
Abstract
Background
Bacterial infections in COVID-19 patients, especially those caused by multidrug-resistant gram-negative strains, are associated with increased morbidity, hospital stay and mortality. However, there is limited data on the epidemiology of extended-spectrum β-lactamase (ESBL)-producing bacteria in COVID-19 patients. Here, we assessed the prevalence and the factors associated with ESBL-producing gram-negative bacterial (GNB) infections among severely ill COVID-19 patients admitted in Kenyatta National Hospital (KNH), Kenya.
Methods
We adopted a descriptive cross-sectional study design for patients admitted between October 2021 and February 2022, purposively recruiting 120 SARS-CoV- 2 infected participants based on clinical presentation. Demographics and clinical characteristics data were collected using structured questionnaires and case report forms. Clinical samples were collected and analyzed by standard microbiological methods in the KNH Microbiology laboratory and the Centre for Microbiology Research, Kenya Medical Research Institute.
Results
GNB infections prevalence was 40.8%, majorly caused by ESBL—producers (67.3%) predominated by Klebsiella pneumoniae (45.5%). Generally, 73% of the ESBL producers harboured our target ESBL genes, mainly CTX-M-type (59%, 17/29) in K. pneumoniae (76.9%, 20/26). GNB harbouring TEM-type (83%, 10/12) and SHV-type (100%, 7/7) genes showed ESBLs phenotypes and inhibitor resistance, mainly involving clavulanate, but most of them remained susceptible to tazobactam (60%, 6/10). SHV-type genes carrying ESBL producers showed resistance to both cefotaxime (CTX) and ceftazidime (CAZ) (K. pneumoniae), CAZ (E. coli) or CTX (E. cloacae complex and K. pneumoniae). About 87% (20/23) of isolates encoding CTX-M-type β-lactamases displayed CTX/ceftriaxone (CRO) resistance phenotype. About 42% of isolates with CTX-M-type β-lactamases only hydrolyzed ceftazidime (CAZ). Isolates with OXA-type β-lactamases were resistant to CTX, CAZ, CRO, cefepime and aztreonam. Patients with comorbidities were 10 times more likely to have an ESBL-producing GNB infection (aOR = 9.86, 95%CI 1.30 – 74.63, p = 0.003).
Conclusion
We report a high prevalence of ESBL-GNB infections in severely ill COVID-19 patients, predominantly due to Klebsiella pneumoniae harbouring CTX-M type ESBL genes. The patient’s underlying comorbidities increased the risk of ESBL-producing GNB infection. In COVID-19 pandemic, enhanced systematic and continuous surveillance of ESBL-producing GNB, strict adherence to infection control measures and antimicrobial stewardship policies are warranted in the current study setting.
Similar content being viewed by others


Background
Infection with Severe Acute Respiratory Syndrome Coronavirus-2 (SARS-CoV-2), the cause of Coronavirus Disease-2019 (COVID-19), suppresses host immunity through aberrant immune system activation and inflammatory cytokines overproduction [1]. Coupled with viral-induced epithelial damage [2], immune suppression favours bacterial colonization and subsequent infection [3]. Bacterial co-infections in COVID-19 patients, especially those caused by multidrug-resistant (MDR) Gram-negative strains, can result in prolonged hospitalization and higher mortality [4,5,6].Factors such as prior hospitalization, underlying medical conditions, immunosuppression, exposure to invasive medical procedures, and admission to intensive care units are associated with increased risk of MDR infections in non-COVID-19 patients [7].
Due to the lack of treatment guidelines at the beginning of the COVID-19 pandemic, most patients received broad-spectrum antibiotics [8]. Even though the impact of increased antibiotic use during the pandemic is still unclear, there was increased geographical distribution of carbapenemases, plasmid-encoded bacterial enzymes that hydrolyse carbapenem [8,9,10] in Latin America and the Caribbean [8]. However, the impact on the epidemiology of extended-spectrum β-lactamases (ESBL), with a similar transmission mechanism to carbapenemases, is unclear.
ESBLs are a group of bacterial enzymes that hydrolyse expanded spectrum β-lactam, thus mediating resistance against penicillins and cephalosporins [11]. These enzymes, produced predominantly by GNB, are worrisome because they can spread rapidly among clinical isolates through mobile genetic elements, which frequently co-harbour other non- β-lactam resistance genes, such as colistin [12, 13] aminoglycosides [14], and quinolones [15]. Surge in ESBL- producing bacterial infections can increase the use of carbapenems, which are among the drugs of last-resort in treatment of multidrug-resistant bacterial infections, posing a serious negative implication in clinical practice.
ESBL-production phenotype is mediated by several ESBL families, such as TEM, SHV, CTX-M, GES, PER, VEB, and BEL [11], with CTX-M-type β-lactamases mostly predominating [11]. Typically, beta-lactam combined with inhibitors, such as clavulanic acid, tazobactam or sulbactam, neutralize ESBL activities. Some TEM and SHV variants are resistant to inhibitors and, similar to other ESBLs, show geographical variation based on human mobility [11, 16, 17]. However, data on co-infections with ESBL-producing bacteria among COVID-19 patients in many developing countries, particularly in Sub-Saharan Africa, is limited. Therefore, we assessed the prevalence and risk factors for co-infection with ESBL-producing GNB among severely ill patients admitted in a Kenyan hospital unit dedicated to COVID-19 patients.
