
Abstract
Background
Malaria remains a major public health concern in Cameroon. Understanding vector distribution and malaria transmission dynamics is of paramount importance for evaluating the performance of control strategies. This study assesses patterns of malaria transmission in four eco-epidemiological settings in Cameroon.
Methods
Adult mosquitoes were collected using Human Landing Catches (HLC) once every 4 months from August 2019 to November 2021 in Kaélé, Tibati, Santchou and Bertoua. Mosquitoes were sorted by genus and Anopheles gambiae sensu lato (s.l.) species complex were identified using PCR. The presence of Plasmodium falciparum circumsporozoite protein (CSP) was measured by ELISA; the entomological inoculation rates (EIR) was estimated in each locality.
Results
A total of 23,536 mosquitoes were collected. Anopheles gambiae and/or Anopheles coluzzii were the main malaria vectors in all sites. Anopheles arabiensis was recorded in low frequency in Kaélé and Tibati. Other species collected included Anopheles funestus, Anopheles pharoensis and Anopheles ziemmani. High anopheline biting rates were recorded outdoor in all sites except in Kaélé. Important differences in species biting dynamics were observed between sites. The sporozoite infection rate varied from 0.36 to 4%. The daily EIR was found to vary from 0.07 in Santchou to 0.26 infected bites/man/night (ib/m/n) in Kaélé).
Conclusion
The study suggests heterogeneous patterns of malaria transmission in different ecoepidemiological settings across the country. The findings stress the need to improve malaria vector control strategies.
Similar content being viewed by others
Background
Despite more than a century of malaria research, the disease remains a major public health concern in Africa [1]. It is estimated that about 241 million people across the world suffer from malaria yearly, with 627,000 associated deaths registered in 2021 [1]. The implementation of key malaria prevention measures such as the massive scale up of ITNs has played significant role in decreasing the burden of malaria between 2010 and 2015 [2]. Since 2017, the downward trend in morbidity and mortality has slowed and even reversed with the COVID-19 pandemic which has limited the delivery of essential commodities used for malaria prevention or case management [1].
In Cameroon, malaria represents a major public health concern with the entire country being at risk of transmission [3]. In 2019, up to 2638191 cases were recorded in health care facilities with 4500 associated deaths. Children under 5 years are the most affected with 65% of deaths among the total number of death caused by malaria [3]. The national vector control policy is based primarily on the widespread use of insecticide-treated nets (ITNS) [3]. It is estimated that, more than 80% of the population possess a treated bed net [4,5,6,7]. Unfortunately, several factors still affect the performance of ITNs, such as changes in vector feeding and resting behaviour and the rapid spread of insecticide resistance in malaria vector populations [8,9,10,11,12]. The success of vector control strategies requires up to date information on vector population distribution, bionomic and malaria transmission pattern [9,10,11].
A high number of species contribute to malaria transmission across the country [8, 13,14,15,16]. Variations in the distribution and behavior of these vector species affect vector control [11, 13, 14]. Species such as Anopheles gambiae sensu lato (s.l.) despite displaying high endophilic and endophagic behavior can become exophilic or exophagic in response to the use of impregnated mosquito nets [17]. It is now clear that the use of indoor based interventions alone will not achieve malaria elimination if outdoor biting or resting vectors are not targeted [18].
Although, increasing efforts have been made to characterize malaria transmission patterns across several epidemiological settings in Cameroon [6, 11, 13,14,15, 19,20,21,22,23,24,25,26,27,28,29], much is still left to be done. There is paucity of data in some region including the deep Eastern forest region, the Adamawa and the western highlands region. This information could be crucial for improving malaria control strategies and filling important knowledge gaps which are impeding control efforts. This study presents data on vector distribution and malaria transmission pattern in four eco-epidemiological settings in Cameroon: the Sahelian zone (Kaélé), the humid savannah area (Tibati), the West highland area covered by grassfields (Santchou), and the forest zone (Bertoua).
Methods
Study sites
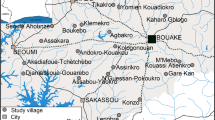
The study was conducted from July 2019 to September 2021 in four epidemiological settings in Cameroon: Kaélé in the Sahelian zone, Tibati in the Sahelo-soudanese area, Bertoua in the forest area and Santchou in the western highlands. The characteristics of the different study sites are presented in Table 1 and Fig. 1.

Map of the study sites
Kaélé is a locality of over 127000, close to the Maga dam on the Logone river. Malaria situation in the area could possibly be affected by the frequent movements of the population between Cameroon and Chad. The region belongs to the Sahelian domain characterize by a long dry season running from October to May with only 4 months of rains, going from June to September.
