
Abstract
Background
Increasing nitric oxide bioavailability may induce physiological effects that enhance endurance exercise performance. This review sought to evaluate the performance effects of consuming foods containing compounds that may promote nitric oxide bioavailability.
Methods
Scopus, Web of Science, Ovid Medline, EMBASE and SportDiscus were searched, with included studies assessing endurance performance following consumption of foods containing nitrate, L-arginine, L-citrulline or polyphenols. Random effects meta-analysis was conducted, with subgroup analyses performed based on food sources, sex, fitness, performance test type and supplementation protocol (e.g. duration).
Results
One hundred and eighteen studies were included in the meta-analysis, which encompassed 59 polyphenol studies, 56 nitrate studies and three L-citrulline studies. No effect on exercise performance following consumption of foods rich in L-citrulline was identified (SMD=-0.03, p=0.24). Trivial but significant benefits were demonstrated for consumption of nitrate and polyphenol-rich foods (SMD=0.15 and 0.17, respectively, p<0.001), including performance in time-trial, time-to-exhaustion and intermittent-type tests, and following both acute and multiple-day supplementation, but no effect of nitrate or polyphenol consumption was found in females. Among nitrate-rich foods, beneficial effects were seen for beetroot, but not red spinach or Swiss chard and rhubarb. For polyphenol-rich foods, benefits were found for grape, (nitrate-depleted) beetroot, French maritime pine, Montmorency cherry and pomegranate, while no significant effects were evident for New Zealand blackcurrant, cocoa, ginseng, green tea or raisins. Considerable heterogeneity between polyphenol studies may reflect food-specific effects or differences in study designs and subject characteristics. Well-trained males (V̇O2max ≥65 ml.kg.min-1) exhibited small, significant benefits following polyphenol, but not nitrate consumption.
Conclusion
Foods rich in polyphenols and nitrate provide trivial benefits for endurance exercise performance, although these effects may be food dependent. Highly trained endurance athletes do not appear to benefit from consuming nitrate-rich foods but may benefit from polyphenol consumption. Further research into food sources, dosage and supplementation duration to optimise the ergogenic response to polyphenol consumption is warranted. Further studies should evaluate whether differential sex-based responses to nitrate and polyphenol consumption are attributable to physiological differences or sample size limitations.
Other
The review protocol was registered on the Open Science Framework (https://osf.io/u7nsj) and no funding was provided.
Similar content being viewed by others


Background
Nitric oxide (NO) is a signalling molecule that is involved in numerous physiological processes including skeletal muscle contraction [1], endothelial function [2], mitochondrial biogenesis and respiration [3], muscle repair [4] and antioxidant defences [5,6,7]. Given these functions are important during exercise, particularly aerobic exercise, there has been considerable interest in enhancing NO production to improve endurance exercise performance. Specifically, increased synthesis of NO is proposed to reduce the oxygen cost of ATP resynthesis, lower the ATP cost of cross-bridge formation and promote vasodilation, thereby enhancing skeletal muscle blood flow and oxygen perfusion which may speed oxygen uptake kinetics [8]. These effects may translate to increased endurance exercise performance by improving exercise efficiency, decreasing the oxygen deficit at exercise onset and reducing the V̇O2 slow component [9].
NO is synthesised through two pathways, one of which is nitric oxide synthase (NOS)-dependent and one which is independent of NOS. NO is synthesised via the NOS-dependent pathway from L-arginine and oxygen in a reaction that is catalysed by various NOS enzymes, including endothelial nitric oxide synthase (eNOS). L-citrulline, an α-amino acid, also contributes to NO synthesis through this NOS-dependent pathway via its conversion to L-arginine. Polyphenols may enhance NO production by increasing eNOS expression and activity [10] and promote NO bioavailability via their antioxidant effects protecting NO from breakdown by reactive oxygen species (ROS). Within the NOS-independent pathway, nitrate (NO3-) is reduced to nitrite (NO2-) and then NO. Thus, it is proposed that NO availability may be improved by increasing the availability of NO3-, NO2-, L-arginine, L-citrulline, or polyphenols. Such changes in NO bioavailability are typically evaluated by plasma or urine NO3- and NO2-, as these are end products of endogenous NO production, but can also be inferred via measures of vascular function such as flow-mediated dilation (FMD).
In addition to the effects of polyphenols on NO synthesis via the NOS-independent pathway, their antioxidant properties have also been proposed to promote exercise performance by helping to maintain redox balance [11]. While the increase in ROS during exercise is an important signalling component that can facilitate acute responses and chronic adaptations to exercise, an imbalance between oxidative stress and antioxidant capacity may lead to impairment in blood flow, calcium handling and sensitivity, and central neural drive [11,12,13]. These derangements may contribute to the development of fatigue during exercise, so increasing antioxidant capacity may assist in inhibiting the onset of fatigue and enhance athletic performance. Thus, increased consumption of dietary polyphenols may enhance endurance exercise performance through effects on the NOS-dependent pathway and antioxidant capacity during exercise.
Augmentation of NO synthesis and bioavailability has been attempted by increasing the dietary intake of NO precursors. NO3- is abundant in beetroot and green leafy vegetables such as lettuce and spinach [14] and L-arginine is found in seafood, nuts, seeds, soy protein isolate and watermelon. Watermelon is also a rich source of L-citrulline, while fruits, vegetables, tea, coffee and cocoa are rich sources of polyphenols [15]. NO2-can be found in some fruits and vegetables, with more pronounced concentrations found in processed meats where it is used as an additive [16]. Many foods contain a combination of these nitric oxide precursors, as well as other antioxidant components such as vitamins, minerals and carotenoids, with interactions between these various phytochemicals and food sources potentially resulting in varied responses to specific foods and/or combinations of foods [17,18,19].
There appear to be considerable inter-individual differences in bioavailability in response to consumption of polyphenols [20] and NO3- [21, 22]. Several factors may influence the content and bioavailability of polyphenols in foods, such as storage and processing methods [23], the food matrix, and in particular, the profile of specific polyphenol subclasses (e.g. anthocyanidins vs quercetin) which vary significantly in their absorptive characteristics [20, 24]. Bioavailability is also affected by background diet, genetic factors, and particularly intestinal microbiota, as most polyphenols are catabolised by bacteria in the large intestine and the catabolites then enter the circulation and exert antioxidant effects [19, 25]. Similarly, the composition of NO3--rich food sources and microflora of the oral and gut microbiome may impact the bioavailability of NO3- from NO3--rich foods [22]. While increasing NO production may improve endurance exercise performance, performance effects may also be influenced by sex and fitness-related differences in NO synthesis [26, 27], vascular function [28, 29] and oxidative damage [30, 31]. Thus it is important to evaluate whether the effects of consuming foods that increase NO production on endurance exercise performance are influenced by these individual factors, as well as other parameters such as type of exercise performance test performed, the intensity at which the test is performed, and the dose and duration of consumption of NO precursors.
The primary aim of this systematic review and meta-analysis was to evaluate whether consumption of foods rich in precursors of NO synthesis improves endurance exercise performance. A secondary aim was to determine the effect of dose and duration of consumption of foods rich in NO precursors, participant characteristics (fitness and sex) and exercise test parameters (type and intensity) on exercise performance.
Methods
Information sources and search strategy
A literature search was conducted using the Scopus, Web of Science, Ovid Medline, EMBASE and SportDiscus databases on 5 September 2019. Title, abstract, keyword and MeSH (where applicable) searches were implemented.
The search terms are provided in Additional Material 1, but in brief, search terms were grouped under:
-
1.
Population (e.g. human, athlete).
-
2.
Compounds that promote nitric oxide synthesis (e.g. NO3-, polyphenols) and food sources of these compounds (e.g. beetroot juice, blackcurrant).
-
3.
Terms related to maximal endurance performance (e.g. exercise tolerance, time-trial).
Articles included in the search were exported to Endnote reference management software (Version 9, Clarivate Analytics, Philadelphia, PA), where duplicates were removed before remaining studies were uploaded to Covidence systematic review management software (Veritas Health Innovation, Melbourne, Australia).
Registration and protocol
A protocol application was submitted to PROSPERO on the 4th of September 2019, but we were informed that it was unsuccessful on the 29th January 2020 as it was not considered within their scope. Thereafter, the authors became aware of the Open Science Framework, where the study was registered on the 5th February 2020 (accessible at https://osf.io/u7nsj), which was after the title and abstract screening had been completed, but before full-text review had been completed by NMAdU or started by HTB.
Eligibility criteria
Studies were limited to human participants and English language, peer-reviewed studies, with no limit set on publication date. Studies needed to increase dietary intake of compounds that promote nitric oxide synthesis and bioavailability, with such compounds including polyphenols, NO2-, NO3-, L-citrulline and L-arginine, and adhere to the inclusion and exclusion criteria listed in Tables 1 and 2 respectively.
Study selection
During title and abstract screening, studies were excluded if they were not consistent with the inclusion and/or exclusion criteria. Antioxidant supplements or nutrients from an unspecified source were deemed eligible at this stage, as during preliminary searches it was noted that these types of articles sometimes specified a polyphenol food source from which the supplements or nutrients were obtained in full texts. Full-text articles were then screened to ensure that studies met the eligibility criteria. For inclusion in the meta-analysis, studies needed to report data that would enable calculation of a standardised mean difference (SMD).
Title and abstract screening and full-text review were conducted independently by two reviewers (NMAdU and other authors excluding JDB for title and abstract, NMAdU and HTB for full text), with disagreements regarding eligibility settled by a third reviewer (JDB). Where full texts were unavailable, they were sought from the authors, and reference lists were also pearled to identify additional relevant studies.
Data collection process and items
The data extraction form was drafted by the lead author (NMAdU), and studies were extracted in duplicate by NMAdU and HTB. Extracted information included date of publication, industry contributions, population, anthropometry, inclusion/exclusion criteria, supplement composition, dosing and duration of supplementation, performance test(s) and outcomes, dietary restriction of other foods or nutrients, and effects on biochemical and physiological markers of nitric oxide production.
Risk of bias assessment
Risk of bias in individual studies was assessed using the Revised Cochrane risk-of-bias tool for randomized trials [33] by the lead author (NMAdU) and a second author (HTB), with partial duplication to ensure consistency. Risk of bias was evaluated through visual inspection of standard and contoured funnel plots.
Effect measures
The effects of nitric-oxide related supplements were assessed using endurance exercise performance outcomes. Studies frequently reported multiple performance outcomes from the same performance test (e.g. mean power and time-to-complete reported for a time-trial). Therefore, where applicable, time-to-complete was the extracted outcome used for data analysis from fixed distance/work time-trials, while total distance/work was preferentially extracted for fixed duration time-trials. Likewise, peak power/speed was designated as the primary outcome from graded exercise tests, and total distance/work was extracted as the primary outcome for intermittent tests. Relative mean or peak power outputs were used in favour of absolute values.
Effect size was quantified as the SMD, where the mean difference between treatments was divided by the pooled standard deviation, with Hedge’s correction factor also applied to adjust for small sample sizes [34]. Where performance was assessed before the active and control interventions in parallel trials, SMD was instead calculated from mean change (from baseline) values and standard deviations. Where SD of these changes was unavailable, it was imputed for each group as \({SD}_{change}=\sqrt{SD_{baseline}^2+{SD}_{final}^2-\Big(2\times 0.80\times {SD}_{baseline}\times {SD}_{final}\Big)\kern1em }\) using a correlation coefficient of 0.80 (35, 36). A factor of -1 was applied to SMDs for time to complete to ensure consistent directions of effect against other outcomes. Where separate populations (e.g. male vs female), performance tests or supplementation protocols (e.g. different doses, durations, timings) were implemented within the same study, data were extracted as separate trials. Where standard deviations were not reported, these were calculated using standard error (SE), confidence intervals or p values, if available, per the Cochrane Handbook for Systematic Reviews of Interventions [37].
-
1.1.
Synthesis of results
Where precise or upper bounds of p-values (e.g. p<0.05) of mean differences were provided, standard error (SE) of effect size was calculated using the equivalent T-statistic and correlation coefficients between treatment groups were estimated in accordance with Elbourne et al [38]. Where these details were unavailable, SE was calculated in crossover trials as \(SE=\frac{1}{N}+\frac{SMD^2}{2N}\times \sqrt{2\left(1-0.72\right)}\), using the lowest correlation coefficient of 0.72 [39]. Effects were defined as trivial (<0.2), small (0.2–0.6), moderate (0.61–1.2), large (1.21–2.0), and very large (>2.0) [40] and precision of the effect size estimate was assessed using 95 % confidence intervals (CIs). Separate meta-analyses were undertaken for crossover and parallel trials.