Methods
Study area, study design and data collection
We conducted this study in the Infectious Disease Unit (IDU), a ward dedicated to COVID-19 patients at Kenyatta National Hospital (KNH), Kenya, between October 2021 and February 2022. We adopted a descriptive cross-sectional study design among severely ill patients with confirmed (real-time reverse transcription and quantitative polymerase chain reaction (RT-qPCR) SARS-CoV-2 infection. Selection of severely ill COVID-19 participants was based on the WHO definition of severe COVID-19 illness; defined as, a critical condition where patients exhibit significant oxygen saturation deficits, impaired oxygen exchange in the lungs, rapid and labored breathing, or extensive lung infiltrates, all of which point to a severe respiratory and medical challenge associated with COVID-19 [18, 19]. This study purposively recruited 120 SARS-CoV-2 infected participants based on patients' clinical presentation suggesting bacteria infection as judged by the treating clinicians, and excluded those who, through their close relatives or legally authorized representative, declined consent to participate.
Data on demographics and clinical characteristics were collected using structured questionnaires and case report forms. Blood samples were collected directly into sterile blood culture bottles (bioMérieux, Marcy l´Etoile, France), observing the standard microbiological operating procedures [20]. Nasopharyngeal (NP) and oropharyngeal (OP) swabs (Sigma-Aldrich, India) and tracheal aspirate samples were collected by a licensed personnel into sterile containers, and transported in an ice box to the hospital Microbiology laboratory for immediate analysis.
Bacteria isolation and identification
Bacterial isolation followed the standard microbiological methods [21]. We cultured NP/OP swabs and tracheal aspirate samples on sheep blood agar (Oxoid, United Kingdom) and MacConkey (Oxoid, United Kingdom), with an overnight incubation at 37 °C. Blood culture bottles were incubated in BACT/ALERT® VIRTUO 3D Microbial Detection Systems (bioMérieux, Marcy l'Etoile, France), followed by sub-culture for the positive samples onto chocolate blood agar (CBA) (Oxoid, United Kingdom), sheep blood agar (Oxoid, United Kingdom) and MacConkey (Oxoid, United Kingdom). After subculture, we incubated the plates in ambient air; and 5–10% CO2 at 37 °C overnight, followed by isolates' identification using VITEK Matrix-assisted laser desorption ionization–time-of-flight mass spectrometry (MALDI-TOF MS) (bioMérieux, Marcy l'Etoile, France). For quality control, we used Escherichia coli ATCC 8739. All the GNB isolates were transported to the Centre for Microbiology Research, Kenya Medical Research Institute (CMR-KEMRI) laboratories for further analysis.
Screening for ESBL production
We screened the isolates for ESBL production using the Double Disk Synergy Test (DDST) [22]. A 0.5 McFarland-equivalent suspension of bacterial isolate were plated on Mueller-Hinton Agar (MHA) and allowed to air dry for 3 min. Antibiotic disks, including cefotaxime (30 µg), ceftazidime (30 µg), and amoxicillin/clavulanic acid (20 µg/10 µg), were added at a 30 mm radius to radius distance and incubated overnight in ambient air at 37 ℃. An inhibition zone around the cefotaxime and/or ceftazidime that increased towards the β-lactam inhibitor was considered an ESBL producer. We used K. pneumoniae ATCC 700603 and E. coli ATCC 25922 for quality control.
ESBL production was also confirmed by the Phenotypic Confirmatory Disc Diffusion Test (PCDDT) [22]. Briefly, 0.5 McFarland of bacterium suspension was inoculated on MHA plate (Oxoid, United Kingdom) and allowed to air dry for 3 min. Antibiotics disks, including cefotaxime (30 µg), ceftazidime (30 µg), cefotaxime/clavulanic acid (30 µg/10 µg), and ceftazidime/clavulanic acid (30 µg/10 µg), were placed on the inoculated MHA plate, at a 30 mm (mm) radius to radius distance. The plates were incubated overnight in ambient air at 37 ℃. ESBL production was confirmed by observing an isolate with a > 5 mm-clear zone formed between the third-generation cephalosporin and the β-lactam inhibitor. Klebsiella pneumoniae ATCC 700603 and Escherichia coli ATCC 25922 were the control strains.