Tibati is a locality of over 36,000 inhabitants, situated midway between the north and the south of the country close to the Mbakaou dam on the Sanaga river. The region belongs to the Sahelo-Sudanese domain characterize by two seasons, a dry season extending from November to February and a rainy season from March to October. Although considered as highly endemic for malaria, there is little data on malaria transmission pattern and vector distribution.
Bertoua is a city of over 200 000 inhabitants situated in the equatorial forest zone with several river systems: the Nyong, the Dja, the Lom, the Kadeï, the Boumba and Ngoko. The region belongs to the Sahelo-Sudanese domain characterized by four seasons including two rainy seasons and two dry season. A long rainy season from September to November, a long dry season from November to March, a short rainy season from March to June and a short dry season from July to August. Previous studies indicated a high frequency of insecticide resistance in vector populations [30, 31]. The region is also one of the most affected by malaria in the country [3].
Santchou is a locality of over 41,000 inhabitants, situated in the highland zone. The region is characterize by four seasons including a long rainy season from mid- August to October; a short rainy season from March to June; a long dry season from mid-October to March and a short dry season from June to Mid-August. Santchou is an area with intensive practice of agriculture and frequent use of pesticides [24, 32]. The river Nkam and its tributaries provide a dense hydrography in the locality.
Mosquito collections and processing
Adult mosquitoes were sampled from August 2019 to November 2021 using Human Landing Catches (HLC). A total of six surveys were conducted in Santchou and Bertoua (August 2019, November 2019, July 2020, November 2020, May 2021, August 2021) and five in Tibati and Kaélé (August 2019, November 2019, November 2020, May 2021, August 2021). In each site, mosquito collection was carried out both indoors and outdoors in four randomly selected houses between 07:00 pm and 06:00 am. Mosquito collection was undertaken during two consecutive nights. Each mosquito collector worked half a night, from 7 pm to 1 am or from 1 to 6 am. A circular permutation of the place of collection and of the time slots was established for each of the collector in order to reduce the effects of individual factors. After mosquito collection, culicines were identified and separated from anophelines. Anopheline species were identified using keys for the morphological identification of Gillies and Coetzee [33]. Anopheles specimens were stored individually in labelled Eppendorf tubes containing a desiccant, then transported to the Malaria Research Laboratory of the OCEAC for further analysis.
Laboratory processing of mosquitoes
Detection of Plasmodium parasite infections in mosquitoes
The heads and thoraces of female anophelines were tested for the presence of circumsporozoite protein using enzyme-linked immunosorbent assay (ELISA) according to Wirtz et al. [34]. A sample was considered positive when its optical density (OD) value was two-fold higher than the mean optical density of negative control.
Molecular identification of species of the Anopheles gambiae complex
A sample of 50 females belonging to the An. gambiae complex were randomly selected after each collection period for DNA extraction. DNA was extracted from the legs, abdomen or wings using the Livak protocol [35]. The SINE PCR [36] was performed to identify members of the An. gambiae complex.
Analysis
The human biting rate (HBR) was estimated as the number of mosquitoes collected per man per night. The sporozoite infection rate (or circumsporozoroite rate) was calculated by dividing the number of female anopheline found infected by the total number of mosquitoes screened. The daily entomological inoculation rate (EIR) was calculated by multiplying the human biting rate by the circum sporozoite rate. Comparisons of proportion were made using the Chi-square test. The level of significance of each test was set at P < 0.05.
Results
Composition of Culicidae fauna in the different study sites
A total of 23,533 mosquitoes were collected during the study period (Fig. 2 and Additional file 1: Table S1). Four genera were recorded after morphological identification namely Culex, Mansonia, Anopheles and Aedes. The genus Culex was the most represented (59.83%; n = 14,079). Anopheline represented 28.31% (6663/23,533) of the total mosquitoes collected. Kaélé had the highest number of Anopheles specimens caught (46.17%; n = 4484) followed by Tibati (29.83%; n = 1449), Santchou (10.63%; n = 446) and Bertoua (5.95%; n = 284).

Distribution of mosquito genera in the different study sites
Composition of the Anophelinae fauna in the different study sites
A total of 6666 Anopheles mosquitoes belonging to six anopheline species, including An. gambiae., Anopheles coluzzii, Anopheles arabiensis, Anopheles ziemmani, Anopheles pharoensis and Anopheles funestus were collected (Table 2). Anopheles gambiae s.l. was the most abundant in all collection sites representing 68.54% (4567/6666) of the total anopheline fauna (Fig. 3 and Additional file 1: Table S2).