Data from included studies were qualitatively synthesised. Where suitable data were available, random effects meta-analysis was conducted using STATA/IC software (version 16.1, College Station, TX, StataCorp LLC.) using the inverse variance model and restricted maximum likelihood method. Statistical heterogeneity between subgroups was assessed via Cochran’s Q, while heterogeneity within groups was evaluated by I2, with heterogeneity classified as low (I2 < 25%), moderate (I2 25–49%) or high (I2 > 50%) [41, 42]. Separate meta-analyses were conducted to investigate the effect of study design, blinding, sex, NO precursor (e.g. polyphenols, NO3-), food source, duration of consumption (i.e. acute vs multiple days), fitness level of study participants, and the mode, type (i.e. time to exhaustion vs time trial) and duration (from time trial and constant load tests only) of exercise performance tests.
For subgroup analyses, fitness was classified by performance level (PL) using the guidelines of De Pauw [43] and Decroix [44] using reported running and cycling maximal oxygen consumption (V̇O2max), as provided in Table 3 below. Mixed-sex trials that did not report separate V̇O2max values for females and males, or featured different PLs between the sexes, were excluded from this fitness subgroup analysis. The influence of fitness was also investigated using meta-regression of outcomes against male V̇O2max values.
Results
Study selection and characteristics
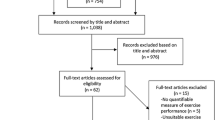
A PRISMA flow diagram of the search and screening results is displayed in Fig. 1. Of the 123 studies that were eligible for inclusion, 103 used a crossover design and 21 used a parallel design. 101 were reported as double-blind, 13 as single-blind, and 10 did not specify blinding. 62 studies were classified as polyphenol studies, 57 were NO3--focused and four studies tested L-citrulline effects, with 40 different food sources represented. While no included studies were specifically focused on L-arginine, it is an additional nutritional component found in watermelon and almonds, and studies that used these foods were classified as L-citrulline and polyphenol studies, respectively. Five studies did not report sufficient data to calculate SMD, thus the meta-analysis incorporated 118 studies. Eleven studies featured multiple performance tests, seven studies assessed the effects of various supplementation durations, five studies evaluated different supplementation dosages, three studies analysed multiple food sources and three studies analysed multiple populations. Thus, the 118 studies with eligible data were considered as 156 separate trials (i.e. with separate effect sizes).

PRISMA flow diagram of search, screening and inclusion outcomes
Data from 1872 participants, from 25 different countries were included, comprising 1482 males, 344 females and 46 participants of unspecified sex, as sex was not reported in four studies. From reported study means, pooled mean age was 27.8 ± 5.9 years, with a height of 177.4 ± 10.0 cm, mass of 74.5 ± 9.9 kg and V̇O2max of 53.9 ± 6.9 ml.kg.min-1. NO3- trials featured a daily dose of 8.4 ± 3.3 mmol NO3-, polyphenol trials had a daily total phenolic content of 817.4 ± 743.1 mg gallic acid equivalents (where reported), and L-citrulline trials reported a daily dose of 1.71 ± 1.0 g citrulline.
Risk of Bias
Risk of Bias results for each study (including specific domain of bias scores) are provided in Additional Material 2. Sixty-six studies had a high risk of bias, with 58 studies having ‘some concerns’ and no studies classified at low risk. Classification of high risk was most frequently given for deviations from intended interventions (53%). ‘Some concerns’ was predominant for bias regarding selective reporting of results (100%), reporting of missing outcome data (70%), and the randomisation process (88%).
Publication Bias
Standard and contoured funnel plots are displayed in Figure 2. There is some asymmetry within L-citrulline trials that is likely due to the limited number of studies, whereas NO3- and polyphenol trials appeared to have good symmetry overall and in the proportion of positive and negative non-significant effects, as assessed by the symmetry within the dark grey shaded areas. Given that 43 trials were designated as receiving financial support, subgroup analyses were also undertaken to help infer whether results from privately funded studies may have been suppressed in the absence of beneficial effects. However, these analyses revealed no significant differences between effects of NO3- studies that did or did not receive financial support (p=0.55), whereas financially supported polyphenol studies had significantly smaller effects (p=0.02) than those without private funding. Overall, these results suggest that publication bias did not play a significant role in the meta-analysis results.

Standard and contoured funnel plots of nitrate, polyphenol, and L-citrulline effects
Results of syntheses
Nitrate consumption (all food sources combined)
A summary of overall and sub-group analyses is provided in Table 4. Based on the analysis of 81 trials and 956 participants, NO3- consumption provided significant, trivial benefits for endurance exercise performance, and trials featured low heterogeneity (I2=6.4%). Significant benefits were evident in both crossover (trivial effects) and parallel (small effects) trials, but study design was not a moderator of effects, and results did not differ between study designs (Qb=2.37, p=0.12).
Effects by supplementation protocol
Sub-group analyses indicated that trivial benefits existed for both acute and multiple-day supplementation, with no significant difference between the two (p=0.08) and no significant influence of the number of supplementation days (p=0.12). Meta-regression indicated that daily NO3-dosage of interventions was not associated with performance effects overall (p=0.93), or within acute (p=0.62) and multiple-day (p=0.49) supplementation trials specifically. Sixteen trials restricted intake of other dietary NO3- sources, but effects within this subgroup were no different from trials without restrictions (Qb=0.55, p=0.46).
Sex and fitness-specific effects
Effects differed by classification of participant sex (Qb=6.94, p=0.03), as trivial benefits were still seen in male-only and mixed-sex studies, but females demonstrated no effects (SMD=0.0, p=0.98) from the results of only four trials. Effects differed by PL overall (Qb = 8.78, p=0.03), but when trials were restricted to males only these differences were not significant (Qb = 6.39, p=0.09) and meta-regression indicated that V̇O2max was not a significant mediator of the size of performance effects (p=0.10). Most trials included participants classified as PL2, who demonstrated small, significant benefits. Male PL2 (V̇O2max: 45-55 ml.kg.min-1) trials exhibited a small, significant improvement overall and within TTE trials specifically, but showed no change in TT performance and insignificant trivial benefits for GXT performance. Within the male PL3 bracket (V̇O2max: 55-65 ml.kg-1.min-1), a significant trivial improvement was seen for overall performance, which was still evident for TT performance, while small but insignificant improvements were observed for TTE performance and trivial insignificant effects were indicated for GXT and ITT trials. No effect was present for males in the PL4 (V̇O2max: 65-71 ml.kg-1.min-1) or PL5 brackets (V̇O2max >71 ml.kg-1.min-1) which were evaluated predominantly through TT performance. Female PL subgroup analyses were not conducted as data were unavailable from female PL2 or PL3 trials and other PLs only included a single study.
Effects by test type and duration
Although effects did not vary between performance test subgroups (Qb=10.63, p=0.059), non-significant trivial effects were evident for intermittent time trial (ITT) tests, while significant trivial effects were identified for intermittent graded exercise tests (IGXT), time trials (TT) and graded exercise tests (GXT). Small, significant improvements were found in time to exhaustion (TTE) tests, whereas small non-significant benefits were observed for intermittent time to exhaustion (ITTE) tests. There was no heterogeneity within IGXT, ITT or TT trials (I2=0%), but moderate heterogeneity was present in TTE (I2=29%) trials and high heterogeneity was present for GXT (I2=47%) and ITTE (I2=60%) trials. Effect sizes within various test duration brackets during TTs and TTE tests are provided in Table 4, and meta-regression found no influence of test duration on performance outcomes for TTs (p = 0.52) or TTE tests (p = 0.48). Since PL2 and PL3 for males were the only categories that demonstrated significant benefits overall, meta-regression was also performed for this subgroup specifically, which found no influence of duration on effects within TTs (p = 0.76) or TTE tests (p = 0.64).
Effects of different nitrate food sources
Sub-group analysis indicated that consumption of (NO3--rich) beetroot demonstrated a significant, trivial improvement in exercise performance (Fig. 3), with studies predominantly using beetroot juice as the delivery method (Additional Material 3). No significant effects were evident for consumption of red spinach or Swiss chard and rhubarb from a limited number of studies, but between-food effects were not statistically different between food sources of NO3- (Qb=1.03, p=0.60).

Nitrate supplementation forest plot. Abbreviations – %Δ, work rate that would achieve x% of difference between V̇O2 at gas exchange threshold and V̇O2peak; CI, confidence interval; GXT, graded exercise test; IGXT, intermittent graded exercise test; ITT, intermittent time-trial; ITTE, intermittent time to exhaustion; k, number of trials; km, kilometre; n, sample size; NO3-, nitrate; PL, performance level; Qb, between-group Q-statistic; rpm, revolutions per minute; s, second; SMD, standardised mean difference (Hedge’s g); supp., supplementation; TT, time-trial; TTE, time to exhaustion.
Polyphenol supplementation effects
The analysis of polyphenol effects included 71 trials and 1227 participants, indicating significant, trivial benefits for consumption of polyphenol-rich foods overall (Table 4), with these effects influenced by high heterogeneity (I2 = 66%). Forty-five (of 71) trials quantified the polyphenol content of the foods consumed, but the overall effects of these trials were not different from trials where polyphenol content was unspecified, and total phenolic content of the foods consumed did not influence effects (p=0.27). There were few differences in subgroup effects (parallel, GXT, TTE, PL1, PL2, and acute trials) in comparison to all polyphenol studies combined (Table 5), and given the similarities between studies that did and did not report polyphenol content, the results presented in-text represent all polyphenol trials. Crossover trials produced trivial, significant benefits, whereas small, insignificant improvements were shown in parallel trials, although subgroup analyses indicated that results between parallel and crossover trials were not significantly different (p=0.55).
Effects by supplementation protocol
Small, significant benefits were evident for acute supplementation and trivial, significant improvements were exhibited after multiple-day consumption, with high heterogeneity for both acute and multiple-day supplementation. Effects did not differ between acute and multiple-day supplementation (p=0.07), nor did the number of supplementation days appear to be a significant moderator of outcomes (p=0.47). Twenty-eight of 56 multiple-day studies provided an acute dose before testing, but effects were not different from multiple-day polyphenol trials that did not include a pre-testing dose (p=0.37). Seventeen trials restricted intake of foods rich in antioxidants and/or polyphenols, and subgroup analyses indicated no between-group differences with polyphenol trials that had no restrictions (Qb=0.74, p=0.39).
Effects by sex and fitness
Small, significant benefits were identified for studies with males only, with no effect in female or mixed-sex studies. Small, significant improvements were evident in males in the PL2, PL3 and PL4 brackets, with no eligible studies for PL5, and V̇O2max did not mediate performance effect sizes (p=0.76). Low heterogeneity (I2=0%) was evident for the male PL2 and PL4 brackets, while moderate heterogeneity (I2= 48%) was evident for male PL3 trials. Two female PL3 (V̇O2max: 48-52 ml.kg-1.min-1) trials indicated no effects, with no other female trials available to classify by PL.
Effects by duration and test type
Significant, trivial improvements were found for TT and TTE performance, and significant moderate improvements were evident for IGXT, with no significant effects for other test types. One IGXT study [100] likely included a reporting error (very low SD), but even with this trial removed, the effect within IGXTs remained moderate. No significant effects were identified for performance tests with durations or <5 minutes or 5-10 minutes, while trivial, significant benefits were seen in tests lasting 10-30 minutes and 30-60 minutes. Small, but significant performance decrements were seen in performance (all TT) with a duration of >60 minutes. Meta-regression did not indicate a significant influence of exercise duration within TTs (p=0.49) or TTE tests (p=0.40).
Polyphenol-rich foods
Trials evaluating effects of polyphenol-rich foods evaluated 36 separate food sources. Effects on exercise performance are displayed in Fig. 4, with significant differences between food sources (Qb=32.64, p=0.002). Moderate, significant benefits were exhibited for the consumption of grape (juice), small significant improvements were seen following consumption of French maritime pine bark extract and Montmorency cherry, and significant, trivial benefits were demonstrated for NO3--depleted beetroot and pomegranate. Small effects were found for the consumption of cocoa/chocolate, but this did not reach statistical significance (p=0.052). No effects (SMD<0.1) were evident for the consumption of American or Siberian ginseng, blueberry, green tea, New Zealand blackcurrant, peanut and mango leaf, or raisins, while a significant, trivial performance decrement was evident following banana consumption. Two Panax ginseng studies [101, 102] and one study investigating the effects of honey consumption [103] did not provide sufficient information for inclusion in the meta-analysis, and each of these studies failed to demonstrate significant effects on exercise performance.