Detection of ESBL resistance genes
ESBL producers were PCR-screened for SHV-, TEM-, OXA-1, and CTX-M- type ESBL genes using primers in Table 1. We extracted bacterial DNA using the heat lysis method [20] and followed the PCR protocol described by Kiiru et al. [22]. Briefly, 2 μl of DNA was added to 22 μl of PCR master mix (Bio-Rad Laboratories, Hercules, USA) with the target ESBL gene primers and loaded to a thermal cycler (Bio-Rad Laboratories, Hercules, USA), programmed as follows: denaturation at 95 °C for 6 min, annealing at 55 °C for 2 min, and final extension at 72 °C for 10 min. The amplification products were separated by gel electrophoresis (1.5% agar rose gel), stained with SYBR green dye and captured images by the Bio-Rad imaging system (Bio-Rad Laboratories, Hercules, USA). Klebsiella pneumoniae ATCC 700603 and Escherichia coli ATCC 25922 were the quality control organisms.
Antimicrobial susceptibility testing
Further, we characterized the ESBL producers based on antimicrobial susceptibility to meropenem (MEM), colistin (COL), gentamicin (GEN), amikacin (AMK), aztreonam (ATM), ceftazidime (CAZ), cefotaxime (CTX), ceftriaxone (CRO), cefepime (FEP), piperacillin/tazobactam (TZP), amoxicillin/clavulanic acid (AMC), ampicillin/sulbactam (SAM), ciprofloxacin (CIP), and trimethoprim/sulfamethoxazole (SXT). We used VITEK 2 COMPACT system (bioMérieux, Marcy l'Etoile, France) to determine the isolates' antimicrobial susceptibility profile using Card AST GN 83, except for colistin and antibiogram interpreted based on CLSI (2021) guidelines [21]. Quality control organisms, Pseudomonas aeruginosa (ATCC 27853) and Escherichia coli (ATCC 25922), were used.
Colistin susceptibility testing was done by the Simple Disk diffusion method [25]. Using Escherichia coli ATCC 25922 and P. aeruginosa ATCC 27853 as quality control strains, we placed a 10 mg colistin disk on a 0.5 McFarland-equivalent bacterium suspension plated on modified Mueller–Hinton agar 30% (5.1 g/L) (Oxoid, United Kingdom), followed by an overnight incubation at 35 ℃ in 5% CO2. Minimum Inhibitory Concentrations (MICs), determined by broth microdilution following CLSI guidelines [21], were used to interpret the resultant inhibition zones. We defined multidrug-resistant organisms (MDRs) by resistance to three or more antibiotic classes [26].
Data analysis and presentation
Statistical analysis was two-sided using STATA version 16. After describing continuous data in means and medians and categorical data in frequencies and percentages, we presented the data in tables and figures. Crude odds ratio (cOR) was analyzed using binary logic regression, with variables giving p-values ≤ 0.2 further computed by multiple regression analysis for adjusted odd ratio (aOR). The level of statistical significance was set at p-value ≤ 0.05, with a 95% Confidence Interval (CI), and statistically significant associations indicated in bold (Table 4).
Results
Demographic and clinical characteristics of patients with GNB infections
In this study, 49 (40.8%) of 120 severely ill COVID-19 patients had GNB infections. Of these, the majority were: adults aged above 60 years (36.7%), males (53.1%), married (73.5%), not vaccinated against COVID-19 (77.6%), comorbid (77.6%), and discharged (77.6%) after hospitalization for 6–10 days (51%), Table 2.
Spectrum of ESBL-producing GNB isolates in COVID-19 patients admitted in KNH-IDU
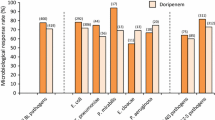
In this study, 49 out of 120 patients had GNB infections (40.8%). Of these, 33 (67.3%) were caused by ESBL – producers, predominated by Klebsiella pneumoniae (45.5%), Enterobacter cloacae complex (21.2%), Acinetobacter baumannii (12.1%), Escherichia coli (9.1%), Pseudomonas aeruginosa (6.1%) and Enterobacter cloacae (6.1%) Fig. 1. All Enterobacter cloacae complex (100%, 7/7) isolates were ESBL producers, whilst the other GNB isolates, including Proteus mirabilis, Acinetobacter calcoaceticus, Serratia marcescens and Stenotrophomonas maltophilia, were all non-ESBL producers Fig. 1.

Spectrum of ESBL-producing GNB isolates in COVID-19 patients admitted in KNH-IDU. ESBL-p extended-spectrum beta-lactamases-producing, non-ESBL-p non-extended-spectrum beta-lactamases-producing
AMR genes carriage in ESBL-producing GNB isolates from COVID-19 patients admitted in KNH-IDU
We determined the occurrence of the most common ESBL genes, including CTX-M-type, TEM-type, SHV-type, and OXA-1, among the ESBL-producing bacterial isolates. All these genes were present in Klebsiella pneumoniae, predominated by CTX-M-type (60.9%, 14/23), and except for isolate 3OP harboring SHV gene only, other isolates encoded CTX-M-type genes, Fig. 2a. In Enterobacter cloacae complex, CTX-M-type and TEM were the principal ESBL genes (75%, 12/16), and similar to K. pneumoniae, AMR genes were present in all the ESBL-producing isolates, with OXA-1 genes as the minority, Fig. 2b. Fifty per cent (1/2) of ESBL-producing- Enterobacter cloacae isolates harbored a single ESBL gene, TEM, (Fig. 2c). About 67% (2/3) of ESBL-producing E. coli isolates encoded AMR genes targeted in this study, except OXA-type (Fig. 2d), whilst 40% (2/5) of ESBL producing- Pseudomonas aeruginosa isolates carried a single ESBL gene, CTX-M-type, Fig. 2e.