Distribution of Anopheles species in the different study sites
A total of 800 out of the 4567 An. gambiae s.l. were further processed by PCR to identify sibling species of the An. gambiae complex in each site. Anopheles gambiae, An. coluzzii and An. arabiensis were recorded. Anopheles gambiae sensu stricto (s.s.) was the most frequent member of the group in Kaélé (50.40%; n = 126/250), Santchou (72%; n = 108/150) and Bertoua (64%; n = 96/150). Anopheles coluzzii was more frequent in Tibati (70%; n = 175/250 (Fig. 4 and Additional file 1: Table S3). Anopheles arabiensis was recorded in low frequencies exclusively in Kaélé (12%; n = 30/250) and Tibati (5.20%; n = 13/250).

Distribution of members of the An. gambiae complex in the different study sites
Indoors and outdoors biting Anopheline densities in different study sites
The proportion of female anopheline mosquitoes caught outdoor was 57.97% (n = 1449) in Tibati; 62.69% (n = 446) in Santchou, 60.35% (n = 285) in Bertoua and 48.48% (n = 4484) in Kaélé. These proportions were significantly higher compared to indoor catches in Tibati (χ2 = 35.27; Df = 1; P < 0.0001), Santchou (χ2 = 26.05; Df = 1; P = 0.001) and Bertoua (χ2 = 10.89; Df = 1; P < 0.0001). The density of anopheline was linked to rainfall and seasons. High densities occur during the rainy season with the predominance of An. gambiae s.l. in all sites (Fig. 5).

Distribution of anopheline species indoor and outdoor according to collection periods (In: indoor; Out: outdoor)
In Kaélé, An. gambiae s.l. was the predominant species during collection done in July 2019, November 2020 and August 2021 with biting rate reaching 122.75 b/m/n. In 2021, An. gambiae and An. pharoensis were recorded at similar densities and were the most predominant species (Fig. 5a). In Tibati, a peack of bite for An. gambiae s.l. (125 b/m/n) was recorded in July 2019 but a total change in biting densities was detected in August 2021 with An. ziemmani becoming the most abundant species (Fig. 5b). In Bertoua and Santchou, An. gambiae s.l. was always the most abundant species with biting densities varying from 0.1 to 8.74 b/m/n in Santchou and 0.1 to 3.91 b/m/n in Bertoua (Fig. 5c, d).
Night biting cycle of Anopheline species
The night biting cycle of anopheline species are presented in Fig. 6. In Santchou (Fig. 6d) and Bertoua (Fig. 6c) most of anopheline bites were recorded during the second half of the night (from 00:00 to 6:00 am) whereas in Tibati and Kaélé (Fig. 6a, b), most of the bites were recorded during the first half of the night (from 7:00 pm to 00:00 am).

Biting cycles of Anophelines species in different study sites
Plasmodium infection
A total of 4889 mosquitoes were screened to detect the presence of Plasmodium infections. After analysis, 36 Anopheline specimens were found infected representing a global infection rate of 0.74%. Both An. gambiae s.l. and An. pharoensis were detected infected in Kaélé with infection rates of 0.36% (6/1645) and 0.20% (2/1028) respectively. In Santchou, both An. gambiae s.l. and An. ziemmani were detected infected with sporozoite rate of 2% (6/309) and 16.70% (1/6) respectively. In Tibati and Bertoua, only An.gambiae s.l. was found infected, with sporozoite rate of 0.80% (10/1285) in Tibati and 4% (6/151) in Bertoua (Table 3).
Entomological inoculation rates
The average daily entomological inoculation rate (EIR) was 0.26 ib/m/n in Kaélé and 0.21 ib/m/n in Tibati, 0.08 ib/m/n in Bertoua and 0.07 ib/m/n in Santchou (Table 4). Seasonal variation of the EIR was observed in all sites. An. gambiae was involved in malaria transmission in all sites across all the periods (Fig. 7).

Monthly variation of the entomological inoculation rates in different study sites
In Kaélé, An. gambiae s.l. was responsible for malaria transmission, except in November 2019, marking the beginning of the dry season. Anopheles pharoensis was found infected during the months of May and August 2021. Annual transmission was estimated at 95 infected bites/man/year (Fig. 7a). The highest transmission occurs in August 2021 during the rainy season.