Polyphenol supplementation forest plot. Abbreviations – CI, confidence interval; GXT, graded exercise test; IGXT, intermittent graded exercise test; ITTE, intermittent time to exhaustion; k, number of trials; n, sample size; PL, performance level; Qb, between-group Q-statistic; SMD, standardised mean difference (Hedge’s g); supp., supplementation; TT, time-trial; TTE, time to exhaustion; Vmax, maximal running velocity.
L-citrulline/Watermelon juice supplementation
L-citrulline (consumed via watermelon juice in all studies) had insignificant trivial detrimental effects on exercise performance based on the results of 4 trials (Figure 5). As shown in Additional Material 4, no significant effects were evident for any subgroup analysis (e.g. exercise mode, type, V̇O2max etc.) and all included studies featured low statistical heterogeneity (I2= 0%). An additional study by Tarazona-Diaz et al. [161] not included in the meta-analysis due to insufficient data reporting also failed to demonstrate any effect of watermelon juice on exercise performance.

L-citrulline supplementation forest plot. Abbreviations – n, sample size; SMD, standardised mean difference (Hedge’s g); CI, confidence interval; GXT, graded exercise test; IGXT, TT, time-trial; TTE, time to exhaustion; Qb, between-group Q-statistic.
Discussion
Key findings
Consumption of foods rich in NO3- and polyphenols exhibited trivial beneficial effects on endurance exercise performance, while no effects of consuming foods rich in L-citrulline were apparent. Grapes, French maritime pine bark and Montmorency cherry, were the most effective food sources of polyphenols, and males may benefit further from polyphenol consumption, with small performance improvements (including small and moderate effects in TT and IGXT performance, respectively). Females do not appear to benefit from consumption of NO3-or polyphenols, although there are sample size limitations. Fitness does not appear to influence the response to polyphenol consumption, while less-trained male athletes (V̇O2max: 45-55 ml.kg.min-1) may obtain the greatest benefits from NO3- consumption and no effects of NO3- are seen in more highly trained athletes (V̇O2max: >65 ml.kg.min-1).
Nitrate
Findings and proposed mechanisms
This meta-analysis provided evidence that consumption of dietary NO3-, particularly via consumption of beetroot, provided trivial but significant benefits for endurance exercise performance. This finding corresponds with other reviews that have investigated the ergogenic potential of NO3-, despite some differences in eligible performance measures (e.g. minimum performance duration, inclusion of non-locomotor performance) and the inclusion of non-food-derived sources such as sodium and potassium NO3- [165, 166]. Within this review, NO3- consumption displayed significant, small benefits for TTE performance and trivial benefits for TT, GXT performance, with effect sizes aligned with those of McMahon et al. [165]. This review also assessed the effects of NO3- on intermittent performance tests, which are more relevant for team-sport athletes given the intermittent nature of their competition, and trivial benefits were found for ITT and IGXT performance. Consistent with Campos et al. [165], we found that effects of NO3- on exercise performance are reduced in more highly conditioned athletes. Sub-group analyses indicated that while no effects were evident for athletes with higher V̇O2max values (>65 ml.kg.min-1), there were significant small and trivial benefits from NO3- consumption in males with a V̇O2max of 45-55 and 55-65 ml.kg.min-1, respectively.
Overall, these results suggest that beetroot juice may confer the greatest benefit in males of lower fitness, particularly during TTE tests. The improvements in exercise performance may have resulted from increases in nitric oxide synthesis and vascular function, as inferred via elevated levels of NO3-and nitrite, and reductions in systolic blood pressure (Additional Material 3), which enhance oxygen delivery and exercise efficiency [8]. These exercise effects were most apparent in less-trained individuals, who demonstrated more frequent reductions in submaximal oxygen consumption and augmentation of tissue oxygenation and oxygen uptake kinetics following NO3- supplementation (Additional Material 3). Improvements in exercise performance with NO3- intake are believed to be a result of improved mechanical efficiency and V̇O2 kinetics that derive from effects on fast-twitch muscle fibres [8], such as enhancement of excitation-contraction coupling. However, the reduced percentage of fast-twitch fibres and increased expression of calcium handling proteins [167] in elite endurance athletes, may limit these effects, in addition to their increased levels of endogenous nitric oxide synthesis [168, 169], vascular function [26, 29] and habitual dietary NO3- intake [170].
Food-specific effects of nitrate consumption
While beetroot juice was the predominant food source of NO3- in the included trials, which demonstrated trivial benefits for performance, a small number of studies evaluated the effect of NO3- from other sources, such as red spinach [97, 98], Swiss chard and rhubarb [99]. These studies reported no benefit for exercise performance; however this may be due to the lower NO3- content (~1.5 mmol vs average of 8.4 mmol within included studies) [97, 98] and the assessment of time-trial performance in trained (PL2) males, for which NO3- consumption overall demonstrated no benefit [99].
Notably, the included (NO3--rich) beetroot studies predominantly used NO3--depleted beetroot juice as the comparator, and these studies are therefore assessing the effects of their high NO3- content, rather than their overall nutritional properties. Beetroot juice, including commercial concentrated shots (e.g. James White Drinks Ltd, Ipswich, UK) typically used in studies, possesses high total phenolic content and antioxidant activity [171,172,173], which may confer additional advantages for exercise performance. Lansley et al. [72] found no significant effects of consuming NO3--depleted beetroot juice, suggesting that ergogenic effects of beetroot juice are attributable to its NO3- content, yet within this review, NO3--depleted beetroot supplementation exhibited trivial but significant benefits for performance [72, 109,110,111]. Thus, the full effects of beetroot may be somewhat underestimated in studies that have used NO3--depleted beetroot juice as a comparator.
Polyphenols
Findings and proposed mechanism
The current analysis identified that consumption of polyphenol-rich foods, both acutely and over several days, resulted in trivial but significant effects on endurance exercise performance. This is in agreement with a previous polyphenol meta-analysis [174], although their included interventions included purified polyphenol extracts from unspecified food sources in addition to whole-food extracts. Responses between and within foods varied significantly in the present analysis, as demonstrated by high heterogeneity, which may reflect the complex interplay between the food matrix, its phytochemical and nutritional composition, background diet and other genetic factors that have been shown to produce significant inter-individual variation in the responses to polyphenol consumption [19, 20]. This variation in the response between different foods is typified by both the non-significant influence of total phenolic content on effect size, as well the differential effects between food sources with similar predominant polyphenolic compounds (e.g. anthocyanins in blueberries, cherries and blackcurrant), as can be seen in Table 6 and the phenolic content of interventions listed in Additional Material 3. Further differences in interventions of included studies such as the consumption of single vs multiple polyphenol-rich food sources, the use of whole foods, juices, powders or extracts, as well as restrictions on specific foods or antioxidant sources, may contribute to such heterogeneous responses. Grape, (NO3--depleted) beetroot, French maritime pine, pomegranate and Montmorency cherry were the only polyphenol-rich foods that demonstrated significant ergogenic effects, while foods sources such as carob, Danggui Buxue Tang, Ecklonia cava, Rhodiola rosea, Spirulina platensis and yerba mate have shown promising effects within single studies and warrant further investigation.
The potential benefits of polyphenols for exercise performance are frequently attributed to their proposed ability to enhance vascular function and limit oxidative damage during exercise by upregulating endogenous antioxidant capacity. However, direct evidence of the physiological mechanisms underpinning the performance changes observed within the included studies was limited. As shown in Additional Material 3, only five polyphenol studies included a direct measure of nitric oxide status (e.g. plasma NO3- or NO2-), with only one study [149] demonstrating a significant increase in any of these biomarkers (plasma NO3-) following polyphenol (pomegranate) consumption. Similarly, only 14 polyphenol studies investigated effects on vascular parameters such as flow-mediated dilation (FMD) and blood pressure. Improvements in resting FMD were only observed following consumption of cocoa [119] and a cranberry and grape seed extract [122] in well-trained and elite athletes, respectively, while two studies found post-exercise (30-90 minutes) improvements in blood flow [151], and systolic blood pressure [138]. Food-derived polyphenol consumption has had inconsistent effects on vascular function in other healthy populations overall, although consumption of specific polyphenol sources such as tea, cocoa/chocolate and soy has demonstrated more consistent improvements in FMD [191, 192], and thus vascular effects may also be food-specific. However, vascular and skeletal muscle neuronal and endothelial NOS are upregulated by chronic exercise [3], fitness is significantly correlated with nitric oxide production [26, 168] and athletes may also have greater NO bioavailability via enhanced antioxidant defences, all of which may limit the vascular benefits seen in other populations. There was also little evidence for other mechanisms of action, such as improvements in markers of oxidative stress and muscle damage, or internal performance parameters (e.g. oxygen consumption, blood lactate accumulation). Thus, the mechanisms by which polyphenol-rich foods improve endurance exercise performance requires further consideration.
Influence of fitness
While the positive effects of NO3- consumption appeared to decrease with greater levels of aerobic fitness, such a relationship was not apparent in polyphenol trials, as small benefits were evident across a range of fitness levels (PL1, PL3 and PL4). When restricted to males only, small, significant benefits were shown in PL2 trials. Of considerable interest is that within males, small effects remained for TT performance within each PL3 & PL4 (only one TT trial in PL1). A 2017 meta-analysis of the performance effects of polyphenols, though not restricted to food sources, also reported that training status did not affect response to supplementation, with polyphenols having a significant ergogenic effect overall [174]. Unlike NO3- trials, no data were available for PL5 athletes within this review, although several studies have been conducted on high-level endurance athletes where V̇O2max data was unavailable, or a PL was not allocated due to test modality differences. No effects were seen following polyphenol supplementation in national-level rowers [105, 112], a cohort of elite athletes (primarily speed-skaters) [122], or national-level runners [147], although the latter two studies [122, 147] utilised a performance test mode that was not specific to their discipline. Given the lack of polyphenol studies evaluating the responses of elite endurance athletes, the efficacy of polyphenol consumption for this population remains unclear and warrants further investigation.
Influence of polyphenol consumption protocol (dosage & supplementation duration)
Several sub-group analyses were used to investigate the possible influences of dose and duration of consumption on the response to polyphenol-rich foods. Regression analysis indicated that total phenolic content was not a significant moderator of effects, suggesting a lack of a dose-response relationship between polyphenol intake and endurance exercise performance. Further, two included studies provided separate effect sizes for responses to differing levels of polyphenol content of the same food, and while these studies indicated no dose-response relationship, neither study induced a significant effect in response to any dosage [108, 148]. There were also no significant differences in the effects between acute and multiple-day consumption, suggesting no influence of duration of consumption. Three studies investigated the responses to polyphenol consumption across different supplementation periods but showed no effect for any supplementation length [113, 148, 156]. Also, while some multiple-day NO3- loading protocols included an additional acute dose approximately 2.5 hours before exercise performance testing, only half of the multiple-day polyphenol trials reported the use of an acute dose (see Additional Material 3). Pharmacokinetic analyses have indicated that plasma polyphenol concentrations typically peak around 2.2 hours post-ingestion [193], yet effects between multiple-day polyphenol trials that did or did not include an additional acute dose before performance testing were equivocal. Given the unclear role of food total phenolic content, variation in pre-exercise consumption timing and similar results between acute and multiple-day supplementation, these results do not provide clear insights into recommendations for consumption protocols that can optimise the ergogenic response to polyphenol-rich foods.
L-Citrulline/Watermelon Juice
Four watermelon juice studies were included within the review, which were conducted in participants of multiple fitness levels (two trials each for PL2 and PL3) and assessed performance through GXTs, ITTs, TTs and TTE tests. However, the included studies had significant differences in the l-citrulline content of the watermelon juice used. Bailey et al. [162] used concentrated watermelon juice containing 2.3-3.4 times the l-citrulline content in the pureed juice used in other included studies [161, 163, 164] and was the also only study to demonstrate an increase in pre-exercise plasma NO3- or any marker of nitric oxide synthesis (Additional Material 3). Despite increases in submaximal tissue oxygenation, Bailey et al. [162] indicated minimal effects on exercise performance, oxygen consumption or oxygen uptake kinetics, and no other studies noted improvements in internal performance indicators. Thus, no effect of watermelon juice consumption on exercise performance was observed overall, or within any individual study.
