
Abstract
Immunotherapy has emerged to play a rapidly expanding role in the treatment of cancers. Currently, many clinical trials of therapeutic agents are on ongoing with majority of immune checkpoint inhibitors (ICIs) especially programmed death receptor 1 (PD-1) and its ligand 1 (PD-L1) inhibitors. PD-1 and PD-L1, two main immune checkpoints, are expressed at high levels in thymic epithelial tumors (TETs) and could be predictors of the progression and immunotherapeutic efficacy of TETs. However, despite inspiring efficacy reported in clinical trials and clinical practice, significantly higher incidence of immune-related adverse events (irAEs) than other tumors bring challenges to the administration of ICIs in TETs. To develop safe and effective immunotherapeutic patterns in TETs, understanding the clinical properties of patients, the cellular and molecular mechanisms of immunotherapy and irAEs occurrence are crucial. In this review, the progress of both basic and clinical research on immune checkpoints in TETs, the evidence of therapeutic efficacy and irAEs based on PD-1 /PD-L1 inhibitors in TETs treatment are discussed. Additionally, we highlighted the possible mechanisms underlying irAEs, prevention and management strategies, the insufficiency of current research and some worthy research insights. High PD-1/PD-L1 expression in TETs provides a rationale for ICI use. Completed clinical trials have shown an encouraging efficacy of ICIs, despite the high rate of irAEs. A deeper mechanism understanding at molecular level how ICIs function in TETs and why irAEs occur will help maximize the immunotherapeutic efficacy while minimizing irAEs risks in TET treatment to improve patient prognosis.
Similar content being viewed by others
Introduction
Thymic epithelial tumors (TETs) are the most common neoplasms of the anterior mediastinum in adults [1]. Histological classification includes thymomas (Type A, AB, B1, B2, B3), thymic carcinomas (Type C), and thymic neuroendocrine tumors, while Massaoka and Tumor-Node-Metastasis staging system concern localization of the involved areas [2]. TET treatment is a paradigm of multidisciplinary cooperation among surgeons, clinicians, and pathologists from establishing the diagnosis to determining therapeutic strategies, especially for complicated cases with autoimmune diseases [3]. Total thymectomy is currently the preferred strategy for TETs. For relapsed or refractory patients with local invasion or distant metastasis, cisplatin combined with anthracycline or paclitaxel has become the first choice [4]. However, the efficacy of chemotherapy is limited, and serious adverse reactions are observed. Targeted drugs such as receptor tyrosine kinase inhibitors and antiangiogenic agents are considered beneficial supplements after chemotherapy for advanced TET patients, but the efficacy is also not satisfactory. And a lack of utilizable genomic alterations in TETs hinders in the development of targeted therapies [5]. Immunotherapy plays an important role in tumor treatment, and the discovery of immune checkpoints drives tumor immunotherapy to a new stage [6]. Among all immune checkpoints, programmed death receptor 1 (PD-1) and its ligand 1 (PD-L1), and cytotoxic T lymphocyte-associated protein 4 (CTLA-4) have attracted increasing attention in TETs [7, 8]. CTLA-4 is expressed in TETs and positively correlates with a poor patient prognosis; moreover, it is associated with the pathogenesis of autoimmune diseases (ADs), such as myasthenia gravis [9]. Several clinical trials of CTLA-4 inhibitors in TETs are in progress, but the efficacy needs to be further examined in clinical practice of TET management. Accumulating evidence has confirmed high PD-1/PD-L1 expression in TETs that is associated with worse clinical characteristics and a poor patient prognosis [10], showing great immunotherapeutic potential. PD-1/PD-L1 inhibitors have been used to treat a variety of tumors, such as melanoma and non-small cell lung cancer (NSCLC), and have shown significant efficacy [11,12,13]. Many clinical trials evaluating the efficacy of PD-1/PD-L1 inhibitors against TETs have also been completed or are in progress. Completed clinical trials reported both encouraging therapeutic benefits and worrisome immune-related adverse events (irAEs), making immunotherapy for TETs highly controversial. The National Comprehensive Cancer Network Guidelines version 2.2022 only recommends pembrolizumab as a second-line therapeutic strategy for thymic carcinoma (TC) with vigilance to the high incidence of irAEs, and no other immune checkpoint inhibitors (ICIs) are recommended for TET treatment [14]. This review summarizes the current research progress in ICIs as TET treatments, highlighting irAEs, the potential mechanisms, and prevention strategies. In addition to the deficiencies of current research, some viewpoints that are worthy of further consideration are noted, which will help lay the foundation and identify directions for future research on ICIs in TETs, accumulating more evidence for clinical practice.
Immunotherapeutic agents for TETs
The emergence and rapid advances in immunotherapies such as ICIs, cancer vaccines, cytokine-based therapies and adoptive cell therapies have significantly changed the treatment of cancers [15] (Fig. 1). They enhance antitumor immunity by blocking inhibitory signaling from immune checkpoints or by enhancing activity of stimulatory signaling, producing T cells with augmented responses toward tumor cells [16, 17]. Wilms’ tumor gene 1 (WT1), a small peptide overexpressed in TETs, can regulate cell reproduction and apoptosis, was reported to be involved in TETs pathogenesis. A phase II clinical trial that enrolled 18 patients with TETs was conducted to examine the efficacy of WT1-peptide vaccine [18] (Fig. 1. D). However, no patients achieved a complete or partial response, although 75% of patients had stable disease. Additionally, an in vivo experiment in mice [19] reported that adoptive transfer of B cells halts thymoma growth, which implies the potential of adoptive cell therapy in TETs. High expression of CD70, a protein belonging to the tumor necrosis family, was reported in TETs, and CD70-targeted CAR T cells were confirmed both in vitro and in vivo to be effective against tumors, indicating the possibility of CD70-targeted CAR T cell therapy for TETs [20,21,22]. Cytokines, such as interleukin (IL)-15 and interferon α can amplify patients’ antitumor immune responses [17], and activators of IL-15 in TETs are being explored in clinical trial (NCT04234113) (Fig. 1 E). Despite theoretically various immunotherapies for TETs, most of them are at experimental or preclinical level. The majority of evidence has been obtained from ICIs, especially PD-1/PD-L1 inhibitors (Fig. 1A), in both clinical trials and clinical practice of TET treatment.

Different forms of anticancer immunotherapy. A. ICIs, especially PD-1/PD-L1 inhibitors, are the main immunotherapies used in TET treatment and have accumulated the most evidence. B-E. Adoptive cell therapies, including CAR-T, TIL, TCR-T and NKC therapies, and different sources of therapeutic cells are obtained, modified or screened and expanded for infusion back into the patients. F. Therapeutic vaccines are designed based on discovered tumor neoantigens. The infused tumor vaccines induce an immune response to tumor cells and enhance antitumor immunity. G. Cytokines with immune activation functions, such as IL-2 and IFN-α/β, are recombined and synthesized in vitro and then infused into patients, which enhance the antitumor immune response. CAR: Chimeric antigen receptor; TIL: Tumor-infiltrating lymphocyte; TCR: T cell receptor; NKC: Natural killer cell
Rationale for using ICIs as TET treatments
The physiology and pathophysiology of the thymus
Primitive progenitor T cells from the embryonic liver or bone marrow hematopoietic system migrate to the thymus, where they develop into naïve T cells that are immune-effective against allogeneic antigens and immune-tolerant to self-antigens after positive and negative selection [23]. Positive selection occurs in the thymic cortex by recognizing autogenic tissue-specific antigens (TSAs) presented by epithelial cells and major histocompatibility complex (MHC) molecules. This process allows CD4−CD8− T cells to become CD4+CD8+ T cells, which also acquire T cell receptor (TCR) rearrangement and gain MHC restriction. Then, CD4+CD8+ T cells enter the thymic medulla, where they undergo negative selection mainly controlled by autoimmune regulator (AIRE) genes. Thymic medullary epithelial cells with high AIRE expression are prone to apoptosis and release TSAs that will be captured by dendritic cells and presented to CD4+CD8+ T cells through MHC. T cells that overreact undergo apoptosis, while surviving T cells recognize TSAs presented by MHC class I/II and develop into CD4+CD8− or CD4−CD8+ T cells. This process is known as central immune tolerance, and these T cells subsequently egress to the peripheral circulation to become recent thymic emigrants, which plays an important role in building a complete immune system [23] (Fig. 2 A). In TETs, however, immune tolerance is rendered dysfunctional because of the decreased expression of AIRE, MHC and the altered thymic architecture. Consequently, an increased number of immature CD8+ T cells but decreased numbers of immature CD4+ T cells and regulatory T cells (Tregs) are observed [24]. These changes promote the release of autoreactive T cells, thus disturbing peripheral homeostasis, which, in turn, predisposes patients to autoimmunity or causes ADs (Fig. 2 B). Additionally, some cytokines, chemokines produced by tumor cells that induce cross-reactions between tumor antigens and TSAs, and the structural similarities between them, which promote autoantibody production, are other proposed mechanisms to explain autoimmunity in TETs [25, 26]. For instance, thymoma overexpresses mid-sized neurofilament gene, which shares sequences encoding acetylcholine receptors and titin epitopes and is often correlated with myasthenia gravis [26].

