
Abstract
Breast cancer is significantly influenced by endoplasmic reticulum (ER) stress, impacting both its initiation and progression. When cells experience an accumulation of misfolded or unfolded proteins, they activate the unfolded protein response (UPR) to restore cellular balance. In breast cancer, the UPR is frequently triggered due to challenging conditions within tumors. The UPR has a dual impact on breast cancer. On one hand, it can contribute to tumor growth by enhancing cell survival and resistance to programmed cell death in unfavorable environments. On the other hand, prolonged and severe ER stress can trigger cell death mechanisms, limiting tumor progression. Furthermore, ER stress has been linked to the regulation of non-coding RNAs (ncRNAs) in breast cancer cells. These ncRNAs, including microRNAs (miRNAs) and long non-coding RNAs (lncRNAs), play essential roles in cancer development by influencing gene expression and cellular processes. An improved understanding of how ER stress and ncRNAs interact in breast cancer can potentially lead to new treatment approaches. Modifying specific ncRNAs involved in the ER stress response might interfere with cancer cell survival and induce cell death. Additionally, focusing on UPR-associated proteins that interact with ncRNAs could offer novel therapeutic possibilities. Therefore, this review provides a concise overview of the interconnection between ER stress and ncRNAs in breast cancer, elucidating the nuanced effects of the UPR on cell fate and emphasizing the regulatory roles of ncRNAs in breast cancer progression.
Similar content being viewed by others


Introduction
Non-coding RNAs (ncRNAs) are RNA molecules that play crucial roles in various cellular processes. They are transcribed from DNA, similar to messenger RNAs (mRNAs), but instead of being translated into proteins, they function as RNA molecules. There are several classes of ncRNAs, each with distinct structures and functions. Some well-known classes include microRNAs (miRNAs), Long Non-coding RNAs (lncRNAs), Small Interfering RNAs (siRNAs), and circular RNAs (circRNAs). miRNAs are small ncRNAs that regulate gene expression by binding to target messenger RNAs, while lncRNAs are longer ncRNAs that regulate gene expression at various levels [1]. siRNAs are double-stranded ncRNAs that guide the silencing machinery to degrade complementary target RNAs or inhibit their translation. circRNAs form covalently closed circular structures and can act as miRNA sponges, sequestering miRNAs and modulating gene expression. The roles of ncRNAs in development, cancer, neurological disorders, and other diseases are actively investigated, with significant potential as diagnostic markers and therapeutic targets in various fields of medicine and biotechnology [2,3,4]. Evidence suggests that endoplasmic reticulum stress (ER stress) can affect the expression and activity of specific ncRNAs in breast cancer cells [5,6,7]. The ER is a vital cellular organelle that plays a crucial role in protein synthesis, folding, and maintaining cellular homeostasis. ER stress can lead to protein homeostasis disorder, resulting from factors like disturbances in calcium homeostasis, nutrient deprivation, oxidative stress, viral infection, or genetic mutations. The ER regulates proteostasis through processes like the UPR, the ubiquitin–proteasome system (UPS), and autophagy. The UPR increases the production of chaperone proteins, reduces protein synthesis, and activates ER-associated degradation (ERAD). When ER stress exceeds the cellular capacity, it can cause cellular dysfunction and contribute to diseases like neurodegenerative disorders, metabolic diseases, and cancer [8,9,10]. Studies have shown that UPR can modulate the expression of miRNAs involved in breast cancer progression. Dysregulation of certain miRNAs can impact cancer cell survival, proliferation, and migration [11]. Furthermore, some lncRNAs, which are regulators of UPR genes, can interact with UPR-associated proteins and affect the expression or activity of ER stress-related genes [12]. Attaining a comprehensive comprehension of the intricate interplay between ncRNAs and ER stress within the context of breast cancer holds the potential to revolutionize treatment strategies significantly. This knowledge possesses the capability to unveil novel therapeutic targets, fostering the development of personalized approaches grounded in distinctive molecular signatures. Enhancing our understanding of how ncRNAs specifically impact ER stress response pathways could enhance the efficacy of current treatments, sensitize cancer cells to standard therapies, and address resistance mechanisms. Additionally, the identification of diagnostic and prognostic biomarkers within the ncRNA-ER stress axis can inform treatment decisions and monitoring processes. Integrating ncRNA-targeted therapies with conventional methods may yield synergistic effects, and manipulating the tumor microenvironment shows promise for augmenting responses to immunotherapies. Ultimately, these further discoveries may play a crucial role in the long-term management of the disease, offering new avenues to sustain control and prevent relapse in breast cancer. The objective of this review is to explore the interaction between ncRNAs and ER stress response pathways in breast cancer cells.