Distribution AMR genes among ESBL-producing GNB isolates from COVID-19 patients admitted in KNH-IDU. a Klebsiella pneumoniae. b Enterobacter cloacae complex. c Enterobacter cloacae. d Escherichia coli. e Pseudomonas aeruginosa. f ESBL gene carriage among all isolates. TEM TEM-type β-lactamase genes, CTX-M CTX-M-type β-lactamase genes, SHV SHV-type β-lactamase genes, OXA-1 Oxacillinases -type β-lactamases 1, ESBL Extended Spectrum Beta Lactamase
In general, 73% (24/33) of the ESBL-producing bacteria carried our study’s target AMR genes, mostly CTX-M-type (59%, 17/29), Fig. 2f. Klebsiella pneumoniae isolates harbored the majority of the study ESBL genes identified (76.9%, 20/26), and none of our target ESBL gene was detected in ESBL-producing Acinetobacter baumannii, Fig. 2f.
Distribution of resistance genes among MDR phenotypes of ESBL-producing GNB
Isolates resistant to three or more antibiotic classes were considered multidrug-resistant organisms (MDRs) [19]. Eighty-three per cent of GNB harbouring TEM-type (10/12) and SHV-type (100%, 7/7) β-lactamases showed ESBL phenotypes and inhibitor resistance, mainly involving clavulanate but most of them remained susceptible to tazobactam (60%, 6/10), Table 3. One of TEM-type β-lactamases (8.3%, 1/12) from E. cloacae complex isolate (410P/NP) seemed to efficiently hydrolyze aztreonam (ATM) than cefotaxime (CTX) or ceftazidime (CAZ) Table 3.
In this study, SHV-type genes carrying ESBL producers showed resistance to both CTX and CAZ (K. pneumoniae isolate, 3OP), CAZ (E. coli, 40BD) or CTX (E. cloacae complex (50OP) and K. pneumoniae (36OP and 44 NP). About 13% (3/24) of bacterial isolates encoding CTX-M-type β-lactamases did not display the typical cefotaxime/ceftriaxone (CTX/CRO) resistance phenotype of the early CTX-M variants. About 42% (5/12) of ESBL-producing GNB with CTX-M-type β-lactamases only hydrolyzed ceftazidime (CAZ), Table 3.
The OXA-1 β-lactamases detected were resistant to third (cefotaxime, CTX; ceftazidime, CAZ; ceftriaxone, CRO) fourth-generation cephalosporins (cefepime, FEP) and monobactam (aztreonam, ATM). One of OXA-type b-lactamases isolated from E. cloacae complex isolates (55OP) showed carbapenemases activity, Table 3.
Factors associated with ESBL-producing GNB infections among COVID-19 patients admitted in KNH-IDU
Multivariable analysis established that severely ill COVID-19 patients with comorbidities were about ten (10) times more likely to have an infection caused by ESBL-producing GNB (aOR = 9.86, 95%CI 1.30 – 74.63, p = 0.003). Male gender was also a risk factor for infection with ESBL-producing GNB, although there was no independent association (cOR = 9.97(2.32–42.85), p = 0.002), Table 4.
Discussion
In this study, we screened 49 gram negative bacterial (GNB) isolates for ESBL production. Of these, 67.3% were ESBL – producers, predominated by K. pneumoniae (30.6%). Contrary to our findings, Lemenand and colleagues reported a decreasing proportion of ESBL among E. coli infections (2.9%) during the COVID-19 pandemic in France [27]. The study by Lemenand et al. focused only on single bacteria, E. coli, and their data might not be generalizable to countries differently impacted by the COVID-19 pandemic. Karataş et al. observed a significant decrease in ESBL-P Enterobacterales during the pandemic period compared to the pre-pandemic era [28]. In the study by Karatas and others, not all participants were COVID-19 patients. Different from Karatas et al., we targeted all GNB in severely ill COVID-19 patients confirmed by real-time reverse transcription and quantitative polymerase chain reaction (RT-qPCR), and admitted in critical care unit. This could possibly explain the high prevalence of ESBL producer isolates in our study. In the current study, the prevalence of ESBL-producing GNB infections among COVID-19 patients was higher than that reported in non-COVID-19 patients in East African (42%) and Kenya (47%) [29]. In a recent study among Kenyan children at the point of hospital discharge, the prevalence of ESBL-producing E. coli was 44.3% [30]. Together, these reports suggest a higher prevalence of ESBL-producing GNB in severely ill COVID-19 patients admitted in ICU in our setting.