Naïve T cell development in the normal thymus and thymoma. A. A healthy thymus with a normal structure and thymic microenvironment determines normal T cell development and maturation and continuously exports normal naïve T cells to establish normal immune function. B. Thymic epithelial tumors with distorted structures and disrupted microenvironments lack components that are necessary for positive and negative selection. Naïve T cells do not complete the central immune tolerance and emigrate to become autoreactive RTEs, which directly or indirectly lead to autoimmune diseases through the cellular or humoral immune systems, respectively. Pro T: Progenitor T cells; AChR: Acetylcholine receptor; AIRE: Autoimmune regulator effectors; RTEs: Recent thymic emigrants; DC: Dendritic cells; SLO: Secondary lymphoid organs
PD-1 and PD-L1 are involved in T cell function
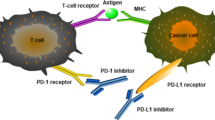
Activation of naïve T cells requires dual signals. The first is a specific antigen recognition signal that is acquired and processed by antigen-presenting cells and then recognized by T cells after an interaction with MHC molecules; the second is a costimulatory signal derived from B7-1/2 expressed by antigen-presenting cells and interacts with CD28 expressed on T cells [27] (Fig. 3 A). Only T cells exposed to both stimuli are activated, including tumor-infiltrating T cells [28]. Moreover, immunologists have discovered negative costimulatory signals, also known as immune checkpoints [6, 29, 30]. Under physiological conditions, immune checkpoints transmit signals that inhibit T cell activation, avoiding autoimmune responses, a process called peripheral immune tolerance. However, in tumor and inflammatory sites, immune checkpoints impair antitumor immunity and antiinfection ability, leading to chronic inflammation and tumor progression. Among all immune checkpoints, PD-1/PD-L1 represents the best studied checkpoint in TETs. PD-1 and PD-L1 are a receptor and ligand, respectively, mediating the cosuppressive signaling of T cells, immunosuppression of T cells and tumor immune escape (Fig. 3B). PD-1 is mainly expressed on immature CD4–/CD8– thymocytes and activated CD4+/CD8+ T cells [31]. PD-L1 is constitutively expressed in different cells, such as dendritic cells, mediating peripheral immune tolerance [6]. However, PD-L1 is also expressed at high levels in inflammatory sites and tumor cells, impairing T cell-mediated immune function. PD-1 aggregates with TCR after binding to PD-L1 and associates with the Src homology 2 domain-containing tyrosine phosphatase 2. They compose negative costimulatory microclusters to induce the dephosphorylation of proximal TCR signaling molecules, inhibiting T cell activation [32] and inducing their differentiation into Tregs or apoptosis. Activation of PD-1/PD-L1 signaling has also been implicated in driving T cell exhaustion by limiting glucose and amino acid metabolism [33]. Therefore, blocking the PD-1/PD-L1 pathway may restore the antitumor effect of T cells (Fig. 3 C, D), which is also the rationale for using ICIs to treat tumors, including TETs that highly express PD-L1 [34].

PD-1/PD-L1 signaling in tumor immune tolerance. A. The mechanism of PD-1/PD-L1-mediated inhibition of T cell activation and PD-1/PD-L1 blocker-mediated T cell function restoration (between APCs and T cells). B. The mechanism of PD-1/PD-L1-mediated tumor immune tolerance and PD-1/PD-L1 blocker-mediated antitumor activity. Blockade of PD-1/PD-L1 alone and in combination with anti-TGF-β restores immune-exhausted T cells. CTLA-4: Cytotoxic T lymphocyte-associated antigen 4; APC: Antigen-presenting cells; MHC: Major histocompatibility complex; TCR: T cell receptor; SHP2: Src homology 2 domain-containing tyrosine phosphatase 2
The clinical significance of PD-1/PD-L1 in TETs
TETs express PD-1 and PD-L1 at high levels ranging from 18–100% [7, 35,36,37,38] (Table 1), which differs between different Masaoka stages and TET subtypes, and usually a higher level is observed in thymic carcinoma (TC) than thymoma [39]. Possible explanations for significantly different expression levels include various detection methods and samples, different proportions of pathological subtypes, different diagnostic criteria for thymoma subtypes at different periods and different positive judgment criteria [40, 41]. In all, PD-L1 is expressed at high levels with a heterogeneous distribution among different subtypes of TETs. Additionally, PD-L1 has been revealed as a predictor of the response to TET immunotherapy. Through genomic and transcriptomic profiling of TET samples in the pembrolizumab treatment cohort, researchers found differentiate gene or molecular alterations associated with PD-L1 expression between responders and nonresponders [42, 43]. The relationship between PD-1/PD-L1 and the pathological characteristics or prognosis of patients with TETs is controversial; however, high PD-1/PD-L1 expression seems to indicate worse pathological characteristics and a poor patient prognosis in terms of both overall survival (OS) and progression-free survival (PFS) [44, 45]. Furthermore, PD-L1 expression is positively correlated with EMT-related indicators, which potentially predict the asymptomatic survival of patients [46].
Clinical exploration of PD-1/PD-L1 inhibitors in TET treatment
Completed clinical trials
PD-1/PD-L1 inhibitors have been shown to be effective antitumor agents and are approved for the treatment of various solid tumors by the Food and Drug Administration [11,12,13]. Clinical trials evaluating the efficacy of PD-1 antibodies (pembrolizumab and nivolumab) and a PD-L1 antibody (avelumab) against TETs have also accumulated some evidence. Four clinical trials have been completed and provide worthwhile information (Table 2), and the others are still in process (Table 3). It seemed that researchers considered the relatively better survival prognosis of TETs compared with other solid tumors, since most clinical trials adopt the overall/object remission rate (ORR) as the primary endpoint instead of PFS or OS.
Pembrolizumab
Giaccone et al. [61] conducted a phase II clinical trial evaluating the efficacy of pembrolizumab in patients with recurrent TC. Patients with a history of ADs or immunodeficiency were excluded. Among 40 evaluable patients, an overall disease control rate of 22.5% was observed, with a disease progression rate of 25%. The median PFS of the PD-L1high group was 4.2 months with an unachieved median OS, which was significantly higher than that of patients in the PD-L1low group, with a median PFS of 2.9 months and a median OS of 15.5 months (p < 0.01), suggesting that the higher the PD-L1 expression level, the better the response to ICIs. However, irAEs were observed in all patients, in which 6 experienced more than one irAE and 6 had severe irAEs, such as myocarditis and hyperglycemia, but no deaths related to irAEs were reported. Giaccone et al. [65] updated the results of the long-term follow-up and reported that pembrolizumab induced durable responses in patients with TET, lasting approximately 3 years with a median survival of more than 2 years, and the 5-year survival rate was 18%. Notably, the incidence of severe irAEs (15%) did not increase significantly over time.
Cho et al. [62] included 33 patients with advanced TET in an open-label phase II clinical trial to evaluate the efficacy of pembrolizumab. Enrolled patients had received at least one first-line therapy, and those with a history of severe autoimmunity were excluded. Of seven patients with thymoma, two achieved partial responses, and five had stable diseases. Of the 26 patients with TC, five achieved partial responses, and 14 had stable diseases. The median duration of response was 9.7 months in patients with TC but was not reached in patients with thymoma. The median PFS was 6.1 months for both groups, and the median OS for patients with TC was 14.5 months but was not reached in patients with thymoma. Consistent with the results reported by Giaccone et al. [61], patients with higher PD-L1 expression had a better response to immunotherapy. Both Cho et al. [62] and Giaccone et al. [60, 61] demonstrated that pembrolizumab yielded encouraging antitumor activity with durable response in refractory, metastatic, or relapsed TETs in their clinical trials and updated follow-up data. They reported a significant correlation between high PD-L1 expression and better response to pembrolizumab in TETs and found that patients with durable responses had high PD-L1 expression. Some research reported durable response to immunotherapy in other solid tumors such as non-small cell lung cancer, breast cancer, and prostate cancer [66,67,68,69], and tumor mutational burden, microsatellite instability and high expression of immune checkpoint might contribute to this phenomenon. Some studies also demonstrated that TETs with microsatellite instability existed and might be sensitive to immunotherapy [70, 71]. Recently, Repetto et al. [72] reported a thymic carcinoma with Lynch syndrome, which had microsatellite instability and achieved durable response to avelumab and axitinib combination therapy. Collectively, TET patients with microsatellite instability and high expression of immune checkpoint might obtain durable response in immunotherapy.
Avelumab
Rajan et al. [63] evaluated the efficacy of avelumab in 8 patients with TETs without a history of autoimmunity. Four of the seven patients with thymoma responded, and the other three had stable disease. Responses were observed after the administration of a single dose of avelumab to four patients, who also discontinued dosing due to severe irAEs. Patient 1 developed grade 3 CPK elevation and grade 1 transaminitis; patient 2 experienced elevated CPK and myositis; patient 3 developed the worst irAEs including grade 2 dysphagia, generalized muscle weakness, CPK elevation and transaminitis, and this patient was admitted to intense care unit; patient 4 developed grade 3 diarrhea. All patients recovered from these abnormalities after either oral or intravenous steroids treatment. The development of a response was accompanied by AEs, and this might be attributed to the disorder of immune homeostasis such as the TCR diversity change, which caused systemic influences. Despite these irAEs, no disease progression was observed for more than 14 weeks in responding patients. The main efficacy was confirmed to be derived from the blockade of PD-L1 signaling, and following the biopsy of two patients after treatment, the authors observed a replacement of thymocytes by activated CD8+ T cells. Additionally, investigators found that patients who responded to avelumab had been treated with the multikinase inhibitor sunitinib. All patients with irAEs had been treated with sunitinib, and two-thirds of patients who did not develop irAEs had never been treated with sunitinib. Sunitinib is an effective targeted drug in TET treatment [73], and it has also been confirmed as an immunomodulator that decreases the populations of Tregs and myeloid-derived suppressor cells [74, 75], possibly leading to the activation of autoimmunity. These findings suggest that sunitinib and other similar kinase inhibitors may alter the efficacy of ICIs and increase the risk of irAEs, but more evidence is needed.
Nivolumab
Not all clinical trials acquired promising efficacy, a PRIMER single-arm, multicenter, phase II trial of 15 patients with unresectable or recurrent TC [64] reported that although treatment with nivolumab achieved a disease control rate of 73% (11/15), no significant tumor shrinkage was observed. Researchers suggested that further development or clinical trials of nivolumab were not recommended in these patients. Nevertheless, the results of this clinical trial were questionable since the sample size of this cohort was small, all patients with TC were from Japan, and the evaluation time was only 12 weeks. Therefore, clinical trials with larger sample sizes and longer evaluation times are needed to obtain more evidence for the efficacy of nivolumab in TET treatment.
Ongoing clinical trials
More clinical trials with larger sample sizes are being conducted in patients with TETs (Table 3). Radiotherapy and/or chemotherapy may affect the in situ immune status and exert ectopic effects, which may improve the immunotherapy response and reduce adverse effects [76]. And the benefits of immunotherapy combined with radiotherapy or chemotherapy have been explored in many tumors, such as breast cancer, melanoma, and small cell lung cancer [77,78,79]. However, there is lack of such evidence in TET treatment. Some studies have documented the potential benefit of combined therapy, and several clinical trials are in progress. For instance, Yuki Katsuya et al. [51] observed a significant increase of PD-1/PD-L1 expression in 30 patients after chemotherapy, suggesting that chemotherapy combined with anti-PD-1/PD-L1 therapy or sequential therapy may provide greater therapeutic benefits. Different immune checkpoints have different functional mechanisms, and combined immunotherapy may produce more overall effects. Clinical trials simultaneously targeting PD-L1 and CTLA-4 have been conducted in patients with TETs. However, this combination may also lead to an increased frequency and severity of irAEs, as reported in clinical trials of anti-PD-1 (nivolumab) and anti-CTLA-4 (ipilimumab) combination therapy for melanoma [80]. In addition, clinical trials combining ICIs and specific targeted drugs such as epidermal growth factor receptor inhibitors, have also been registered. Notably, even for the combination of regimens with completely different functional mechanisms, the therapy-related adverse events may be more severe. For example, the combination of durvalumab and gefitinib results in high-grade liver enzyme elevations in 40–70% of patients, which is higher than that reported with either drug alone [81, 82]. These ongoing clinical trials will provide more information about the monotherapy or combination therapy using ICIs and facilitate the development of novel therapeutic strategies for the systemic treatment of patients with TETs.
