
Abstract
Background
Escherichia. coli is the most frequent host for New Delhi metallo-β-lactamase (NDM) which hydrolyzes almost all β-lactams except aztreonam. The worldwide spread of blaNDM-carrying E. coli heavily threatens public health.
Objective
This study aimed to explore the global genomic epidemiology of blaNDM- carrying E. coli isolates, providing information for preventing the dissemination of such strains.
Methods
Global E. coli genomes were downloaded from NCBI database and blaNDM was detected using BLASTP. Per software was used to extract meta information on hosts, resources, collection data, and countries of origin from GenBank. The sequence types (STs) and distribution of antimicrobial resistance gene (ARG) were analyzed by CLC Workbench; Plasmid replicons, serotypes and virulence genes (VFs) were analyzed by submitting the genomes to the websites. Statistical analyses were performed to access the relationships among ARGs and plasmid replicons.
Results
Until March 2023, 1,774 out of 33,055 isolates collected during 2003–2022 were found to contain blaNDM in total. Among them, 15 blaNDM variants were found with blaNDM-5 (74.1%) being most frequent, followed by blaNDM-1 (16.6%) and blaNDM-9 (4.6%). Among the 213 ARGs identified, 27 blaCTX-M and 39 blaTEM variants were found with blaCTX-M-15 (n = 438, 24.7%) and blaTEM-1B (n = 1092, 61.6%) being the most frequent ones, respectively. In addition, 546 (30.8%) plasmids mediated ampC genes, 508 (28.6%) exogenously acquired 16 S rRNA methyltransferase encoding genes and 262 (14.8%) mcr were also detected. Among the 232 distinct STs, ST167 (17.2%) were the most prevalent. As for plasmids, more than half of isolates contained IncFII, IncFIB and IncX3. The VF terC, gad, traT and iss as well as the serotypes O101:H9 (n = 231, 13.0%), O8:H9 (n = 115, 6.5%) and O9:H30 (n = 99, 5.6%) were frequently observed.
Conclusions
The study delves into the intricate relationship between plasmid types, virulence factors, and ARGs, which provides valuable insights for clinical treatment and public health interventions, and serves as a critical resource for guiding future research, surveillance, and implementation of effective strategies to address the challenges posed by blaNDM-carrying E. coli. The findings underscore the urgent need for sustained global collaboration, surveillance efforts, and antimicrobial stewardship to mitigate the impact of these highly resistant strains on public health.
Similar content being viewed by others

Introduction
Escherichia coli, a rod-shaped, gram-negative bacterium, predominantly resides in the lower intestinal tract of warm-blooded animals, including humans. Known as one of the most frequent opportunistic pathogens, it is a leading cause of urinary, bloodstream, and wound infections in both community and hospital settings. Within the realm of antibiotic resistance, a significant concern is the emergence of New Delhi metallo-β-lactamase (NDM), a member of the β1 metallo-β-lactamase class, capable of hydrolyzing almost all β-lactams except monobactams. It’s initially identified in a Swedish patient in New Delhi, India, in 2008 [1]. At present, the blaNDM variants have now spread across more than 60 species in 11 bacterial families, with E. coli being the predominant carrier of the blaNDM gene [2]. Strains belonging to the ST167, ST410, and ST617 lineages are the most prevalent clones [3, 4]. Geographically, the Indian subcontinent, the Middle East, and the Balkans are the most epidemic regions [5]. The global dissemination of blaNDM-carrying strains poses a considerable challenge for clinical management and public health, due to the heightened mortality rates associated with infections caused by these strains. Besides, various antimicrobial resistance genes (ARGs), such as non-NDM carbapenem hydrolyzing β-lactamases (CHβLs), extended-spectrum β-lactamases (ESBLs), plasmid-mediated quinolone resistance genes (PMQRs), and exogenously acquired 16 S rRNA methyltransferase (16 S-RMTase) genes, are often co-harbored with blaNDM in E. coli, leading to multi-drug resistance or pan-drug resistance [6, 7], thereby limiting antimicrobial treatment options for E. coli infection in clinical settings.
The escalating prevalence of blaNDM worldwide can be attributed to global travel and extensive antibiotic use, recognized as key population risk factors linked to the dispersal of blaNDM. Notably, the spread of blaNDM genes is primarily facilitated by mobile genetic elements (MGEs), with plasmids being the most common carriers. These blaNDM-carrying plasmids typically fall under limited replicon types, such as IncX3, IncFII, or IncC [8]. Despite numerous studies on the epidemiological characteristics of Carbapenem-resistant E. coli (CREC), comprehensive data on the virulence factors (VFs), serotypes, and sequence types (STs) of blaNDM-carrying E. coli remain limited.
