
Abstract
Background
Neonicotinoids are potential alternatives for controlling pyrethroid-resistant mosquitoes, but their efficacy against malaria vector populations of sub-Saharan Africa has yet to be investigated. The aim of the present study was to test the efficacy of four neonicotinoids against adult populations of the sibling species Anopheles gambiae and Anopheles coluzzii sampled along an urban-to-rural gradient.
Methods
The lethal toxicity of three active ingredients for adults of two susceptible Anopheles strains was assessed using concentration–response assays, and their discriminating concentrations were calculated. The discriminating concentrations were then used to test the susceptibility of An. gambiae and An. coluzzii mosquitoes collected from urban, suburban and rural areas of Yaoundé, Cameroon, to acetamiprid, imidacloprid, clothianidin and thiamethoxam.
Results
Lethal concentrations of neonicotinoids were relatively high suggesting that this class of insecticides has low toxicity against Anopheles mosquitoes. Reduced susceptibility to the four neonicotinoids tested was detected in An. gambiae populations collected from rural and suburban areas. By contrast, adults of An. coluzzii that occurred in urbanized settings were susceptible to neonicotinoids except acetamiprid for which 80% mortality was obtained within 72 h of insecticide exposure. The cytochrome inhibitor, piperonyl butoxide (PBO), significantly enhanced the activity of clothianidin and acetamiprid against An. gambiae mosquitoes.
Conclusions
These findings corroborate susceptibility profiles observed in larvae and highlight a significant variation in tolerance to neonicotinoids between An. gambiae and An. coluzzii populations from Yaoundé. Further studies are needed to disentangle the role of exposure to agricultural pesticides and of cross-resistance mechanisms in the development of neonicotinoid resistance in some Anopheles species.
Similar content being viewed by others

Explore related subjects
Find the latest articles, discoveries, and news in related topics.Background
The scale-up of vector control has largely contributed to reducing malaria burden over the last two decades in sub-Saharan Africa [1]. Long-lasting insecticidal nets and indoor residual spraying are the cornerstone of malaria prevention and rely on the use of eight classes of chemical insecticides [2]. Prior to the recent prequalification of formulations based on new active ingredients (i.e., chlorfenapyr, clothianidin, etofenprox and broflanilide) by the World Health Organization (WHO), neurotoxic insecticides that disrupt a sodium channel or inhibit acetylcholinesterase in the insect’s nervous system were widely applied [3]. The similarity of modes of action and the intensive use of a limited number of active ingredients have created ideal conditions for the emergence and spread of resistance [4, 5]. Indeed, resistance to any of the classes of neurotoxic insecticides used in interventions has been reported posing a challenge to the sustainability of vector control [6, 7]. As a result, the search for new insecticides has become an urgent necessity [8, 9]. In the quest for new active ingredients, alternatives to sodium channel and acetylcholinesterase inhibitors have drawn considerable attention because their new modes of action are more suitable for targeting populations that are currently resistant to existing insecticides [10,11,12].
Formulations of two neonicotinoid insecticides repurposed from the agricultural sector (clothianidin and imidacloprid) are prequalified for indoor residual spraying (IRS) and space spraying targeting malaria-carrying mosquitoes [2]. Clothianidin is an active ingredient used alone or in combination with deltamethrin in four new IRS formulations [2, 13,14,15]. Imidacloprid is combined with prallethrin in Cielo ULV®, a formulation manufactured for space spraying. Neonicotinoids act as agonist of acetylcholine, selectively target the insect nicotinic acetylcholine receptor (nAChR) and disrupt excitatory cholinergic neurotransmission leading to paralysis and death [16].
Cross-resistance which occurs when insects develop resistance to several active ingredients that have a similar mode of action is a pervasive driver of tolerance to neonicotinoids in agricultural pests [17,18,19,20]. Importantly, significant research has demonstrated that detoxification enzymes, primarily cytochrome monooxygenases (CYPs), play a key role in the development of neonicotinoid resistance in insect pests [21,22,23,24,25]. Due to chronic exposure to public health and agricultural pesticides and other xenobiotic compounds, anopheline populations from sub-Saharan Africa have evolved an impressive array of detoxification mechanisms including overexpression and duplication of CYPs [26,27,28,29,30]. These enzymes represent a threat to the efficacy of neonicotinoids as vector populations can use preexisting metabolic and excretion mechanisms selected by previous exposure to adapt to new active ingredients.
Agricultural spray of pesticides is also known to be a major factor driving resistance to public health insecticides [31, 32]. Neonicotinoids are intensively used in agriculture and represented more than 25% of the global insecticide sales share in 2014 [33]. In some sub-Saharan African countries, between 100 and 200 formulations of thiacloprid, imidacloprid, acetamiprid and thiamethoxam are registered for agricultural pest management [34]. Neonicotinoids sprayed to protect crops from insect pests are highly water-soluble and are prone to leach in aquatic habitats that are also used as breeding sites by Anopheles larvae in farming areas [35, 36]. This unintentional exposure may contribute to the development of cross-resistance to neonicotinoids in larval populations [37,38,39,40].