IrAEs of ICI therapy in patients with TETs
IrAEs reported in clinical trials and practice
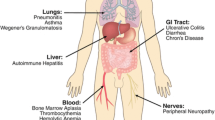
All clinical trials and most case reports in TETs immunotherapy reported irAEs of varying severity. They are very diverse and affect almost all organ systems (Fig. 4). Notably, irAEs seem more common in patients with thymoma. Cho et al. [62] reported irAEs in 15.4% of patients with TC compared with 71.6% of patients with thymoma. They suggested that immune checkpoint inhibitors should be avoided in patients with thymomas, and in patients with thymic carcinoma, immunotherapy should be considered with careful monitoring. All irAEs were graded according to the Common Terminology Criteria for Adverse Events, version 5.0 [83], listing them of different grades that occurred in the four completed clinical trials (Table 4). Patients with TETs usually have ADs or are in a preautoimmune state, and the irAEs caused by ICIs seem to be significantly higher than those in patients with other tumors [84, 85]. Although most patients only experienced mild irAEs (grade 1–2), a unique pattern of grade 3–4 irAEs was observed, including myocarditis, myositis and severe muscle weakness, which is rarely observed in patients with other tumors [61, 63, 85]. Remarkably, the development of irAEs appears to be associated with better therapeutic efficacy. Giaccone et al. [61] reported that four of nine patients (44.4%) who developed severe irAEs achieved partial responses, much higher than those without developing irAEs. Rajan et al. [63] also found that all responders developed irAEs, while only one of the four patients without response developed irAEs. Similar results have also been reported in patients with melanoma and NSCLC, with significantly higher response rates, PFS and OS in patients with irAEs than in those without irAEs [86,87,88]. In addition, some irAEs may occur several weeks after treatment, but the cumulative incidence of irAEs does not appear to increase with long-term follow-up [65], suggesting that a proportion of patients with TC are not at high risk of immune-mediated toxicity. Although irAE-related deaths are rare, further explorations of the mechanisms and more evidence that guide the identification of patients who might benefit from ICIs without developing severe irAEs are urgently needed [89].

An overview of irAEs in patients with TETs receiving PD-1/PD-L1 inhibitors. Immunotherapy is systemically administered to affect not only tumor progression but also the whole immune system, including central tolerance in the thymus and peripheral immune homeostasis. irAEs: Immune-related adverse events; TETs: Thymic epithelial tumors
Molecular mechanisms of immunotherapy and irAEs
Although tumor immunotherapy is well-studied, little is known about the mechanisms of irAEs. Accumulating evidence indicates that some irAEs at least represent a decrease in self-tolerance mediated by abnormalities in T cells development, genetic susceptibility, B cells and other possible mechanisms (Fig. 5) [90]. Recently, Chen et al. [91] appealed more mechanistic studies on ICI resistance rather than performing additional clinical trials with combinations of different treatment schemes. Similarly, explorations aiming to elucidate the mechanisms of irAEs during ICI therapy for TETs should be strengthened before more clinical trials are conducted.

Some key issues requiring further mechanistic exploration. Comprehensive studies based on these issues will help researchers develop new biomarkers to prevent, monitor and manage irAEs during ICI therapy in patients with TETs. Autoimmune T cells and a disturbed TCR repertoire seem most relevant to irAEs, and a preclinical mouse model of irAEs is urgently needed to provide an ideal platform for mechanistic studies of irAEs. irAEs: Immune-related adverse events; ICIs: Immune checkpoint inhibitors; TETs: Thymic epithelial tumors
Disturbed T cell development mediated by PD1/PD-L1 blockade
Systemic administration of ICIs inevitably affects T cells residing in normal thymic tissues, not only affecting central immune tolerance but also disrupting the homeostasis of the peripheral T cell receptor repertoire, leading to the production of autoreactive T cells. Firsly, thymic stromal cells and epithelial cells were confirmed to express PD-L1, especially in lymphocyte-rich thymoma, and the PD-1/PD-L1 interaction regulated both positive and negative selection of T cells in the thymus [47, 48]. Secondly, recent thymic emigrants in the peripheral blood are continuously activated via TCR signaling after encountering either autoantigen or alloantigen. And then, these activated naïve T cells will gradually express high-level PD-1 until they become exhausted T cells (Fig. 6 A). Under physiological conditions, normal function of PD-1/PD-L1 help establish immunologic homeostasis and protect normal tissues from attacked by exhausted T cells. However, blockade of PD-1/PD-L1 signaling may lead to disinhibition of effector T cells that induce thymic epithelial cell apoptosis and overcome either central immune tolerance to TSAs expressed by the thymic epithelium [92] or peripheral immune tolerance to circulating antigens. Moreover, PD-1 affects the CD8+ T cell status through intrinsic mechanisms such as functional inactivation or developmental regulation and promotes the differentiation of CD4+ T cells into Tregs [93]. In addition, the prognostic impact of the density and spatial architecture of tumor infiltrating lymphocytes was explored in TETs [94], and researchers demonstrated that high infiltration of stromal T helper and cytotoxic lymphocytes played a crucial role in anti-tumor immunity and might be potential marker predicting the efficacy of immunotherapy in TETs. The link between ADs and PD-1 has also been confirmed in a PD-1-deficient mouse model [95] that develops glomerulonephritis, inflammatory arthritis and lupus-like disease. In summary, blockade of PD-1/PD-L1 may impair their regulatory roles in both central and peripheral immune tolerance, thus leading to immune-related toxicities.

Cellular and molecular mechanisms of immune imbalance in TETs immunotherapy. A. PD-1/PD-L1 interaction in the thymic tissue regulates T cell development, and antigen-activated T cells upregulate PD-1 to avoid overactivation mediated tissue attack. B. Activated T cells release IFN-γ which upregulate PD-L1 expression in tumor cells via ERK/JAK2-STAT signaling pathways. Additionally, some circRNAs expressed by tumor cells may be involved in regulation of immune imbalance in TETs. C. Some lncRNAs, LncXLOC_003810, for example, can inhibit PD-1/PD-L1signaling and may be involved in regulating T cell differentiation. PD-1/PD-L1 signaling can inhibit PI3K/AKT pathway, which, however, can be activated by some immune related miRNAs. D. The immune microenvironment in TETs is distorted. PD-1/PD-L1 and their directly or indirectly interacting molecules play a vital role in maintaining immune homeostasis either inside or outside the thymus. However, this balance is disrupted in TETs especially after immunotherapy, which make TETs susceptible to irAEs during immunotherapy. TCR: T cell receptor; SHP2: Src homology 2 domain-containing tyrosine phosphatase 2; irAEs: Immune-related adverse events; ICIs: Immune checkpoint inhibitors; TETs: Thymic epithelial tumors
Gene alterations and TCR diversity changes
Different haplotypes and polymorphisms of human leukocyte antigen in immune regulatory genes (such as CTLA-4 and PD-1) are associated with multiple ADs, and may play an important role in the development of irAEs [96,97,98]. Through gene sequencing analysis, researchers found enrichment of specific gene mutations in patients who developed irAEs compared to those without irAEs [99]. And significantly higher IFN-γ expression in T cells from patients who responded to pembrolizumab than in nonresponders was observed [100]. Notably, autoreactive T cells recognize autoantigens expressed by TET cells and release IFN-γ, and IFN-γ can upregulate PD-L1 expression via ERK/JAK2-STAT signaling pathways in tumor cells, which may be one of the reasons why patients with irAEs respond better to ICIs [101] (Fig. 6 A, B). These findings suggest that specific gene mutations may contribute to the development of irAEs and that TET mutational patterns should be considered when evaluating the risks of developing irAEs in patients receiving immunotherapy. Additionally, through TCR sequencing, Rajan et al. [63] found a trend in patients with irAEs toward a higher level of TCR diversity prior to therapy that decreased after treatment with steroids. The patient with the highest level of TCR diversity experienced the most severe irAEs. Läubli et al. [102] observed T cell infiltration in the irAE-related foci of patients treated with a PD-1 inhibitor, with characteristics similar to those of tumor-infiltrating T cells. TCR expansion may be a pharmacodynamic effect of ICIs, which reflects overall immune activation. However, the mobilization of abundant T cells and the increase in clonal diversity may lead to the production of autoreactive T cells and antibodies, increasing the risk of irAEs [103].
B cells and antibodies
In NSCLC and renal cell carcinoma, researchers observed decreased numbers of circulating B cells and increased numbers of CD21low B cells and plasmablasts after a course of ICI therapy. CD21low B cells express higher levels of PD-1, and B cell receptor sequencing revealed greater clonality and a higher frequency of clones than CD21high cells. Compared with patients with no B cell changes, patients with a 30% or greater reduction in the number of total circulating B cells and a two-fold or greater increase in the numbers of CD21low B cells or plasmablasts were more sensitive to grade 3–4 irAEs [104,105,106]. Similar research in TET immunotherapy is also worth conducting because early changes in circulating B cells following immunotherapy may be biomarkers to identify patients who are at high risk of irAEs, and preemptive strategies targeting B cells may reduce toxicities in these patients. The analysis of baseline serum antibody profiles can also provide some insights into the mechanism of irAEs. Gowen et al. [107] identified a panel of specific antibodies that were differentially expressed in patients with severe irAEs. Autoantibodies against thyroid antigens or islet cell antigens were also detected in patients with thyroid dysfunction or diabetes after ICI treatment, but serological tests in patients with rheumatoid arthritis were usually negative [108,109,110].