Klebsiella pneumoniae (30.6%) was the predominant ESBL producer among GNB isolates from severely ill COVID-19 patients. Even though data on ESBL-producing bacteria in COVID-19 patients is limited, in the general population, E. coli and K. pneumoniae [31,32,33,34] seems to be the most common ESBL producers. Our finding may infer similarity in ESBL-producing bacteria profiles among COVID-19 and non-COVID patients. In the hospital environment, GNB can acquire and transfer ESBL genes via mobile genetic elements such as plasmids and transposons, and predominating ESBL producers may vary geographically depending on environmental sanitary status, adherence to infection prevention and control protocols and antimicrobial stewardship policies [35, 36].
To decipher the antimicrobial resistance (AMR), AMR gene carriage among ESBL phenotypes, the commonly reported ESBL genes in bacteria, (CTX-M- type, TEM, SHV and OXA-1) were detected by PCR. About 73% of the ESBL-producing GNB encoded our target AMR genes, mostly CTX-M-type (59%, 17/29), with K. pneumoniae harbouring the majority (76.9%) of the genes. Emeraud et al. documented a nosocomial outbreak of ESBL producing K. pneumoniae carrying CTX-M-15 in a French intensive care unit dedicated to COVID-19 during the first wave of the pandemic [37], however, information on ESBL gene carriage among ESBL producing GNB that cause infections in COVID-19 patients is limited. Before the year 2000, SHV- and TEM–type enzymes were the most predominant ESBLs worldwide [38] but have since been outnumbered by CTX-M ESBLs in non-COVID19 patients [39, 40, 11, 34]. Therefore, our findings suggest a similar ESBL gene carriage among bacterial isolates from COVID-19 patients and the general population. Clinically, the CTX-M-producing bacterial infections are treated using carbapenems, thus promoting the spread of potentially untreatable carbapenemase-producing bacterial infections [38].
In this work, 83% of TEM-type and SHV-type (100%) β-lactamases showed ESBLs phenotypes and inhibitor resistance, mainly involving clavulanate, but mostly remained susceptible to tazobactam (60%, 6/10). Inhibitor-resistant variants emerge following mutations that result in one, two or three amino acid substitutions in the parental enzymes [40]. These mutations confer resistance to clavulanate and sulbactam but not tazobactam and avibactam [11, 41,42,43]. TEM-type β-lactamases (8.3%) from E. cloacae complex isolate, 410P/NP, seemed to efficiently hydrolyze aztreonam (ATM) than cefotaxime (CTX) or ceftazidime (CAZ), a phenotype that was reported in TEM-184 with Q6K, E104K, I127V, R164S and M182T amino acid substitutions [44, 45]. SHV-type ESBL producers showed resistance to both CTX and CAZ (K. pneumoniae isolate, 3OP), CAZ (E. coli, 40BD) or CTX (E. cloacae complex (50OP) and K. pneumoniae (36OP and 44 NP).
SHV-type enzymes mutations that result in the substitution of lysine (Lys238) with serine (Ser) and lysine (Lys240) with glutamic acid (Glu) play a critical role in the efficient hydrolysis of ceftazidime and cefotaxime, respectively [11]. About 87% (3/24) of isolates harboring CTX-M-type β-lactamases in our study displayed the typical cefotaxime/ceftriaxone (CTX/CRO) resistance phenotype observed with the early CTX-M variants [11], and about 42% of the isolates showed ceftazidime (CAZ) resistant phenotypes. The CAZ- resistance has been reported in CTX-M-15 [46, 47] and CTX-M-27 variants [48].
In our study, the OXA-1 β-lactamases detected were resistant to third (cefotaxime, CTX; ceftazidime, CAZ; ceftriaxone, CRO) and fourth (cefepime, FEP)-generation cephalosporins and monobactam (aztreonam, ATM). One of OXA-1 b-lactamases isolated from E. cloacae complex isolates (55OP) had carbapenemase activity. These enzymes are known to have hydrolytic activity against penicillins and cephalosporins, including third-and/or fourth-generations [49]. OXA-1/OXA-30 [50, 51] and OXA-31 [52] variants is associated with FEP resistance. OXA-48 derivative, OXA-163 and OXA-405, OXA-58, OXA-143, and OXA-235 [52] have carbapenemase activity. OXA-48 β-lactamases mediating carbapenem resistance among ESBL-producing Escherichia coli and Klebsiella pneumoniae isolates were describe in a Turkish university hospital [53]. Though we did not elucidate molecular variants of the ESBL genes, these published reports suggest the possible inhibitor-resistance mechanisms among bacterial isolates in our study.
Severely ill COVID-19 patients with comorbidities were at higher risk of infection by ESBL-producing bacteria. Greco and others found that COVID-19 patients with comorbidities, such as diabetes mellitus and hypertension, were at increased risk of co-infections in Italy [54]. In a multi-centre study by He and others on clinical characteristics of COVID-19 patients with clinically diagnosed bacterial co-infection, patients with cardiovascular comorbidities were more likely to have clinically diagnosed bacterial co-infection [55]. In the current study, the most common comorbidities were cancer (17%), kidney disease (16%), diabetes (14.9%), hypertension (11.7%), haematological disorders (7.4%) and HIV/AIDS (6.4%).