To better evaluate the risk of cross-resistance to neonicotinoids in Anopheles mosquitoes, it is vital to assess baseline susceptibility of vector populations to a wide range of neonicotinoid insecticides. Notably, addressing the fine-scale variations in susceptibility between species and geographic areas could provide insights into the candidate environmental and genetic drivers of resistance. Larvae of the sibling species Anopheles gambiae and Anopheles coluzzii from urban and suburban areas of Yaoundé display contrasting patterns of resistance to neonicotinoids [38]. In contrast to An. coluzzii, third instar larvae of An. gambiae can growth and emerge in water containing a concentration of neonicotinoid that kill immature stages of a susceptible strain within 24 h [39]. Therefore, comparing patterns of resistance and susceptibility between populations of the two sibling species could help decipher the key players involved in the development of neonicotinoid resistance in anopheline populations.
The present study aimed to test the efficacy of four agricultural neonicotinoids against An. gambiae and An. coluzzii adult mosquitoes along an urban-to-rural gradient in Yaoundé, Cameroon. The lethal toxicity of three active ingredients was tested, and the susceptibility of wild female adults to clothianidin, acetamiprid, imidacloprid and thiamethoxam assessed. Anopheles coluzzii populations that thrive in urban areas of Yaoundé were globally susceptible to neonicotinoids, while An. gambiae adults collected from rural and sub-urban environments were more tolerant. The potential efficacy of neonicotinoid formulations against malaria mosquitoes could be impacted by fine-scale geographic variations in susceptibility among wild populations as well as the low toxicity of some active ingredients.
Methods
Lethal concentrations determination
Center for Disease Control and prevention (CDC) bottle bioassays were used to assess the lethal toxicity of neonicotinoids [41]. Four active ingredients were tested: acetamiprid, imidacloprid, thiamethoxam and clothianidin. Acetamiprid, imidacloprid and thiamethoxam are commonly used by farmers in Cameroon to protect several types of crops from insect pests [34, 42]. Clothianidin is an agrochemical, which is not registered in Cameroon, but whose formulations have been approved for malaria mosquito control [2]. All four neonicotinoids tested were technical-grade material (Sigma Aldrich, Pestanal®). The insecticides were dissolved in absolute ethanol except imidacloprid for which acetone was used. A range of concentrations of the active ingredient was tested against a susceptible strain (An. gambiae Kisumu or An. coluzzii Ngousso) to determine LC50 and LC99 corresponding to the lowest concentrations required to kill 50% and 99% of susceptible individuals, respectively. By contrast to clothianidin whose toxicity has been tested with at least one susceptible strain [8, 12, 42, 43], information on the lethal concentrations of acetamiprid, imidacloprid and thiamethoxam against African malaria mosquitoes is lacking. The three active ingredients were tested using the following gradients: imidacloprid (12.5, 50, 100, 200 and 250 µg/ml); acetamiprid (12.5, 25, 50, 75 and 150 µg/ml); thiamethoxam (3, 50, 150, 250 and 300 µg/ml).
250-ml Wheaton bottles were coated with 1 ml of a given concentration of the insecticide and 25 female adult mosquitoes, 3 to 5 days old, were exposed for 1 h in the bottles. After the exposure period, mosquitoes were removed from the bottles and released into net-covered paper cups on top of which cotton imbibed with 10% sugar solution was placed. Mortality was observed at 24 h and 72 h, respectively. Bioassays were performed under a controlled environment of 25–27 °C, 70–90% relative humidity and a 12:12 h light/dark photoperiod. Four replicates were tested per concentration together with two controls where mosquitoes were exposed to 1 ml of solvent, ethanol or acetone.
Susceptibility evaluation in wild populations
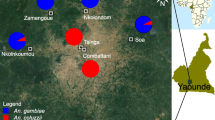
Wild An. gambiae sensu lato (s.l.) (the An. gambiae complex) mosquito populations were collected from several field surveys and tested between September 2019 and September 2022. Mosquito larvae were sampled using a dipper from breeding sites identified in nine locations along an urban-to-rural gradient in Yaoundé. Two locations surveyed within the city were densely urbanized (Combattant, Etoa Meki), while seven sampling sites belonged to suburban and rural areas (Fig. 1). One of the suburban sites (Nkolondom) was situated in a neighbourhood where marshy areas are being used for intensive cultivation of food crops (Fig. 1) [37].

Map of the sampling sites where neonicotinoid susceptibility was monitored in An. gambiae and An. coluzzii adults. The nine sites are located along an urban-to-rural gradient centered on Yaoundé. The city (brown areas) situated in the equatorial zone of Cameroon is surrounded by degraded forests (green areas)
Larvae collected from typical An. gambiae s.l. breeding sites were transported in plastic containers to the insectary where they were identified using reference morphological identification keys [44, 45]. Larvae were reared in trays containing 200 ml borehole water and fed with TetraMin® daily. Adults that emerged were maintained in 30 cm-by-30 cm cages and provided with 10% sugar solution. Previous studies have established that two cryptic species of the An. gambiae complex: An. gambiae sensu stricto (hereafter referred to as An. gambiae) and An. coluzzii segregate along an urbanization gradient in Yaoundé [46, 47]. Anopheles coluzzii is found exclusively in densely urbanized settings, while An. gambiae occurs in sub-urban and rural environments. The two species were identified by subjecting a subset of 50 female adults from each sampling site to a molecular diagnostic method described in [48]. Female adults were tested to assess their level of susceptibility to neonicotinoids using bottle bioassays and a discriminating concentration of the insecticide. The discriminating concentration, defined as the lowest concentration required to kill 100% from a susceptible strain (An. gambiae Kisumu or An. coluzzii Ngousso), was chosen within the confidence intervals of LC99 for acetamiprid, imidacloprid and thiamethoxam. The discriminating concentration of clothianidin was obtained from Agumba et al. [42].