Immune-related epigenetic alterations in TETs
Large scale RNA sequencing data helps deeper understand the molecular mechanisms of TET development and their relationship with autoimmune diseases, which also provides insights to explore molecular mechanisms of TET immunotherapy and irAEs. Researchers identified some immune-related microRNAs (miRNAs) that are correlated with immune cell infiltration and type 2 macrophage polarization, some of which can regulate IFN-γor IL-10 signaling pathway to influence TETs microenvironment and PD-L1 expression [111]. They may also be involved in the pathogenesis of irAEs. For example, the downregulation of miRNA-146a was reported to be associated with an increased risk of irAEs [112]. In addition, miR-34 is confirmed to be highly expressed in early-stage thymomas but virtually absent in TC [50]. In NSCLC, P53 can downregulate PD-L1 via miR-34 [113], coincidently, TCs express PD-L1 more frequently than thymoma, and this may be caused by a decrease of miR-34. Except for non-clustered miRNAs, miRNA clusters-C19MC was also differentially expressed in thymoma and TC. In thymoma, miRNAs of C19MC are strongly expressed and can activate PI3K/AKT signaling pathway [114], which however, is inhibited by PD-1/PD-L1 axis [115], regulating TETs development and immune microenvironment (Fig. 6 B, C). Some other miRNAs were also reported to activate PI3K-AKT, FoxO, HIF-1 and Rap-1signaling pathways [116], indicating a synergistic role combining pathway inhibitors and immunotherapy, which will reduce the risk of irAEs. Considering the autoimmune correlation of thymoma rather than TC and the differential expression pattern of some miRNAs, miRNAs may be involved in autoimmunity related pathways in patients with thymoma. Although these miRNAs are absent in TC and patients with TC seldom suffer autoimmunity, the administration of ICIs can also activate autoimmunity related pathways. This may partially explain why patients with TC also appear irAEs after immunotherapy.
Moreover, researchers reported some differentially expressed circRNAs such as circ_0001173 and circ_0007291 in normal thymic tissue and TETs. Their upregulation in TETs was confirmed to be positively correlated with MAPK and TNF signaling, which mediate immune disorder in TETs and influence efficacy of immunotherapy [117]. Also, LncRNA XLOC_003810 was reported to regulate Th17/Treg balance in TETs with myasthenia gravis, and high expression of LncRNA XLOC_003810 results in a decrease of Treg cells and immune regulating factors [118], making patients more susceptible to irAEs. Interestingly, another study reported that LncRNA XLOC_003810 can promote the activation of T cells and inhibit the expression of PD-1/PD-L1, resulting in the upregulation of proinflammatory cytokines and a decrease of proportion of CD4+PD1+/CD4+PD-L1+ monocytes [119], influencing immunotherapeutic efficacy and the risk of irAEs (Fig. 6 B, D). These findings indicate that noncoding RNAs are potential biomarkers to predict and monitor immunotherapeutic efficacy and the occurrence of irAEs.
Other potential mechanisms
Some cytokines, such as CXCL2/9/10 and IL-17, have been confirmed to be associated with irAEs induced by nivolumab in NSCLC or ipilimumab in melanoma. A higher level of IL-17 or a lower level of IL-6 correlates with a higher risk of developing irAEs [120,121,122,123]. Further investigations are also necessary to evaluate the roles of these cytokines in irAEs of TETs immunotherapy. The microbiota composition may be another indicator of irAEs in patients receiving ICI therapy. Routy et al. [124] showed that the effect of antibiotics on the gut microbiota is associated with adverse responses to PD-1 blockade, but the mechanism requires further investigation. In patients with melanoma, CTLA-4 inhibitor-induced irAEs were reported to be negatively associated with a high proportion of the Bacteroidetes phylum but positively associated with the Faecalibacterium genus and other Firmicutes species [99]. In patients with NSCLC, baseline enrichment of Bifidobacterium and Desulfovibrio in the gut microbiota was reported to be significantly associated with a lower incidence of irAEs during ICI therapy [125]. Similar research is needed for TETs to explore the relationship between the microbiota composition and the occurrence of irAEs.
Strategies for the prevention and management of irAEs
Although ICIs improve the survival of patients with unresectable cancers, inevitable off-target conditions lead to autoimmune events at nontumor sites. Certain strategies and interventions may help prevent and manage irAEs that occur during TET immunotherapy. Rigorous evaluation of the indications for immunotherapy, especially the immune status of patients is needed since patients with immunity changes caused by any factors, such as aging, long-term immunomodulators and chronic virus infection, may be prone to side effects [90, 126, 127]. For patients with active immune instability or a history of ADs, immunotherapy is not recommended due to a lack of evidence from clinical trials. The administration of immunomodulators or immunosuppressants, such as steroids, concurrently with ICIs may be useful. Rajan et al. [63] reported that TCR diversity decreased after treatment with steroids in patients who developed irAEs. However, researchers have drawn inconsistent conclusions regarding whether the use of steroids to prevent or treat irAEs alters the efficacy of ICIs [128,129,130]. Furthermore, severe steroid-related toxicity was reported, including fatal infections [131]. Therefore, more research is needed to explore the effect of immunosuppressants on the efficacy of ICIs and to find their ideal regimen and timing. In addition, the dose of ICIs for TET immunotherapy must be optimized to minimize the risks of irAEs while maintaining efficacy. Although a higher incidence of irAEs was observed in patients treated with higher doses of ICIs [132], some studies reported that a lower dose of pembrolizumab also achieved significant efficacy with acceptable toxicity in patients with NSCLC and TETs [133, 134]. Importantly, irAES are variable and unpredictable that occur at the initial or later stage of treatment and even several months after the final course. Some patients experience a single irAE, while others may develop a series of irAEs simultaneously [135, 136].
Thus, a more reasonable approach seems to be closely monitoring irAEs via specific biomarkers and implementing timely measures to prevent their occurrence or minimize their risk. Radiomics provides comprehensive visualization and characterization of tissues of interest and associated microenvironments by automatically extracting high-fidelity, high-dimensional medical imaging features from standard images and has been shown to predict clinical outcomes, including irAEs, especially pneumonia [137]. Early changes in B cells may identify patients at high risk of irAEs, and strategies targeting B cells are worthy of development to reduce toxicity in these patients [105]. Cytokines, which are involved in the occurrence of irAEs, also serve as both predictive biomarkers and intervention targets [120, 121]. When irAEs occur, a timely severity assessment and the necessary management, such as oral or intravenous steroids, are needed. Discontinuation of immunotherapy is required for severe irAEs such as myocarditis and neuromuscular complications, and sometimes a gamma globulin infusion or plasma exchange should be performed.
Readministration of ICIs in TETs
The readministration of ICIs after initial irAEs is challenging and controversial, which requires a careful assessment of the risk and potential clinical benefits. Evidence from case reports is insufficient [101, 102], and rigorous clinical research with a large sample size is needed, particularly to evaluate the ICI dose correlation with the occurrence of second-time irAEs. The antitumor activity and tolerability of readministering the same ICI at different doses was explored in other tumors, including NSCLC, renal cancer, and melanoma. Considering different immunological mechanisms of different ICIs, switching to another ICI after irAEs occur has also been evaluated, and readministration of ICIs appears feasible for low-grade irAEs [138,139,140,141,142,143,144]. A study examined the effect of a PD-1 inhibitor on patients with melanoma who had experienced ipilimumab (CTLA-4 inhibitor)-associated immune toxicity and found that most irAEs observed were new rather than recurrent [129]. Clinical trials in patients with NSCLC and melanoma have reported a dose correlation between the second-time irAEs with CTLA4 inhibitor, but not PD-1/PD-L1 inhibitor. And an adjustment of drug dose or course interval was not recommended for PD-1/PD-L1 inhibitor, but required for CTLA4 inhibitor after the initial irAEs [145, 146]. In addition, concurrent immunosuppressors or immunomodulators, such as vedolizumab (an integrin inhibitor) and TNF-α inhibitors, were administered to decrease the risk of re-emergence of irAEs, and a lower risk of second-time irAEs was confirmed than in patients who were administered ICIs alone [141, 147]. In patients with TETs, concurrent use of cyclosporine A with avelumab was also reported to prevent the development of second-time irAEs in patients with previous immune-mediated myositis [63]. These findings indicate that readministration of ICI with either the same/different doses or different types is feasible in carefully selected patients after balancing risks and benefits. However, for patients who have experienced life-threatening immunotoxicity, reintroduction of ICIs may not be reasonable, and further research is needed to accumulate more evidence.
Insights and future perspectives on ICIs in TETs
PD-1/PD-L1-based ICIs show promising prospects in TET treatment, but high-frequency irAEs pose a challenge. Several issues merit further study to maximize the therapeutic benefits while minimizing the risks of irAEs. First, new biomarkers are urgently needed to screen patients who might experience potential benefits. The main criterion for evaluating patients who might benefit from ICIs is PD-1/PD-L1 expression levels, but they are not perfect biomarkers [148] and do not reflect the propensity to develop irAEs. Second, clinical trials of patients with unstable immune status should be conducted. Patients with ADs are usually excluded from clinical trials of ICIs for tumors, which is particularly restrictive for patients with TETs [61,62,63], resulting in a serious lack of evidence for the guidance of immunotherapy. Clinical trials of ICIs in patients with melanoma and NSCLC presenting with ADs have reported mild irAEs and similar response rates to those without ADs, and irAEs usually do not lead to a discontinuation of ICIs [128]. Therefore, clinical trials of ICIs for patients with TETs complicated with ADs are also needed to accumulate more evidence for medication. Third, changes in some biomarkers related to irAEs are noted, which are presumed to be involved in the mechanisms of irAE occurrence and have the potential to serve as monitoring factors [90, 105, 121]. Nevertheless, much more research is needed before the clinical application of these biomarkers. A study combining radiotherapy and immunotherapy was conducted in mouse models of subcutaneous in situ thymomas to study the changes in cytokine levels in the tumor immune microenvironment and explore the effect of combined therapy [149]. However, no preclinical mouse models that mimic the autoimmune and toxic events observed in patients have been developed to understand the biological mechanisms of irAEs [150]. Fourth, standardization of the management of irAEs of different types and severity, assessments of the criteria for readministration of ICIs after initial irAEs, and recommendations for the time interval and corresponding precautions are needed. With an increasing number of clinicians trying to use individual ICIs, the standardized guiding framework will help to improve the efficacy of ICIs in patients with TETs and reduce the occurrence of severe irAEs. In addition, many researchers focus on irAEs, but mechanisms of nonresponse and resistance are also needed. Some potential mechanisms have been reported, such as defects in class I antigen presentation, defects in the Wnt/β-catenin and interferon signaling pathways [151], and overexpression of alternative immune checkpoints, such as T cell immunoglobulin and mucin domain-containing molecule-3 [152]. In all, more explorations of irAE mechanisms combined with evidence from clinical trials will increase the prospects of immunotherapy especially ICIs for patients with TETs.
In conclusion, although ICI therapy is at the early stage of TETs treatment, it has been proven the ability to induce durable response in a subset of patients with TETs. However, the risk of developing life-threatening irAEs hampers their adoption. An understanding of the mechanisms of irAEs, the identification of predictive biomarkers, and development of risk countermeasures to make immunotherapy a safe and effective regimen are urgently needed, which will improve survival and the quality of life of patients with TETs.