Role of non-coding RNAs in the pathogenesis of breast cancer
In the year 2020, breast cancer emerged as the most commonly diagnosed cancer on a globally, registering over 2.3 million new cases and resulting in 685,000 associated deaths. Notably, a majority of these cases were documented in transitioned countries, although a disproportionately high number of breast cancer-related deaths also occurred in transitioning countries. The terms “transitioning”, and “emerging” are used interchangeably to describe nations classified as low or medium Human Development Index (HDI). Conversely, “transitioned” is used to refer to those classified as high or very high HDI. Projections indicate a significant rise in the burden of breast cancer in the coming years, with estimations surpassing 3 million new cases and 1 million deaths by the year 2040. These projections raise serious concerns, emphasizing the critical need for the implementation of comprehensive and effective global policies aimed at combating the devastating consequences of breast cancer and mitigating its escalating impact [13]. The etiological underpinnings of breast cancer are inexorably intertwined with the dysregulation of ncRNAs, a phenomenon that has captured the attention of researchers in recent years [14]. ncRNAs, when their expression deviates from the normal levels, play a crucial role in regulating essential cellular processes such as cell proliferation, apoptosis, migration, and invasion, actively contributing to the initiation and progression of breast cancer [15, 16]. Extensive research efforts have been made to unravel the functional relevance of ncRNAs in the pathogenesis of breast cancer. In a 2019 study, researchers identified a significant overexpression of miR-21 in the breast in patient with breast cancer. Additionally, a reduction in the expression of programmed cell death protein 4 (PDCD4) was observed in these patients. The PDCD4 gene can inhibit tumor growth and promote cell death via the Sp1 transcription factor. The increased expression of miR-21 was found to accelerate mammary cell transformation and the growth of mammary tumors by inducing translational suppression of PDCD4 [17]. In 2023, Huang, Xie et al. identified elevated levels of miR-1260b and reduced mRNA expression of secretory coil domain-containing protein 134 (CCDC134) in human breast cancer tissue and a panel of human breast cancer cell lines. Silencing miR-1260b resulted in a decrease in the migration and invasion capabilities of breast cancer cells. Furthermore, the findings demonstrated that upregulating CCDC134 hindered the motility and invasion of breast cancer. These results suggest that miR-1260b functions as an oncogene in breast cancer, potentially promoting the motility and invasion of BRCA cells by suppressing the target gene CCDC134 and activating the MAPK signaling pathway. Additionally, it may play a role in suppressing immunological function and enhancing the immune system [18]. Furthermore, lncRNAs play a vital role in promoting the growth of breast cancer. A study in 2018 revealed a novel connection between the lncRNA LINC00538 (YIYA) and cancer metabolism in breast cancer. In a cell cycle-independent manner, YIYA interacts with cytosolic cyclin-dependent CDK6 kinase and controls CDK6-dependent phosphorylation of fructose bisphosphatase PFK2 (PFKFB3). These events increase the catalysis of glucose 6-phosphate to fructose-2,6-bisphosphate/fructose-1,6-bisphosphate in breast cancer cells. Silencing YIYA or CDK6 by CRISPR/Cas9-mediated deletion can disrupt glycolysis and tumor development in vivo. Approximately 40% of clinical breast cancer samples showed the presence of YIYA, which is associated with poor survival outcomes and CDK6 expression. These findings underscore the functional significance of lncRNAs in metabolic reprogramming [19]. A recent study highlighted that lncSNHG5 is significantly expressed in breast cancer-associated fibroblasts (CAFs) and plays a crucial role in the formation of the pre-metastatic niche. This is achieved by enhancing angiogenesis and vascular leakage through the regulation of Zinc finger protein 281 (ZNF281) in CAFs. The interaction between lncSNHG5 and the IGF2BP2 m6A reader contributes to the increased stability of ZNF281 mRNA. In endothelial cells, ZNF281 is responsible for the transcriptional control of CCL2 and CCL5 expression, as well as the activation of MAPK p38 signaling. The overexpression of CCL2 and CCL5 is associated with tumor metastasis and a poor prognosis in patients. Ultimately, the study findings underscore the critical role of the lncSNHG5-ZNF281-CCL2/CCL5 signaling axis in promoting the establishment