Synergistic effect of piperonyl butoxide
A bioassay was performed to test if the synergistic effect of the cytochrome P450 inhibitor, piperonyl butoxide (PBO) could restore neonicotinoid susceptibility. Female adults that emerged from the same pool of larvae were first exposed to 4% PBO in CDC bottles, for 1 h before being released into other bottles coated with the neonicotinoid insecticide. Susceptibility tests were conducted as described above. Mortality values obtained with or without prior exposure to the synergist were compared after 72 h of holding period.
Data analysis
The mortality rate was calculated as a percentage across all four test bottles. Abbott’s formula was used to correct mortality rates when 5–20% of individuals died in the corresponding control tests [49]. For each insecticide, a log-logistic model were used to fit the concentration–response curve with the drc package in R (version 4.2.2) [50]. A probit model was applied to determine LC50 and LC99 and their 95% confidence intervals for each insecticide using the ecotox package. To compare LC50 values at 24 and 72 h, a ratio test implemented in Wheeler et al. [51] was used. Mortality in wild populations was interpreted based on the WHO criteria which suggest that 98%–100% mortality indicates susceptibility, 90%–97% mortality reflects the possibility of resistance that needs to be confirmed and less than 90% mortality corresponds to resistance [52].
Results
Lethal concentrations of three neonicotinoids against An. gambiae and An. coluzzii
The acute toxicity of three neonicotinoids against Anopheles mosquitoes was tested using concentration–response bioassays with mortality recorded within 24 h and 72 h of exposure. Results of LC50 and LC99 of acetamiprid, thiamethoxam and imidacloprid are presented in Table 1 and Fig. 2. 24-h LC99 obtained with the susceptible strain An. gambiae Kisumu were 69.0 µg/ml, confidence interval CI95% (54.40, 98.1), for acetamiprid compared to 152.0 µg/ml (112.0, 235.1) for imidacloprid. Meanwhile, 24-h lethal concentrations of thiamethoxam detected using the susceptible strain An. coluzzii Ngousso were 9.6 µg/ml (6.8, 13.5) for LC50 and 133.0 µg/ml (79.4, 277.0) for LC99, respectively. Low lethal concentrations reflect high toxicity since smaller doses of insecticide would be required to kill a given number of exposed individuals. Based on the lowest LC50, thiamethoxam [LC50: 9.6 µg/ml (6.8, 13.5)] was the most toxic neonicotinoid to Anopheles mosquitoes within 24 h followed by acetamiprid [LC50: 13.6 µg/ml (11.5, 15.5)] and imidacloprid [LC50: 18.6 µg/ml (15.2, 20.0)]. To compare lethal toxicity within 24 h and 72 h, a ratio test was applied. The toxicity was considered different between 24 and 72 h if there was a significant difference between LC50 values. Based on the ratio test, the difference in LC50 at 24 h and 72 h for the three active ingredients tested was not significant (p > 0.05), which suggested that extending the holding period did not increase toxicity (Table 1).

Concentration–response curves with the standard error of the regression model (grey bands) depicting the 24-h toxicity of three neonicotinoids against Anopheles mosquitoes. LC50 and LC99 with 95% confidence intervals of acetamiprid and imidacloprid were determined using the susceptible strain An. gambiae Kisumu while lethal concentrations of thiamethoxam were estimated with the An. coluzzii Ngousso strain
Variation in susceptibility within and between species
The baseline susceptibility of wild populations to acetamiprid, imidacloprid, and thiamethoxam, was evaluated using discriminating concentrations of 75 µg/ml, 200 µg/ml and 250 µg/ml, respectively. The concentrations were chosen within the 95% confidence intervals for 24-h LC99 with the aim to balance the risk of not detecting low-level resistance while limiting the risk of reporting false positives. In addition to acetamiprid, imidacloprid and thiamethoxam, we tested a fourth neonicotinoid (clothianidin) using a discriminating concentration of 150 µg/ml. The efficacy of the discriminating concentrations against the insecticide susceptible strains An. gambiae Kisumu and An. coluzzii Ngousso was confirmed. For both laboratory strains, 100% of female adults exposed to the discriminating concentration of each of the four neonicotinoids died within 24 h.
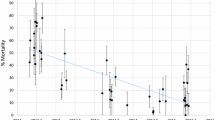
Molecular analyses confirmed that An. gambiae s.l. larvae collected from two sites situated in urbanized areas (Etoa Meki and Combattant) were 100% An. coluzzii. Across two sites located in the sub-urban area (Nkolkoumou and Soa), the relative frequencies of An. gambiae and An. coluzzii were ~ 80% and 20%, respectively. Anopheles gambiae was the only member of the species complex collected from the remaining five sites. Anopheles coluzzii mosquitoes that occurred in urban areas were susceptible to neonicotinoids except acetamiprid for which signs of reduced susceptibility were apparent (Fig. 3). 100% mortality was observed within 72 h of exposure to thiamethoxam, imidacloprid and clothianidin whereas the average 72-h mortality for acetamiprid was 80%. Mosquito populations from Nkolnkoumou were susceptible to imidacloprid and clothianidin and resistant to thiamethoxam and acetamiprid. Samples collected from Soa were tested only with clothianidin and were susceptible. None of the populations from typical An. gambiae habitats (rural/sub-urban areas) were susceptible to acetamiprid or clothianidin. Populations from the agricultural site (Nkolondom) were the least susceptible to neonicotinoids, with mortality rates lower than 50% for acetamiprid and clothianidin. It was also the only site where resistance to all four neonicotinoids was detected.