Availability of data and materials
Not applicable.
Abbreviations
- PD1:
-
Programmed death receptor 1
- PD-L1:
-
Programmed death receptor 1 ligand 1
- TETs:
-
Thymic epithelial tumors
- ICIs:
-
Immune checkpoint inhibitors
- irAEs:
-
Immune-related adverse events
- ADs:
-
Autoimmune diseases
- CTLA-4:
-
Cytotoxic T lymphocyte-associated protein 4
- NSCLC:
-
Non-small cell lung cancer
- OS:
-
Overall survival
- PFS:
-
Progression-free survival
References
Harnath T, Marx A, Ströbel P, Bölke E, Willers R, Gripp S. Thymoma-a clinico-pathological long-term study with emphasis on histology and adjuvant radiotherapy dose. J Thorac Oncol. 2012;7:1867–71.
Marx A, Chan JKC, Chalabreysse L, Dacic S, Detterbeck F, French CA, Hornick JL, Inagaki H, Jain D, Lazar AJ, et al. The 2021 WHO classification of tumors of the thymus and mediastinum: what is new in thymic epithelial, germ cell, and mesenchymal tumors? J Thorac Oncol. 2022;17:200–13.
Berardi R, De Lisa M, Pagliaretta S, Onofri A, Morgese F, Savini A, Ballatore Z, Caramanti M, Santoni M, Mazzanti P, Cascinu S. Thymic neoplasms: an update on the use of chemotherapy and new targeted therapies. A literature review Cancer Treat Rev. 2014;40:495–506.
Berghmans T, Durieux V, Holbrechts S, Jungels C, Lafitte JJ, Meert AP, Moretti L, Ocak S, Roelandts M, Girard N. Systemic treatments for thymoma and thymic carcinoma: a systematic review. Lung Cancer. 2018;126:25–31.
Merveilleux du Vignaux C, Maury JM, Girard N. Novel agents in the treatment of thymic malignancies. Curr Treat Options Oncol. 2017;18:52.
Bardhan K, Anagnostou T, Boussiotis VA. The PD1:PD-L1/2 pathway from discovery to clinical implementation. Front Immunol. 2016;7:550.
Zhao C, Rajan A. Immune checkpoint inhibitors for treatment of thymic epithelial tumors: how to maximize benefit and optimize risk? Mediastinum. 2019;3:35.
Santoni G, Amantini C, Morelli MB, Tomassoni D, Santoni M, Marinelli O, Nabissi M, Cardinali C, Paolucci V, Torniai M, et al. High CTLA-4 expression correlates with poor prognosis in thymoma patients. Oncotarget. 2018;9:16665–77.
Zheng K, Zhang J, Zhang P, Guo Y. PTPN22 and CTLA-4 gene polymorphisms in resected thymomas and thymus for myasthenia gravis. Thorac Cancer. 2012;3:307–12.
Berardi R, Goteri G, Brunelli A, Pagliaretta S, Paolucci V, Caramanti M, Rinaldi S, Refai M, Pompili C, Morgese F, et al. Prognostic relevance of programmed cell death protein 1/programmed death-ligand 1 pathway in thymic malignancies with combined immunohistochemical and biomolecular approach. Expert Opin Ther Targets. 2020;24:937–43.
Franzin R, Netti GS, Spadaccino F, Porta C, Gesualdo L, Stallone G, Castellano G, Ranieri E. The use of immune checkpoint inhibitors in oncology and the occurrence of AKI: where do we stand? Front Immunol. 2020;11: 574271.
Vikas P, Borcherding N, Zhang W. The clinical promise of immunotherapy in triple-negative breast cancer. Cancer Manag Res. 2018;10:6823–33.
Goodman A, Patel SP, Kurzrock R. PD-1-PD-L1 immune-checkpoint blockade in B-cell lymphomas. Nat Rev Clin Oncol. 2017;14:203–20.
NCCN Guidelines Version 1.2022 Thymomas and Thymic Carcinomas [https://www.nccn.org]
Ballman M, Zhao C, McAdams MJ, Rajan A. Immunotherapy for management of thymic epithelial tumors: a double-edged sword. Cancers (Basel). 2022;14(9):2060.
Velcheti V, Schalper K. Basic overview of current immunotherapy approaches in cancer. Am Soc Clin Oncol Educ Book. 2016;35:298–308.
Conlon KC, Miljkovic MD, Waldmann TA. Cytokines in the treatment of cancer. J Interferon Cytokine Res. 2019;39:6–21.
Oji Y, Inoue M, Takeda Y, Hosen N, Shintani Y, Kawakami M, Harada T, Murakami Y, Iwai M, Fukuda M, et al. WT1 peptide-based immunotherapy for advanced thymic epithelial malignancies. Int J Cancer. 2018;142:2375–82.
Zhang Y, Morgan R, Podack ER, Rosenblatt J. B cell regulation of anti-tumor immune response. Immunol Res. 2013;57:115–24.
Shaffer DR, Savoldo B, Yi Z, Chow KK, Kakarla S, Spencer DM, Dotti G, Wu MF, Liu H, Kenney S, Gottschalk S. T cells redirected against CD70 for the immunotherapy of CD70-positive malignancies. Blood. 2011;117:4304–14.
Hishima T, Fukayama M, Hayashi Y, Fujii T, Ooba T, Funata N, Koike M. CD70 expression in thymic carcinoma. Am J Surg Pathol. 2000;24:742–6.
Kashima J, Hishima T, Okuma Y, Horio H, Ogawa M, Hayashi Y, Horiguchi SI, Motoi T, Ushiku T, Fukayama M. CD70 in thymic squamous cell carcinoma: potential diagnostic markers and immunotherapeutic targets. Front Oncol. 2021;11: 808396.
Ao YQ, Jiang JH, Gao J, Wang HK, Ding JY. Recent thymic emigrants as the bridge between thymoma and autoimmune diseases. Biochim Biophys Acta Rev Cancer. 2022;1877: 188730.
Burbelo PD, Browne SK, Sampaio EP, Giaccone G, Zaman R, Kristosturyan E, Rajan A, Ding L, Ching KH, Berman A, et al. Anti-cytokine autoantibodies are associated with opportunistic infection in patients with thymic neoplasia. Blood. 2010;116:4848–58.
Girard N. Immune checkpoints in thymic epithelial tumors: challenges and opportunities. Immunooncol Technol. 2019;3:8–14.
Radovich M, Pickering CR, Felau I, Ha G, Zhang H, Jo H, Hoadley KA, Anur P, Zhang J, McLellan M, et al. The integrated genomic landscape of thymic epithelial tumors. Cancer Cell. 2018;33:244-258.e210.
Lenschow DJ, Walunas TL, Bluestone JA. CD28/B7 system of T cell costimulation. Annu Rev Immunol. 1996;14:233–58.
Farhood B, Najafi M, Mortezaee K. CD8(+) cytotoxic T lymphocytes in cancer immunotherapy: a review. J Cell Physiol. 2019;234:8509–21.
Twomey JD, Zhang B. Cancer immunotherapy update: FDA-approved checkpoint inhibitors and companion diagnostics. Aaps j. 2021;23:39.
Li X, Hu W, Zheng X, Zhang C, Du P, Zheng Z, Yang Y, Wu J, Ji M, Jiang J, Wu C. Emerging immune checkpoints for cancer therapy. Acta Oncol. 2015;54:1706–13.
Arasanz H, Gato-Cañas M, Zuazo M, Ibañez-Vea M, Breckpot K, Kochan G, Escors D. PD1 signal transduction pathways in T cells. Oncotarget. 2017;8:51936–45.
Yokosuka T, Takamatsu M, Kobayashi-Imanishi W, Hashimoto-Tane A, Azuma M, Saito T. Programmed cell death 1 forms negative costimulatory microclusters that directly inhibit T cell receptor signaling by recruiting phosphatase SHP2. J Exp Med. 2012;209:1201–17.
Patsoukis N, Bardhan K, Chatterjee P, Sari D, Liu B, Bell LN, Karoly ED, Freeman GJ, Petkova V, Seth P, et al. PD-1 alters T-cell metabolic reprogramming by inhibiting glycolysis and promoting lipolysis and fatty acid oxidation. Nat Commun. 2015;6:6692.
Robainas M, Otano R, Bueno S, Ait-Oudhia S. Understanding the role of PD-L1/PD1 pathway blockade and autophagy in cancer therapy. Onco Targets Ther. 2017;10:1803–7.
Weissferdt A, Fujimoto J, Kalhor N, Rodriguez J, Bassett R, Wistuba II, Moran CA. Expression of PD-1 and PD-L1 in thymic epithelial neoplasms. Mod Pathol. 2017;30:826–33.
Arbour KC, Naidoo J, Steele KE, Ni A, Moreira AL, Rekhtman N, Robbins PB, Karakunnel J, Rimner A, Huang J, et al. Expression of PD-L1 and other immunotherapeutic targets in thymic epithelial tumors. PLoS ONE. 2017;12: e0182665.
Suster D, Pihan G, Mackinnon AC, Suster S. Expression of PD-L1/PD-1 in lymphoepithelioma-like carcinoma of the thymus. Mod Pathol. 2018;31:1801–6.
Higuchi R, Goto T, Hirotsu Y, Nakagomi T, Yokoyama Y, Otake S, Amemiya K, Oyama T, Omata M. PD-L1 expression and tumor-infiltrating lymphocytes in thymic epithelial neoplasms. J Clin Med. 2019;8(11):1833.
Chen HF, Wu LX, Li XF, Zhu YC, Pan WW, Wang WX, Xu CW, Huang JH, Wu MH, Du KQ. PD-L1 expression level in different thymoma stages and thymic carcinoma: a meta-analysis. Tumori. 2020;106:306–11.
Doroshow DB, Bhalla S, Beasley MB, Sholl LM, Kerr KM, Gnjatic S, Wistuba II, Rimm DL, Tsao MS, Hirsch FR. PD-L1 as a biomarker of response to immune-checkpoint inhibitors. Nat Rev Clin Oncol. 2021;18:345–62.
Terra S, Mansfield AS, Vrana JA, Roden AC. Heterogeneity of programmed death-ligand 1 expression in thymic epithelial tumours between initial specimen and synchronous or metachronous metastases or recurrences. Histopathology. 2019;74:364–7.
He Y, Ramesh A, Gusev Y, Bhuvaneshwar K, Giaccone G. Molecular predictors of response to pembrolizumab in thymic carcinoma. Cell Rep Med. 2021;2: 100392.