Areas for future research
Sex differences in effects of nitric oxide-related supplementation
While trivial effects were evident for the consumption of NO3- and polyphenol-rich foods, neither nutrient was effective in enhancing female endurance exercise performance (g=0.0 and 0.01, respectively) based on data from four trials. Polyphenol mixed-sex trials also demonstrated no effect on exercise performance, whereas trials of males only demonstrated small, significant effects. This finding is in agreement with that of a polyphenol meta-analysis by Somerville, Bringans and Braakhuis [174], who noted an attenuated effect (magnitude not specified) of exercise performance in studies that included both females and males.
Regarding the lack of efficacy of NO3- consumption for improving exercise performance in females, it is difficult to determine whether the lack of effect is attributable to biological differences between the sexes or the contexts of the studies. All four trials that assessed the effect of NO3- on exercise performance in females assessed time-trial performance, and were predominantly conducted in high-level athletes, with two studies conducted in participants classified as PL4 [62] and PL5 [70], and one study conducted in national-level water polo players. Hence, the absence of effects in these trials may be attributable to the decreased efficacy demonstrated for both time-trial performance and fitter athletes within this review, although an included study by de Castro, de Assis Manoel, and Machado [59] found no effects in untrained females. Overall, the small number of female trials, particularly in less well-trained athletes, combined with the similar effect sizes between mixed-sex (although these were predominantly comprised of males) trials and male-only trials, makes it difficult to infer whether there are genuine sex differences in the response to NO3- consumption.
If sex differences in the response to polyphenol and NO3- consumption do exist, several potential mechanisms may be responsible. Females have increased endothelium-dependent dilation [28], which may be attributable to elevated levels of plasma nitrite as a result of enhanced NO3--reducing activity by oral bacteria [194] and an augmenting role of estrogen in eNOS expression [27]. The increased proportion of slow-twitch muscle fibres in females [195] may be an additional factor, given that NO3- consumption proposedly confers its ergogenic potential primarily via effects on fast-twitch muscle fibres [8]. Females also appear to have reduced levels of oxidative stress compared with males, which may be due to a variety of factors including the antioxidant properties of estrogen [30]. While these factors may provide mechanistic underpinnings for the reduced response observed in females, studies in females are underrepresented in the literature, comprising only 18% of the participants within the meta-analysis and female-only studies were even more limited. Thus, further research of potential sex differences in the response to NO3- and polyphenol consumption is certainly still warranted.
Effects of exercise intensity on responses to supplementation
Metabolic acidosis is proposed to inhibit NO synthesis through the NOS-dependent pathway while enhancing synthesis through the NOS-independent pathway [196], which would suggest distinct intensity-dependent effects of NO3- and polyphenol consumption. However, there appeared to be no moderating influence of test duration on effect sizes for TT or TTE tests following both NO3- and polyphenol consumption. Several studies assessed the effects of NO3- and polyphenol consumption across multiple exercise intensities, with mixed results. Investigating the effects of NO3- on TTE at increasing exercise intensities, Kelly et al. [67] reported decreasing effect sizes (Hedge’s g = 1.06, 0.64, 0.62 and 0.48, respectively) as intensity increased and no significant effect at maximal intensity (V̇O2peak). Similarly, [151] Trexler et al. found significant TTE improvements at 90% and 100%, but not 110%, of peak aerobic velocity following pomegranate extract consumption. Two studies conducted multiple distance TTs following NO3- consumption, with Shannon et al. [85] noting positive effects in a 1.5 km but not 10 km running TT, whereas no differences between improvements were seen between 4 and 16.1 km cycling TT performance by Lansley et al. [71]. Regarding intermittent performance, Wylie et al. [95] found significant increases in mean power output during 24 repeated six-second sprints, but no effect on exercise performance across six 60-second efforts. Overall, these studies do not provide a clear indication of whether the effectiveness of consuming foods that promote synthesis of nitric oxide may be influenced by the intensity/duration of the performance test.
Potential influence of other dietary factors
The bioavailability of polyphenols is dependent on several factors including the phytochemical and overall composition of their food source matrix, background diet and genetic factors, particularly intestinal microbiota [25, 191]. Most included studies increased polyphenol intake through supplementation with a specific product or food, rather than a more holistic dietary intervention that increases polyphenol intake from various food sources. Notable exceptions included Knab et al. (2014), who utilised a fruit and vegetable juice blend powder, while a study by Ueberschlag et al. (2016) used Protandim, a mix of milk thistle, bacopa, ashwagandha root, turmeric and green tea, but no significant performance improvements were seen in either of these studies. There is evidence that interactions between specific combinations of polyphenols and their food sources can have both synergistic and inhibitory effects on antioxidant activity [18, 197], but whether this may influence exercise performance is not yet established. It has also been postulated that the co-ingestion of NO3- and polyphenols could have a synergistic effect on nitric oxide status [18, 191], which may contribute to increased efficacy of NO3- administered as beetroot juice in comparison to sodium NO3- [198], although limited evidence is available in support of this [17]. Baker et al. [199] found significant improvements in TT performance following a four-day Mediterranean diet intervention where consumption of olive oil, fruits, nuts, seeds and vegetables was significantly increased, although there were also changes in intake of fish and red meat. Thus, while food-derived polyphenol consumption was shown to have significant overall benefits within this review, further research is still warranted into whether polyphenol consumption through a more holistic, whole foods-based approach may still enhance endurance exercise performance and whether the co-consumption of polyphenols NO3--rich foods could confer any additional ergogenic effects.
Given the potential interactions of NO3- and polyphenols with other dietary factors, it is also of interest as to whether nitric oxide-related supplements may affect the responses to other ergogenic aids such as caffeine. Four beetroot studies investigated the effects of beetroot juice both alone and in combination but indicated that beetroot juice had neither a positive effect independently, nor did it influence the ergogenic effect of caffeine [62, 63, 70, 80]. These studies were all conducted on highly-trained athletes (all ≥PL4) and three of the four studies assessed performance via a TT, with both factors having reduced ergogenic effects within this review. No included polyphenol studies investigated any interactional effects with caffeine, although an excluded study investigating the effects of consuming coffee rich in caffeine and chlorogenic acid demonstrated no effects on TT performance despite comparison against a decaffeinated placebo [200].
Effects on training adaptations
While it appears that NO3- and polyphenol consumption can enhance endurance exercise performance, it is also important to evaluate whether their consumption may influence an athlete’s response to training. While most studies controlled each individual’s training performed during the interventions, several studies investigated responses to polyphenol consumption during a training program. NO3--rich beetroot juice consumption enhanced adaptions to sprint interval training [35, 89], which may be related to enhancing exercise capacity during training [201], as well as remodelling of skeletal muscle towards oxidative phenotypes [35], although this has not been consistently found [89]. While Kuo et al. [134] found no effects of green tea consumption on its own, an ergogenic response was shown when it was combined with an endurance training protocol. This interactional effect has not been replicated by other polyphenols studies however, with no discernible effects evident following consumption of ginger and New Zealand blackcurrant in combination with high-intensity interval training [128, 140], although both of these studies were in females. It has been proposed that there is a hormetic relationship between the oxidative stress induced by training and subsequent adaptations, whereby an optimal amount of oxidative damage is needed to maximise training adaptations, whereas inadequate or excess levels can both result in negative responses [202]. Indeed, chronic antioxidant supplementation with vitamin E has been linked to impaired performance [203], and varied responses have been demonstrated following vitamin C supplementation [204]. Presently, the lack of negative effects overall and in response to the same training suggests that consumption of polyphenol and NO3--rich foods does not impair adaptations to training, although their ability to augment these adaptations requires further investigation.
Limitations
Polyphenol inclusion criteria
One limitation of identifying polyphenol-rich foods within the present study was that there is no set definition of what this entails, and often studies did not specify phenolic content or reported only the composition of specific polyphenolic compounds and total phenolic content was unclear. This issue was identified during the development of the search strategy, and thus it was decided that a separate meta-analysis would also be conducted for studies that did report phenolic content. However, as seen in Table 6, results largely did not differ between studies that did and did not report phenolic content.
Several foods featured in the included studies also contained other bioactive compounds (e.g. other antioxidants such as polysaccharides, carotenoids, ginsenosides, vitamins C and E) that may confound the effects of their phenolic content. While there are some exceptions [184], in many such foods, antioxidant activity remains very strongly linked to total phenolic content [178, 205,206,207], suggesting that polyphenols are the primary driver of their antioxidant properties. Also, in the interest of evaluating the effectiveness of these foods overall and maintaining ecological validity, foods were deemed eligible unless such compounds were added to foods separately.
Risk of Bias
In contrast to older reviews that used the previous Cochrane RoB Tool, no studies within this review were classified as low risk using the RoB Tool 2.0, reflecting its more explicit, detailed requirements that were not aligned well with sports nutrition and exercise science reporting standards. Future publications should provide more specific details to better ascertain risk, particularly regarding specification of the randomisation process used, allocation sequence concealment, compliance with dietary interventions, and reference to pre-specified data analysis protocols, as these were poorly addressed by included studies.
Conclusion
Consumption of foods rich in NO3- and polyphenols may provide trivial beneficial effects for endurance exercise performance, while consumption of foods rich in L-citrulline, currently limited only to studies of watermelon juice, does not appear to affect performance. Beetroot juice has been extensively studied and its NO3- content confers ergogenic effects in various exercise types in populations that are not considered well-trained. Other food sources of NO3- require further investigation of their ergogenic capacity. Food-derived polyphenols appear to have the potential to enhance TT performance to a similar extent as beetroot juice, although more research is needed regarding its efficacy for use in highly trained athletes. No effects were evident for the consumption of polyphenols from New Zealand blackcurrant, cocoa, ginseng, green tea and raisins, but significant benefits were shown for the consumption of grape, beetroot (NO3--depleted), French maritime pine, Montmorency cherry and pomegranate across multiple studies. However, caution should be exercised in translating these ergogenic effects to other food sources of polyphenols, as there seems to be considerable variation in the effects between foods that cannot be attributed to differences in total phenolic content or key polyphenolic compounds. Distinct responses to NO3- and polyphenol supplementation were also observed between males and females, with females not demonstrating any benefit for exercise performance. This may be due to sex-based differences in nitric oxide synthesis, vascular function and oxidative stress, and/or the limited number of female studies and the training status of the participants. NO3--rich food consumption increases nitric oxide synthesis, and its physiological effects are more clearly linked to increases in muscle oxygen delivery and exercise efficiency, whereas polyphenol-rich foods have less clearly established effects on nitric oxide synthesis, vascular function and physiological variables during exercise. Future studies should evaluate effects of NO3- and polyphenol consumption on training performance and adaptations, as well as optimising protocols for consuming polyphenol-rich foods, and establishing the individual and test-related (e.g. intensity) factors that influence the ergogenic response to consuming NO3- and polyphenol-rich foods.
Availability of data and materials
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- GXT:
-
graded exercise test
- IGXT:
-
intermittent graded exercise test
- ITT:
-
intermittent time-trial
- ITTE:
-
intermittent time to exhaustion
- k:
-
number of trials
- km:
-
kilometre
- n:
-
sample size
- NO2 - :
-
nitrite
- NO3 - :
-
nitrate
- PL:
-
performance level
- Qb :
-
between-group Q-statistic
- SMD:
-
standardised mean difference (Hedge’s g)
- TT:
-
time-trial
- TTE:
-
time to exhaustion
References
Coggan AR, Peterson LR. Dietary Nitrate Enhances the Contractile Properties of Human Skeletal Muscle. Exerc Sport Sci Rev. 2018;46(4):254–61.
Bailey JC, Feelisch M, Horowitz JD, Frenneaux MP, Madhani M. Pharmacology and therapeutic role of inorganic nitrite and nitrate in vasodilatation. Pharmacol Ther. 2014;144(3):303–20.
Stamler JS, Meissner G. Physiology of Nitric Oxide in Skeletal Muscle. Physiol Rev. 2001;81(1):209–37.
Radak Z, Zhao Z, Koltai E, Ohno H, Atalay M. Oxygen consumption and usage during physical exercise: the balance between oxidative stress and ROS-dependent adaptive signaling. Antioxid Redox Signal. 2013;18(10):1208–46.