Percent mortality in An. gambiae and An. coluzzii female adults exposed to four neonicotinoids. Error bars indicate the standard error of the mean and (n) the total number of adult mosquitoes tested from several pools of larvae collected between 2019 and 2022. *Fisher’s exact test indicated a significantly lower mortality in An. gambiae compared to An. coluzzii (p < 0.05)
Using Fisher’s exact test, the susceptibility profiles of wild populations of An. gambiae (n = 3364) and An. coluzzii (n = 754) to the four neonicotinoids was compared. Mortality rates within 72 h of insecticide exposure was significantly lower in An. gambiae compared to An. coluzzii for acetamiprid (p < 2.2E−16), clothianidin (p < 2.2E−16), imidacloprid (p = 1.46E−08) and thiamethoxam (p = 1.86E−12) (Fig. 3).
The susceptibility profiles of populations from the agricultural site (Nkolondom, n = 1793) and those from the rural and sub-urban areas (Soa, Nkolnkoumou, Evogo, Nkolngock, Eyang and Lobo, n = 1571) revealed significant differences. Female adults from Nkolondom exhibited significantly lower mortality compared to rural/sub-urban populations for clothianidin (p < 2.2E−16, Fisher’s exact test), imidacloprid (p = 5.71E−08) and thiamethoxam (p = 3.12E−14), but not for acetamiprid (p = 0.1156) (Fig. 4). While thiamethoxam was the most toxic to Anopheles mosquitoes based on the lowest LC50, imidacloprid was the most effective considering the mortality rate induced in wild populations. Clothianidin was the least effective in An. gambiae populations from rural/sub-urban and agricultural areas.

Mortality rates in An. gambiae female adults exposed to a discriminating concentration of a neonicotinoid. n = sample size, *Fisher’s exact test indicated a significantly lower mortality in the agricultural site compared to rural/sub-urban populations (p < 0.05)
PBO enhances neonicotinoid activity in An. gambiae
Bioassays were carried out to determine if inhibition of cytochromes mediated by PBO could improve the efficacy of neonicotinoids against Anopheles mosquitoes. The synergistic effect of PBO was tested using the agricultural population that was resistant to the four neonicotinoids. Clothianidin, acetamiprid and thiamethoxam were tested because mortality rates within 72 h of exposure were less than 75% allowing a more accurate evaluation of synergism. Susceptibility was fully restored for acetamiprid (100% vs 35.55 ± 5.25%, p < 2.2E−16, Fisher’s exact test) and partially restored for clothianidin (74.33 ± 3.82 vs 30.98 ± 3.49, p < 2.2E−16) in the presence of PBO at 72 h post-exposure (Fig. 5). On the contrary, pre-exposure to PBO reduced mortality to thiamethoxam (58.0% ± 8.2 vs 71.5% ± 7.7, p = 0.04149, Fisher’s exact test).

Effects of piperonyl butoxide (PBO) on the efficacy of three neonicotinoids against resistant An. gambiae mosquitoes. *Mortality was significantly higher in the presence of PBO (p < 0.05, Fisher’s exact test). Black diamond suit: mortality was significantly lower in the presence of PBO (p < 0.05)
Discussion
Most insecticides used in mosquito control come from the agricultural sector [5]. A new generation of active ingredients are being evaluated to control anopheline mosquitoes that have developed resistance to existing vector control insecticides [8, 9]. In the present study, neonicotinoids, a class of insecticides including clothianidin and imidacloprid whose formulations are proposed for indoor residual spraying and space spraying were tested [13,14,15].
The lethal concentrations of three different neonicotinoids were determined using two susceptible laboratory strains. 24-h lethal concentration indicated that thiamethoxam was the most toxic neonicotinoid to anopheline mosquitoes as it had the lowest LC50, followed by acetamiprid [42]. However, the neonicotinoids tested relatively low toxicity against Anopheles mosquitoes based on LC values. For example, in comparison with the pyrethroid deltamethrin, the 24-h LC99 of the most potent neonicotinoid was approximately sevenfold lower [41] A large-scale screening conducted to search for candidate pesticides that could be used for malaria vector control revealed that three neonicotinoids were among the least active insecticides against adults of Aedes aegypti and Anopheles stephensi in a list of nearly 100 compounds tested [9]. LC80 of > 200 µg/ml was obtained in 24 h when an insecticide-susceptible strain of An. stephensi was exposed to imidacloprid or thiamethoxam while LC80 for acetamiprid was ~ 20 µg/ml [9]. In the current study, it was also noted that extending the holding period post-exposure did not improve the toxicity of neonicotinoids as there was no significant difference between LC50 at 24 h and 72 h. Two laboratory strains (An. gambiae Kisumu and An. coluzzii Ngousso) were used to determine the discrimination concentrations that were applied to evaluate susceptibility in wild populations. Although the values detected provided sufficient discriminating power, a more comprehensive survey involving multiple laboratory colonies is still needed to establish guidelines on neonicotinoid susceptibility testing in malaria vectors [43, 53].