Umemura S, Zhu J, Chahine JJ, Kallakury B, Chen V, Kim IK, Zhang YW, Goto K, He Y, Giaccone G. Downregulation of CYLD promotes IFN-γ mediated PD-L1 expression in thymic epithelial tumors. Lung Cancer. 2020;147:221–8.
Wei YF, Chu CY, Chang CC, Lin SH, Su WC, Tseng YL, Lin CC, Yen YT. Different pattern of PD-L1, IDO, and FOXP3 Tregs expression with survival in thymoma and thymic carcinoma. Lung Cancer. 2018;125:35–42.
Duan J, Liu X, Chen H, Sun Y, Liu Y, Bai H, Wang J. Impact of PD-L1, transforming growth factor-β expression and tumor-infiltrating CD8(+) T cells on clinical outcome of patients with advanced thymic epithelial tumors. Thorac Cancer. 2018;9:1341–53.
Funaki S, Shintani Y, Fukui E, Yamamoto Y, Kanzaki R, Ose N, Kanou T, Minami M, Mori E, Okumura M. The prognostic impact of programmed cell death 1 and its ligand and the correlation with epithelial-mesenchymal transition in thymic carcinoma. Cancer Med. 2019;8:216–26.
Katsuya Y, Fujita Y, Horinouchi H, Ohe Y, Watanabe S, Tsuta K. Immunohistochemical status of PD-L1 in thymoma and thymic carcinoma. Lung Cancer. 2015;88:154–9.
Padda SK, Riess JW, Schwartz EJ, Tian L, Kohrt HE, Neal JW, West RB, Wakelee HA. Diffuse high intensity PD-L1 staining in thymic epithelial tumors. J Thorac Oncol. 2015;10:500–8.
Marchevsky AM, Walts AE. PD-L1, PD-1, CD4, and CD8 expression in neoplastic and nonneoplastic thymus. Hum Pathol. 2017;60:16–23.
Enkner F, Pichlhöfer B, Zaharie AT, Krunic M, Holper TM, Janik S, Moser B, Schlangen K, Neudert B, Walter K, et al. Molecular profiling of thymoma and thymic carcinoma: genetic differences and potential novel therapeutic targets. Pathol Oncol Res. 2017;23:551–64.
Katsuya Y, Horinouchi H, Asao T, Kitahara S, Goto Y, Kanda S, Fujiwara Y, Nokihara H, Yamamoto N, Watanabe S, et al. Expression of programmed death 1 (PD-1) and its ligand (PD-L1) in thymic epithelial tumors: impact on treatment efficacy and alteration in expression after chemotherapy. Lung Cancer. 2016;99:4–10.
Yokoyama S, Miyoshi H, Nishi T, Hashiguchi T, Mitsuoka M, Takamori S, Akagi Y, Kakuma T, Ohshima K. Clinicopathologic and prognostic implications of programmed death ligand 1 expression in thymoma. Ann Thorac Surg. 2016;101:1361–9.
Tiseo M, Damato A, Longo L, Barbieri F, Bertolini F, Stefani A, Migaldi M, Gnetti L, Camisa R, Bordi P, et al. Analysis of a panel of druggable gene mutations and of ALK and PD-L1 expression in a series of thymic epithelial tumors (TETs). Lung Cancer. 2017;104:24–30.
Owen D, Chu B, Lehman AM, Annamalai L, Yearley JH, Shilo K, Otterson GA. Expression patterns, prognostic value, and intratumoral heterogeneity of PD-L1 and PD-1 in thymoma and thymic carcinoma. J Thorac Oncol. 2018;13:1204–12.
Hakiri S, Fukui T, Mori S, Kawaguchi K, Nakamura S, Ozeki N, Kato T, Goto M, Yatabe Y, Yokoi K. Clinicopathologic features of thymoma with the expression of programmed death ligand 1. Ann Thorac Surg. 2019;107:418–24.
Guleria P, Husain N, Shukla S, Kumar S, Parshad R, Jain D. PD-L1 immuno-expression assay in thymomas: study of 84 cases and review of literature. Ann Diagn Pathol. 2018;34:135–41.
Chen Y, Zhang Y, Chai X, Gao J, Chen G, Zhang W, Zhang Y. Correlation between the expression of PD-L1 and clinicopathological features in patients with thymic epithelial tumors. Biomed Res Int. 2018;2018:5830547.
Bagir EK, Acikalin A, Avci A, Gumurdulu D, Paydas S. PD-1 and PD-L1 expression in thymic epithelial tumours and non-neoplastic thymus. J Clin Pathol. 2018;71:637–41.
Rouquette I, Taranchon-Clermont E, Gilhodes J, Bluthgen MV, Perallon R, Chalabreysse L, De Muret A, Hofman V, Marx A, Parrens M, et al. Immune biomarkers in thymic epithelial tumors: expression patterns, prognostic value and comparison of diagnostic tests for PD-L1. Biomark Res. 2019;7:28.
Ishihara S, Okada S, Ogi H, Kodama Y, Shimomura M, Tsunezuka H, Itoh K, Marx A, Inoue M. Programmed death-ligand 1 expression profiling in thymic epithelial cell tumors: Clinicopathological features and quantitative digital image analyses. Lung Cancer. 2020;145:40–7.
Giaccone G, Kim C, Thompson J, McGuire C, Kallakury B, Chahine JJ, Manning M, Mogg R, Blumenschein WM, Tan MT, et al. Pembrolizumab in patients with thymic carcinoma: a single-arm, single-centre, phase 2 study. Lancet Oncol. 2018;19:347–55.
Cho J, Kim HS, Ku BM, Choi YL, Cristescu R, Han J, Sun JM, Lee SH, Ahn JS, Park K, Ahn MJ. Pembrolizumab for patients with refractory or relapsed thymic epithelial tumor: an open-label phase II trial. J Clin Oncol. 2019;37:2162–70.
Rajan A, Heery CR, Thomas A, Mammen AL, Perry S, O’Sullivan Coyne G, Guha U, Berman A, Szabo E, Madan RA, et al. Efficacy and tolerability of anti-programmed death-ligand 1 (PD-L1) antibody (Avelumab) treatment in advanced thymoma. J Immunother Cancer. 2019;7:269.
Katsuya Y, Horinouchi H, Seto T, Umemura S, Hosomi Y, Satouchi M, Nishio M, Kozuki T, Hida T, Sukigara T, et al. Single-arm, multicentre, phase II trial of nivolumab for unresectable or recurrent thymic carcinoma: PRIMER study. Eur J Cancer. 2019;113:78–86.
Giaccone G, Kim C. Durable response in patients with thymic carcinoma treated with pembrolizumab after prolonged follow-up. J Thorac Oncol. 2021;16:483–5.
Frigola J, Navarro A, Carbonell C, Callejo A, Iranzo P, Cedrés S, Martinez-Marti A, Pardo N, Saoudi-Gonzalez N, Martinez D, et al. Molecular profiling of long-term responders to immune checkpoint inhibitors in advanced non-small cell lung cancer. Mol Oncol. 2021;15:887–900.
Giroux Leprieur E, Hélias-Rodzewicz Z, Takam Kamga P, Costantini A, Julie C, Corjon A, Dumenil C, Dumoulin J, Giraud V, Labrune S, et al. Sequential ctDNA whole-exome sequencing in advanced lung adenocarcinoma with initial durable tumor response on immune checkpoint inhibitor and late progression. J Immunother Cancer. 2020;8(1):e000527.
Chumsri S, Sokol ES, Soyano-Muller AE, Parrondo RD, Reynolds GA, Nassar A, Thompson EA. Durable complete response with immune checkpoint inhibitor in breast cancer with high tumor mutational burden and APOBEC signature. J Natl Compr Canc Netw. 2020;18:517–21.
Lee L, Ali S, Genega E, Reed D, Sokol E, Mathew P. Aggressive-variant microsatellite-stable POLE mutant prostate cancer with high mutation burden and durable response to immune checkpoint inhibitor therapy. JCO Precis Oncol. 2018;2:1–8.
Radovich M, Solzak JP, Hancock BA, Conces ML, Atale R, Porter RF, Zhu J, Glasscock J, Kesler KA, Badve SS, et al. A large microRNA cluster on chromosome 19 is a transcriptional hallmark of WHO type A and AB thymomas. Br J Cancer. 2016;114:477–84.
Inoue M, Marx A, Zettl A, Ströbel P, Müller-Hermelink HK, Starostik P. Chromosome 6 suffers frequent and multiple aberrations in thymoma. Am J Pathol. 2002;161:1507–13.
Repetto M, Conforti F, Pirola S, Calvello M, Pala L, Bonanni B, Catania C, Curigliano G, De Pas T. Thymic carcinoma with Lynch syndrome or microsatellite instability, a rare entity responsive to immunotherapy. Eur J Cancer. 2021;153:162–7.
Thomas A, Rajan A, Berman A, Tomita Y, Brzezniak C, Lee MJ, Lee S, Ling A, Spittler AJ, Carter CA, et al. Sunitinib in patients with chemotherapy-refractory thymoma and thymic carcinoma: an open-label phase 2 trial. Lancet Oncol. 2015;16:177–86.
Ozao-Choy J, Ma G, Kao J, Wang GX, Meseck M, Sung M, Schwartz M, Divino CM, Pan PY, Chen SH. The novel role of tyrosine kinase inhibitor in the reversal of immune suppression and modulation of tumor microenvironment for immune-based cancer therapies. Cancer Res. 2009;69:2514–22.
Finke JH, Rini B, Ireland J, Rayman P, Richmond A, Golshayan A, Wood L, Elson P, Garcia J, Dreicer R, Bukowski R. Sunitinib reverses type-1 immune suppression and decreases T-regulatory cells in renal cell carcinoma patients. Clin Cancer Res. 2008;14:6674–82.
Bracci L, Schiavoni G, Sistigu A, Belardelli F. Immune-based mechanisms of cytotoxic chemotherapy: implications for the design of novel and rationale-based combined treatments against cancer. Cell Death Differ. 2014;21:15–25.
Yu WD, Sun G, Li J, Xu J, Wang X. Mechanisms and therapeutic potentials of cancer immunotherapy in combination with radiotherapy and/or chemotherapy. Cancer Lett. 2019;452:66–70.
Schmid P, Adams S, Rugo HS, Schneeweiss A, Barrios CH, Iwata H, Diéras V, Hegg R, Im SA, Shaw Wright G, et al. Atezolizumab and nab-paclitaxel in advanced triple-negative breast cancer. N Engl J Med. 2018;379:2108–21.