Wink DA, Miranda KM, Espey MG, Pluta RM, Hewett SJ, Colton C, et al. Mechanisms of the antioxidant effects of nitric oxide. Antioxid Redox Signal. 2001;3(2):203–13.
Menezes EF, Peixoto LG, Teixeira RR, Justino AB, Puga GM, Espindola FS. Potential Benefits of Nitrate Supplementation on Antioxidant Defense System and Blood Pressure Responses after Exercise Performance. Oxid Med Cell Longev. 2019;2019:7218936.
Clifford T, Howatson G, West DJ, Stevenson EJ. The potential benefits of red beetroot supplementation in health and disease. Nutrients. 2015;7(4):2801–22.
Jones AM, Thompson C, Wylie LJ, Vanhatalo A. Dietary Nitrate and Physical Performance. Annu Rev Nutr. 2018;38(1):303–28.
Jones AM, Grassi B, Christensen PM, Krustrup P, Bangsbo J, Poole DC. Slow component of VO2 kinetics: mechanistic bases and practical applications. Med Sci Sports Exerc. 2011;43(11):2046–62.
Stoclet JC, Chataigneau T, Ndiaye M, Oak MH, El Bedoui J, Chataigneau M, et al. Vascular protection by dietary polyphenols. Eur J Clin Pharmacol. 2004;500(1-3):299–313.
Reid MB. Redox interventions to increase exercise performance. J Physiol. 2016;594(18):5125–33.
Powers SK, Deminice R, Ozdemir M, Yoshihara T, Bomkamp MP, Hyatt H. Exercise-induced oxidative stress: Friend or foe? J Sport Health Sci. 2020;9(5):415–25.
Bowtell J, Kelly V. Fruit-Derived Polyphenol Supplementation for Athlete Recovery and Performance. Sports Med. 2019;49(Suppl 1):3–23.
Blekkenhorst LC, Prince RL, Ward NC, Croft KD, Lewis JR, Devine A, et al. Development of a reference database for assessing dietary nitrate in vegetables. Mol Nutr Food Res. 2017;61(8).
Shahidi F, Ambigaipalan P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects – A review. J Funct Foods. 2015;18:820–97.
Hord NG, Tang Y, Bryan NS. Food sources of nitrates and nitrites: the physiologic context for potential health benefits. Am J Clin Nutr. 2009;90(1):1–10.
Bondonno CP, Yang X, Croft KD, Considine MJ, Ward NC, Rich L, et al. Flavonoid-rich apples and nitrate-rich spinach augment nitric oxide status and improve endothelial function in healthy men and women: a randomized controlled trial. Free Radic Biol Med. 2012;52(1):95–102.
Phan MAT, Paterson J, Bucknall M, Arcot J. Interactions between phytochemicals from fruits and vegetables: Effects on bioactivities and bioavailability. Crit Rev Food Sci Nutr. 2018;58(8):1310–29.
Bohn T. Dietary factors affecting polyphenol bioavailability. Nutr Rev. 2014;72(7):429–52.
Scalbert A, Williamson G. Dietary Intake and Bioavailability of Polyphenols. J Nutr. 2000;130(8):2073S–85S.
McIlvenna LC, Monaghan C, Liddle L, Fernandez BO, Feelisch M, Muggeridge DJ, et al. Beetroot juice versus chard gel: A pharmacokinetic and pharmacodynamic comparison of nitrate bioavailability. Nitric Oxide. 2017;64:61–7.
James PE, Willis GR, Allen JD, Winyard PG, Jones AM. Nitrate pharmacokinetics: Taking note of the difference. Nitric Oxide. 2015;48:44–50.
Rothwell JA, Medina-Remón A, Pérez-Jiménez J, Neveu V, Knaze V, Slimani N, et al. Effects of food processing on polyphenol contents: A systematic analysis using Phenol-Explorer data. Mol Nutr Food Res. 2015;59(1):160–70.
Kay CD, Pereira-Caro G, Ludwig IA, Clifford MN, Crozier A. Anthocyanins and Flavanones Are More Bioavailable than Previously Perceived: A Review of Recent Evidence. Annu Rev Food Sci Technol. 2017;8(1):155–80.
Kawabata K, Yoshioka Y, Terao J. Role of Intestinal Microbiota in the Bioavailability and Physiological Functions of Dietary Polyphenols. Molecules. 2019;24(2):370.
Vassalle C, Lubrano V, Domenici C, L'Abbate A. Influence of chronic aerobic exercise on microcirculatory flow and nitric oxide in humans. Int J Sports Med. 2003;24(1):30–5.
Hayashi T, Yamada K, Esaki T, Kuzuya M, Satake S, Ishikawa T, et al. Estrogen Increases Endothelial Nitric Oxide by a Receptor Mediated System. Biochem Biophys Res Commun. 1995;214(3):847–55.
Stanhewicz AE, Wenner MM, Stachenfeld NS. Sex differences in endothelial function important to vascular health and overall cardiovascular disease risk across the lifespan. Am J Physiol Heart Circ Physiol. 2018;315(6):H1569–h88.
Franzoni F, Galetta F, Morizzo C, Lubrano V, Palombo C, Santoro G, et al. Effects of age and physical fitness on microcirculatory function. Clin Sci (Lond). 2004;106(3):329–35.
Kander MC, Cui Y, Liu Z. Gender difference in oxidative stress: a new look at the mechanisms for cardiovascular diseases. J Cell Mol Med. 2017;21(5):1024–32.
Mason SA, Trewin AJ, Parker L, Wadley GD. Antioxidant supplements and endurance exercise: Current evidence and mechanistic insights. Redox Biol. 2020;35:101471.
Pickering C, Kiely J. Are low doses of caffeine as ergogenic as higher doses? A critical review highlighting the need for comparison with current best practice in caffeine research. Nutrition. 2019;67-68:110535.
Sterne JAC, Savović J, Page MJ, Elbers RG, Blencowe NS, Boutron I, et al. RoB 2: a revised tool for assessing risk of bias in randomised trials. BMJ. 2019;366:l4898.
Lakens D. Calculating and reporting effect sizes to facilitate cumulative science: a practical primer for t-tests and ANOVAs. Front Psychol. 2013;4(863).
Thompson C, Wylie LJ, Blackwell JR, Fulford J, Black MI, Kelly J, et al. Influence of dietary nitrate supplementation on physiological and muscle metabolic adaptations to sprint interval training. J Appl Physiol (1985). 2017;122(3):642-652.
Higgins J, Eldridge S, Li T. Chapter 23: Including variants on randomized trials. 2021. In: Cochrane Handbook for Systematic Reviews of Interventions. Cochrane. Version 6.2. Available from: http://www.training.cochrane.org/handbook.
Higgins JPT, Li T, Deeks J. Chapter 6: Choosing effect measures and computing estimates of effect. 2020. In: Cochrane Handbook for Systematic Reviews of Interventions. Cochrane. Version 6.1.
Elbourne DR, Altman DG, Higgins JP, Curtin F, Worthington HV, Vail A. Meta-analyses involving cross-over trials: methodological issues. Int J Epidemiol. 2002;31(1):140–9.
Martin BJ, Tan RB, Gillen JB, Percival ME, Gibala MJ. No effect of short-term green tea extract supplementation on metabolism at rest or during exercise in the fed state. Int J Sport Nutr Exerc Metab. 2014;24(6):656–64.
Cohen J. Statistical power analysis for the behavioral sciences. 2nd ed. Hillsdale, N.J.: L. Erlbaum Associates; 1988.
Higgins JP, Thompson SG. Quantifying heterogeneity in a meta-analysis. Stat Med. 2002;21(11):1539–58.
Higgins JPT, Thompson SG, Deeks JJ, Altman DG. Measuring inconsistency in meta-analyses. BMJ. 2003;327(7414):557–60.
De Pauw K, Roelands B, Cheung SS, de Geus B, Rietjens G, Meeusen R. Guidelines to classify subject groups in sport-science research. Int J Sports Physiol Perform. 2013;8(2):111–22.
Decroix L, De Pauw K, Foster C, Meeusen R. Guidelines to Classify Female Subject Groups in Sport-Science Research. Int J Sports Physiol Perform. 2016;11(2):204–13.
Aucouturier J, Boissière J, Pawlak-Chaouch M, Cuvelier G, Gamelin FX. Effect of dietary nitrate supplementation on tolerance to supramaximal intensity intermittent exercise. Nitric Oxide. 2015;49:16–25.
Bailey SJ, Winyard P, Vanhatalo A, Blackwell JR, DiMenna FJ, Wilkerson DP, et al. Dietary nitrate supplementation reduces the O2 cost of low-intensity exercise and enhances tolerance to high-intensity exercise in humans. J Appl Physiol. 2009;107(4):1144–55.
Bailey SJ, Varnham RL, DiMenna FJ, Breese BC, Wylie LJ, Jones AM. Inorganic nitrate supplementation improves muscle oxygenation, O2 uptake kinetics, and exercise tolerance at high but not low pedal rates. J Appl Physiol. 2015;118(11):1396–405.
Balsalobre-Fernández C, Romero-Moraleda B, Cupeiro R, Peinado AB, Butragueño J, Benito PJ. The effects of beetroot juice supplementation on exercise economy, rating of perceived exertion and running mechanics in elite distance runners: A double-blinded, randomized study. PLoS One. 2018;13(7).
Bernardi BB, Schoenfeld BJ, Alves RC, Urbinati KS, McAnulty SR, Junior TPS. Acute Supplementation with Beetroot Juice Does Not Enhance Exercise Performance among Well-trained Athletes: A Randomized Crossover Study. J Exerc Physiol Online. 2018;21(3):1–12.
Boorsma RK, Whitfield J, Spriet LL. Beetroot juice supplementation does not improve performance of elite 1500-m runners. Med Sci Sports Exerc. 2014;46(12):2326–34.
Breese BC, McNarry MA, Marwood S, Blackwell JR, Bailey SJ, Jones AM. Beetroot juice supplementation speeds O2 uptake kinetics and improves exercise tolerance during severe-intensity exercise initiated from an elevated metabolic rate. Am J Physiol Regul Integr Comp Physiol. 2013;305(12):R1441–50.
Callahan MJ, Parr EB, Hawley JA, Burke LM. Single and combined effects of beetroot crystals and sodium bicarbonate on 4-km cycling time trial performance. Int J Sport Nutr Exerc Metab. 2017;27(3):271–8.
Cermak NM, Res P, Stinkens R, Lundberg JO, Gibala MJ, Van Loon LJC. No improvement in endurance performance after a single dose of beetroot juice. Int J Sport Nutr Exerc Metab. 2012;22(6):470–8.
Cermak NM, Gibala MJ, Van Loon LJC. Nitrate supplementation's improvement of 10-km time-trial performance in trained cyclists. Int J Sport Nutr Exerc Metab. 2012;22(1):64–71.
Christensen PM, Petersen NK, Friis SN, Weitzberg E, Nybo L. Effects of nitrate supplementation in trained and untrained muscle are modest with initial high plasma nitrite levels. Scand J Med Sci Sports. 2017;27(12):1616–26.
Christensen PM, Nyberg M, Bangsbo J. Influence of nitrate supplementation on VO2 kinetics and endurance of elite cyclists. Scand J Med Sci Sports. 2013;23(1):e21–31.
de Castro TF, de Assis MF, Figueiredo DH, Figueiredo DH, Machado FA. Effects of chronic beetroot juice supplementation on maximum oxygen uptake, velocity associated with maximum oxygen uptake, and peak velocity in recreational runners: a double-blinded, randomized and crossover study. Eur J Appl Physiol. 2019;119(5):1043–53.
de Castro TF, Manoel FA, Figueiredo DH, Figueiredo DH, Machado FA. Effect of beetroot juice supplementation on 10-km performance in recreational runners. Appl Physiol Nutr Metab. 2019;44(1):90–4.
de Castro TF, de Assis MF, Machado FA. Beetroot juice supplementation does not modify the 3-km running performance in untrained women. Sci Sports. 2018;33(4):e167–e70.
Esen O, Nicholas C, Morris M, Bailey SJ. No Effect of Beetroot Juice Supplementation on 100-m and 200-m Swimming Performance in Moderately Trained Swimmers. Int J Sports Physiol Perform. 2019;14(6):706–10.
