
Abstract
Background
This study is to investigate the association of fibroblast growth factor receptor 2 (FGFR2) rs2981582, trinucleotide-repeat-containing 9 (TNRC9) rs3803662, rs12443621, and leukocyte-specific protein 1 (LSP1) rs3817198 polymorphisms with breast cancer and mammographic density in Han Chinese population.
Methods
TaqMan Single Nucleotide Polymorphism (SNP) Genotyping Assays and unconditional logistic regression analysis were used to examine these SNPs in 105 breast cancer cases and 382 controls.
Results
The genotype frequencies of rs12443621 and rs2981582 were significantly different between controls and cases (P = 0.017 and 0.006, respectively). Subjects carrying G allele of rs12443621 had increased breast cancer risk (AG vs AA: OR = 2.017, 95 % CI = 0.910–4.471; GG vs AA: OR = 2.684, 95 % CI = 1.318–5.463). Subjects carrying an allele of rs2981582 had reduced breast cancer risk (GA vs GG: OR = 0.444, 95 % CI = 0.262–0.752; AA vs GG: OR = 0.579, 95 % CI = 0.342–0.983). rs3803662 and rs3817198 SNPs did not significantly differ between cases and controls (P = 0.408 and 0.116, respectively). Interestingly, the AA genotype of rs2981582 was also associated with reduced mammographic densities (P = 0.0092, 95 % CI = 0.334–0.926).
Conclusions
Our findings indicate that the GG genotype of rs12443621 is associated with increased breast cancer risk whereas the GA and AA genotypes of rs2981582 are reduced risk in Han Chinese population.
Similar content being viewed by others

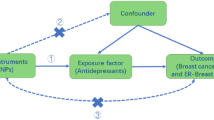

Background
Breast cancer is one of the most common malignancies and a leading cause of cancer death in women worldwide, with over one million new cases each year [1]. In China, breast cancer has increased rapidly in recent years and become the most common cancer for women in major cities [2]. In Heilongjiang Province, which is located in Northeast China, there are nearly 6000 new cases of breast cancer every year accounting for 15 % of female malignancies in the area [3]. Therefore, it is necessary to identify risk factors for breast cancer in this Chinese population.
The development and progression of breast cancer is a multi-step complicated process that involves both genetic and epigenetic factors. Although significant advancement has been made in understanding the genetic susceptibility to breast cancer, known susceptibility genes account for less than a quarter of familial breast cancer risk. To further identify novel susceptibility alleles associated with breast cancer, genome-wide association studies (GWAS) have been performed. One of these studies was published on Nature [4], which identifies trinucleotide-repeat-containing 9 (TNRC9), fibroblast growth factor receptor 2 (FGFR2), mitogen-activated protein kinase 1 (MAP3K1), and leukocyte-specific protein 1 (LSP1) as four novel loci with consistent evidence of association with breast cancer.
Since then, a number of case-control or cohort studies have been carried out to further examine the association of the single nucleotide polymorphisms (SNPs) with breast cancer, but the results are not completely consistent. For example, a cohort study in American population found that the AG/GG genotypes of TNRC9 rs12443621 and the CT/TT genotype of FGFR2 rs2981582 had a decreased risk for death in breast cancer patients [5]. In contrast, another cohort study in Swedish population found that the AG/GG genotypes of TNRC9 rs12443621 and the AA genotypes of FGFR2 rs2981582 both had increased breast cancer risk [6]. A case-control study in Tunisian population reported that subjects with AA genotype of FGFR2 rs2981582 had increased risk of breast cancer [7], but TNRC9 rs12443621 was not significantly associated with breast cancer risk. Strong ethnic differences have been observed in breast cancer risk. Racial/ethnic differences and environmental factors that vary among populations may affect the associations between SNPs and risk of breast cancer by modulating complex interactions between various genes. Therefore, more studies are needed to clarify the relationship between these SNPs and breast cancer risk.