In Tibati, transmission was mainly due to An. gambiae s.l. except August 2021. Annual transmission was estimated at 76.65 ib/m/y (Fig. 7b). The highest transmission was recorded in July 2019 during the pick of the rainy season.
In Santchou, malaria transmission was ensured by An. gambiae s.l. and An. ziemmani during the periods of July 2020 and May 2021. Annual transmission was estimated at 25.55 ib/m/y. The month of May 2021 in the mid of the rainy season was associated with high EIR (11.66 ib/m/month) maintain by An. gambiae (Fig. 7d).
In Bertoua, only An. gambiae s.l. was recorded as vector and transmission was registered in July 2020, May 2021 and August 2021. Annual transmission rate in the locality was estimated at 29.20 ib/m/y. The month of July 2020 during the short dry season was associated with the highest EIR (7.50 ib/m/y) (Fig. 7c).
Discussion
The main objective of the study was to assess the dynamic of malaria transmission in sentinel sites within four ecoepidemiological settings in Cameroon including the Sahelian (Kaélé), Soudano-sahelian (Tibati), highland (Santchou) and forest zone (Bertoua). Despite high possession and usage of ITNs in studied sites [6], continuous transmission of malaria was recorded. Transmission estimates recorded during this study were similar to data recorded in other parts of the country [8, 13,14,15]. Yet very high transmission intensity was found to occur in both Kaélé and Tibati situated in Sahelian and Soudano-sahelian zones, respectively, compared with Santchou and Bertoua situated in the highland and forest area, respectively. This may result from the high biting anopheline densities in those areas particularly during the rainy season. The two sites are situated close to dams which might contribute in the maintenance of vector populations all year long. The EIR in Santchou was low, which is in line with previous findings supporting moderate malaria endemicity in the highland region [24, 37]. Malaria transmission intensity in Bertoua was also relatively moderate. Bertoua is situated in the East region of Cameroon considered as one of the most affected by malaria in the country with high morbidity and high mortality rates [38]. The moderate transmission pattern in the city of Bertoua may owe to the development of infrastructures such as drains and roads during past years.
Six Anopheline species were identified in this study, namely An. gambiae s.s., An. coluzzii, An. arabiensis, An. ziemmani, An. pharoensis, and An. funestus. The presence of different species was consistent with the diversity of aquatic habitats for mosquitoes in the studied sites including permanent water sources, puddles, dams, rivers. Species diversity was poor compared to previous reports [15] and probably reflects the effects of climatic or environmental changes on species distribution. Studies conducted so far in Cameroon point at a temperature increase of + 0.4 °C compared with the 1961–1990 period, and a rainfall reduction ranging from − 10 to − 20% [39]. Anopheles gambiae s.l. was the most abundant species and the major malaria vector in all sites. Previous studies indicated the high adaptation capacity of members of the complex to environmental conditions [32, 40,41,42]. Three members of the An. gambiae complex were detected, including An. gambiae, An. coluzzii and An. arabiensis. Both An. gambiae and An. coluzzii were found in sympatry in the four sites, whereas An. arabiensis was only detected in Kaélé and Tibati, with a low frequency. The distribution of members of the An. gambiae complex was consistent with previous studies mapping the distribution of these species in the country [40].
Other anopheline species, including An. funestus, An. pharoensis and An. ziemmani; were collected alongside members of the An. gambiae complex. Despite the high vectorial capacity of An. funestus this species was not found infected in any of the sites. However, this species has usually been found to display a high vectorial capacity compared to An. gambiae in different epidemiological settings [15, 19]. Anopheles pharoensis and An. ziemmani were found infected both in Kaélé and Santchou respectively. Considered as secondary vectors, these species were actively involved in malaria transmission. These findings are consistent with previous studies in Cameroon where up to 17 species were reported involved in malaria transmission in various places [15, 43, 44]. Anopheles pharoensis was collected in very high densities in Kaélé. The species is considered to breed in shallow, sunlight, freshwater pools with high organic content; the presence of rice paddies in Kaélé and a lake reservoir with a high vegetation on it edges could possibly constitute preferential breeding habitats for the species [45, 46]. The distribution of anopheline species in Kaélé could be highly dynamic since previous studies indicated high densities of An. funestus in the area [47], whereas no An. funestus was collected during the present study. It is possible that the reduction of land surface used for rice cultivation, extension of human settlements following the severe drought which affected the region since 2017 and population displacement following Boko-haram insurgency alongside new agricultural practices are shaping species distribution and malaria transmission patterns within the area [48,49,50]. The situation deserves further investigations. Anopheles ziemmani was recorded in high density in both Tibati and Kaélé this species was mostly collected outdoors. Anopheles ziemmani has so far received less attention despite its implication in malaria transmission in Cameroon [15]. The species belongs to a group composed of Anopheles coustani, Anopheles namibiensis, Anopheles obscurus and Anopheles implexus with some being highly zoophilic and not involved in malaria transmission at all [51].