Flueck JL, Gallo A, Moelijker N, Bogdanov N, Bogdanova A, Perret C. Influence of Equimolar Doses of Beetroot Juice and Sodium Nitrate on Time Trial Performance in Handcycling. Nutrients. 2019;11(7).
Glaister M, Pattison JR, Muniz-Pumares D, Patterson SD, Foley P. Effects of dietary nitrate, caffeine, and their combination on 20-km cycling time trial performance. J Strength Cond Res. 2015;29(1):165–74.
Handzlik MK, Gleeson M. Likely additive ergogenic effects of combined preexercise dietary nitrate and caffeine ingestion in trained cyclists. ISRN Nutr. 2013;2013:396581.
Hoon MW, Hopkins WG, Jones AM, Martin DT, Halson SL, West NP, et al. Nitrate supplementation and high-intensity performance in competitive cyclists. Appl Physiol Nutr Metab. 2014;39(9):1043–9.
Hoon MW, Jones AM, Johnson NA, Blackwell JR, Broad EM, Lundy B, et al. The effect of variable doses of inorganic nitrate-rich beetroot juice on simulated 2000-m rowing performance in trained athletes. Int J Sports Physiol Perform. 2014;9(4):615–20.
Jonvik KL, Van Dijk JW, Senden JMG, Van Loon LJC, Verdijk LB. The effect of beetroot juice supplementation on dynamic apnea and intermittent sprint performance in elite female water polo players. Int J Sport Nutr Exerc Metab. 2018;28(5):468–73.
Kelly J, Vanhatalo A, Wilkerson DP, Wylie LJ, Jones AM. Effects of nitrate on the power-duration relationship for severe-intensity exercise. Med Sci Sports Exerc. 2013;45(9):1798–806.
Kelly J, Vanhatalo A, Bailey SJ, Wylie LJ, Tucker C, List S, et al. Dietary nitrate supplementation: effects on plasma nitrite and pulmonary O2 uptake dynamics during exercise in hypoxia and normoxia. Am J Physiol Regul Integr Comp Physiol. 2014;307(7):R920–30.
Kent GL, Dawson B, Cox GR, Burke LM, Eastwood A, Croft KD, et al. Dietary nitrate supplementation does not improve cycling time-trial performance in the heat. J Sports Sci. 2018;36(11):1204–11.
Lane SC, Hawley JA, Desbrow B, Jones AM, Blackwell JR, Ross ML, et al. Single and combined effects of beetroot juice and caffeine supplementation on cycling time trial performance. Appl Physiol Nutr Metab. 2014;39(9):1050–7.
Lansley KE, Winyard PG, Bailey SJ, Vanhatalo A, Wilkerson DP, Blackwell JR, et al. Acute dietary nitrate supplementation improves cycling time trial performance. Med Sci Sports Exerc. 2011;43(6):1125–31.
Lansley KE, Winyard PG, Fulford J, Vanhatalo A, Bailey SJ, Blackwell JR, et al. Dietary nitrate supplementation reduces the O2 cost of walking and running: A placebo-controlled study. J Appl Physiol. 2011;110(3):591–600.
Lowings S, Shannon OM, Deighton K, Matu J, Barlow MJ. Effect of Dietary Nitrate Supplementation on Swimming Performance in Trained Swimmers. Int J Sport Nutr Exerc Metab. 2017;27(4):377–84.
MacLeod KE, Nugent SF, Barr SI, Koehle MS, Sporer BC, MacInnis MJ. Acute Beetroot Juice Supplementation Does Not Improve Cycling Performance in Normoxia or Moderate Hypoxia. Int J Sport Nutr Exerc Metab. 2015;25(4):359–66.
McQuillan JA, Dulson DK, Laursen PB, Kilding AE. Dietary nitrate fails to improve 1 and 4 km cycling performance in highly trained cyclists. Int J Sport Nutr Exerc Metab. 2017;27(3):255–63.
McQuillan JA, Dulson DK, Laursen PB, Kilding AE. The Effect of Dietary Nitrate Supplementation on Physiology and Performance in Trained Cyclists. Int J Sports Physiol Perform. 2017;12(5):684–9.
Mosher SL, Gough LA, Deb S, Saunders B, Mc Naughton LR, Brown DR, et al. High dose Nitrate ingestion does not improve 40 km cycling time trial performance in trained cyclists. Res. Sports Med. 2019.
Muggeridge DJ, Howe CCF, Spendiff O, Pedlar C, James PE, Easton C. The effects of a single dose of concentrated beetroot juice on performance in trained flatwater kayakers. Int J Sport Nutr Exerc Metab. 2013;23(5):498–506.
Murphy M, Eliot K, Heuertz RM, Weiss E. Whole Beetroot Consumption Acutely Improves Running Performance. J Acad Nutr Diet. 2012;112(4):548–52.
Oskarsson J, McGawley K. No individual or combined effects of caffeine and beetroot-juice supplementation during submaximal or maximal running. Appl Physiol Nutr Metab. 2018;43(7):697–703.
Pawlak-Chaouch M, Boissiere J, Munyaneza D, Gamelin F-X, Cuvelier G, Berthoin S, et al. Beetroot Juice Does Not Enhance Supramaximal Intermittent Exercise Performance in Elite Endurance Athletes. J Am Coll Nutr. 2019;1-10.
Peeling P, Cox GR, Bullock N, Burke LM. Beetroot juice improves on-water 500 M time-trial performance, and laboratory-based paddling economy in national and international-level kayak athletes. Int J Sport Nutr Exerc Metab. 2015;25(3):278–84.
Pinna M, Roberto S, Milia R, Marongiu E, Olla S, Loi A, et al. Effect of beetroot juice supplementation on aerobic response during swimming. Nutrients. 2014;6(2):605–15.
Rokkedal-Lausch T, Franch J, Poulsen MK, Thomsen LP, Weitzberg E, Kamavuako EN, et al. Chronic high-dose beetroot juice supplementation improves time trial performance of well-trained cyclists in normoxia and hypoxia. Nitric Oxide. 2019;85:44–52.
Shannon OM, Barlow MJ, Duckworth L, Williams E, Wort G, Woods D, et al. Dietary nitrate supplementation enhances short but not longer duration running time-trial performance. Eur J Appl Physiol. 2017;117(4):775–85.
Tan R, Wylie LJ, Thompson C, Blackwell JR, Bailey SJ, Vanhatalo A, et al. Beetroot juice ingestion during prolonged moderate-intensity exercise attenuates progressive rise in O-2 uptake. J Appl Physiol. 2018;124(5):1254–63.
Thompson KG, Turner L, Prichard J, Dodd F, Kennedy DO, Haskell C, et al. Influence of dietary nitrate supplementation on physiological and cognitive responses to incremental cycle exercise. Respir Physiol Neurobiol. 2014;193(1):11–20.
Thompson C, Vanhatalo A, Jell H, Fulford J, Carter J, Nyman L, et al. Dietary nitrate supplementation improves sprint and high-intensity intermittent running performance. Nitric Oxide. 2016;61:55–61.
Thompson C, Vanhatalo A, Kadach S, Wylie LJ, Fulford J, Ferguson SK, et al. Discrete physiological effects of beetroot juice and potassium nitrate supplementation following 4-wk sprint interval training. J Appl Physiol (1985). 2018;124(6):1519-1528.
Vanhatalo A, Bailey SJ, Blackwell JR, DiMenna FJ, Pavey TG, Wilkerson DP, et al. Acute and chronic effects of dietary nitrate supplementation on blood pressure and the physiological responses to moderate-intensity and incremental exercise. Am J Physiol Regul Integr Comp Physiol. 2010;299(4):R1121–R31.
Vasconcellos J, Silvestre DH, Baiao DD, Werneck-de-Castro JP, Alvares TS, Paschoalin VMF. A Single Dose of Beetroot Gel Rich in Nitrate Does Not Improve Performance but Lowers Blood Glucose in Physically Active Individuals. Med J Nutrition Metab. 2017.
Wilkerson DP, Hayward GM, Bailey SJ, Vanhatalo A, Blackwell JR, Jones AM. Influence of acute dietary nitrate supplementation on 50 mile time trial performance in well-trained cyclists. Eur J Appl Physiol. 2012;112(12):4127–34.
Wylie LJ, Kelly J, Bailey SJ, Blackwell JR, Skiba PF, Winyard PG, et al. Beetroot juice and exercise: Pharmacodynamic and dose-response relationships. J Appl Physiol. 2013;115(3):325–36.
Wylie LJ, Mohr M, Krustrup P, Jackman SR, Ermdis G, Kelly J, et al. Dietary nitrate supplementation improves team sport-specific intense intermittent exercise performance. Eur J Appl Physiol. 2013;113(7):1673–84.
Wylie LJ, Bailey SJ, Kelly J, Blackwell JR, Vanhatalo A, Jones AM. Influence of beetroot juice supplementation on intermittent exercise performance. Eur J Appl Physiol. 2016;116(2):415–25.
Wylie LJ, Park JW, Vanhatalo A, Kadach S, Black MI, Stoyanov Z, et al. Human skeletal muscle nitrate store: influence of dietary nitrate supplementation and exercise. J Physiol. 2019.
Gonzalez AM, Accetta MR, Spitz RW, Mangine GT, Ghigiarelli JJ, Sell KM. Red Spinach Extract Supplementation Improves Cycle Time Trial Performance in Recreationally Active Men and Women. J Strength Cond Res. 2019.
Moore AN, Haun CT, Kephart WC, Holland AM, Mobley CB, Pascoe DD, et al. Red Spinach Extract Increases Ventilatory Threshold during Graded Exercise Testing. Sports. 2017;5(4).
Muggeridge DJ, Sculthorpe N, Grace FM, Willis G, Thornhill L, Weller RB, et al. Acute whole body UVA irradiation combined with nitrate ingestion enhances time trial performance in trained cyclists. Nitric Oxide. 2015;48:3–9.
Boussetta N, Abedelmalek S, Khouloud A, Ben anes A, Souissi N. Does red orange juice supplementation has a protective effect on performance, cardiovascular parameters, muscle damage and oxidative stress markers following the Yo-Yo Intermittent Recovery Test Level-1 under polluted air? Int J Environ Health Res. 2019.
Allen JD, McLung J, Nelson AG, Welsch M. Ginseng Supplementation Does Not Enhance Healthy Young Adults’ Peak Aerobic Exercise Performance. J Am Coll Nutr. 1998;17(5):462–6.
Engels H-J, Said JM, Wirth JC. Failure of chronic ginseng supplementation to affect work performance and energy metabolism in healthy adult females. Nutr Res. 1996;16(8):1295–305.
Abbey EL, Rankin JW. Effect of ingesting a honey-sweetened beverage on soccer performance and exercise-induced cytokine response. Int J Sport Nutr Exerc Metab. 2009;19(6):659–72.
Yi M, Fu J, Zhou L, Gao H, Fan C, Shao J, et al. The effect of almond consumption on elements of endurance exercise performance in trained athletes. J Int Soc Sports Nutr. 2014;11(1).
Basta P, Pilaczynska-Szczesniak L, Woitas-Slubowska D, Skarpanska-Stejnborn A. Influence of aloe arborescens Mill. Extract on selected parameters of pro-oxidant-antioxidant equilibrium and cytokine synthesis in rowers. Int J Sport Nutr Exerc Metab. 2013;23(4):388–98.
Hsu CC, Ho MC, Lin LC, Su B, Hsu MC. American ginseng supplementation attenuates creatine kinase level induced by submaximal exercise in human beings. World J Gastroenterol. 2005;11(34):5327–31.
Morris AC, Jacobs I, McLellan TM, Klugerman A, Wang LC, Zamecnik J. No ergogenic effect of ginseng ingestion. Int J Sport Nutr. 1996;6(3):263–71.
Nieman DC, Gillitt ND, Sha W, Esposito D, Ramamoorthy S. Metabolic recovery from heavy exertion following banana compared to sugar beverage or water only ingestion: A randomized, crossover trial. PLoS One. 2018;13(3):e0194843.
Montenegro CF, Kwong DA, Minow ZA, Davis BA, Lozada CF, Casazza GA. Betalain-rich concentrate supplementation improves exercise performance and recovery in competitive triathletes. Appl Physiol Nutr Metab. 2017;42(2):166–72.
Mumford PW, Kephart WC, Romero MA, Haun CT, Mobley CB, Osburn SC, et al. Effect of 1-week betalain-rich beetroot concentrate supplementation on cycling performance and select physiological parameters. Eur J Appl Physiol. 2018;118(11):2465–76.