This study aims to characterize the global epidemiological features of blaNDM-carrying E. coli by leveraging genomic data from GenBank. The investigation includes an analysis of the distribution of other ARGs, plasmid replicons, VFs, serotypes, and STs to provide a thorough genomic characterization. Additionally, the study explores the consistency in the distribution of plasmid replicons and VFs, as well as VFs and ARGs, shedding light on potential associations between these key elements.
Materials and methods
Download of E. coli genome data
Sequence files of all 33,055 E. coli genomes (updated to 2023.03.09) were downloaded in batch from NCBI (https://www.ncbi.nlm.nih.gov/genome/browse/#!/overview/) using the aspera high-speed download tool. For all annotated genomes, the protein coding gene sequence of each genome is obtained in batch from GenBank file by using self-made Perl script. All genomes were qualified with completeness > 90%, contamination < 5%, and contig quantity ≤ 500 and N50 ≥ 40,000.
blaNDM identification
All blaNDM sequences were obtained from the NCBI Biological Resistance Reference Gene Database (https://www.ncbi.nlm.nih.gov/pathogens/refgene/#gene_family:(blaNDM). BLASTP was performed, with thresholds being set as expected value = 1e-5, coverage ≥ 60%, identity ≥ 90%, and match length = subject gene length. Finally, the results were processed by Perl program to obtain the detailed distribution of blaNDM gene in E. coli genomes.
Extraction of meta-information on the blaNDM-carrying E. coli
Meta information on hosts, resources, collection data, and countries of origin was extracted from GenBank using Per software. This information was integrated with the ARGs, STs, VFs and serotypes into the same excel for further analysis.
Investigation into the prevalence of antimicrobial resistance genes
The distribution of other ARGs was investigated using CLC workbench version 21.0.1. The fasta file was input into the files in CLC Workbench using standard import, and after the consensus sequence was extracted, the prevalence of ARGs was analyzed using the ResFinder database for comparison. The results were exported as scv files, and further sorted for analysis.
Sequence types
STs of blaNDM-carrying E. coli were identified using CLC workbench 21.0.1. After consensus sequences were extracted, STs were analyzed using multi-locus sequence typing (MLST) with E. coli (Oxford) as the reference database.
Distributions of plasmid replicons, VFs, and serotypes
Genomes were submitted to the Center for Genomic Epidemiology (http://www.Genomicepidemiology.org/). Plasmid Finder 2.1 was used to identify plasmid replicons, and VFs were analyzed by Virulence Finder 2.0. Moreover, serotypes were identified by Serotype Finder 2.0.
Statistical analyses
Correlation analyses were performed using SPSS 22.0. The distribution consistency of VFs and ARGs was tested by McNimar analysis, and a p-value of > 0.05 was taken as the consistency between them.
Results
The prevalent characteristics of blaNDM-carrying E. coli
Totally, 33,055 E. coli isolates were downloaded from NCBI database and the time span was from 2003 to 2022. Of which, 1,774 were identified to be positive for the blaNDM gene (Additional file 1). These isolates were obtained from 43 countries across 6 continents (Fig. 1), which were as follows: Asia (n = 1376, 77.6%), Europe (n = 196, 11.0%), North America (n = 63, 3.6%), Africa (n = 29, 1.6%), South America (n = 29, 1.6%), and Oceania (n = 2, 0.1%). Of concern, China (n = 1127, 63.5%), India (n = 115, 6.5%), and France (n = 83, 4.7%) were the primary contributors, submitting the highest number of genomes, following closely were the USA (n = 56), Germany (n = 33), Thailand (n = 30), United Kingdom (n = 21), Bangladesh (n = 20), South Korea (n = 19), Lebanon (n = 16), and Pakistan (n = 11). The origin of the remaining 79 isolates was unspecified.

Geographical distribution of blaNDM and STs of blaNDM-carrying Escherichia coli worldwide. Hollow and solid pie charts of each continent represent blaNDM and STs, respectively
Of 1,774 blaNDM genes, 15 distinct blaNDM variants were identified. While the number of blaNDM-carrying E. coli showed a gradual increase each year, it surged significantly in 2015 (n = 350). This elevated level was sustained from 2016 to 2019, followed by a decline from 2020 to 2022 (Fig. 2).