This study revealed a reduction in susceptibility to neonicotinoids in wild anopheline mosquito populations. Contrasting patterns of baseline susceptibility to neonicotinoids between the two sibling species An. gambiae and An. coluzzii were observed. These results corroborated findings from earlier surveys indicating that clothianidin resistance is emerging in An. gambiae [38]. In contrast to clothianidin which is not registered for crop protection, formulations of acetamiprid and imidacloprid are intensively used in agricultural areas in some African countries including Cameroon [34, 38, 54]. Agricultural pesticides have historically play a key role in the spread of resistance to insecticides used in public health programmes among malaria-carrying mosquito species [31, 55]. As neonicotinoids are highly soluble in water and are likely to leach in aquatic habitats, it has been suggested that exposure of larval populations to pesticide residues may contribute to resistance selection [35, 55]. Indeed, a complimentary study has revealed that An. gambiae larvae collected from rural and suburban areas of Yaoundé displayed high fitness when reared in water containing concentrations of neonicotinoids that were lethal to susceptible strains [38]. For instance, at least 40% of larvae collected from Nkolondom and Nkolnkoumou were able to survive and emerge in water containing a lethal concentration of acetamiprid or imidacloprid [39]. Although the environmental and genetic bases of neonicotinoid tolerance in female adult mosquitoes have yet to be investigated, the present study suggests that selection at larval stage could be a driver of cross-resistance to several active ingredients [38,39,40]. Anopheles coluzzii populations collected from urbanized areas of Yaoundé had sub-optimal mortality (~ 80%) to acetamiprid, but this species was generally susceptible to neonicotinoids. This finding is also consistent with larval tests, which showed that third instars of An. coluzzii from Yaoundé had low survival and barely emerged in water containing a lethal concentration of a neonicotinoid [38]. However, in Ivory Coast, reduced susceptibility to acetamiprid and imidacloprid has been reported in adults of An. coluzzii sampled from agricultural areas suggesting that populations of this species can develop resistance to neonicotinoid [37].
Although our data show that neonicotinoid resistance was stronger among populations collected from an agricultural site, it is premature to establish a link between residual pesticide exposure in farming areas and the development of resistance. The role of cross-resistance mechanisms notably the contribution of agricultural pesticides and of detoxification enzymes on resistance selection has yet to be elucidated. In this study, it was noted that pre-exposure of resistant Anopheles mosquitoes to PBO restored their susceptibility to acetamiprid suggesting that cytochrome P450 monooxygenases (CYPs) are involved in resistance to this insecticide. Indeed, overexpression of CYPs is an important mechanism underlying neonicotinoid resistance in a variety of insect pests including mosquitoes, white fly and aphids [22, 38, 56].
Gene expression analysis revealed overexpression of multiple CYP genes in an acetamiprid-resistant strain of the melon aphid, Aphis gossypii, indicating a role of P450-mediated detoxification in acetamiprid resistance [57]. Acetamiprid resistance has also been shown to depend strongly on monooxygenases in white flies and Aedes mosquitoes [23, 24]. Consistent with past surveys, the current study confirms that pre-exposure to PBO also drastically improve the efficacy of clothianidin against Anopheles mosquitoes [38]. Conversely, PBO had a slightly negative effect on the activity of thiamethoxam in wild populations. This could be explained by the fact that thiamethoxam is a pro-insecticide which needs to be converted to clothianidin before being active in insects. This conversion is catalyzed by enzymes which might have been inhibited by PBO, leading to a slight reduction in insecticidal activity when mosquitoes were pre-exposed to PBO [58].
Some surfactants have been shown to enhance the efficacy of neonicotinoids against malaria vectors [8, 37, 40, 59]. This synergistic action could be harnessed to improve neonicotinoid formulations that may be used vector control. In the present study, it has been observed that PBO is a synergist of neonicotinoids, which offers additional options to enhance the potency of some active ingredients against Anopheles mosquitoes.
The contrasting pattern of susceptibility observed between An. gambiae and An. coluzzii from Yaoundé highlights the need for a more comprehensive survey of tolerance to neonicotinoids in malaria vectors species [60]. The vectorial system in sub-Saharan Africa comprises at least a dozen major species that occupy diverse niches at larval and adult stages [61, 62]. The results show that the susceptibility profile could vary significantly within and between Anopheles species, even on a small geographic scale. Species including An. gambiae, An. coluzzii and Anopheles arabiensis whose larvae are more likely to be exposed to neonicotinoid residues in man-made habitats such as temporary breeding sites created in farms should be the focus of extensive monitoring efforts [63, 64]. Even if Anopheles funestus larval habitats are less prone to anthropogenic disturbance, populations of this species have developed strong pesticide resistance based on overexpression of detoxification enzymes [27, 65, 66]. Preselected enzymes can contribute to the detoxification of some neonicotinoid insecticides and to the emergence of resistance in An. funestus [66].
Conclusions
Although neonicotinoids have low acute toxicity and reduced efficacy in some Anopheles mosquito populations, the use of more potent formulations can still provide alternatives for controlling pyrethroid-resistant malaria mosquitoes. The current study and complimentary investigations revealed that synergists and adjuvants such as PBO and surfactants could be used to enhance the efficacy of neonicotinoid insecticides against Anopheles mosquitoes.
Availability of data and materials
The data for this study have been presented within this article.
References
WHO. World malaria report 2022. Geneva: World Health Organization; 2022.
WHO. List of WHO prequalified vector control products. Geneva: World Health Organization; 2023. https://extranet.who.int/pqweb/vector-control-products/prequalified-product-list.
Hemingway J, Ranson H. Insecticide resistance in insect vectors of human disease. Annu Rev Entomol. 2000;45:371–91.
Ranson H, Lissenden N. Insecticide resistance in African Anopheles mosquitoes: a worsening situation that needs urgent action to maintain malaria control. Trends Parasitol. 2016;32:187–96.
Hemingway J. The way forward for vector control. Science. 2017;358:998–9.