Van Hoorebeke JS, Trias CO, Davis BA, Lozada CF, Casazza GA. Betalain-Rich Concentrate Supplementation Improves Exercise Performance in Competitive Runners. Sports. 2016;4(3).
Skarpańska-Stejnborn A, Basta P, Pilaczyńska-Szcześniak Ł. The Influence Of Supplementation With The Black Currant (Ribes Nigrum) Extract On Selected Prooxidative-Antioxidative Balance Parameters In Rowers. Stud Phys Cult Tourism. 2006;13(2):51–8.
Brandenburg JP, Giles LV. Four Days of Blueberry Powder Supplementation Lowers the Blood Lactate Response to Running But Has No Effect on Time-Trial Performance. Int J Sport Nutr Exerc Metab. 2019;1-7.
Gaamouri N, Zouhal H, Hammami M, Hackney AC, Abderrahman AB, Saeidi A, et al. Effects of polyphenol (carob) supplementation on body composition and aerobic capacity in taekwondo athletes. Physiol Behav. 2019;205:22–8.
Pospieszna B, Wochna K, JerszyŃSki D, GowaCinna K, Czapski J. Ergogenic effects of dietary nitrates in female swimmers. Trends Sport Sci. 2016;23(1):13–20.
Overdevest E, Wouters JA, Wolfs KHM, Van Leeuwen JJM, Possemiers S. Citrus flavonoid supplementation improves exercise performance in trained athletes. J Sports Sci Med. 2018;17(1):24–30.
Allgrove J, Farrell E, Gleeson M, Williamson G, Cooper K. Regular dark chocolate consumption's reduction of oxidative stress and increase of free-fatty-acid mobilization in response to prolonged cycling. Int J Sport Nutr Exerc Metab. 2011;21(2):113–23.
Decroix L, Tonoli C, Soares DD, Descat A, Drittij-Reijnders M-J, Weseler AR, et al. Acute cocoa Flavanols intake has minimal effects on exercise-induced oxidative stress and nitric oxide production in healthy cyclists: a randomized controlled trial. J Int Soc Sports Nutr. 2017;14:28.
Decroix L, Tonoli C, Lespagnol E, Balestra C, Descat A, Drittij-Reijnders MJ, et al. One-week cocoa flavanol intake increases prefrontal cortex oxygenation at rest and during moderate-intensity exercise in normoxia and hypoxia. J Appl Physiol. 2018;125(1):8–18.
Patel RK, Brouner J, Spendiff O. Dark chocolate supplementation reduces the oxygen cost of moderate intensity cycling. J Int Soc Sports Nutr. 2015;12:47.
Ostojic S, Stojanovic M, Djordjevic B, Jourkesh M, Vasiljevic N. The Effects of a 4-week Coffeeberry Supplementation on Antioxidant Status, Endurance, and Anaerobic Performance in College Athletes. Res Sports Med. 2008;16(4):281–94.
Labonté K, Couillard C, Motard-Bélanger A, Paradis M-E, Couture P, Lamarche B. Acute Effects of Polyphenols from Cranberries and Grape Seeds on Endothelial Function and Performance in Elite Athletes. Sports. 2013;1(3):55–68.
Chang CW, Chen CY, Yen CC, Wu YT, Hsu MC. Repressed exercise-induced hepcidin levels after Danggui Buxue Tang supplementation in male recreational runners. Nutrients. 2018;10(9).
Oh JK, Shin YO, Yoon JH, Kim SH, Shin HC, Hwang HJ. Effect of Supplementation With Ecklonia cava Polyphenol on Endurance Performance of College Students. Int J Sport Nutr Exerc Metab. 2010;20(1):72–9.
Bentley D, Dank S, Coupland R, Midgley A, Spence I. Acute Antioxidant Supplementation Improves Endurance Performance in Trained Athletes. Res Sports Med. 2012;20(1):1–12.
Clifford T, Mitchell N, Scott A. The influence of different sources of polyphenols on submaximal cycling and time trial performance. J Athl Enhanc. 2013;2:S10.
Mach J, Midgley AW, Dank S, Grant RS, Bentley DJ. The effect of antioxidant supplementation on fatigue during exercise: potential role for NAD+(H). Nutrients. 2010;2(3):319–29.
Nayebifar S, Afzalpour ME, Kazemi T, Eivary SHA, Mogharnasi M. The effect of a 10-week high-intensity interval training and ginger consumption on inflammatory indices contributing to atherosclerosis in overweight women. J Res Med Sci. 2016;21(8):116.
O'Connor PJ, Caravalho AL, Freese EC, Cureton KJ. Grape consumption's effects on fitness, muscle injury, mood, and perceived health. Int J Sport Nutr Exerc Metab. 2013;23(1):57–64.
Toscano LT, Tavares RL, Toscano LT, Silva CSO, Almeida AEM, Biasoto ACT, et al. Potential ergogenic activity of grape juice in runners. Appl Physiol Nutr Metab. 2015;40(9):899–906.
Deley G, Guillemet D, Allaert F-A, Babault N. An Acute Dose of Specific Grape and Apple Polyphenols Improves Endurance Performance: A Randomized, Crossover, Double-Blind versus Placebo Controlled Study. Nutrients. 2017;9(8).
Dean S, Braakhuis A, Paton C. The effects of EGCG on fat oxidation and endurance performance in male cyclists. Int J Sport Nutr Exerc Metab. 2009;19(6):624–44.
Eichenberger P, Mettler S, Arnold M, Colombani PC. No effects of three-week consumption of a green tea extract on time trial performance in endurance-trained men. Int J Vitam Nutr Res. 2010;80(1):54–64.
Kuo Y-C, Lin J-C, Bernard JR, Liao Y-H. Green tea extract supplementation does not hamper endurance-training adaptation but improves antioxidant capacity in sedentary men. Appl Physiol Nutr Metab. 2015;40(10):990–6.
Knab AM, Nieman DC, Gillitt ND, Shanely RA, Cialdella-Kam L, Henson D, et al. Effects of a freeze-dried juice blend powder on exercise-induced inflammation, oxidative stress, and immune function in cyclists. Appl Physiol Nutr Metab. 2014;39(3):381–5.
Bell PG, Walshe IH, Davison GW, Stevenson E, Howatson G. Montmorency cherries reduce the oxidative stress and inflammatory responses to repeated days high-intensity stochastic cycling. Nutrients. 2014;6(2):829–43.
Bell PG, Walshe IH, Davison GW, Stevenson EJ, Howatson G. Recovery facilitation with Montmorency cherries following high-intensity, metabolically challenging exercise. Appl Physiol Nutr Metab. 2015;40(4):414–23.
Keane KM, Bailey SJ, Vanhatalo A, Jones AM, Howatson G. Effects of montmorency tart cherry (L. Prunus Cerasus) consumption on nitric oxide biomarkers and exercise performance. Scand J Med Sci Sports. 2018;28(7):1746–56.
Morgan PT, Barton MJ, Bowtell JL. Montmorency cherry supplementation improves 15-km cycling time-trial performance. Eur J Appl Physiol. 2019;119(3):675–84.
Braakhuis AJ, Hopkins WG, Lowe TE. Effects of dietary antioxidants on training and performance in female runners. Eur J Sport Sci. 2014;14(2):160–8.
Cook MD, Myers SD, Blacker SD, Willems MET. New Zealand blackcurrant extract improves cycling performance and fat oxidation in cyclists. Eur J Appl Physiol. 2015;115(11):2357–65.
Murphy CA, Cook MD, Willems MET. Effect of New Zealand Blackcurrant Extract on Repeated Cycling Time Trial Performance. Sports. 2017;5(2).
Perkins I, Vine S, Blacker S, Willems M. New Zealand blackcurrant extract improves high-intensity intermittent running performance. Int J Sport Nutr Exerc Metab. 2015;25(Supplement 1):7.
Potter JA, Hodgson CI, Broadhurst M, Howell L, Gilbert J, Willems MET, et al. Effects of New Zealand blackcurrant extract on sport climbing performance. Eur J Appl Physiol. 2019.
Willems MET, Myers SD, Gault ML, Cook MD. Beneficial physiological effects with blackcurrant intake in endurance athletes. Int J Sport Nutr Exerc Metab. 2015;25(4):367–74.
Willems M, Cousins L, Williams D, Blacker S. Beneficial effect of New Zealand blackcurrant on maximal sprint speeds during the Loughborough intermittent shuttle test. Int J Sport Nutr Exerc Metab. 2016;26(Supplement 1):S7–8.
Esquius L, Garcia-Retortillo S, Balagué N, Hristovski R, Javierre C. Physiological- and performance-related effects of acute olive oil supplementation at moderate exercise intensity. J Int Soc Sports Nutr. 2019;16(1).
Gelabert-Rebato M, Wiebe JC, Martin-Rincon M, Galvan-Alvarez V, Curtelin D, Perez-Valera M, et al. Enhancement of exercise performance by 48 hours, and 15-day supplementation with mangiferin and luteolin in men. Nutrients. 2019;11(2).
Crum EM, Barnes MJ, Stannard SR. Multiday Pomegranate Extract Supplementation Decreases Oxygen Uptake During Submaximal Cycling Exercise, but Cosupplementation With N-acetylcysteine Negates the Effect. Int J Sport Nutr Exerc Metab. 2018;28(6):586–92.
Torregrosa-García A, Ávila-Gandía V, Luque-Rubia AJ, Abellán-Ruiz MS, Querol-Calderón M, López-Román FJ. Pomegranate extract improves maximal performance of trained cyclists after an exhausting endurance trial: A randomised controlled trial. Nutrients. 2019;11(4).
Trexler ET, Smith-Ryan AE, Melvin MN, Roelofs EJ, Wingfield HL. Effects of pomegranate extract on blood flow and running time to exhaustion. Appl Physiol Nutr Metab. 2014;39(9):1038–42.
Ueberschlag SL, Seay JR, Roberts AH, DeSpirito PC, Stith JM, Folz RJ, et al. The Effect of Protandim Supplementation on Athletic Performance and Oxidative Blood Markers in Runners. PLoS One. 2016;11(8):e0160559.
Kern M, Heslin CJ, Rezende RS. Metabolic and performance effects of raisins versus sports gel as pre-exercise feedings in cyclists. J Strength Cond Res. 2007;21(4):1204–7.
Rietschier HL, Henagan TM, Earnest CP, Baker BL, Cortez CC, Stewart LK. Sun-dried raisins are a cost-effective alternative to sports jelly beans in prolonged cycling. J Strength Cond Res. 2011;25(11):3150–6.
Jowko E, Sadowski J, Dlugolecka B, Gierczuk D, Opaszowski B, Cieslinski I. Effects of Rhodiola rosea supplementation on mental performance, physical capacity, and oxidative stress biomarkers in healthy men. J Sport Health Sci. 2018;7(4):473–80.
Dowling EA, Redondo DR, Branch JD, Jones S, McNabb G, Williams MH. Effect of Eleutherococcus senticosus on submaximal and maximal exercise performance. Med Sci Sports Exerc. 1996;28(4):482–9.
Eschbach LC, Webster MJ, Boyd JC, McArthur PD, Evetovich TK. The Effect of Siberian Ginseng (Eleutherococcus Senticosus) on Substrate Utilization and Performance during Prolonged Cycling. Int J Sport Nutr Exerc Metab. 2000;10(4):444.
Kalafati M, Jamurtas AZ, Nikolaidis MG, Paschalis V, Theodorou AA, Sakellariou GK, et al. Ergogenic and antioxidant effects of spirulina supplementation in humans. Med Sci Sports Exerc. 2010;42(1):142–51.
Wasuntarawat C, Pengnet S, Walaikavinan N, Kamkaew N, Bualoang T, Toskulkao C, et al. No effect of acute ingestion of Thai ginseng (Kaempferia parviflora) on sprint and endurance exercise performance in humans. J Sports Sci. 2010;28(11):1243–50.
Areta JL, Austarheim I, Wangensteen H, Capelli C. Metabolic and Performance Effects of Yerba Mate on Well-trained Cyclists. Med Sci Sports Exerc. 2018;50(4):817–26.
Tarazona-Díaz MP, Alacid F, Carrasco M, Martínez I, Aguayo E. Watermelon juice: potential functional drink for sore muscle relief in athletes. J Agric Food Chem. 2013;61(31):7522–8.