Anopheline species were frequently caught biting outdoors in almost all sites except in Kaélé. This behaviour might result from the high ownership and usage of insecticide treated bed nets by the population [6]. This is in line with previous studies in Cameroon, indicating high transmission of malaria both indoor and outdoor [11, 27]. This trend was also described in many other African countries [14, 52, 53]. In Kaélé, anopheline were found to bite mostly indoor. Due to the high nuisance in the area the population usually stay indoor during the night and this behaviour might have influenced mosquito biting behaviour. It is possible that the high frequency of resistant mosquito in the area could also explain why mosquitoes in the area are not affected by the excito-repellency effects of insecticide treated bed nets.
Mosquitoes were found to bites all night long with picks in Kaélé and Tibati recorded during the first half of the night and during the second half of the night in Bertoua and Santchou. Anopheles pharoensis and An. ziemmani were found to bite predominantly during the first part of the night, whereas An. gambiae s.l. was found to bite both in the first and second part of the night. Studies conducted so far in Cameroon indicated that mosquitoes could bite up to 8 am [54] although for the moment there is still no data on the epidemiological consequences of this behaviour. This calls for further actions to improve malaria vector control in the country.
Conclusion
This study of the dynamics of malaria transmission in different eco-epidemiological settings shows a heterogeneous pattern of malaria transmission. Malaria transmission was found to be high in all eco-epidemiological settings with different factors contributing to transmission. The high presence of vectors biting outdoors calls for urgent actions in order to improve the fight against malaria in Cameroon. Additional tools to control outdoor biting mosquitoes should be implemented (larviciding, environmental management). Integrated vector management could constitute an efficient way to fight against continuous malaria in the country and achieve elimination goals.
Availability of data and materials
The datasets supporting the findings of this paper are included in this paper.
References
WHO. World malaria report 2021. Geneva: World Health Organization; 2021. https://apps.who.int/iris/handle/10665/350147
WHO. World Malaria Report 2015. Geneva: World Health Organization. 2015.
PNLP. Cameroun – Programme National de Lutte contre le Paludisme. https://pnlp.cm/.
Ndo C, Menze-Djantio B, Antonio-Nkondjio C. Awareness, attitudes and prevention of malaria in the cities of Douala and Yaoundé (Cameroon). Parasit Vectors. 2011;4:181.
Abdou T, Ngadjeu C, Doumbe B, Djamouko-Djonkam L, Sonhafouo-Chiana N, Kopya E, et al. Malaria prevention in the city of Yaoundé: Knowledge and practices of urban dwellers. Malar J. 2019;18:167.
Kala Chouakeu NA, Ngingahi LG, Bamou R, Talipouo A, Ngadjeu CS, Mayi MPA, et al. Knowledge, attitude, and practices (KAP) of human populations towards malaria control in four ecoepidemiological settings in Cameroon. J Trop Med. 2021;2021:9925135.
WHO. Strategies for delivering insecticide-treated nets at scale for malaria control: a systematic review. Geneva, World Health Organization. https://www.who.int/bulletin/volumes/90/9/11-094771/en/
Antonio-Nkondjio C, Ndo C, Njiokou F, Bigoga JD, Awono-Ambene P, Etang J, et al. Review of malaria situation in Cameroon: technical viewpoint on challenges and prospects for disease elimination. Parasit Vectors. 2019;12:501.
Antonio-Nkondjio C, Sonhafouo-Chiana N, Ngadjeu CS, Doumbe-Belisse P, Talipouo A, Djamouko-Djonkam L, et al. Review of the evolution of insecticide resistance in main malaria vectors in Cameroon from 1990 to 2017. Parasit Vectors. 2017;10:472.
Talom AD, Essoung MA, Gbankoto A, Tchigossou G, Akoton R, Sahabi BBA, et al. A preliminary analysis on the effect of copper on Anopheles coluzzii insecticide resistance in vegetable farms in Benin. Sci Rep. 2020;10:6392.
Bamou R, Mbakop L, Kopya E, Ndo C, Awono-Ambene P, Tchuinkam T, et al. Changes in malaria vector bionomics and transmission patterns in the equatorial forest region of Cameroon between 2000 and 2017. Parasit Vectors. 2018;11:464.