Number of global blaNDM-carrying Escherichia coli isolates submitted per year
Regarding the sources of the blaNDM-carrying E. coli isolates (Table 1), it was observed that Homo sapiens accounted for the majority, constituting 59.6% (n = 1,057) of the total. These isolates were predominantly sourced from urine (n = 232), blood (n = 162), rectal/anal swabs (n = 149), sputum (n = 81), and fecal samples (n = 86). Animals comprised 27.4% (n = 486) of the isolates, with chickens (n = 182), pigs (n = 59), and flies (n = 46) being the most prevalent species. The primary sources for animal isolates included fecal samples, cloaca swabs, and various organs (intestine/liver/other). Notably, blaNDM-carrying strains were also detected in the environment, accounting for 13.0% (n = 231) of the isolates. These environmental sources included water, hospitals, and various food items.
Wide distribution of various resistance genes among blaNDM-carrying E. coli
Among the 1,774 blaNDM-carrying isolates, 15 distinct variants were identified, with blaNDM-5 being the most prevalent (n = 1,315, 74.1%), followed by blaNDM-1 (n = 295, 16.6%) and blaNDM-9 (n = 82, 4.6%). Other variants included blaNDM-7, blaNDM-4, blaNDM-6, blaNDM-13, blaNDM-3, blaNDM-15, blaNDM-19, blaNDM-16, blaNDM-20, blaNDM-21, blaNDM-22, and blaNDM-24, each with lower frequencies. Geographically, blaNDM-5 dominated in Asia, Europe, Africa, and North America, constituting 62.0-77.0% of cases (Fig. 1). Notably, South America exhibited a distinct pattern, with blaNDM-1 being the most common variant, representing 93.0% of cases.
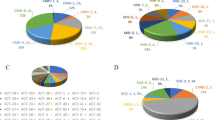
A comprehensive analysis of ARGs in blaNDM-carrying strains revealed 213 different types. Among them, CHßLs encoding genes including 8 blaKPC-2, 1 blaIMP-1, 34 blaOXA-181, 10 blaOXA-232, 5 blaOXA-244 and 5 blaOXA-48 were identified. Moreover, 27 blaCTX-M and 39 blaTEM variants were detected with blaCTX-M-15 (n = 438, 24.7%), blaCTX-M-55 (n = 300, 16.9%), blaCTX-M-14 (n = 204, 11.5%), blaCTX-M-65 (n = 147, 8.3%) and blaTEM-1B (n = 1092, 61.6%) being the most frequent ones, respectively. In addition, 546 (30.8%) plasmids mediated ampC genes, including 501 blaCMY and 45 blaDHA as well as 508 (28.6%) 16S-RMTase encoding genes, including 409 rmtB, 55 rmtC and 44 armA were found. Of significant concern, 262 (14.8%) co-existing mcr genes were also detected, with mcr1.1 being the most prevalent genotype (n = 249, 14.0%). Furthermore, 467 (26.3%) fosfomycin resistance genes including 459 fosA3 and 8 fosA4 as well as 546 (30.8%) aac(6’)-ib-cr conferring resistance to amikacin and fluoroquinolones in addition to 805 (45.4%) PMQRs including 239 oqxAB, 126 qepA, and 440 qnr were identified. Other main ARGs detected were shown in Fig. 3.
Multiple distinct sequence types were identified with several high-risk clones being prevalent
A total of 232 distinct STs were identified among the blaNDM-carrying E. coli isolates. The most prevalent was ST167 (n = 306, 17.2%), followed by ST410 (n = 174, 9.8%), ST361 (n = 108, 6.1%), ST405 (n = 85, 4.8%), ST156 (n = 74, 4.2%), ST10 (n = 73, 4.1%), ST48 (n = 59, 3.3%), ST617 (n = 53, 3.0%), ST101 (n = 50, 2.8%), ST648 (n = 44, 2.5%), and ST746 (n = 38, 2.1%). Several other STs were also identified, with each less than 30. Geographically, the distribution of STs varied. ST167 was predominant in both Asia and North America, ST361 was endemic in Europe, and ST410 emerged as the predominant clone in Africa, with ST10 dominating in South America (Fig. 1).

The antimicrobial resistance genes identified among 1774 blaNDM-carrying Escherichia coli. 3 A. all the antimicrobial resistance genes; 3B. mcr variants; 3 C, blaOXA variants; 3D. blaCMY variants; 3E. blaNDM variants; 3 F. blaCTX-M variants; 3G. blaTEM variants
Virulence factors
A total of 170 distinct VFs were identified among the blaNDM-carrying E. coli isolates. The most prevalent VF was terC (n = 1,766, 99.5%). Notably, more than half of the isolates carried four specific VFs: gad (83.4%, n = 1,480), traT (67.9%, n = 1,205), and iss (51.3%, n = 910). Several other VFs were also frequently detected, including sitA (n = 756, 42.6%), hra (n = 689, 38.8%), lpfA (n = 631, 35.6%), fyuA (n = 561, 31.6%), and irp2 (n = 560, 31.6%).