Mammographic density has been shown to be strongly linked to breast cancer risk [8]. Women with high mammographic density have increased risk for breast cancer as compared to women with lower mammographic density. Because both breast cancer and mammographic density are influenced by genetic factors, they may share some genetic determinants. A recent study showed that women with at least one G allele of rs12443621 in TRC9 had higher mammographic density than women with two alleles [9]. They also found that women with at least one T allele of rs2981582 in FGFR2 had non-significantly decreased mammographic density than women with two C alleles.
To have a better understanding of the association of FGFR2 rs2981582, TNRC9 rs3803662, rs12443621, and LSP1 rs3817198 polymorphisms with breast cancer, we investigated these SNPs in Han Chinese women in Heilongjiang Province and analyzed their association with breast cancer risk and mammographic density.
Methods
Study population
All samples were collected at the First Affiliated Hospital of Harbin Medical University in 2010 between June and November. A total of 487 participants were recruited in this study. The case group was comprised of 105 female patients with histopathologically diagnosed breast cancer. Patients with a history of cancer, tumor chemotherapy, or radiotherapy were excluded from this study. The control group was comprised of 382 age-matched healthy women. Ninety of the breast cancer cases and 229 of controls had mammographic X-ray. All subjects gave written consent for participation in this research. This study was approved by the Ethical Committee of the First Hospital Affiliated to Haerbing Medical University.
DNA extraction and TaqMan SNP Genotyping Assays
EDTA-anti-coagulated venous blood samples were preserved at −70 °C. Genomic DNA was isolated from whole blood using the Wizard® kit (Promega, Madison, WI, USA) according to the manufacturer’s protocol. Blinded genotyping of FGFR2 rs2981582, TNRC9 rs3803662, rs12443621, and LSP1 rs3817198 was carried out using TaqMan SNP Genotyping Assays (Applied Biosystems, Foster City, CA, USA) with Mini Option2 Real Time PCR System (BioRad, Hercules, California, USA). Assays were performed with TaqMan Universal Master Mix, TaqMan probe, and 50 ng of DNA per reaction. PCR conditions were provided by the manufacturer: 5 min initial denaturation at 94 °C followed by 45 cycles of 94 °C denaturation for 15 s and 60 °C annealing/extension for 1 min.
Reproductive factors
Information on age, age at menarche, age at menopause, breast-feeding, parity, and miscarriages/abortions was assessed from the questionnaire. The number of miscarriages/abortions was categorized into three groups: 0, 1–2, and 3 or more.
Mammographic density
Mediolateral oblique (MLO) and craniocaudal (CC) view digital mammograms were evaluated by three radiologists specialized in mammographic diagnosis. Mammographic density was described by using the Breast Imaging Reporting and Data System (BI-RADS, American College of Radiology) four-category terminology [10]: D1, less than 25 % glandular (category 1), D2, 25–50 % glandular (category 2), D3, 51–75 % glandular (category 3), and D4 greater than 75 % glandular (category 4). The evaluation was blinded to ensure accuracy. For a few films on which there was disagreement in reporting, final reporting was made after the discussion among the radiologists.
Statistical analysis
Quantitative data were expressed as mean ± SD and analyzed using ANOVA or rank sum test. Categorical data were analyzed using chi-square test. The association between the genotypes and breast cancer risk or mammographic density was evaluated by unconditional logistic regression analysis and expressed as odds ratio (OR) and their 95 % confidence intervals (CI) using SPSS 18.0 software (SPSS Inc., Chicago, IL, USA). Throughout the analysis, a two-sided P value less than 0.05 was considered to be statistically significant.
Results
Distribution of TNRC9, FGFR2, and LSP1 SNPs in subjects
Using TaqMan SNP Genotyping Assays, we evaluated FGFR2 rs2981582, TNRC9 rs3803662, TNRC9 rs12443621, and LP1 rs3817198 polymorphisms in 487 subjects, including 105 breast cancer cases and 382 controls. The genotype frequencies for these SNPs in cases and controls and the allele frequencies in all subjects are listed in Table 1. All four SNPs conformed to the Hardy-Weinberg equilibrium.