Horn L, Mansfield AS, Szczęsna A, Havel L, Krzakowski M, Hochmair MJ, Huemer F, Losonczy G, Johnson ML, Nishio M, et al. First-line atezolizumab plus chemotherapy in extensive-stage small-cell lung cancer. N Engl J Med. 2018;379:2220–9.
Wolchok JD, Chiarion-Sileni V, Gonzalez R, Rutkowski P, Grob JJ, Cowey CL, Lao CD, Wagstaff J, Schadendorf D, Ferrucci PF, et al. Overall survival with combined nivolumab and ipilimumab in advanced melanoma. N Engl J Med. 2017;377:1345–56.
Ahn MJ, Sun JM, Lee SH, Ahn JS, Park K. EGFR TKI combination with immunotherapy in non-small cell lung cancer. Expert Opin Drug Saf. 2017;16:465–9.
Oxnard GR, Yang JC, Yu H, Kim SW, Saka H, Horn L, Goto K, Ohe Y, Mann H, Thress KS, et al. TATTON: a multi-arm, phase Ib trial of osimertinib combined with selumetinib, savolitinib, or durvalumab in EGFR-mutant lung cancer. Ann Oncol. 2020;31:507–16.
Yu Y, Ruddy KJ, Tsuji S, Hong N, Liu H, Shah N, Jiang G. Coverage evaluation of CTCAE for capturing the immune-related adverse events leveraging text mining technologies. AMIA Jt Summits Transl Sci Proc. 2019;2019:771–8.
Abdel-Wahab N, Shah M, Lopez-Olivo MA, Suarez-Almazor ME. Use of immune checkpoint inhibitors in the treatment of patients with cancer and preexisting autoimmune disease: a systematic review. Ann Intern Med. 2018;168:121–30.
Mammen AL, Rajan A, Pak K, Lehky T, Casciola-Rosen L, Donahue RN, Lepone LM, Zekeridou A, Pittock SJ, Hassan R, et al. Pre-existing antiacetylcholine receptor autoantibodies and B cell lymphopaenia are associated with the development of myositis in patients with thymoma treated with avelumab, an immune checkpoint inhibitor targeting programmed death-ligand 1. Ann Rheum Dis. 2019;78:150–2.
Hua C, Boussemart L, Mateus C, Routier E, Boutros C, Cazenave H, Viollet R, Thomas M, Roy S, Benannoune N, et al. Association of vitiligo with tumor response in patients with metastatic melanoma treated with pembrolizumab. JAMA Dermatol. 2016;152:45–51.
Toi Y, Sugawara S, Kawashima Y, Aiba T, Kawana S, Saito R, Tsurumi K, Suzuki K, Shimizu H, Sugisaka J, et al. Association of immune-related adverse events with clinical benefit in patients with advanced non-small-cell lung cancer treated with nivolumab. Oncologist. 2018;23:1358–65.
Fujimura T, Sato Y, Tanita K, Kambayashi Y, Otsuka A, Fujisawa Y, Yoshino K, Matsushita S, Funakoshi T, Hata H, et al. Serum levels of soluble CD163 and CXCL5 may be predictive markers for immune-related adverse events in patients with advanced melanoma treated with nivolumab: a pilot study. Oncotarget. 2018;9:15542–51.
Rajan A. Immunotherapy for thymic cancers: a convoluted path toward a cherished goal. J Thorac Oncol. 2021;16:352–4.
Khan S, Gerber DE. Autoimmunity, checkpoint inhibitor therapy and immune-related adverse events: a review. Semin Cancer Biol. 2020;64:93–101.
Kim TK, Vandsemb EN, Herbst RS, Chen L. Adaptive immune resistance at the tumour site: mechanisms and therapeutic opportunities. Nat Rev Drug Discov. 2022;21:529–40.
Anderson G, Takahama Y. Thymic epithelial cells: working class heroes for T cell development and repertoire selection. Trends Immunol. 2012;33:256–63.
Sckisel GD, Mirsoian A, Minnar CM, Crittenden M, Curti B, Chen JQ, Blazar BR, Borowsky AD, Monjazeb AM, Murphy WJ. Differential phenotypes of memory CD4 and CD8 T cells in the spleen and peripheral tissues following immunostimulatory therapy. J Immunother Cancer. 2017;5:33.
Bocchialini G, Schiefer AI, Müllauer L, Thanner J, Bauer J, Thaler F, Laggner M, Veraar C, Klepetko W, Hötzenecker K, et al. Tumour immune microenvironment in resected thymic carcinomas as a predictor of clinical outcome. Br J Cancer. 2022;127:1162–71.
Nishimura H, Nose M, Hiai H, Minato N, Honjo T. Development of lupus-like autoimmune diseases by disruption of the PD-1 gene encoding an ITIM motif-carrying immunoreceptor. Immunity. 1999;11:141–51.
Gough SC, Walker LS, Sansom DM. CTLA4 gene polymorphism and autoimmunity. Immunol Rev. 2005;204:102–15.
Lee J, Phong B, Egloff AM, Kane LP. TIM polymorphisms–genetics and function. Genes Immun. 2011;12:595–604.
Sharpe AH, Wherry EJ, Ahmed R, Freeman GJ. The function of programmed cell death 1 and its ligands in regulating autoimmunity and infection. Nat Immunol. 2007;8:239–45.
Wells K, Amato C, Hintzsche J, Robinson W. Identification of somatic mutations and neoantigens to predict development of autoimmune adverse events to immune therapy in melanoma. J Clin Oncol. 2017;35:19–19.
Ayers M, Lunceford J, Nebozhyn M, Murphy E, Loboda A, Kaufman DR, Albright A, Cheng JD, Kang SP, Shankaran V, et al. IFN-γ-related mRNA profile predicts clinical response to PD-1 blockade. J Clin Invest. 2017;127:2930–40.
Kim TK, Herbst RS, Chen L. Defining and understanding adaptive resistance in cancer immunotherapy. Trends Immunol. 2018;39:624–31.
Läubli H, Koelzer VH, Matter MS, Herzig P, Dolder Schlienger B, Wiese MN, Lardinois D, Mertz KD, Zippelius A. The T cell repertoire in tumors overlaps with pulmonary inflammatory lesions in patients treated with checkpoint inhibitors. Oncoimmunology. 2018;7: e1386362.
Oh DY, Cham J, Zhang L, Fong G, Kwek SS, Klinger M, Faham M, Fong L. Immune toxicities elicted by CTLA-4 blockade in cancer patients are associated with early diversification of the T-cell repertoire. Cancer Res. 2017;77:1322–30.
Nishimura K, Konishi T, Ochi T, Watanabe R, Noda T, Fukumoto T, Miura N, Miyauchi Y, Kikugawa T, Takenaka K, Saika T. CD21(lo) B cells could be a potential predictor of immune-related adverse events in renal cell carcinoma. J Pers Med. 2022;12(6):888.
Das R, Bar N, Ferreira M, Newman AM, Zhang L, Bailur JK, Bacchiocchi A, Kluger H, Wei W, Halaban R, et al. Early B cell changes predict autoimmunity following combination immune checkpoint blockade. J Clin Invest. 2018;128:715–20.
Liudahl SM, Coussens LM. B cells as biomarkers: predicting immune checkpoint therapy adverse events. J Clin Invest. 2018;128:577–9.
Gowen MF, Giles KM, Simpson D, Tchack J, Zhou H, Moran U, Dawood Z, Pavlick AC, Hu S, Wilson MA, et al. Baseline antibody profiles predict toxicity in melanoma patients treated with immune checkpoint inhibitors. J Transl Med. 2018;16:82.
Osorio JC, Ni A, Chaft JE, Pollina R, Kasler MK, Stephens D, Rodriguez C, Cambridge L, Rizvi H, Wolchok JD, et al. Antibody-mediated thyroid dysfunction during T-cell checkpoint blockade in patients with non-small-cell lung cancer. Ann Oncol. 2017;28:583–9.
Stamatouli AM, Quandt Z, Perdigoto AL, Clark PL, Kluger H, Weiss SA, Gettinger S, Sznol M, Young A, Rushakoff R, et al. Collateral damage: insulin-dependent diabetes induced with checkpoint inhibitors. Diabetes. 2018;67:1471–80.
Kostine M, Rouxel L, Barnetche T, Veillon R, Martin F, Dutriaux C, Dousset L, Pham-Ledard A, Prey S, Beylot-Barry M, et al. Rheumatic disorders associated with immune checkpoint inhibitors in patients with cancer-clinical aspects and relationship with tumour response: a single-centre prospective cohort study. Ann Rheum Dis. 2018;77:393–8.
Wang B, Xiao H, Yang X, Zeng Y, Zhang Z, Yang R, Chen H, Chen C, Chen J. A novel immune-related microRNA signature for prognosis of thymoma. Aging (Albany NY). 2022;14:4739–54.
Marschner D, Falk M, Javorniczky NR, Hanke-Müller K, Rawluk J, Schmitt-Graeff A, Simonetta F, Haring E, Dicks S, Ku M, et al. MicroRNA-146a regulates immune-related adverse events caused by immune checkpoint inhibitors. JCI Insight. 2020;5(6):e132334.
Cortez MA, Ivan C, Valdecanas D, Wang X, Peltier HJ, Ye Y, Araujo L, Carbone DP, Shilo K, Giri DK, et al: PDL1 Regulation by p53 via miR-34. J Natl Cancer Inst 2016, 108.
Oberndorfer F, Müllauer L. Genomic alterations in thymoma-molecular pathogenesis? J Thorac Dis. 2020;12:7536–44.
Gaikwad S, Agrawal MY, Kaushik I, Ramachandran S, Srivastava SK. Immune checkpoint proteins: signaling mechanisms and molecular interactions in cancer immunotherapy. Semin Cancer Biol. 2022;86:137–50.
Ji G, Ren R, Fang X. Identification and characterization of non-coding RNAs in thymoma. Med Sci Monit. 2021;27: e929727.
Wu Q, Luo X, Li H, Zhang L, Su F, Hou S, Yin J, Zhang W, Zou L. Identification of differentially expressed circular RNAs associated with thymoma. Thorac Cancer. 2021;12:1312–9.
Niu L, Jiang J, Yin Y, Hu B. LncRNA XLOC_003810 modulates thymic Th17/Treg balance in myasthenia gravis with thymoma. Clin Exp Pharmacol Physiol. 2020;47:989–96.
Hu B, Niu L, Jiang Z, Xu S, Hu Y, Cao K. LncRNA XLOC_003810 promotes T cell activation and inhibits PD-1/PD-L1 expression in patients with myasthenia gravis-related thymoma. Scand J Immunol. 2020;92: e12886.