Of significant concern is the differential distribution of the predominantly prevalent VFs among the endemic clones. Notably, lpfA was primarily distributed among ST410 and ST156, while iss was predominantly concentrated in ST167 and ST156. The dominance of fyuA and irp2 was observed among ST405, with ST361 and ST405 being the most frequent carriers for sitA.
Multiple Plasmid replicons were found among blaNDM-carrying E. coli which may facilitate the spread of antimicrobial resistance genes
Various plasmid replicons were identified, with IncFII (n = 1,163, 65.6%) emerging as the most prevalent. Following closely were IncFIB (n = 1,157, 65.2%), IncX3 (n = 888, 50.1%), IncFIA (n = 756, 42.6%), COL (n = 440, 24.8%), IncY (n = 338, 19.1%), IncI1-I (n = 321, 18.1%), P0111 (n = 306, 17.2%), IncHI2 (n = 287, 16.2%), IncQ1 (n = 194, 10.9%), IncI (n = 190, 10.7%), IncFIC (n = 179, 10.1%), IncC (n = 174, 9.8%), IncR (n = 114, 6.4%), IncX1 (n = 128, 7.2%), IncN (n = 77, 4.3%), and several other rare plasmid replicons.
Analysis revealed that 44.7–66.8% of blaNDM-5-carrying strains carried IncFII, IncFIB, IncX3, and IncFIA plasmids (Table 2). For blaNDM-1-carrying strains, over half carried IncFII and IncFIB plasmids, with IncFIA and Col being relatively highly prevalent. IncFIB, IncFII, and Col were frequently carried by blaNDM-9 positive strains. Notably, IncFIA was particularly prevalent, reaching 80.9%, in blaNDM-7-carrying strains, followed by IncFIB, IncFII, and IncX3.
Analyzing the consistency between prevalent plasmids and resistant/virulent genes (Table 3) revealed correlations between the prevalence of mph(A) and tra(T) and the plasmid replicons IncFII and IncFIB (p > 0.05). The incidences of aph(6)-Id, aadA2, aph(3’’)-Ib, dfrA12, and iss were all correlated with the presence of plasmid IncX3. Additionally, the distribution of blaOXA-1, iucC, and capU was consistent with the presence of plasmid Col.
The prevalence of ARGs and VFs among epidemic clones ST167, ST410, ST361, ST405 and ST156 were demonstrated in the Fig. 4. More than 42.5% of them carried blaTEM, blaCTX-M, blaNDM-5, traT, gad and terC, constituting a basic resistant and virulent profile. In detail, ST167 clones exhibited high incidences of iss and hra; S156 showed a high carriage of mcr, rmt, lpdA, iss, and hra. Notably, ST410 clones demonstrated nearly 100% carriage of lpfA and high incidences of aac(6’)-ib-cr and blaOXA, while ST405 showed a high prevalence of fyuA, irp2, sitA, traT, gad and rmt. This highlights the distinct prevalence patterns of ARGs among different STs, emphasizing the diversity in resistance and virulence profiles associated with specific E. coli lineages.

The prevalence of prevalent antimicrobial resistance genes and virulent factors among epidemic clones. Antimicrobial resistance genes were aac(6’)-ib-cr, mcr, rmt, qnr, blaOXA, blaTEM, blaCTX-M, blaNDM-5; Virulence factors were terC, gad, traT, iss, sitA, hra, lpfA, fyuA, irp2
A large number of serotypes detected among blaNDM-carrying E. coli with O101:H9 and O8:H9 being the predominate
A total of 91 distinct O types were identified, with O101 (n = 391, 22.0%), O8 (n = 179, 10.1%), O9 (n = 154, 8.7%), and O102 (n = 92, 5.2%) emerging as the dominant ones. Among the 43 H types, H9 (n = 470, 26.5%) was the most prevalent, followed by H30 (n = 140, 7.9%), H6 (n = 130, 7.3%), H10 (n = 121, 6.8%), and H5 (n = 114, 6.4%). Over 100 distinct serotypes were identified, with O101:H9 (n = 231, 13.0%), O8:H9 (n = 115, 6.5%), O9:H30 (n = 99, 5.6%), O102:H6 (n = 86, 4.8%), and O101:H10 (n = 77, 4.3%) being the most frequent.
Discussion
The increasing prevalence of blaNDM-carrying E. coli presents significant challenges to clinical treatment and global public health, prompting a critical need to bolster infection control measures in hospitals. The epidemiological and genomic characterization of 1,774 blaNDM-carrying E. coli isolates from 43 countries (2003–2022) provides valuable insights for guiding clinical treatment strategies and implementing preventative measures.