of the pre-metastatic niche, thereby facilitating breast cancer metastasis [20]. circRNAs, akin to miRNAs and lncRNAs, play an integral role in the pathophysiology of breast cancer. Research has demonstrated significant expression of circSKA3 in breast cancer tissues and cells, directly correlating with the invasive capacity of breast cancer cells through the formation of invadopodia. The binding partners of circSKA3 in invadopodia derived from tumors were identified as Tks5 (tyrosine kinase substrate with 5 SH3 domains) and integrinb1. Furthermore, abnormal expression of circSKA3 was found to enhance tumor invasiveness both in vitro and in vivo [21]. In 2022, Qi et al. uncovered the involvement of circ_RPPH1 and Rho GTPase activator protein 1 (ARHGAP1), coupled with the decreased expression of miR-542-3p, in breast cancer (BC) tissues of cancer patients. The knockdown of circ_RPPH1 or the introduction of miR-542-3p resulted in a reduction in breast cancer cell growth and metastasis, accompanied by the induction of apoptosis. circ_RPPH1 acts by inhibiting miR-542-3p, which, in turn, regulates ARHGAP1 expression, thereby influencing the progression of breast cancer. Additionally, the downregulation of circ_RPPH1 was observed to inhibit tumor growth in vivo [22]. In conclusion, the findings from these studies highlight the intricate involvement of ncRNAs in essential cellular processes like proliferation, apoptosis, migration, and invasion, playing an active role in the onset and advancement of breast cancer. By comprehending the impact of these ncRNAs on diverse pathways, valuable insights can be gained regarding potential targets for therapeutic interventions and diagnostic markers in breast cancer.
Role of the endoplasmic reticulum stress in the pathogenesis of breast cancer
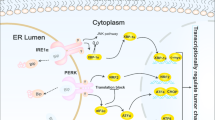
The ER is a cellular organelle responsible for protein synthesis and homeostasis. ER stress occurs when the ER struggles to fold proteins correctly, resulting in the buildup of misfolded proteins. To counter this, the UPR is activated as a cellular mechanism to restore equilibrium (Fig. 1). Under normal conditions, cells employ a process known as the UPR to manage stress within the ER. The UPR mechanism involves several strategies to address ER stress effectively. Firstly, it promotes the synthesis of chaperone proteins, which aid in the proper folding of newly synthesized proteins, ensuring their functionality. Additionally, the UPR reduces the overall production of proteins, alleviating the burden on the ER. This reduction in protein synthesis helps restore ER homeostasis. Moreover, the UPR activates a process called ERAD. ERAD is responsible for identifying and eliminating proteins that fail to fold correctly, preventing the accumulation of misfolded or dysfunctional proteins within the ER. Overall, the UPR acts as a protective mechanism to maintain ER function and cellular homeostasis in response to ER stress [8, 9]. The UPR is regulated by three primary sensors: inositol-requiring enzyme 1 (IRE1), protein RNA-activated kinase (PKR)-like ER kinase (PERK), and activating transcription factor 6 (ATF6). Its primary role is to restore normal functioning of the endoplasmic reticulum (ER) by repairing or degrading proteins. However, if the stress persists, it can trigger cell death, including apoptosis, and contribute to the development of diseases such as cancer. ER stress-induced apoptosis serves as a crucial cellular mechanism for eliminating damaged cells, maintaining protein balance, and ensuring proper tissue development. Understanding and effectively managing this process are essential in various scenarios. Targeting the UPR, particularly the aspect related to ER stress-induced apoptosis, holds potential as a therapeutic strategy, particularly in conditions like cancer [10, 23]. ER stress can occur in cancer cells because of their rapid growth, insufficient nutrients, and improper protein folding. This susceptibility can be exploited for targeted therapy, aiming to kill cancer cells while sparing healthy ones. Proteasome inhibitors and combination therapies disrupting protein folding and ER function are among the medications and therapeutic approaches designed to induce ER stress and promote apoptosis in cancer cells [24]. Evidence from studies suggests that the upregulation of UPR components contributes to the proliferation, progression, and chemotherapy resistance in breast cancer cells [25]. The activation of PERK-ATF4 is pivotal for the advancement and rapid growth of breast cancer cells, as supported by experimental results obtained both in vitro and in vivo by Feng in 2017 [26]. Dysregulation