This study has some limitations. As a single centre study, the data obtained may not be generalizable to other hospitals within our locality and therefore, a larger study is recommended to determine this epidemiology against the general patient population. Additionally, the purposive sampling may have subjected it to selection bias, and due to resource constrains, we were unable to elucidate the molecular variants of the ESBL genes detected. However, this study highlights the need for systematic and continuous surveillance of multidrug-resistant bacteria among SARS-CoV-2 infected persons in the hospital to inform AMR prevention interventions in line with national and global action plans.
Conclusion
We report a high prevalence of ESBL-GNB infections in severely ill COVID-19 patients, predominantly due to Klebsiella pneumoniae harbouring CTX-M type ESBL genes. The patient’s underlying comorbidities increased the risk of ESBL-producing GNB infection. In COVID-19 pandemic, enhanced systematic and continuous surveillance of ESBL-producing GNB, strict adherence to infection control measures and antimicrobial stewardship policies are warranted in the current study setting.
Availability of data and materials
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- AMR:
-
Antimicrobial Resistance
- CTX-M:
-
Cefotaxime-hydrolysing β-lactamase isolated in Munich
- ESBL:
-
Extended Spectrum β-Lactamases
- MDR:
-
Multidrug Resistant
- OXA:
-
Oxacillinases
- SARS-CoV-2:
-
Severe Acute Respiratory Syndrome-Corona Virus 2
- SHV:
-
Sulfhydryl reagent variable
- TEM:
-
Temoneira, the patient infected with the first isolate expressing TEM-1
References
Gu W, et al. The molecular mechanism of SARS-CoV-2 evading host antiviral innate immunity. Virol J. 2022;19(1):49.
Deinhardt-Emmer S, et al. SARS-CoV-2 causes severe epithelial inflammation and barrier dysfunction. J Virol. 2021;95(10):e00110-e121.
Cyprian F, et al. SARS-CoV-2 and immune-microbiome interactions: lessons from respiratory viral infections. Int J Infect Dis. 2021;105:540–50.
Sahu C, et al. Bacterial coinfections in COVID: Prevalence, antibiotic sensitivity patterns and clinical outcomes from a tertiary institute of Northern India. J Fam Med Prim care. 2022;11(8):4473–8.
Alqahtani A, et al. Bacterial coinfections increase mortality of severely Ill COVID-19 patients in Saudi Arabia. Int J Environ Res Public Health. 2022;19(4):2424.
Cogliati Dezza F, et al. Clinical impact of COVID-19 on multi-drug-resistant gram-negative bacilli bloodstream infections in an intensive care unit setting: two pandemics compared. Antibiotics. 2022;11(7):926.
Mai HTT, Espinoza JL. The impact of COVID-19 pandemic on ESBL-producing enterobacterales infections: a scoping review. Antibiotics. 2023;12(6):1064.
Eaton L. Antimicrobial use in the pandemic. Bull World Heal Organ. 2022;100(5):298–9.
Aurilio C, et al. Mechanisms of action of carbapenem resistance. Antibiotics. 2022;11(3):421.
León-Sampedro R, et al. Pervasive transmission of a carbapenem resistance plasmid in the gut microbiota of hospitalized patients. Nat Microbiol. 2021;6(5):606–16.
Castanheira M, Simner PJ, Bradford PA. Extended-spectrum β-lactamases: an update on their characteristics, epidemiology and detection. JAC-antimicrobial Resist. 2021;3(3):dlab092.
Yang Y-Q, et al. Co-occurrence of mcr-1 and ESBL on a single plasmid in Salmonella enterica. J Antimicrob Chemother. 2016;71(8):2336–8.
Caspar Y, et al. mcr-1 colistin resistance in ESBL-producing Klebsiella pneumoniae, France. Emerg Infect Dis. 2017;23(5):874.
Wangkheimayum J, et al. Occurrence of diverse aminoglycoside modifying enzymes with co-existing extended-spectrum-β-lactamases within Enterobacteriaceae isolated in India. J Glob Antimicrob Resist. 2020;21:369–74.
Salah FD, et al. Distribution of quinolone resistance gene (qnr) in ESBL-producing Escherichia coli and Klebsiella spp. in Lomé, Togo. Antimicrob Resist Infect Control. 2019;8:1–8.
Canton R, Morosini MI, Martin O, De la Maza S, De La Pedrosa EGG. IRT and CMT β-lactamases and inhibitor resistance. Clin Microbiol Infect. 2008;14:53–62.
Moirongo RM, et al. Regional variation of extended-spectrum beta-lactamase (ESBL)-producing Enterobacterales, fluoroquinolone-resistant Salmonella enterica and methicillin-resistant Staphylococcus aureus among febrile patients in sub-Saharan Africa. Front Microbiol. 2020;11: 567235.
Del Sole F, et al. Features of severe COVID‐19: a systematic review and meta‐analysis. Eur J Clin Invest, 2020;50(10), e13378.