Our study showed that prevalence rates of blaNDM-carrying E. coli varied globally, with Asia reporting the highest, followed by Europe (11.0%), America (5.2%), Africa (1.6%), and Oceania (0.01%). These rates closely align with a meta-analysis of 110 studies from 2008 to 2018 [9], confirming the high prevalence of such strain in Asia. In addition, we identified a notable surge of blaNDM in E. coli isolates during 2015–2019, with blaNDM-5 emerging as the most frequent variant. Consistent with this, an upward trend in blaNDM-carrying Enterobacterale has been observed in France since 2012, in Switzerland since 2013, and in Germany from 2013 to 2019 [10,11,12,13]. Particularly concerning is the rapid global spread of blaNDM-5 producing E. coli, contributing to an epidemic situation in India, China, and sub-Saharan Africa [5, 14]. Furthermore, the global prevalence of blaNDM experienced a decline in 2020 and 2021, likely attributed to some sequence data not being released at the time of our data retrieval in 2023. Moreover, our study showed that animals and environments constituted 40% of the studied resources, which may relate with the use of carbapenem antibiotics in poultry breeding. Notably, as observed in other studies, there was a substantial prevalence of blaNDM-5 in poultry and farm environments, which may be spread from healthcare settings [15].
Of great note, our findings align with previous research, affirming that all blaNDM-positive E. coli isolates in our study co-carried additional ARGs. The most prevalent among these were sul1 and sul2, encoding an alternative dihydropteroate synthase, emphasizing the need for cautious prescription of sulfonamides targeting CREC. Furthermore, we observed the coexistence of blaESBL, particularly blaTEM-1b and blaCTX-M-15, with the blaNDM-5 gene, consistent with prior reports [12, 16]. This underscores the complexity of resistance profiles and emphasizes the importance of comprehensive antimicrobial stewardship. It is noteworthy that in our study, 61.1% of blaNDM-carrying E. coli concurrently carried the tet(A) gene, known to result in reduced susceptibility to tigecycline, signaling a need for cautious use of tigecycline in treating such strains [17, 18]. Additionally, a significant 14.0% of blaNDM-carrying E. coli harbored the mcr1.1 gene, a horizontally transmitted colistin resistance gene [19]. This finding is of great concern as it poses a challenge to the effectiveness of colistin in treating infections caused by these strains. Further complicating matters, our study identified nine derivatives of mcr (mcr-2 to mcr-10) [20], suggesting a potential reduction in polymyxin sensitivity among blaNDM-positive strains due to the widespread presence of mcr. Furthermore, 46.6% of blaNDM-positive strains simultaneously carried the Trimethoprim-resistance gene dfrA12 and Florfenicol resistance genes floR, indicating a multi-drug resistance profile. Notably, in strains carrying both mcr-1 and blaNDM-5, our study revealed IncFIB as the main plasmid carrying both genes, deviating from previous observations [21, 22], possibly due to the extensive inclusion of strains in our analysis. This emphasizes the intricate interplay of resistance mechanisms and highlights the necessity for vigilant antimicrobial management strategies.
ST analysis revealed a diverse array of STs among blaNDM-carrying E. coli. Notably, ST167 dominated in China and India, while ST361 was predominant in Europe, diverging from previous reports which indicated that ST101 and ST131 are the most prevalent clones in Asia, ST101 and ST405 are the dominant STs in Europe [9]. This discrepancy may arise from the focus on blaNDM-carrying E. coli in our study, reflecting distinct strain compositions. Consistent with other studies, ST167 and ST410 emerged as international epidemic clones linked with blaNDM-5 [23, 24]. Contrary to these findings, no specific ST linkage was observed for blaNDM-1 [5], although some strains were assigned to ST410, ST156, ST167, ST10, and ST101. Remarkably, ST13, recognized as the pandemic clone associated with global dissemination of the blaCTX-M-15 [25, 26], was identified in only 17 blaNDM-carrying E. coli isolates, underscoring a divergence in the dissemination patterns of different ARGs.
Among various VFs, over half of the strains carried terC, gad, traT, and iss. terC, encoding a tellurium iron resistance protein [27]. The gad gene, facilitating survival in acidic environments, and the traT gene, which encodes protections, along with iss, sitA, and hra genes being associated with urinary tract infection, were also prevalent [28, 29]. Importantly, the distribution of predominant VFs varied significantly among the prevailing clones, highlighting the intricate relationship between genetic backgrounds and virulence traits [30].