Mandeng SE, Awono-Ambene HP, Bigoga JD, Ekoko WE, Binyang J, Piameu M, et al. Spatial and temporal development of deltamethrin resistance in malaria vectors of the Anopheles gambiae complex from North Cameroon. PLoS ONE. 2019;14: e0212024.
Mbakop L, Awono-Ambene P, Mandeng S, Ekoko W, Fesuh B, Antonio-Nkondjio C, et al. Malaria transmission around the Memve’ele hydroelectric dam in South Cameroon: a combined retrospective and prospective study, 2000–2016. Int J Environ Res Public Health. 2019;16:1618.
Bamou R, Rono M, Degefa T, Midega J, Mbogo C, Ingosi P, et al. Entomological and anthropological factors contributing to persistent malaria transmission in Kenya, Ethiopia, and Cameroon. J Infect Dis. 2021;223:S155–70.
Antonio-Nkondjio C, Simard FDR, Awono-Ambene P, Chouaibou M, Tchuinkam TO, Fontenille D. Complexity of the malaria vectorial system in Cameroon: contribution of secondary vectors to malaria transmission. J Med Entomol. 2006;43:1215–21.
Awono-ambene HP, Kengne P, Simard F, Antonio-Nkondjio C, Fontenille D. Description and bionomics of Anopheles (Cellia) ovengensis (Diptera: Culicidae), a new malaria vector species of the Anopheles nili group from South Cameroon. J Med Entomol. 2004;41:561–8.
Sougoufara S, Diédhiou SM, Doucouré S, Diagne N, Sembène PM, Harry M, et al. Biting by Anopheles funestus in broad daylight after use of long-lasting insecticidal nets: a new challenge to malaria elimination. Malar J. 2014;13:125.
Gnanguenon V, Govoetchan R, Agossa FR, Ossè R, Oke-Agbo F, Azondekon R, et al. Transmission patterns of Plasmodium falciparum by Anopheles gambiae in Benin. Malar J. 2014;13:444.
Antonio-Nkondjio C, Awono-Ambene P, Toto J-C, Meunier J-Y, Zebaze-Kemleu S, Nyambam R, et al. High malaria transmission intensity in a village close to Yaounde, the capital city of Cameroon. J Med Entomol. 2002;39:350–5.
Antonio-Nkondjio C, Simard F, Awono-Ambene P, Ngassam P, Toto J-C, Tchuinkam T, et al. Malaria vectors and urbanization in the equatorial forest region of south Cameroon. Trans R Soc Trop Med Hyg. 2005;99:347–54.
Bigoga JD, Manga L, Titanji VP, Coetzee M, Leke RG. Malaria vectors and transmission dynamics in coastal south-western Cameroon. Malar J. 2007;6:5.
Wanji S, Tanke T, Atanga SN, Ajonina C, Nicholas T, Fontenille D. Anopheles species of the mount Cameroon region: biting habits, feeding behaviour and entomological inoculation rates. Trop Med Int Health. 2003;8:643–9.
Ayala D, Costantini C, Ose K, Kamdem GC, Antonio-Nkondjio C, Agbor J-P, et al. Habitat suitability and ecological niche profile of major malaria vectors in Cameroon. Malar J. 2009;8:307.
Tchuinkam T, Simard F, Lélé-Defo E, Téné-Fossog B, Tateng-Ngouateu A, Antonio-Nkondjio C, et al. Bionomics of Anopheline species and malaria transmission dynamics along an altitudinal transect in Western Cameroon. BMC Infect Dis. 2010;10:119.
Tia E, Chouaibou M, Gbalégba CNG, Boby AMO, Koné M, Kadjo AK. Distribution des espèces et de la fréquence du gène Kdr chez les populations d’Anopheles gambiae s.s. et d’Anopheles coluzzii dans cinq sites agricoles de la Côte d’Ivoire. Bull Soc Path Exot. 2017;110:130–4.
Menze BD, Wondji MJ, Tchapga W, Tchoupo M, Riveron JM, Wondji CS. Bionomics and insecticides resistance profiling of malaria vectors at a selected site for experimental hut trials in central Cameroon. Malar J. 2018;17:317.
Doumbe B, Ngadjeu C, Sonhafouo-Chiana N, Abdou T, Djamouko-Djonkam L, Kopya E, et al. High malaria transmission sustained by Anopheles gambiae sl occurring both indoors and outdoors in the city of Yaoundé. Wellcome Open Res Cameroon. 2018. https://doi.org/10.1688/wellcomeopenres.14963.1.