Hemingway J, Ranson H, Magill A, Kolaczinski J, Fornadel C, Gimnig J, et al. Averting a malaria disaster: will insecticide resistance derail malaria control? Lancet. 2016;387:1785–8.
Strode C, Donegan S, Garner P, Enayati AA, Hemingway J. The impact of pyrethroid resistance on the efficacy of insecticide-treated bed nets against African Anopheline mosquitoes: systematic review and meta-analysis. PLoS Med. 2014;11: e1001619.
Lees R, Praulins G, Davies R, Brown F, Parsons G, White A, et al. A testing cascade to identify repurposed insecticides for next-generation vector control tools: screening a panel of chemistries with novel modes of action against a malaria vector. Gates Open Res. 2019;3:1464.
Hoppé M, Hueter OF, Bywater A, Wege P, Maienfisch P. Evaluation of commercial agrochemicals as new tools for malaria vector control. Chimia (Aarau). 2016;70:721–9.
N’Guessan R, Boko P, Akogbeto M, Yates A, Rowland M. Chlorfenapyr: a pyrrole insecticide for the control of pyrethroid or DDT resistant Anopheles gambiae (Diptera: Culicidae) mosquitoes. Acta Trop. 2007;102:69–78.
Raghavendra K, Barik TK, Sharma P, Bhatt RM, Srivastava HC, Sreehari U, et al. Chlorfenapyr: a new insecticide with novel mode of action can control pyrethroid resistant malaria vectors. Malar J. 2011;10:16.
Dagg K, Irish S, Wiegand RE, Shililu J, Yewhalaw D, Messenger LA. Evaluation of toxicity of clothianidin (neonicotinoid) and chlorfenapyr (pyrrole) insecticides and cross-resistance to other public health insecticides in Anopheles arabiensis from Ethiopia. Malar J. 2019;18:49.
Uragayala S, Kamaraju R, Tiwari SN, Sreedharan S, Ghosh SK, Valecha N. Village-scale (phase III) evaluation of the efficacy and residual activity of SumiShield 50® WG (clothianidin 50%, w/w) for indoor spraying for the control of pyrethroid-resistant Anopheles culicifacies Giles in Karnataka state, India. Trop Med Int Health. 2018;23:605–15.
Agossa FR, Padonou GG, Fassinou AJYH, Odjo EM, Akuoko OK, Salako A, et al. Small-scale field evaluation of the efficacy and residual effect of Fludora® Fusion (mixture of clothianidin and deltamethrin) against susceptible and resistant Anopheles gambiae populations from Benin, West Africa. Malar J. 2018;17:484.
Kweka E, Mahande A, Ouma J, Msangi S, Karanja W, Temba V, et al. Novel indoor residual spray insecticide with extended mortality effect: a case of SumiShield 50WG against wild resistant populations of Anopheles arabiensis in Northern Tanzania. Glob Health Sci Pract. 2018;6:758–65.
Matsuda K, Ihara M, Sattelle DB. Neonicotinoid insecticides: molecular targets, resistance, and toxicity. Annu Rev Pharmacol Toxicol. 2020;60:241–55.
Wu S, He M, Xia F, Zhao X, Liao X, Li R, et al. The cross-resistance pattern and the metabolic resistance mechanism of acetamiprid in the Brown Planthopper, Nilaparvata lugens (Stål). Int J Mol Sci. 2022;23:9429.
Balkan T, Kara K. Neonicotinoid resistance in adults and nymphs of Bemisia tabaci (Genn., 1889) (Hemiptera: Aleyrodidae) populations in tomato fields from Tokat, Turkey. Turk Entomol Derg. 2020;44:319–31.
Zhang H, Chen A, Shan T, Dong W, Shi X, Gao X. Cross-resistance and fitness cost analysis of resistance to thiamethoxam in Melon and Cotton Aphid (Hemiptera: Aphididae). J Econ Entomol. 2020;113:1946–54.
Wei X, Pan Y, Xin X, Zheng C, Gao X, Xi J, et al. Cross-resistance pattern and basis of resistance in a thiamethoxam-resistant strain of Aphis gossypii Glover. Pestic Biochem Physiol. 2017;138:91–6.
Kaplanoglu E, Chapman P, Scott IM, Donly C. Overexpression of a cytochrome P450 and a UDP-glycosyltransferase is associated with imidacloprid resistance in the Colorado potato beetle, Leptinotarsa decemlineata. Sci Rep. 2017;7:1762.
Puinean AM, Foster SP, Oliphant L, Denholm I, Field LM, Millar NS, et al. Amplification of a cytochrome P450 gene is associated with resistance to neonicotinoid insecticides in the aphid Myzus persicae. PLoS Genet. 2010;6: e1000999.
Zhou C-S, Cao Q, Li G-Z, Ma D-Y. Role of several cytochrome P450s in the resistance and cross-resistance against imidacloprid and acetamiprid of Bemisia tabaci (Hemiptera: Aleyrodidae) MEAM1 cryptic species in Xinjiang, China. Pestic Biochem Physiol. 2020;163:209–15.
Samal RR, Panmei K, Lanbiliu P, Kumar S. Reversion of CYP450 monooxygenase-mediated acetamiprid larval resistance in dengue fever mosquito, Aedes aegypti L. Bull Entomol Res. 2022;112:557–66.
Jin R, Mao K, Liao X, Xu P, Li Z, Ali E, et al. Overexpression of CYP6ER1 associated with clothianidin resistance in Nilaparvata lugens (Stål). Pestic Biochem Physiol. 2019;154:39–45.