Bailey SJ, Blackwell JR, Williams E, Vanhatalo A, Wylie LJ, Winyard PG, et al. Two weeks of watermelon juice supplementation improves nitric oxide bioavailability but not endurance exercise performance in humans. Nitric Oxide. 2016;59:10–20.
Cutrufello PT, Gadomski SJ, Zavorsky GS. The effect of l-citrulline and watermelon juice supplementation on anaerobic and aerobic exercise performance. J Sports Sci. 2015;33(14):1459–66.
Shanely RA, Nieman DC, Perkins-Veazie P, Henson DA, Meaney MP, Knab AM, et al. Comparison of watermelon and carbohydrate beverage on exercise-induced alterations in systemic inflammation, immune dysfunction, and plasma antioxidant capacity. Nutrients. 2016;8(8).
McMahon NF, Leveritt MD, Pavey TG. The Effect of Dietary Nitrate Supplementation on Endurance Exercise Performance in Healthy Adults: A Systematic Review and Meta-Analysis. Sports Med. 2017;47(4):735–56.
Senefeld JW, Wiggins CC, Regimbal RJ, Dominelli PB, Baker SE, Joyner MJ. Ergogenic Effect of Nitrate Supplementation: A Systematic Review and Meta-analysis. Med Sci Sports Exerc. 2020;52(10):2250–61.
Tesch PA, Karlsson J. Muscle fiber types and size in trained and untrained muscles of elite athletes. J Appl Physiol (1985). 1985;59(6):1716-1720.
Totzeck M, Hendgen-Cotta UB, Rammos C, Frommke LM, Knackstedt C, Predel HG, et al. Higher endogenous nitrite levels are associated with superior exercise capacity in highly trained athletes. Nitric Oxide. 2012;27(2):75–81.
Maroun MJ, Mehta S, Turcotte R, Cosio MG, Hussain SN. Effects of physical conditioning on endogenous nitric oxide output during exercise. J Appl Physiol (1985). 1995;79(4):1219-1225.
Van der Avoort CMT, Van Loon LJC, Hopman MTE, Verdijk LB. Increasing vegetable intake to obtain the health promoting and ergogenic effects of dietary nitrate. Eur J Clin Nutr. 2018;72(11):1485–9.
Granato D, Karnopp AR, van Ruth SM. Characterization and comparison of phenolic composition, antioxidant capacity and instrumental taste profile of juices from different botanical origins. J Sci Food Agric. 2015;95(10):1997–2006.
Wootton-Beard PC, Ryan L. A beetroot juice shot is a significant and convenient source of bioaccessible antioxidants. J Funct Foods. 2011;3(4):329–34.
Wootton-Beard PC, Ryan L. Combined use of Multiple Methodologies for the Measurement of Total Antioxidant Capacity in UK Commercially Available Vegetable Juices. Plant Foods Hum Nutr. 2012;67(2):142–7.
Somerville V, Bringans C, Braakhuis A. Polyphenols and Performance: A Systematic Review and Meta-Analysis. Sports Med. 2017;47(8):1589–99.
Azuma T, Tanaka Y, Kikuzaki H. Phenolic glycosides from Kaempferia parviflora. Phytochemistry. 2008;69(15):2743–8.
da Silveira TFF, Meinhart AD, de Souza TCL, Teixeira Filho J, Godoy HT. Phenolic compounds from yerba mate based beverages – A multivariate optimisation. Food Chem. 2016;190:1159–67.
Załuski D, Olech M, Galanty A, Verpoorte R, Kuźniewski R, Nowak R, et al. Phytochemical Content and Pharma-Nutrition Study on Eleutherococcus senticosus Fruits Intractum. Oxid Med Cell Longev. 2016;2016:9270691-.
Chung IM, Lim JJ, Ahn MS, Jeong HN, An TJ, Kim SH. Comparative phenolic compound profiles and antioxidative activity of the fruit, leaves, and roots of Korean ginseng (Panax ginseng Meyer) according to cultivation years. J Ginseng Res. 2016;40(1):68–75.
Kochan E, Szymańska G, Wielanek M, Wiktorowska-Owczarek A, Jóźwiak-Bębenista M, Grzegorczyk-Karolak I. The content of triterpene saponins and phenolic compounds in American ginseng hairy root extracts and their antioxidant and cytotoxic properties. Plant Cell Tissue Organ Cult. 2019;138(2):353–62.
Sidhu JS, Zafar TA. Bioactive compounds in banana fruits and their health benefits. Food Qual Saf. 2018;2(4):183–8.
Lucini L, Pellizzoni M, Pellegrino R, Molinari GP, Colla G. Phytochemical constituents and in vitro radical scavenging activity of different Aloe species. Food Chem. 2015;170:501–7.
Kwan KKL, Huang Y, Leung KW, Dong TTX, Tsim KWK. Danggui Buxue Tang, a Chinese Herbal Decoction Containing Astragali Radix and Angelicae Sinensis Radix, Modulates Mitochondrial Bioenergetics in Cultured Cardiomyoblasts. Front Pharmacol. 2019;10(614).
Cianciosi D, Forbes-Hernández TY, Afrin S, Gasparrini M, Reboredo-Rodriguez P, Manna PP, et al. Phenolic Compounds in Honey and Their Associated Health Benefits: A Review. Molecules. 2018;23(9):2322.
Al-Dhabi NA, Valan AM. Quantification of Phytochemicals from Commercial Spirulina Products and Their Antioxidant Activities. Evid Based Complement Alternat Med. 2016;2016:7631864.
Lucini L, Kane D, Pellizzoni M, Ferrari A, Trevisi E, Ruzickova G, et al. Phenolic profile and in vitro antioxidant power of different milk thistle [Silybum marianum (L.) Gaertn.] cultivars. Ind Crops. Prod. 2016;83:11–6.
Muszyńska B, Łojewski M, Sułkowska-Ziaja K, Szewczyk A, Gdula-Argasińska J, Hałaszuk P. In vitro cultures of Bacopa monnieri and an analysis of selected groups of biologically active metabolites in their biomass. Pharm Biol. 2016;54(11):2443–53.
Maimoona A, Naeem I, Saddiqe Z, Jameel K. A review on biological, nutraceutical and clinical aspects of French maritime pine bark extract. J Ethnopharmacol. 2011;133(2):261–77.
Papagiannopoulos M, Wollseifen HR, Mellenthin A, Haber B, Galensa R. Identification and quantification of polyphenols in carob fruits (Ceratonia siliqua L.) and derived products by HPLC-UV-ESI/MSn. J Agric Food Chem. 2004;52(12):3784–91.
Koirala P, Jung HA, Choi JS. Recent advances in pharmacological research on Ecklonia species: a review. Arch Pharm Res. 2017;40(9):981–1005.
Panossian A, Wikman G, Sarris J. Rosenroot (Rhodiola rosea): Traditional use, chemical composition, pharmacology and clinical efficacy. Phytomedicine. 2010;17(7):481–93.
Bondonno CP, Croft KD, Ward N, Considine MJ, Hodgson JM. Dietary flavonoids and nitrate: effects on nitric oxide and vascular function. Nutr Rev. 2015;73(4):216–35.
Rees A, Dodd GF, Spencer JPE. The Effects of Flavonoids on Cardiovascular Health: A Review of Human Intervention Trials and Implications for Cerebrovascular Function. Nutrients. 2018;10(12):1852.
Rothwell JA, Urpi-Sarda M, Boto-Ordoñez M, Llorach R, Farran-Codina A, Barupal DK, et al. Systematic analysis of the polyphenol metabolome using the Phenol-Explorer database. Mol Nutr Food Res. 2016;60(1):203–11.
Kapil V, Rathod KS, Khambata RS, Bahra M, Velmurugan S, Purba A, et al. Sex differences in the nitrate-nitrite-NO(•) pathway: Role of oral nitrate-reducing bacteria. Free Radic Biol Med. 2018;126:113–21.
Haizlip KM, Harrison BC, Leinwand LA. Sex-based differences in skeletal muscle kinetics and fiber-type composition. Physiology (Bethesda). 2015;30(1):30–9.
Lundberg JO, Weitzberg E, Gladwin MT. The nitrate–nitrite–nitric oxide pathway in physiology and therapeutics. Nat Rev Drug Discov. 2008;7(2):156–67.
Olszowy-Tomczyk M. Synergistic, antagonistic and additive antioxidant effects in the binary mixtures. Phytochemistry Rev. 2020;19(1):63–103.
Lorenzo Calvo J, Alorda-Capo F, Pareja-Galeano H, Jiménez SL. Influence of Nitrate Supplementation on Endurance Cyclic Sports Performance: A Systematic Review. Nutrients. 2020;12(6):1796.
Baker ME, DeCesare KN, Johnson A, Kress KS, Inman CL, Weiss EP. Short-Term Mediterranean Diet Improves Endurance Exercise Performance: A Randomized-Sequence Crossover Trial. J Am Coll Nutr. 2019;38(7):597–605.
Nieman DC, Goodman CL, Capps CR, Shue ZL, Arnot R. Influence of 2-Weeks Ingestion of High Chlorogenic Acid Coffee on Mood State, Performance, and Postexercise Inflammation and Oxidative Stress: A Randomized, Placebo-Controlled Trial. Int J Sport Nutr Exerc Metab. 2018;28(1):55–65.
Muggeridge DJ, Sculthorpe N, James PE, Easton C. The effects of dietary nitrate supplementation on the adaptations to sprint interval training in previously untrained males. J Sci Med Sport. 2017;20(1):92–7.
Radak Z, Ishihara K, Tekus E, Varga C, Posa A, Balogh L, et al. Exercise, oxidants, and antioxidants change the shape of the bell-shaped hormesis curve. Redox Biol. 2017;12:285–90.
Braakhuis AJ, Hopkins WG. Impact of Dietary Antioxidants on Sport Performance: A Review. Sports Med. 2015;45(7):939–55.
Nikolaidis MG, Kerksick CM, Lamprecht M, McAnulty SR. Does vitamin C and E supplementation impair the favorable adaptations of regular exercise? Oxid Med Cell Longev. 2012;2012:707941.
Proteggente AR, Pannala AS, Paganga G, Lv B, Wagner E, Wiseman S, et al. The Antioxidant Activity of Regularly Consumed Fruit and Vegetables Reflects their Phenolic and Vitamin C Composition. Free Radic Res. 2002;36(2):217–33.
Cardarelli M, Rouphael Y, Pellizzoni M, Colla G, Lucini L. Profile of bioactive secondary metabolites and antioxidant capacity of leaf exudates from eighteen Aloe species. Ind Crops Prod. 2017;108:44–51.
Deng G-F, Lin X, Xu X-R, Gao L-L, Xie J-F, Li H-B. Antioxidant capacities and total phenolic contents of 56 vegetables. J Funct Foods. 2013;5(1):260–6.
Funding
NMAdU and HTB are supported by Australian Government Research Training Program Scholarships. No other funding was provided.
Author information
Authors and Affiliations
Contributions
NMAdU, AMH, AMC, MJN and JDB conceived the review, all authors conducted title and abstract screening, NMAdU and HTB conducted full-text review, data extraction and risk of bias assessment, NMAdU drafted the manuscript, which was critically revised by all authors. The author(s) read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
NMAdU, AMH, AMC, MJN and JDB are currently conducting a randomised controlled trial assessing the effects of consuming almonds, sultanas (raisins) and cranberries on endurance exercise performance and recovery, which is funded by a grant from the International Nut and Dried Fruit Council (INC) Foundation. However, the INC did not provide support for this review and were not involved in any aspect of its conception, collection, analysis or interpretation.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1.
Database search strategies. Verbatim search strategy used in each database.
Additional file 2.
Cochrane Risk of Bias Tool 2.0 Summary. Assessments of overall and domain-specific bias of included studies.
Additional file 3.
Study Characteristics and Summary Table. Description: Key characteristics and results of included studies.
Additional file 4.
L-citrulline meta-analysis and sub-group analyses. Description: L-citrulline meta-analysis and sub-group analyses table.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
d’Unienville, N.M.A., Blake, H.T., Coates, A.M. et al. Effect of food sources of nitrate, polyphenols, L-arginine and L-citrulline on endurance exercise performance: a systematic review and meta-analysis of randomised controlled trials. J Int Soc Sports Nutr 18, 76 (2021). https://doi.org/10.1186/s12970-021-00472-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12970-021-00472-y