Reproductive factors and breast cancer risk
Reproductive characteristics of cases and controls are summarized in Table 2. Case group had a significantly younger age at menarche compared to control group (P = 0.022). Controls had longer average breast-feeding duration (0.000) and higher rate of breast-feeding history (0.009). Controls also had a larger number of parity (0.03) and higher rate of childbearing history compared to cases (0.006). These factors were hence included in the multivariate analyses. All the other factors were similarly distributed between cases and controls.
TNRC9, FGFR2, and LSP1 SNPs and breast cancer risk
We examined the association between FGFR2 rs2981582, TNRC9 rs3803662, TNRC9 rs12443621, and LP1 rs3817198 and breast cancer risk, and the results are shown in Table 1. FGFR2 rs2981582 and TNRC9 rs12443621 genotype frequencies were significantly different between case and control groups (P = 0.006 and 0.017, respectively). The AG and GG genotypes of rs12443621 in TNRC9 were more prevalent among cases than among controls (AG vs AA: OR = 2.017, 95 % CI = 0.910–4.471; GG vs AA: OR = 2.684, 95 % CI = 1.318–5.463). The GA and AA genotypes of rs2981582 in FGFR2 were more prevalent among controls than among cases (GA vs GG: OR = 0.444, 95 % CI = 0.262–0.752; AA vs GG: OR = 0.579, 95 % CI = 0.342–0.983). After adjusting for age and reproductive risk factors, subjects carrying G allele of rs12443621 in TNRC9 were found to have a higher breast cancer risk compared with those carrying AA genotype (AG vs AA: OR = 0.621, 95 % CI = 0.075–5.129; GG vs AA: OR = 1.569, 95 % CI = 0.233–10.556).
TNRC9 rs3803662 and LSP1 rs3817198 genotype distribution in cases and controls was not significantly different (P = 0.408 and 0.116, respectively).
TNRC9, FGFR2, and LSP1 SNPs and mammographic density
Analysis of mammographic density using rank sum test showed a significant difference between cases and controls (Table 3), with higher mammographic density in case patients (χ 2 = 4.4530, P = 0.0348). As shown in Table 4, the distribution of FGFR2 rs2981582 polymorphism, but not the other three SNPs, was significantly different among the four mammographic density categories (P = 0.0342). Subjects carrying AA genotype of rs2981582 in FGFR2 were associated with reduced mammographic density (Table 5, P = 0.0092, 95 % CI = 0.334–0.926). After adjusting for age and reproductive risk factors, this remained significant (Table 6, P = 0.0154, 95 % CI = 0.294–0.893).
Discussion
In the present study, we found that early age at menarche is associated with increased breast cancer risk, whereas breast-feeding and parity are associated with reduced breast cancer risk. Multiple studies have examined the relationship between reproductive factors and breast cancer risk, but the results are somewhat inconsistent. A case-control study in Japanese population reported that early age at menarche, late age at first birth, and premenopausal status are significantly associated with breast cancer risk, while high parity is a protective factor [11]. A meta-analysis revealed that induced abortion is a risk factor for breast cancer [12]. Stuver and coworkers failed to observe any protective effects of lactation or duration of lactation against breast cancer risk [13], although there are other studies showing that lactation is linked to reduced risk of breast cancer [14]. Breast cancer is a complicated disease affected by genetic, environmental, and economic conditions as well as lifestyles. Variations in these factors may contribute to the inconsistency in research findings on the association between reproductive factors and breast cancer.
Our study revealed a significant association between TNRC9 rs12443621 and breast cancer risk. Specifically, the GG genotype of TNRC9 rs12443621 had increased risk of breast cancer. The rs12443621 SNP of TNRC9 (also named TOX3) is located at chromosome 16q12 [4]. The function of TNRC9 is unclear, but a recent paper has reported that TNRC9 down-regulates BRCA1 expression and promotes breast cancer aggressiveness [7]. The association between TNRC9 polymorphisms and breast cancer risk remains controversial. A meta-analysis in 2011 found that TNRC9 rs3803662, but not TNRC9 rs12443621, polymorphism was significantly correlated with breast cancer risk [15], which differs from our results. However, our findings agree with a recently published cohort study in Swedish population, which reports an increased risk of breast cancer for subjects carrying AG/GG genotypes of TRNC9 rs12443621 [6]. The reason for the inconsistency among published data regarding the association between TRNC9 rs12443621 and breast cancer risk is hard to decipher but could be related to the multifaceted characteristics of breast cancer.