Edi CV, Djogbenou L, Jenkins AM, Regna K, Muskavitch MAT, Poupardin R, et al. CYP6 P450 enzymes and ACE-1 duplication produce extreme and multiple insecticide resistance in the malaria mosquito Anopheles gambiae. PLoS Genet. 2014;10: e1004236.
Weedall GD, Mugenzi LMJ, Menze BD, Tchouakui M, Ibrahim SS, Amvongo-Adjia N, et al. A cytochrome P450 allele confers pyrethroid resistance on a major African malaria vector, reducing insecticide-treated bednet efficacy. Sci Transl Med. 2019;11: eaat7386.
Ibrahim SS, Ndula M, Riveron JM, Irving H, Wondji CS. The P450 CYP6Z1 confers carbamate/pyrethroid cross-resistance in a major African malaria vector beside a novel carbamate-insensitive N485I acetylcholinesterase-1 mutation. Mol Ecol. 2016;25:3436–52.
Kamdem C, Fouet C, Gamez S, White BJ. Pollutants and insecticides drive local adaptation in African malaria mosquitoes. Mol Biol Evol. 2017;34:1261–75.
Miles A, Harding NJ, Bottà G, Clarkson CS, Antão T, Kozak K, et al. Genetic diversity of the African malaria vector Anopheles gambiae. Nature. 2017;552:96–100.
Lines JD. Do agricultural insecticides select for insecticide resistance in mosquitoes? A look at the evidence. Parasitol Today. 1988;4:S17-20.
Georghiou GP, Breeland SG, Ariaratnam V. Seasonal escalation of organophosphorus and carbamate resistance in Anopheles albimanus by agricultural sprays. Environ Entomol. 1973;2:369–74.
Simon-Delso N, Amaral-Rogers V, Belzunces L, Bonmatin JM, Chagnon M, Downs C, et al. Systemic insecticides (neonicotinoids and fipronil): trends, uses, mode of action and metabolites. Environ Sci Pollut Res. 2015;22:5–34.
Working Group Meeting. Neonicotinoids and their impact on ecosystem service for agriculture and biodiversity in Africa. Proceedings report, icipe, Nairobi. 2018.
Ramadevi R, Ramachandraiah C, Reddy GVS. A review on contamination of soil and water by neonicotinoid pesticides and trends it’s in soil and water samples with chromatographic analytical techniques. Orient J Chem. 2022;38:259–67.
Morrissey CA, Mineau P, Devries JH, Sanchez-Bayo F, Liess M, Cavallaro MC, et al. Neonicotinoid contamination of global surface waters and associated risk to aquatic invertebrates: a review. Environ Int. 2015;74:291–303.
Chouaïbou MS, De SSS, Fodjo BK, Zoh MG, Bli NK, Koudou BG. Evidence of insecticide resistance selection in wild Anopheles coluzzii mosquitoes due to agricultural pesticide use. Infect Dis Poverty. 2019;8:64.
Fouet C, Ashu FA, Ambadiang MM, Tchapga W, Wondji CS, Kamdem C. Clothianidin-resistant Anopheles gambiae adult mosquitoes from Yaoundé, Cameroon, display reduced susceptibility to SumiShield® 50WG, a neonicotinoid formulation for indoor residual spraying. BMC Infect Dis. 2024;24:133.
Ambadiang MM, Fouet C, Ashu FA, Penlap-Beng V, Kamdem C. Anopheles gambiae larvae’s ability to grow and emerge in water containing lethal concentrations of clothianidin, acetamiprid or imidacloprid is consistent with cross-resistance to neonicotinoids. bioRxiv. 2023. https://biorxiv.org/cgi/content/short/2023.04.18.537423v1.
Ashu FA, Fouet C, Ambadiang MM, Penlap-Beng V, Kamdem C. Vegetable oil-based surfactants are adjuvants that enhance the efficacy of neonicotinoid insecticides and can bias susceptibility testing in adult mosquitoes. PLoS Negl Trop Dis. 2023;17: e0011737.
CDC. Guideline for evaluating insecticide resistance in vectors using the CDC bottle bioassay. 2012. https://stacks.cdc.gov/view/cdc/21777.
Agumba S, Gimnig JE, Ogonda L, Ombok M, Kosgei J, Munga S, et al. Diagnostic dose determination and efficacy of chlorfenapyr and clothianidin insecticides against Anopheles malaria vector populations of western Kenya. Malar J. 2019;18:243.
Corbel V, Kont MD, Ahumada ML, Andréo L, Bayili B, Bayili K, et al. A new WHO bottle bioassay method to assess the susceptibility of mosquito vectors to public health insecticides: results from a WHO-coordinated multi-centre study. Parasit Vectors. 2023;16:21.
Gillies MT, Coetzee M. A supplement to the Anophelinae of Africa south of the Sahara. Johannesburg: The South African Institute for Medical Research; 1987.
Gillies MT, De Meillon B. The Anophelinae of Africa South of the Sahara. 2nd ed. Johannesburg: Publ S Afr Inst Med Res; 1968.
Kamdem C, Tene Fossog B, Simard F, Etouna J, Ndo C, Kengne P, et al. Anthropogenic habitat disturbance and ecological divergence between incipient species of the malaria mosquito Anopheles gambiae. PLoS ONE. 2012;7: e39453.