Tchouakui M, Chiang M-C, Ndo C, Kuicheu CK, Amvongo-Adjia N, Wondji MJ, et al. A marker of glutathione S-transferase-mediated resistance to insecticides is associated with higher Plasmodium infection in the African malaria vector Anopheles funestus. Sci Rep. 2019;9:5772.
Mieguim Ngninpogni D, Ndo C, Ntonga Akono P, Nguemo A, Nguepi A, Metitsi DR, et al. Insights into factors sustaining persistence of high malaria transmission in forested areas of sub-Saharan Africa: the case of Mvoua. South Cameroon Parasit Vectors. 2021;14:2.
Reimer L, Fondjo E, Patchoké S, Diallo B, Lee Y, Ng A, et al. Relationship Between kdr mutation and resistance to pyrethroid and DDT insecticides in natural populations of Anopheles gambiae. J Med Entomol. 2008;45:260–6.
Etang J, Fondjo E, Chandre F, Morlais I, Brengues C, Nwane P, et al. First report of knockdown mutations in the malaria vector Anopheles gambiae from Cameroon. Am J Trop Med Hyg. 2006;74:795–7.
Tene Fossog B, Ayala D, Acevedo P, Kengne P, Ngomo Abeso Mebuy I, Makanga B, et al. Habitat segregation and ecological character displacement in cryptic African malaria mosquitoes. Evol Appl. 2015;8:326–45.
Gillies MT. A supplement to the Anophelinae of Africa south of the Sahara (Afrotropical Region). Publ South Afr Inst Med Res. 1987;55:1–143.
Wirtz RA, Zavala F, Charoenvit Y, Campbell GH, Burkot TR, Schneider I, et al. Comparative testing of monoclonal antibodies against Plasmodium falciparum sporozoites for ELISA development. Bull World Health Organ. 1987;65:39–45.
Livak KJ. Organization and mapping of a sequence on the Drosophila melanogaster X and Y chromosomes that is transcribed during spermatogenesis. Genetics. 1984;107:611–34.
Santolamazza F, Mancini E, Simard F, Qi Y, Tu Z, della Torre A. Insertion polymorphisms of SINE200 retrotransposons within speciation islands of Anopheles gambiae molecular forms. Malar J. 2008;7:163.
Amvongo Adjia N, Riveron Miranda J, Njiokou F, Wanji S, Wondji C. Influence of a major mountainous landscape barrier (Mount Cameroon) on the spread of metabolic (GSTe2) and target-site (Rdl) resistance alleles in the African malaria vector Anopheles funestus. Genes. 2020;11:1492.
Données clés sur le paludisme au Cameroun—PNLP Cameroun. 2021. https://pnlp.cm/donnees-cles-sur-le-paludisme-au-cameroun/
Sighomnou D. Analyse et redéfinition des régimes climatiques et hydrologiques du Cameroun Perspectives d’évolution des ressources en eau. Ph.D. Thesis, Université of Yaoundé I, Yaoundé, 2004. Scientific Research Publishing. https://www.scirp.org/%28S%28lz5mqp453edsnp55rrgjct55%29%29/reference/referencespapers.aspx?referenceid=1787943
Wondji C, Simard FDR, Petrarca V, Etang J, Santolamazza F, Torre AD, et al. Species and populations of the Anopheles gambiae complex in Cameroon with special emphasis on chromosomal and molecular forms of Anopheles gambiae ss. J Med Entomol. 2005;42(8):998.
Tene Fossog B, Kopya E, Ndo C, Menze-Djantio B, Costantini C, Njiokou F, et al. Water quality and Anopheles gambiae larval tolerance to pyrethroids in the cities of Douala and Yaoundé (Cameroon). J Trop Med. 2012;2012: 429817.
Fossog Tene B, Poupardin R, Costantini C, Awono-Ambene P, Wondji CS, Ranson H, et al. Resistance to DDT in an urban setting: common mechanisms implicated in both M and S forms of Anopheles gambiae in the city of Yaoundé Cameroon. PLoS ONE. 2013;8: e61408.
Awono-Ambene PH, Etang J, Antonio-Nkondjio C, Ndo C, Eyisap WE, Piameu MC, et al. The bionomics of the malaria vector Anopheles rufipes Gough, 1910 and its susceptibility to deltamethrin insecticide in North Cameroon. Parasit Vectors. 2018;11:253.