IncFII and IncFIB emerged as the predominant plasmids in all blaNDM-carrying isolates, showcasing a correlation with the mph(A) and tra(T) genes. Following closely were IncX3 and IncFIA. Notably, it’s reported that IncX3-type plasmids are recognized as key contributors to horizontal transmission of blaNDM in CREC [31] and IncFIB is prevalent especially in blaNDM-1-carrying E. coli in Greece [32]. Serotype O101, associated with both animal and human diseases, is commonly detected among pathogenic E. coli [33]. In our study, O101:H9 emerged as the most prevalent, even though reports of this specific serotype are limited. Interestingly, O101:H9 has been linked to possible international dissemination via migratory birds [34], emphasizing the importance of employing techniques such as multiplex PCR or whole-genome sequencing that go beyond focusing solely on serotype O157:H7 in clinical laboratories. These diverse molecular findings underscore the necessity for ongoing surveillance of blaNDM-carrying E. coli strains.
To our best knowledge, this study represents the largest comprehensive report on the prevalence and genetic characterization of blaNDM-carrying E. coli worldwide, utilizing whole-genome data. However, it is crucial to acknowledge certain limitations. Firstly, the inclusion criteria were limited to data available on the NCBI database, potentially introducing selection bias. Secondly, the study’s ability to fully capture the global prevalence of blaNDM-carrying E. coli is constrained by the insufficient availability of genome data from many countries. Thirdly, the absence of detailed epidemiological data limits the exploration of factors such as prior exposure to healthcare settings and environmental influences, which may play a significant role in the global transmission of blaNDM-carrying E. coli. Despite these limitations, this study offers valuable insights into the epidemiology and genomic characteristics of blaNDM-carrying E. coli on a global scale.
Conclusions
This study offers a comprehensive understanding of the prevalence, genetic diversity, and associated factors of blaNDM-carrying E. coli, submitted by 43 countries over the period 2003–2022. blaNDM-5 is the predominant variant, identified in diverse ST and O: H serotypes, highlighting the complexity and adaptability of these multidrug-resistant strains. The study delves into the intricate relationship between plasmid types, virulence factors, and ARGs, providing valuable insights for clinical treatment and public health interventions. As the global threat of antimicrobial resistance continues to escalate, this study serves as a critical resource for guiding future research, surveillance, and implementation of effective strategies to address the challenges posed by blaNDM-carrying E. coli. The findings underscore the urgent need for sustained global collaboration, surveillance efforts, and antimicrobial stewardship to mitigate the impact of these highly resistant strains on public health.
Data availability
No datasets were generated or analysed during the current study.
References
Yong D, Toleman MA, Giske CG, Cho HS, Sundman K, Lee K, et al. Characterization of a new metallo-beta-lactamase gene, bla(NDM-1), and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob Agents Chemother. 2009;53(12):5046–54. https://doi.org/10.1128/aac.00774-09.
Farhat N, Khan AU. Evolving trends of New Delhi Metallo-Betalactamse (NDM) variants: a threat to antimicrobial resistance. Infect Genet Evol. 2020;86:104588. https://doi.org/10.1016/j.meegid.2020.104588.
Grundmann H, Glasner C, Albiger B, Aanensen DM, Tomlinson CT, Andrasević AT, et al. Occurrence of carbapenemase-producing Klebsiella pneumoniae and Escherichia coli in the European survey of carbapenemase-producing Enterobacteriaceae (EuSCAPE): a prospective, multinational study. Lancet Infect Dis. 2017;17(2):153–63. https://doi.org/10.1016/s1473-3099(16)30257-2.
Hu X, Xu X, Wang X, Xue W, Zhou H, Zhang L, et al. Diversity of New Delhi metallo-beta-lactamase-producing bacteria in China. Int J Infect Dis. 2017;55:92–5. https://doi.org/10.1016/j.ijid.2017.01.011.
Wu W, Feng Y, Tang G, Qiao F, McNally A, Zong Z. NDM Metallo-β-Lactamases and their bacterial producers in Health Care settings. Clin Microbiol Rev. 2019;32(2). https://doi.org/10.1128/cmr.00115-18.
Zhong LL, Zhang YF, Doi Y, Huang X, Zhang XF, Zeng KJ, et al. Coproduction of MCR-1 and NDM-1 by Colistin-Resistant Escherichia coli isolated from a healthy individual. Antimicrob Agents Chemother. 2017;61(1). https://doi.org/10.1128/aac.01962-16.
Lin D, Xie M, Li R, Chen K, Chan EW, Chen S. IncFII Conjugative plasmid-mediated transmission of blaNDM-1 elements among animal-borne Escherichia coli strains. Antimicrob Agents Chemother. 2017;61(1). https://doi.org/10.1128/aac.02285-16.