Cassone BJ, Kamdem C, Cheng C, Tan JC, Hahn MW, Costantini C, et al. Gene expression divergence between malaria vector sibling species Anopheles gambiae and An. coluzzii from rural and urban Yaoundé Cameroon. Mol Ecol. 2014;23:2242–59.
Fanello C, Santolamazza F, DellaTorre A. Simultaneous identification of species and molecular forms of the Anopheles gambiae complex by PCR-RFLP. Med Vet Entomol. 2002;16:461–4.
Abbott WS. A method for computing the effectiveness of an insecticide. J Econ Entomol. 1925;18:265–7.
R Core Team. R: a language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing; 2016.
Wheeler MW, Park RM, Bailer AJ. Comparing median lethal concentration values using confidence interval overlap or ratio tests. Environ Toxicol Chem. 2009;25:1441–4.
WHO. Standard operating procedure for testing insecticide susceptibility of adult mosquitoes in WHO bottle bioassays. Geneva: World Health Organization; 2022.
Portwood NM, Mbewe NJ, Tungu PK, Mlay G, Small G, Snetselaar J, et al. Multi-centre discriminating concentration determination of broflanilide and potential for cross-resistance to other public health insecticides in Anopheles vector populations. Sci Rep. 2022;12:22359.
Okolle NJ, Monono EY, Tabikam AI, Kinge MB, Rodrique MP. Chapter 1. Insecticide use and application in Cameroon. In: Ranz RER, editor. Insecticides: impact and benefits of its use for humanity. London: IntechOpen; 2022.
Nauen R, Bass C, Feyereisen R, Vontas J. The role of cytochrome P450s in insect toxicology and resistance. Annu Rev Entomol. 2022;67:105–24.
Ullah F, Gul H, Tariq K, Desneux N, Gao X, Song D. Functional analysis of cytochrome P450 genes linked with acetamiprid resistance in melon aphid, Aphis gossypii. Pestic Biochem Physiol. 2020;170: 104687.
Nauen R, Ebbinghaus-Kintscher U, Salgado VL, Kaussmann M. Thiamethoxam is a neonicotinoid precursor converted to clothianidin in insects and plants. Pestic Biochem Physiol. 2003;76:55–69.
Tchouakui M, Assatse T, Mugenzi LMJ, Menze BD, Nguiffo-Nguete D, Tchapga W, et al. Comparative study of the effect of solvents on the efficacy of neonicotinoid insecticides against malaria vector populations across Africa. Infect Dis Poverty. 2022;11:35.
Oxborough RM, Seyoum A, Yihdego Y, Dabire R, Gnanguenon V, Wat F, et al. Susceptibility testing of Anopheles malaria vectors with the neonicotinoid insecticide clothianidin; results from 16 African countries, in preparation for indoor residual spraying with new insecticide formulations. Malar J. 2019;18:264.
Antonio-Nkondjio C, Kerah CH, Simard F, Awono-Ambene P, Chouaibou M, Tchuinkam T, et al. Complexity of the malaria vectorial system in Cameroon: contribution of secondary vectors to malaria transmission. J Med Entomol. 2006;43:1215–21.
Stevenson J, St. Laurent B, Lobo NF, Cooke MK, Kahindi SC, Oriango RM, et al. Novel vectors of malaria parasites in the western highlands of Kenya. Emerg Infect Dis. 2012;18:1547–9.
Costantini C, Ayala D, Guelbeogo WM, Pombi M, Some CY, Bassole IH, et al. Living at the edge: biogeographic patterns of habitat segregation conform to speciation by niche expansion in Anopheles gambiae. BMC Ecol. 2009;9:16.
Simard F, Ayala D, Kamdem GC, Pombi M, Etouna J, Ose K, et al. Ecological niche partitioning between Anopheles gambiae molecular forms in Cameroon: the ecological side of speciation. BMC Ecol. 2009;9:17.
Coetzee M, Koekemoer L. Molecular systematics and insecticide resistance in the major African malaria vector Anopheles funestus. Annu Rev Entomol. 2013;58:393–412.
Assatse T, Tchouakui M, Mugenzi L, Menze B, Nguiffo-nguete D, Tchapga W, et al. Anopheles funestus populations across Africa are broadly susceptible to neonicotinoids but with signals of possible cross-resistance from the GSTe2 gene. Trop Med Infect Dis. 2023;8:244.
Riveron JM, Huijben S, Tchapga W, Tchouakui M, Wondji MJ, Tchoupo M, et al. Escalation of pyrethroid resistance in the malaria vector Anopheles funestus induces a loss of efficacy of piperonyl butoxide-based insecticide-treated nets in Mozambique. J Infect Dis. 2019;220:467–75.
Curtis GF, Miller JE, Hodjati MH, Kolaczinski JH, Kasumba I. Can anything be done to maintain the effectiveness of pyrethroid-impregnated bednets against malaria vectors? Philos Trans R Soc B Biol Sci. 1998;353:1769–75.
Funding
This study was supported by a National Institutes of Health grant (R01AI150529) to CK. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
CF and CK conceived and designed the study. FA, MA, CF and CK conducted fieldwork, performed laboratory tests and analysed the data. VPB contributed resources and supervision. AF and CK wrote the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Ashu, F.A., Fouet, C., Ambadiang, M.M. et al. Adult mosquitoes of the sibling species Anopheles gambiae and Anopheles coluzzii exhibit contrasting patterns of susceptibility to four neonicotinoid insecticides along an urban-to-rural gradient in Yaoundé, Cameroon. Malar J 23, 65 (2024). https://doi.org/10.1186/s12936-024-04876-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12936-024-04876-4