WHO. Clinical care severe acute respiratory infection. Toolkit: COVID-19 adaptation. Geneva: World Healt Organisation; 2022.
Swaminathan S, Balaji V, Parija SC, Kapil A, Gautam V, Ray P. Standard operating procedures bacteriology antimicrobial resistance surveillance and research network. New Delhi: ICMR; 2015.
Weistein M, et al. M100 Performance Standards for Antimicrobial. Wayne: Clinical Laboratory Standard Institute; 2021.
Kiiru J, Kariuki S, Goddeeris BM, Butaye P. Analysis of β-lactamase phenotypes and carriage of selected β-lactamase genes among Escherichia coli strains obtained from Kenyan patients during an 18-year period. BMC Microbiol. 2012;12(1):1–11.
Wanja FK, Omwenga EO, Ngugi CW, Maina JN, Kiiru JN. Genotypic characterization of urinary tract infections causing bacteria isolates among adults at Kiambu level 5 hospital, Kenya: selected extended spectrum β-lactamase genes and biofilm formation. MedRxiv. 2022;2(5):1.
Maina J, Ndung’u P, Muigai A, Kiiru J. Antimicrobial resistance profiles and genetic basis of resistance among non-fastidious gram-negative bacteria recovered from ready-to-eat foods in Kibera informal housing in Nairobi. Kenya Access Microbiol. 2021;3(6):000236. https://doi.org/10.1099/acmi.0.000236.
Uwizeyimana JD, Kim D, Lee H, Byun JH, Yong D. Determination of colistin resistance by simple disk diffusion test using modified Mueller-Hinton agar. Ann Lab Med. 2020;40(4):306–11. https://doi.org/10.3343/alm.2020.40.4.306.
Wang M, et al. Analysis of multidrug-resistant bacteria in 3223 patients with hospital-acquired infections (HAI) from a tertiary general hospital in China. Bosn J basic Med Sci. 2019;19(1):86.
Lemenand O, Coeffic T, Thibaut S, Cotinat MC, Caillon J, Birgand G. Decreasing proportion of extended-spectrum beta-lactamase among E. coli infections during the COVID-19 pandemic in France. J Infect. 2021;83(6):664–70.
Karataş M, Yaşar-Duman M, Tünger A, Çilli F, Aydemir Ş, Özenci V. Secondary bacterial infections and antimicrobial resistance in COVID-19: comparative evaluation of pre-pandemic and pandemic-era, a retrospective single center study. Ann Clin Microbiol Antimicrob. 2021;20:1–8.
Sonda T, et al. Meta-analysis of proportion estimates of Extended-Spectrum-Beta-Lactamase-producing Enterobacteriaceae in East Africa hospitals. Antimicrob Resist Infect Control. 2016;5(1):1–9.
Tornberg-Belanger SN, et al. Antimicrobial resistance including extended spectrum beta lactamases (ESBL) among E. coli isolated from kenyan children at hospital discharge. PLoS Negl Trop Dis. 2022;16(3):e0010283. https://doi.org/10.1371/journal.pntd.0010283.
Diriba K, Awulachew E, Gemede A, Anja A. The magnitude of extended-spectrum beta-lactamase-producing Enterobacteriaceae from clinical samples in Ethiopia: a systematic review and meta-analysis. Access Microbiol. 2021;3(3):000195.
Kateregga JN, Kantume R, Atuhaire C, Lubowa MN, Ndukui JG. Phenotypic expression and prevalence of ESBL-producing Enterobacteriaceae in samples collected from patients in various wards of Mulago Hospital, Uganda. BMC Pharmacol Toxicol. 2015;16:1–6.
Alsamawi M, et al. Epidemiology of extended-spectrum beta-lactamase producing Enterobacteriaceae in Qatar: a three-year hospital-based study. Front Antibiot. 2022;7(1):980686.
Li Z, et al. High Carriage of Extended-Spectrum, Beta Lactamase-Producing, and Colistin-Resistant Enterobacteriaceae in Tibetan Outpatients with Diarrhea. Antibiotics. 2022;11(4):508.
Larsson DGJ, Flach C-F. Antibiotic resistance in the environment. Nat Rev Microbiol. 2022;20(5):257–69.
Aleem A, Samad ABA, Slenker AK. Emerging variants of SARS-CoV-2 and novel therapeutics against coronavirus (COVID-19). Treasure Island: StatPearls Publishing; 2021.
Emeraud C, et al. Outbreak of CTX-M-15 extended-spectrum β-lactamase-producing Klebsiella pneumoniae ST394 in a French intensive care unit dedicated to COVID-19. Pathog. 2021;10(11):1426. https://doi.org/10.3390/pathogens10111426.
Bevan ER, Jones AM, Hawkey PM. Global epidemiology of CTX-M β-lactamases: temporal and geographical shifts in genotype. J Antimicrob Chemother. 2017;72(8):2145–55.
Tacconelli E, et al. Analysis of the challenges in implementing guidelines to prevent the spread of multidrug-resistant gram-negatives in Europe. BMJ Open. 2019;9(5): e027683.