of the PERK arm leads to the modification of signaling pathways, establishing an adaptive mechanism that enables cell survival in challenging circumstances. This suggests that disruptions in the normal functioning of the PERK pathway prompt cells to adapt and find alternative means to ensure survival in unfavorable environments or circumstances. Understanding these adaptive mechanisms is crucial, especially in diseases like cancer, where these changes may contribute to the advancement and resilience of cancer cells [27, 28]. Recent studies have shown that inhibiting the PERK pathway can increase the susceptibility of breast cancer cells to apoptosis, indicating its potential as a treatment strategy. However, it is important to note that separate research has indicated that ATF4 can independently influence the regulation of autophagy, irrespective of PERK. This suggests that while inhibiting PERK may promote apoptosis, the involvement of ATF4 and autophagy regulation adds complexity, requiring a comprehensive understanding of the overall cellular response in breast cancer. The interplay between these pathways highlights the intricate nature of cellular processes and the challenges in developing targeted therapies [29]. The activation of PERK-ATF4 in autophagy has the potential to tilt the balance from pro-survival signals to pro-death signals, disrupting the equilibrium of apoptosis [30]. Other components of the UPR include chaperones like GRP78 (BiP) and GRP94 (gp96). GRP78, a crucial molecular chaperone, aids in the folding, assembly, and secretion of proteins within the ER, playing a vital role in the UPR. Another ER-resident chaperone, GRP94, interacts specifically with secretory and transmembrane proteins. Notably, GRP94’s immunogenic characteristics have led to its exploration in immunotherapy for diseases such as cancer. Overexpression of both GRP78 and GRP94 is often observed in malignancies, including breast cancer, highlighting their significance in disease development. Given their role in maintaining protein homeostasis and their association with diseases characterized by protein misfolding, targeting these chaperones presents a promising therapeutic approach [31, 32]. In the absence of GRP78, which is an ER chaperone protein, CHOP initiates a notable impact on impeding the formation of malignant tumors. This is achieved by triggering cell death in precancerous cells, and the mechanism involves the downstream modulation of the PERK signaling pathway. The disruption of the GRP78 gene, leading to the activation of CHOP and subsequent modulation of PERK signaling, acts as a deterrent to the development of cancerous growths, emphasizing the intricate relationship between ER stress response, cell survival, and tumorigenesis [33]. The PERK signaling pathway exhibits a dual role, capable of both promoting and suppressing tumor progression. In the context of breast tumors, this pathway holds the potential to impede tumor progression. However, it is crucial to note that prolonged activation of the PERK pathway may lead to the induction of breast cancer by fostering genomic instability [34]. Abundant research has accumulated compelling evidence affirming that therapeutic interventions targeting PERK, GRP78, and IRE1/XBP1 have substantial potential to enhance the effectiveness of breast cancer treatment. The utilization of inhibitors directed at these proteins, combined with drugs inducing resistance through the UPR, has shown promise in reducing the growth of breast tumors in vivo [35]. ER stress exerts effects beyond the proteins directly involved in its pathway. A 2014 study demonstrated that inhibiting ER stress-induced heparinase expression in breast cancer cells yields positive outcomes, reducing tumor invasion and metastasis. This inhibition also upregulates CCL5 expression, subsequently reducing transmigration. Notably, CCL5 is recognized for inducing tumor angiogenesis, impacting tumor cell motility, invasion, and metastasis [35]. Certainly, ER stress exhibits a dual role in breast cancer progression. Firstly, it can hinder breast cancer development by triggering cell death pathways during prolonged stress conditions. Conversely, ER stress sustains the progression of breast cancer by fostering adaptive mechanisms that aid the survival of cancer cells. This dual nature emphasizes the significance of targeting ER stress in therapeutic interventions for breast cancer treatment. Approaches focused on modulating ER stress-induced pathways, including the inhibition of PERK or other UPR components, show promise in improving treatment effectiveness and potentially restraining breast cancer metastasis.