FGFR2 is a member of the FGFR family of receptor tyrosine kinases that has been found to be overexpressed in some breast cancer cell lines [16]. Amplification of FGFR2 gene occurs in a small subset of breast cancer [17]. Our study reveals an interesting finding that the GA and AA genotypes of FGFR2 rs2981582 are associated with reduced risk of breast cancer. This is surprising, since most studies reported that the T allele of FGFR2 rs2981582 is associated with increased breast cancer risk [18, 19]. However, our results are consistent with another case-control study in the Chinese population, which also reveals the AA genotype of rs2981582 as a protective factor against breast cancer [20]. It is worthwhile to mention that we also observed a strong association between the GA and AA genotypes of FGFR2 rs2981582 and decreased mammographic density. Meanwhile, our data showed that lower mammographic density is linked to lower risk of breast cancer. These findings are consistent with each other. One limitation of the current study was that the sample size for breast cancer cases was relatively small (105 cases were included in this study), and all patients were from Heilongjiang Province in Northeast China. We cannot rule out the potential instability rising from a relatively small number of patients. Further studies are necessary to elude the reasons for the disparity between our results and some of the earlier reports.
Conclusions
In conclusion, we detected a significant association between the GG genotype of TNRC9 rs12443621 and elevated breast cancer risk. Moreover, the GA and AA genotypes of FGFR2 rs2981582 appear to be associated with lower mammographic density and reduced breast cancer risk. These data provide support for the potential importance of these polymorphisms in breast cancer biology. Further studies are needed to define the role of these SNPs in breast cancer and test the possibility of using them as biomarkers for diagnostic, prognostic, or therapeutic purposes in Chinese Han population.
Abbreviations
- CC:
-
craniocaudal
- CI:
-
confidence intervals
- FGFR2 :
-
fibroblast growth factor receptor 2
- GWAS:
-
genome-wide association studies
- LSP1 :
-
leukocyte-specific protein 1
- MAP3K1:
-
mitogen-activated protein kinase 1
- MLO:
-
mediolateral oblique
- OR:
-
odds ratio
- SNPs:
-
single nucleotide polymorphisms
- TNRC9 :
-
trinucleotide-repeat-containing 9
References
Chu D, Lu J. Novel therapies in breast cancer: what is new from ASCO 2008. J Hematol Oncol. 2008;1:16. doi:10.1186/1756-8722-1-16.
Hong W, Dong E. The past, present and future of breast cancer research in China. Cancer Lett. 2014. doi:10.1016/j.canlet.2014.04.007.
Ying-ji L, Xu-dong D, Xi-wen S, Hui-li H, Lu A-M, Lu Y, et al. The incidence and mortality analysis malignancies in 1995 and 2000 year in major cities of Heilongjiang Province. Bull Chin Cancer. 2002;11(1):29–30.
Easton DF, Pooley KA, Dunning AM, Pharoah PD, Thompson D, Ballinger DG, et al. Genome-wide association study identifies novel breast cancer susceptibility loci. Nature. 2007;447(7148):1087–93. doi:10.1038/nature05887.
Bayraktar S, Thompson PA, Yoo SY, Do KA, Sahin AA, Arun BK, et al. The relationship between eight GWAS-identified single-nucleotide polymorphisms and primary breast cancer outcomes. Oncologist. 2013;18(5):493–500. doi:10.1634/theoncologist.2012-0419.
Butt S, Harlid S, Borgquist S, Ivarsson M, Landberg G, Dillner J, et al. Genetic predisposition, parity, age at first childbirth and risk for breast cancer. BMC Res Notes. 2012;5:414. doi:10.1186/1756-0500-5-414.
Shan J, Mahfoudh W, Dsouza SP, Hassen E, Bouaouina N, Abdelhak S, et al. Genome-wide association studies (GWAS) breast cancer susceptibility loci in Arabs: susceptibility and prognostic implications in Tunisians. Breast Cancer Res Treat. 2012;135(3):715–24. doi:10.1007/s10549-012-2202-6.