Zhang Z, Guo H, Li X, Li W, Yang G, Ni W, et al. Genetic diversity and characteristics of bla (NDM)-Positive plasmids in Escherichia coli. Front Microbiol. 2021;12:729952. https://doi.org/10.3389/fmicb.2021.729952.
Dadashi M, Yaslianifard S, Hajikhani B, Kabir K, Owlia P, Goudarzi M, et al. Frequency distribution, genotypes and prevalent sequence types of New Delhi metallo-β-lactamase-producing Escherichia coli among clinical isolates around the world: a review. J Glob Antimicrob Resist. 2019;19:284–93. https://doi.org/10.1016/j.jgar.2019.06.008.
Dortet L, Cuzon G, Ponties V, Nordmann P. Trends in carbapenemase-producing Enterobacteriaceae, France, 2012 to 2014. Euro Surveill. 2017;22(6). https://doi.org/10.2807/1560-7917.Es.2017.22.6.30461.
Ramette A, Gasser M, Nordmann P, Zbinden R, Schrenzel J, Perisa D, et al. Temporal and regional incidence of carbapenemase-producing enterobacterales, Switzerland, 2013 to 2018. Euro Surveill. 2021;26(15). https://doi.org/10.2807/1560-7917.Es.2021.26.15.1900760.
Hans JB, Pfennigwerth N, Neumann B, Pfeifer Y, Fischer MA, Eisfeld J, et al. Molecular surveillance reveals the emergence and dissemination of NDM-5-producing Escherichia coli high-risk clones in Germany, 2013 to 2019. Euro Surveill. 2023;28(10). https://doi.org/10.2807/1560-7917.Es.2023.28.10.2200509.
Kremer K, Kramer R, Neumann B, Haller S, Pfennigwerth N, Werner G, et al. Rapid spread of OXA-244-producing Escherichia coli ST38 in Germany: insights from an integrated molecular surveillance approach; 2017 to January 2020. Euro Surveill. 2020;25(25). https://doi.org/10.2807/1560-7917.Es.2020.25.25.2000923.
Bi R, Kong Z, Qian H, Jiang F, Kang H, Gu B, et al. High prevalence of Bla (NDM) variants among Carbapenem-Resistant Escherichia coli in Northern Jiangsu Province, China. Front Microbiol. 2018;9:2704. https://doi.org/10.3389/fmicb.2018.02704.
Wei H, Kong L, Wang Y, Huang Z, Yang X, Zhou C, et al. Characterization and Public Health Insights of the New Delhi Metallo-β-Lactamase-producing enterobacterales from laying hens in China. Microorganisms. 2022;10(4). https://doi.org/10.3390/microorganisms10040800.
Yin C, Yang W, Lv Y, Zhao P, Wang J. Clonal spread of carbapenemase-producing Enterobacteriaceae in a region, China. BMC Microbiol. 2022;22(1):81. https://doi.org/10.1186/s12866-022-02497-y.
Chiu SK, Huang LY, Chen H, Tsai YK, Liou CH, Lin JC, et al. Roles of ramR and tet(A) mutations in conferring Tigecycline Resistance in Carbapenem-resistant Klebsiella pneumoniae clinical isolates. Antimicrob Agents Chemother. 2017;61(8). https://doi.org/10.1128/aac.00391-17.
Yao H, Cheng J, Li A, Yu R, Zhao W, Qin S, et al. Molecular characterization of an IncFII(k) plasmid co-harboring bla (IMP-26) and tet(A) variant in a clinical Klebsiella pneumoniae isolate. Front Microbiol. 2020;11:1610. https://doi.org/10.3389/fmicb.2020.01610.
Liu YY, Wang Y, Walsh TR, Yi LX, Zhang R, Spencer J, et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. Lancet Infect Dis. 2016;16(2):161–8. https://doi.org/10.1016/s1473-3099(15)00424-7.
Hussein NH, Al-Kadmy IMS, Taha BM, Hussein JD. Mobilized colistin resistance (mcr) genes from 1 to 10: a comprehensive review. Mol Biol Rep. 2021;48(3):2897–907. https://doi.org/10.1007/s11033-021-06307-y.
Mediavilla JR, Patrawalla A, Chen L, Chavda KD, Mathema B, Vinnard C, et al. Colistin- and Carbapenem-Resistant Escherichia coli Harboring mcr-1 and blaNDM-5, causing a complicated urinary tract infection in a patient from the United States. mBio. 2016;7(4). https://doi.org/10.1128/mBio.01191-16.
Yu H, Qu F, Shan B, Huang B, Jia W, Chen C, et al. Detection of the mcr-1 Colistin Resistance Gene in Carbapenem-Resistant Enterobacteriaceae from different hospitals in China. Antimicrob Agents Chemother. 2016;60(8):5033–5. https://doi.org/10.1128/aac.00440-16.