Suh KJ, Kim SH, Kim YJ, Kim M, Keam B, Kim TM, Kim DW, Heo DS, Lee JS. Post-treatment neutrophil-to-lymphocyte ratio at week 6 is prognostic in patients with advanced non-small cell lung cancers treated with anti-PD-1 antibody. Cancer Immunol Immunother. 2018;67:459–70.
Khan S, Khan SA, Luo X, Fattah FJ, Saltarski J, Gloria-McCutchen Y, Lu R, Xie Y, Li Q, Wakeland E, Gerber DE. Immune dysregulation in cancer patients developing immune-related adverse events. Br J Cancer. 2019;120:63–8.
Tarhini AA, Zahoor H, Lin Y, Malhotra U, Sander C, Butterfield LH, Kirkwood JM. Baseline circulating IL-17 predicts toxicity while TGF-β1 and IL-10 are prognostic of relapse in ipilimumab neoadjuvant therapy of melanoma. J Immunother Cancer. 2015;3:39.
Valpione S, Pasquali S, Campana LG, Piccin L, Mocellin S, Pigozzo J, Chiarion-Sileni V. Sex and interleukin-6 are prognostic factors for autoimmune toxicity following treatment with anti-CTLA4 blockade. J Transl Med. 2018;16:94.
Routy B, Le Chatelier E, Derosa L, Duong CPM, Alou MT, Daillère R, Fluckiger A, Messaoudene M, Rauber C, Roberti MP, et al. Gut microbiome influences efficacy of PD-1-based immunotherapy against epithelial tumors. Science. 2018;359:91–7.
Chau J, Yadav M, Liu B, Furqan M, Dai Q, Shahi S, Gupta A, Mercer KN, Eastman E, Hejleh TA, et al. Prospective correlation between the patient microbiome with response to and development of immune-mediated adverse effects to immunotherapy in lung cancer. BMC Cancer. 2021;21:808.
Bouchlaka MN, Sckisel GD, Chen M, Mirsoian A, Zamora AE, Maverakis E, Wilkins DE, Alderson KL, Hsiao HH, Weiss JM, et al. Aging predisposes to acute inflammatory induced pathology after tumor immunotherapy. J Exp Med. 2013;210:2223–37.
Kennedy LB, Salama AKS. A review of cancer immunotherapy toxicity. CA Cancer J Clin. 2020;70:86–104.
Haanen J, Ernstoff MS, Wang Y, Menzies AM, Puzanov I, Grivas P, Larkin J, Peters S, Thompson JA, Obeid M. Autoimmune diseases and immune-checkpoint inhibitors for cancer therapy: review of the literature and personalized risk-based prevention strategy. Ann Oncol. 2020;31:724–44.
Menzies AM, Johnson DB, Ramanujam S, Atkinson VG, Wong ANM, Park JJ, McQuade JL, Shoushtari AN, Tsai KK, Eroglu Z, et al. Anti-PD-1 therapy in patients with advanced melanoma and preexisting autoimmune disorders or major toxicity with ipilimumab. Ann Oncol. 2017;28:368–76.
Postow MA, Sidlow R, Hellmann MD. Immune-related adverse events associated with immune checkpoint blockade. N Engl J Med. 2018;378:158–68.
Naidoo J, Wang X, Woo KM, Iyriboz T, Halpenny D, Cunningham J, Chaft JE, Segal NH, Callahan MK, Lesokhin AM, et al. Pneumonitis in patients treated with anti-programmed death-1/programmed death ligand 1 therapy. J Clin Oncol. 2017;35:709–17.
Haanen J, Carbonnel F, Robert C, Kerr KM, Peters S, Larkin J, Jordan K. Management of toxicities from immunotherapy: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann Oncol. 2017;28:iv119–42.
Low JL, Huang Y, Sooi K, Ang Y, Chan ZY, Spencer K, Jeyasekharan AD, Sundar R, Goh BC, Soo R, Yong WP. Low-dose pembrolizumab in the treatment of advanced non-small cell lung cancer. Int J Cancer. 2021;149:169–76.
Shen L, Chen H, Wei Q. Immune-therapy-related toxicity events and dramatic remission after a single dose of pembrolizumab treatment in metastatic thymoma: a case report. Front Immunol. 2021;12: 621858.
Michot JM, Bigenwald C, Champiat S, Collins M, Carbonnel F, Postel-Vinay S, Berdelou A, Varga A, Bahleda R, Hollebecque A, et al. Immune-related adverse events with immune checkpoint blockade: a comprehensive review. Eur J Cancer. 2016;54:139–48.
Mandalà M, Merelli B, Indriolo A, Tondini C. Late-occurring toxicity induced by an immune checkpoint blockade in adjuvant treatment of a stage III melanoma patient. Eur J Cancer. 2018;95:130–2.
Sun R, Limkin EJ, Vakalopoulou M, Dercle L, Champiat S, Han SR, Verlingue L, Brandao D, Lancia A, Ammari S, et al. A radiomics approach to assess tumour-infiltrating CD8 cells and response to anti-PD-1 or anti-PD-L1 immunotherapy: an imaging biomarker, retrospective multicohort study. Lancet Oncol. 2018;19:1180–91.
Dolladille C, Ederhy S, Sassier M, Cautela J, Thuny F, Cohen AA, Fedrizzi S, Chrétien B, Da-Silva A, Plane AF, et al. Immune checkpoint inhibitor rechallenge after immune-related adverse events in patients with cancer. JAMA Oncol. 2020;6:865–71.
Haanen J, Ernstoff M, Wang Y, Menzies A, Puzanov I, Grivas P, Larkin J, Peters S, Thompson J, Obeid M. Rechallenge patients with immune checkpoint inhibitors following severe immune-related adverse events: review of the literature and suggested prophylactic strategy. J Immunother Cancer. 2020;8(1):e000604.
Abou Alaiwi S, Xie W, Nassar AH, Dudani S, Martini D, Bakouny Z, Steinharter JA, Nuzzo PV, Flippot R, Martinez-Chanza N, et al. Safety and efficacy of restarting immune checkpoint inhibitors after clinically significant immune-related adverse events in metastatic renal cell carcinoma. J Immunother Cancer. 2020;8(1):e000144.
Abu-Sbeih H, Ali FS, Naqash AR, Owen DH, Patel S, Otterson GA, Kendra K, Ricciuti B, Chiari R, De Giglio A, et al. Resumption of immune checkpoint inhibitor therapy after immune-mediated colitis. J Clin Oncol. 2019;37:2738–45.
Pollack MH, Betof A, Dearden H, Rapazzo K, Valentine I, Brohl AS, Ancell KK, Long GV, Menzies AM, Eroglu Z, et al. Safety of resuming anti-PD-1 in patients with immune-related adverse events (irAEs) during combined anti-CTLA-4 and anti-PD1 in metastatic melanoma. Ann Oncol. 2018;29:250–5.
Simonaggio A, Michot JM, Voisin AL, Le Pavec J, Collins M, Lallart A, Cengizalp G, Vozy A, Laparra A, Varga A, et al. Evaluation of readministration of immune checkpoint inhibitors after immune-related adverse events in patients with cancer. JAMA Oncol. 2019;5:1310–7.
Santini FC, Rizvi H, Plodkowski AJ, Ni A, Lacouture ME, Gambarin-Gelwan M, Wilkins O, Panora E, Halpenny DF, Long NM, et al. Safety and efficacy of re-treating with immunotherapy after immune-related adverse events in patients with NSCLC. Cancer Immunol Res. 2018;6:1093–9.
Shankar B, Zhang J, Naqash AR, Forde PM, Feliciano JL, Marrone KA, Ettinger DS, Hann CL, Brahmer JR, Ricciuti B, et al. Multisystem immune-related adverse events associated with immune checkpoint inhibitors for treatment of non-small cell lung cancer. JAMA Oncol. 2020;6:1952–6.
Ramos-Casals M, Brahmer JR, Callahan MK, Flores-Chávez A, Keegan N, Khamashta MA, Lambotte O, Mariette X, Prat A, Suárez-Almazor ME. Immune-related adverse events of checkpoint inhibitors. Nat Rev Dis Primers. 2020;6:38.
Badran YR, Cohen JV, Brastianos PK, Parikh AR, Hong TS, Dougan M. Concurrent therapy with immune checkpoint inhibitors and TNFα blockade in patients with gastrointestinal immune-related adverse events. J Immunother Cancer. 2019;7:226.
Bellesoeur A, Torossian N, Amigorena S, Romano E. Advances in theranostic biomarkers for tumor immunotherapy. Curr Opin Chem Biol. 2020;56:79–90.
Basler L, Kowalczyk A, Heidenreich R, Fotin-Mleczek M, Tsitsekidis S, Zips D, Eckert F, Huber SM. Abscopal effects of radiotherapy and combined mRNA-based immunotherapy in a syngeneic, OVA-expressing thymoma mouse model. Cancer Immunol Immunother. 2018;67:653–62.
Wei SC, Duffy CR, Allison JP. Fundamental mechanisms of immune checkpoint blockade therapy. Cancer Discov. 2018;8:1069–86.
LaFleur MW, Muroyama Y, Drake CG, Sharpe AH. Inhibitors of the PD-1 pathway in tumor therapy. J Immunol. 2018;200:375–83.
Koyama S, Akbay EA, Li YY, Herter-Sprie GS, Buczkowski KA, Richards WG, Gandhi L, Redig AJ, Rodig SJ, Asahina H, et al. Adaptive resistance to therapeutic PD-1 blockade is associated with upregulation of alternative immune checkpoints. Nat Commun. 2016;7:10501.
Acknowledgements
Figdraw (www.figdraw.com) and SMART (smart.servier.com) were acknowledged since the main images and elements used to draw the figures were downloaded from these two websites.
Funding
This study was supported by the National Natural Science Foundation of China (81972168), Strategic Priority Research Program of the Chinese Academy of Sciences (XDB29030103) and National Key Research and Development Program of China (2021YFA1301402).
Author information
Authors and Affiliations
Contributions
Y.A. and J.G. collected relevant literature, drafted manuscripts, and prepared figures and tables. S.W., J.J. and J.D reviewed and made significant revisions to the manuscript. J.D., B.X. and H.W. conceived and designed the research study and the manuscript preparation. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no potential conflicts of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Ao, YQ., Gao, J., Wang, S. et al. Immunotherapy of thymic epithelial tumors: molecular understandings and clinical perspectives. Mol Cancer 22, 70 (2023). https://doi.org/10.1186/s12943-023-01772-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12943-023-01772-4