Antonio-Nkondjio C, Atangana J, Ndo C, Awono-Ambene P, Fondjo E, Fontenille D, et al. Malaria transmission and rice cultivation in Lagdo, northern Cameroon. Trans R Soc Trop Med Hyg. 2008;102:352–9.
Fondjo E, Robert V, Le Goff G, Toto JC, Carnevale P. [Urban malaria in Yaounde (Cameroon) 2 entomologic study in 2 suburban districts] (in French). Bull Soc Path Exot. 1990;1992(85):57–63.
Atangana J, Fomena A, Tamesse JL, Fondjo E. [Agricultural activities and epidemiology of malaria in Soudano-Sahelian zone in Cameroon] (in French). Bull Société Path Exot. 1990;2012(105):23–9.
Feufack-Donfack LB, Sarah-Matio EM, Abate LM, Bouopda Tuedom AG, Ngano Bayibéki A, Maffo Ngou C, et al. Epidemiological and entomological studies of malaria transmission in Tibati, Adamawa region of Cameroon 6 years following the introduction of long-lasting insecticide nets. Parasit Vectors. 2021;14:247.
Paul P, Kangalawe RYM, Mboera LEG. Land-use patterns and their implication on malaria transmission in Kilosa District. Tanzan Trop Dis Travel Med Vaccin. 2018;4:6.
Shah HA, Carrasco LR, Hamlet A, Murray KA. Exploring agricultural land-use and childhood malaria associations in sub-Saharan Africa. Sci Rep. 2022;12:4124.
Fornace KM, Abidin TR, Alexander N, Brock P, Grigg MJ, Murphy A, et al. Association between landscape factors and spatial patterns of Plasmodium knowlesi infections in Sabah. Malaysia Emerg Infect Dis. 2016;22:201–9.
Gillies MT, De Meillon B. The Anophelinae of Africa south of the Sahara (Ethiopian Zoogeographical Region). Johannesburg: South African Institute for Medical Research. 1968. https://www.cabdirect.org/cabdirect/abstract/19692900946
Assouho KF, Adja AM, Guindo-Coulibaly N, Tia E, Kouadio AMN, Zoh DD, et al. Vectorial transmission of malaria in major districts of Côte d’Ivoire. J Med Entomol. 2020;57:908–14.
Mwangangi JM, Mbogo CM, Orindi BO, Muturi EJ, Midega JT, Nzovu J, et al. Shifts in malaria vector species composition and transmission dynamics along the Kenyan coast over the past 20 years. Malar J. 2013;12:13.
PSNLP-2019–2023-Consolide-transmis.pdf. https://lnsp-cam.org/wp-content/uploads/2021/10/PSNLP-2019-2023-CONSOLIDE-TRANSMIS.pdf
Acknowledgements
We are grateful to Djoufouna Joel, Mayi Marie Paul Audrey, Ningahi Laura Gilberine and Joko Steve for their assistance during the field work. We are also grateful to administrative and traditional authorities, mosquito collectors and the population of Kaélé, Tibati, Santchou and Bertoua for their participation in the study.
Funding
This work received financial support from Bill & Melinda Gates and Panafrican Mosquito Association (ID: OPP1210340) to CAN. The funding body did not have any role in the experimental design, collection of data, analysis, and interpretation of data, or in the writing of the manuscript.
Author information
Authors and Affiliations
Contributions
KCNA: participated in data collection; performed laboratory analysis, analyzed and interpreted the data; wrote the manuscript; revised the manuscript. TT: critically revised the manuscript. BR: participated in data collection, laboratory analysis and revised the manuscript. KE: participated in data collection, laboratory analysis. TA, BM: revised the manuscript. PAA: critically revised the manuscript. CAN: Conceived and designed the study protocol; critically revised the manuscript. All authors read and approved the final maunscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The study was conducted under the ethical clearance N°2020/04/1209/CE/CNERSH/SP delivered by the Cameroon National Ethics Committee for Research on Human Health (CNERSH). Signed authorizations from District Health Officers (DHO) and owners of houses that served as collection points were also obtained.
Consent for Publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1:
Table S1. Distribution of mosquito genera in the different study sites. Table S2. Distribution of Anopheles species in the different study sites. Table S3. Distribution of members of the An. gambiae complex in different study sites.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Chouakeu, N.A.K., Tchuinkam, T., Bamou, R. et al. Malaria transmission pattern across the Sahelian, humid savanna, highland and forest eco-epidemiological settings in Cameroon. Malar J 22, 116 (2023). https://doi.org/10.1186/s12936-023-04544-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12936-023-04544-z