Zhang R, Liu L, Zhou H, Chan EW, Li J, Fang Y, et al. Nationwide Surveillance of Clinical Carbapenem-resistant Enterobacteriaceae (CRE) strains in China. EBioMedicine. 2017;19:98–106. https://doi.org/10.1016/j.ebiom.2017.04.032.
Peirano G, Chen L, Nobrega D, Finn TJ, Kreiswirth BN, DeVinney R, et al. Genomic epidemiology of global carbapenemase-producing Escherichia coli, 2015–2017. Emerg Infect Dis. 2022;28(5):924–31. https://doi.org/10.3201/eid2805.212535.
Nicolas-Chanoine MH, Blanco J, Leflon-Guibout V, Demarty R, Alonso MP, Caniça MM, et al. Intercontinental emergence of Escherichia coli clone O25:H4-ST131 producing CTX-M-15. J Antimicrob Chemother. 2008;61(2):273–81. https://doi.org/10.1093/jac/dkm464.
Rogers BA, Sidjabat HE, Paterson DL. Escherichia coli O25b-ST131: a pandemic, multiresistant, community-associated strain. J Antimicrob Chemother. 2011;66(1):1–14. https://doi.org/10.1093/jac/dkq415.
Maltman C, Yurkov V. Extreme environments and high-level bacterial Tellurite Resistance. Microorganisms. 2019;7(12). https://doi.org/10.3390/microorganisms7120601.
Schwan WR, Flohr NL, Multerer AR, Starkey JC. GadE regulates fliC gene transcription and motility in Escherichia coli. World J Clin Infect Dis. 2020;10(1):14–23. https://doi.org/10.5495/wjcid.v10.i1.14.
Giovannercole F, Mérigoux C, Zamparelli C, Verzili D, Grassini G, Buckle M, et al. On the effect of alkaline pH and cofactor availability in the conformational and oligomeric state of Escherichia coli glutamate decarboxylase. Protein Eng Des Sel. 2017;30(3):235–44. https://doi.org/10.1093/protein/gzw076.
Ovi F, Zhang L, Nabors H, Jia L, Adhikari P. A compilation of virulence-associated genes that are frequently reported in avian pathogenic Escherichia coli (APEC) compared to other E. Coli. J Appl Microbiol. 2023;134(3). https://doi.org/10.1093/jambio/lxad014.
Xu J, Guo H, Li L, He F. Molecular epidemiology and genomic insights into the transmission of carbapenem-resistant NDM-producing Escherichia coli. Comput Struct Biotechnol J. 2023;21:847–55. https://doi.org/10.1016/j.csbj.2023.01.004.
Tsilipounidaki K, Florou Z, Skoulakis A, Fthenakis GC, Miriagou V, Petinaki E. Diversity of bacterial clones and plasmids of NDM-1 producing Escherichia coli Clinical isolates in Central Greece. Microorganisms. 2023;11(2). https://doi.org/10.3390/microorganisms11020516.
Chirila F, Tabaran A, Fit N, Nadas G, Mihaiu M, Tabaran F, et al. Concerning increase in Antimicrobial Resistance in Shiga Toxin-Producing Escherichia coli isolated from young animals during 1980–2016. Microbes Environ. 2017;32(3):252–9. https://doi.org/10.1264/jsme2.ME17023.
He WY, Zhang XX, Gao GL, Gao MY, Zhong FG, Lv LC, et al. Clonal spread of Escherichia coli O101: H9-ST10 and O101: H9-ST167 strains carrying fosA3 and bla (CTX-M-14) among diarrheal calves in a Chinese farm, with Australian Chroicocephalus as the possible origin of E. Coli O101: H9-ST10. Zool Res. 2021;42(4):461–8. https://doi.org/10.24272/j.issn.2095-8137.2021.153.
Acknowledgements
We would like to express our gratitude to everyone who participated in this research study.
Funding
This study was supported by the National Natural Science Foundation of China (81902124).
Author information
Authors and Affiliations
Contributions
XC contributed to the experimental design of the study. CX, RY, JZ, JZ and XC performed data acquisition and statistical analysis. CX, RY and XC performed bioinformatics analysis and writing. All authors contributed to the article and approved the submitted version.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable (All the data were downloaded from NCBI database).
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Xia, C., Yan, R., Liu, C. et al. Epidemiological and genomic characteristics of global blaNDM-carrying Escherichia coli. Ann Clin Microbiol Antimicrob 23, 58 (2024). https://doi.org/10.1186/s12941-024-00719-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12941-024-00719-x