Gutiérrez-Gutiérrez B, Rodríguez-Baño J. Current options for the treatment of infections due to extended-spectrum beta-lactamase-producing Enterobacteriaceae in different groups of patients. Clin Microbiol Infect. 2019;25(8):932–42.
Russ D, et al. Escape mutations circumvent a tradeoff between resistance to a beta-lactam and resistance to a beta-lactamase inhibitor. Nat Commun. 2020;11(1):2029.
Chaibi EB, Sirot D, Paul G, Labia R. Inhibitor-resistant TEM β-lactamases: phenotypic, genetic and biochemical characteristics. J Antimicrob Chemother. 1999;43(4):447–58.
Drawz SM, Bonomo RA. Three decades of β-lactamase inhibitors. Clin Microbiol Rev. 2010;23(1):160–201.
Piccirilli A, et al. TEM-184, a novel TEM-derived extended-spectrum β-lactamase with enhanced activity against aztreonam. Antimicrob Agents Chemother. 2018;62(9):e00688-e718.
Piccirilli G, et al. Infectious meningitis/encephalitis: evaluation of a rapid and fully automated multiplex PCR in the microbiological diagnostic workup. New Microbiol. 2018;41(2):118–25.
Poirel L, Gniadkowski M, Nordmann P. Biochemical analysis of the ceftazidime-hydrolysing extended-spectrum β-lactamase CTX-M-15 and of its structurally related β-lactamase CTX-M-3. J Antimicrob Chemother. 2002;50(6):1031–4.
Poirel L, de la Rosa J-MO, Richard A, Aires-de-Sousa M, Nordmann P. CTX-M-33 is a CTX-M-15 derivative conferring reduced susceptibility to carbapenems. Antimicrob Agents Chemother. 2019;63(12):e01515-e1519.
Bonnet R, et al. Effect of D240G substitution in a novel ESBL CTX-M-27. J Antimicrob Chemother. 2003;52(1):29–35.
Yoon E-J, Jeong SH. Class D β-lactamases. J Antimicrob Chemother. 2021;76(4):836–64.
Dubois V, Arpin C, Quentin C, Texier-Maugein J, Poirel L, Nordmann P. Decreased susceptibility to cefepime in a clinical strain of Escherichia coli related to plasmid-and integron-encoded OXA-30 β-lactamase. Antimicrob Agents Chemother. 2003;47(7):2380–1.
Beceiro A, et al. False extended-spectrum β-lactamase phenotype in clinical isolates of Escherichia coli associated with increased expression of OXA-1 or TEM-1 penicillinases and loss of porins. J Antimicrob Chemother. 2011;66(9):2006–10.
Aubert D, Poirel L, Chevalier J, Leotard S, Pages J-M, Nordmann P. Oxacillinase-mediated resistance to cefepime and susceptibility to ceftazidime in Pseudomonas aeruginosa. Antimicrob Agents Chemother. 2001;45(6):1615–20.
Nazik H, et al. Carbapenem resistance due to Bla (sub oxa-48) among ESBL-producing Escherichia coli and Klebsiella pneumoniae isolates in a univesity hospital, turkey. Southeast Asian J Trop Med Public Health. 2012;43(5):1178.
Greco R, Panetta V, Della Rocca MT, Durante A, Di Caprio G, Maggi P. Profile of co-infection prevalence and antibiotics use among COVID-19 patients. Pathogens. 2022;11(11):1250.
He S, et al. Clinical characteristics of COVID-19 patients with clinically diagnosed bacterial co-infection: a multi-center study. PLoS ONE. 2021;16(4):e0249668.
Acknowledgements
We acknowledge the valuable work of Mr. Kelvin Nyongesa from KNH Medical Research Department in performing statistical analysis for the data of this study, and we appreciate the technical help provided by Mr. John Maina and Ms. Susan Kiiru both from Centre for Microbiology Research, Kenya Medical Research Institute (CMR-KEMRI).
Funding
This work was partially funded by the Kenyatta National Hospital (KNH) Medical Research and Programs. Ref. No. KNH/R&P/23 K/23/6.
Author information
Authors and Affiliations
Contributions
All authors significantly contributed to the research reported. JMM and AMM conceptualized and designed the study, interpreted the data and drafted the manuscript. JMN supervised the study execution and all laboratory work. JMM performed all laboratory analyses directed by JNM. JMN reviewed the manuscript. All authors read, substantially revised and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The procedures in this study which were involving human materials were approved by the Kenyatta National Hospital-University of Nairobi Ethics and Research Committee (KNH-UoN ERC), Ref KNH-ERC/A/280. We sought consent for patient’s participation in the study through the patient’s guardians or their legally approved representatives.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Mutua, J.M., Njeru, J.M. & Musyoki, A.M. Extended-spectrum β-lactamase- producing gram-negative bacterial infections in severely ill COVID-19 patients admitted in a national referral hospital, Kenya. Ann Clin Microbiol Antimicrob 22, 91 (2023). https://doi.org/10.1186/s12941-023-00641-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12941-023-00641-8