McCormack VA, dos Santos Silva I. Breast density and parenchymal patterns as markers of breast cancer risk: a meta-analysis. Cancer Epidemiol Biomarkers Prev. 2006;15(6):1159–69. doi:10.1158/1055-9965.EPI-06-0034.
Woolcott CG, Maskarinec G, Haiman CA, Verheus M, Pagano IS, Le Marchand L, et al. Association between breast cancer susceptibility loci and mammographic density: the multiethnic cohort. Breast Cancer Res. 2009;11(1):R10. doi:10.1186/bcr2229.
American College of Radiology. Breast imaging reporting and data system (BI-RADS). 4th ed. Reston: American College of Radiology; 2003.
Nagata C, Hu YH, Shimizu H. Effects of menstrual and reproductive factors on the risk of breast cancer: meta-analysis of the case-control studies in Japan. Jpn J Cancer Res. 1995;86(10):910–5.
Brind J, Chinchilli VM, Severs WB, Summy-Long J. Induced abortion as an independent risk factor for breast cancer: a comprehensive review and meta-analysis. J Epidemiol Community Health. 1996;50(5):481–96.
Stuver SO, Hsieh CC, Bertone E, Trichopoulos D. The association between lactation and breast cancer in an international case-control study: a re-analysis by menopausal status. Int J Cancer. 1997;71(2):166–9.
do Carmo Franca-Botelho A, Ferreira MC, Franca JL, Franca EL, Honorio-Franca AC. Breastfeeding and its relationship with reduction of breast cancer: a review. Asian Pac J Cancer Prev. 2012;13(11):5327–32.
Chen MB, Wu XY, Shen W, Wei MX, Li C, Cai B, et al. Association between polymorphisms of trinucleotide repeat containing 9 gene and breast cancer risk: evidence from 62,005 subjects. Breast Cancer Res Treat. 2011;126(1):177–83. doi:10.1007/s10549-010-1114-6.
Tannheimer SL, Rehemtulla A, Ethier SP. Characterization of fibroblast growth factor receptor 2 overexpression in the human breast cancer cell line SUM-52PE. Breast Cancer Res. 2000;2(4):311–20. doi:10.1186/bcr73.
Jain VK, Turner NC. Challenges and opportunities in the targeting of fibroblast growth factor receptors in breast cancer. Breast Cancer Res. 2012;14(3):208. doi:10.1186/bcr3139.
Jia C, Cai Y, Ma Y, Fu D. Quantitative assessment of the effect of FGFR2 gene polymorphism on the risk of breast cancer. Breast Cancer Res Treat. 2010;124(2):521–8. doi:10.1007/s10549-010-0872-5.
Murillo-Zamora E, Moreno-Macias H, Ziv E, Romieu I, Lazcano-Ponce E, Angeles-Llerenas A, et al. Association between rs2981582 polymorphism in the FGFR2 gene and the risk of breast cancer in Mexican women. Arch Med Res. 2013;44(6):459–66. doi:10.1016/j.arcmed.2013.08.006.
Chen F, Lu M, Xue Y, Zhou J, Hu F, Chen X, et al. Genetic variants of fibroblast growth factor receptor 2 (FGFR2) are associated with breast cancer risk in Chinese women of the Han nationality. Immunogenetics. 2012;64(1):71–6. doi:10.1007/s00251-011-0564-2.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
YC and CS carried out the molecular genetic studies, participated in the sequence alignment, and drafted the manuscript. CS carried out the immunoassays. YC participated in the sequence alignment. YC and CS participated in the design of the study and performed the statistical analysis. QG conceived of the study, participated in its design and coordination, and helped to draft the manuscript. All authors read and approved the final manuscript.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Chen, Y., Shi, C. & Guo, Q. TNRC9 rs12443621 and FGFR2 rs2981582 polymorphisms and breast cancer risk. World J Surg Onc 14, 50 (2016). https://doi.org/10.1186/s12957-016-0795-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12957-016-0795-7