part A: Physiol. 1977;56:457–60.
Boldyrev AA, Dupin AM, Pindel EV, Severin SE. Antioxidative properties of histidine-containing dipeptides from skeletal muscles of vertebrates. Comp Biochem Physiol B. 1988;89:245–50.
Abe H. Chapter 14 Histidine-related dipeptides: distribution, metabolism, and physiological function. Biochem Mol Biol Fishes. 1995;4:309–33.
Speers-Roesch B, Treberg JR. The unusual energy metabolism of elasmobranch fishes. Comp Biochem Physiol A Mol Integr Physiol. 2010;155:417–34.
Soengas JL, Strong EF, Fuentes J, Veira JAR, Andrés DMD. Food deprivation and refeeding in Atlantic salmon, Salmo salar: effects on brain and liver carbohydrate and ketone bodies metabolism. Fish Physiol Biochem. 1996;15:491–511.
Naya DE, Veloso C, Sabat P, Bozinovic F. The effect of short- and long-term fasting on digestive and metabolic flexibility in the Andean toad, Bufo spinulosus. J Exp Biol. 2009;212:2167–75.
van Beurden EK. Energy metabolism of dormant Australian water-holding frogs (Cyclorana platycephalus). Copeia. 1980;1980:787–99.
Baic D, Ladewski BG, Frye BE. Quantitative ultrastructural studies of hepatocytes from fed and starved frogs. J Exp Zool. 1979;210:381–405.
Duveau A, Piery Y. Prolonged starvation and ultrastructure of the frog liver. C R Seances Soc Biol Fil. 1973;167:491.
Viscarra JA, Ortiz RM. Cellular mechanisms regulating fuel metabolism in mammals: role of adipose tissue and lipids during prolonged food deprivation. Metabolism. 2013;62:889–97.
Cahill GF. Fuel metabolism in starvation. Annu Rev Nutr. 2006;26:1–22.
Boutilier RG. Mechanisms of metabolic defense against hypoxia in hibernating frogs. Respir Physiol. 2001;128:365–77.
Nurjhan N, Bucci A, Perriello G, Stumvoll M, Dailey G, Bier DM, Toft I, Jenssen TG, Gerich JE: Glutamine: a major gluconeogenic precursor and vehicle for interorgan carbon transport in man. J Clin Investig 1995, 95:272–277.
Hakvoort TBM, He Y, Kulik W, Vermeulen JLM, Duijst S, Ruijter JM, Runge JH, Deutz NEP, Koehler SE, Lamers WH. Pivotal role of glutamine synthetase in ammonia detoxification. Hepatology. 2017;65:281–93.
Beck CW, Congdon JD. Energetics of metamorphic climax in the southern toad (Bufo terrestris). Oecologia. 2003;137:344–51.
Kulkarni SS, Buchholz DR. Corticosteroid signaling in frog metamorphosis. Gen Comp Endocrinol. 2014;203:225–31.
Denver RJ. Chapter seven - Neuroendocrinology of amphibian metamorphosis. In: Shi Y-B, editor. Current Topics in Developmental Biology. Volume 103. New York: Academic Press; 2013. p. 195–227.
Denver RJ. Hormonal correlates of environmentally induced metamorphosis in the western spadefoot toad, Scaphiopus hammondii. Gen Comp Endocrinol. 1998;110:326–36.
Enriquez-Urzelai U, San Sebastian O, Garriga N, Llorente GA. Food availability determines the response to pond desiccation in anuran tadpoles. Oecologia. 2013;173:117–27.
Audo MC, Mann TM, Polk TL, Loudenslager CM, Diehl WJ, Altig R. Food deprivation during different periods of tadpole (Hyla chrysoscelis) ontogeny affects metamorphic performance differently. Oecologia. 1995;103:518–22.
D'Angelo SA, Gordon AS, Charipper HA. The role of the thyroid and pituitary glands in the anomalous effect of inanition on amphibian metamorphosis. J Exp Zool. 1941;87:259–77.
Denver RJ, Mirhadi N, Phillips M. Adaptive plasticity in amphibian metamorphosis: response of Scaphiopus hammondii tadpoles to habitat desiccation. Ecology. 1998;79:1859–72.
Crespi EJ, Denver RJ. Roles of stress hormones in food intake regulation in anuran amphibians throughout the life cycle. Comp Biochem Physiol A Mol Integr Physiol. 2005;141:381–90.
Morey S, Reznick D. A comparative analysis of plasticity in larval development in three species of spadefoot toads. Ecology. 2000;81:1736–49.
Boorse GC, Denver RJ. Endocrine mechanisms underlying plasticity in metamorphic timing in spadefoot toads. Integr Comp Biol. 2003;43:646–57.
Acknowledgements
Thanks to Guocheng Shu, Xungang Wang and Chengxin Hu from Chengdu Institute of Biology, Chinese Academy of Science for helping collecting egg clutches in the field.
Funding
This study was financially supported by the Strategic Priority Research Program of the Chinese Academy of Sciences, Grant No. XDA19050201, National Natural Science Foundation of China (31900327, 31700353), Construction of Basic Conditions Platform of Sichuan Science and Technology Department (2019JDPT0020), Important Research Project of Chinese Academy of Sciences (KJZG-EW-L13), Western Light Talent Culture Project of the Chinese Academy of Sciences (2016XBZG_XBQNXZ_B_008).
Author information
Authors and Affiliations
Contributions
WZ, JJ, TZ and HZ conceived the study. WZ, MZ, LC and WZ conducted the experiments and analyzed the data. WZ and MZ drafted the manuscript. WZ, MZ, JJ, TZ, CL and FX wrote the paper. All authors reviewed, edited and approved the final version of the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
All animal work in the present study was conducted according to relevant national and international guidelines. All animal care and experimental procedures were approved by the Chengdu Institute of Biology Animal Care and Use Committee [CIB2015003].
Consent for publication
All data generated or analyzed during this study are included in this published article [and its supplementary information files].
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Additional file 1.
Liver and tail metabolite tables.
Additional file 2.
Numeric data used in graphs.
Additional file 3: Figure S1.
Metabolic profile of the tail of R. omeimontis tadpoles (Stage 30–31). (A) Metabolites with an abundance higher than 1% of all identified metabolites. (B) Profile of identified soluble carbohydrates. (C) Profile of identified FFAs. (D) Profile of identified amino acids. (E) Proportion of free amino acids and dipeptides in the total amino acid pool. Each column in these two figures represents the mean ± SE of 7 samples. Figure S2. Variation of intermediates in the TCA cycle after 10 days of starvation in liver (A) and tail (B). Each column represents mean ± SE (n = 5–7), ***: p < 0.001, **: p < 0.01, *: p < 0.05. Figure S3. Sufficiently fed tadpoles showed traits of accelerated metamorphosis after starvation. Major morphological traits of accelerated metamorphosis included evacuated and shortened intestine, tail apoptosis (rounded tail) and accelerated development of hind limb (toe development). These three traits did not appear in any tadpoles before starvation treatment (either sufficiently fed or insufficiently fed ones), as well as in insufficiently fed tadpoles after starvation.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Zhu, W., Zhang, M., Chang, L. et al. Characterizing the composition, metabolism and physiological functions of the fatty liver in Rana omeimontis tadpoles. Front Zool 16, 42 (2019). https://doi.org/10.1186/s12983-019-0341-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12983-019-0341-x