Endoplasmic reticulum (ER) stress is the result of an imbalance or disturbance in the normal functioning of the ER, a cellular organelle involved in protein production and folding. An excess of misfolded proteins, a calcium imbalance, a viral infection, or dietary deficiency can all contribute to this. When ER stress occurs, a cellular signaling cascade called the unfolded protein response (UPR) is triggered. It aims to reduce stress and return to ER equilibrium. IRE1, PERK, and ATF6 are the three primary branches of the UPR that are mediated by distinct ER transmembrane proteins. IRE1 pathway: IRE1 splices and processes XBP1 mRNA to produce a transcription factor that activates genes related to ER-associated degradation and protein folding. PERK pathway: eIF2α is phosphorylated by PERK, which causes an overall decrease in protein synthesis. It preferentially stimulates the translation of ATF4, which controls the expression of genes related to the metabolism of amino acids, apoptosis, and antioxidant responses. ATF6 pathway: after being delivered to the Golgi apparatus, ATF6 cleaves and releases a fragment of a transcription factor. After translocating to the nucleus, this fragment activates genes related to lipid metabolism, ER-associated degradation, and protein folding. Stress reduction and ER function restoration are the primary objectives of the UPR. Normality is restored to cellular activities upon resolution of the stress. On the other hand, apoptotic mechanisms may cause cell death in response to extended or severe ER stress. Created by https://www.biorender.com/
Non-coding RNAs and ER stress
Non-coding RNAs involved in the folding and modification of proteins
Proteins must be accurately folded into precise, three-dimensional forms to function effectively, which is an essential biological activity. The ER is a cellular compartment dedicated to the process of protein folding. As noted, the failure of proteins to fold correctly leads to ER stress. Calreticulin (CALR), BIP/GRP78, and protein disulfide isomerases (PDIs) are indeed vital chaperone proteins located in the ER lumen. They play critical roles in preserving protein homeostasis, ensuring the proper folding, and maintaining the quality control of newly synthesized proteins [36]. To investigate the role of miR-455 in the protective effect of H2S on lung epithelial cells against CoCl2-induced apoptosis through the modulation of ER stress-related genes, experiments were performed using human lung epithelial cells (BEAS-2B). These experiments included exposing the cells to hypoxic injury, both with and without prior H2S preconditioning. Researchers identified that CALR, GRP78, CHOP, and Caspase-12 protein expression were all downregulated by the miR-455 mimic, whereas their expression was increased by the miR-455 inhibitor [37]. In another study, it has been shown that miR-663a may act as a CALR regulator in patients with rectal cancer [38]. GRP78 and CALR, two ER stress indicators, are downregulated by miR-124 in angiotensin II (AngII)-induced myocardial hypertrophy. This suggests that the observed inhibition of cardiac hypertrophy resulting from miR-124 downregulation may be attributed to the reduction of ER stress [39]. Based on the data currently available, miR-455 has binding sites in CALR mRNA, and microRNA sequencing discovered that miR-455 is downregulated in the hearts of mice with constitutively active ATF6. Additionally, the induction of the UPR with tunicamycin or the overexpression of constitutively active ATF6 in neonatal rat ventricular myocytes (NRVMs) results in reduced levels of miR-455 and increased expression of CALR. Furthermore, CALR abundance is reduced as a result of overexpressing pre-miR-455, leading to elevated levels of miR-455. Conversely, CALR abundance increases when miR-455 antagonistic oligonucleotides are transfected, thereby reducing their levels. This indicates that, by decreasing the levels of miR-455, ER stress at least partially contributes to the activation of CALR [40, 41]. Soft tissue sarcomas, known as liposarcomas, are characterized by poor adipocyte development, especially in the dedifferentiated subtype. The dysregulation of CALR, possibly regulated by miR-1257, may contribute to the inhibition of adipocyte development, potentially explaining the dedifferentiated phenotype observed in liposarcomas [42]. Regulation of BIP is essential for maintaining ER Ca2+ homeostasis and protein folding because it is a master regulator of the UPR, which is vital for ER Ca2+ storage [43]. Heat shock 70 kDa protein 5 (HSPA5) encodes the protein GRP78. Recent research has revealed a significant presence of GRP78 on the cell surface of cancer cells. Consequently, GRP78 has emerged as a crucial regulator of signaling pathways associated with tumor cell viability. This discovery has prompted the investigation of therapeutic strategies targeting cell surface GRP78 for cancer treatment [44]. In laboratory experiments conducted both in vitro and in vivo, the introduction of a miR-181a mimic has been shown to decrease GRP78 protein expression, indicating negative regulation by miR-181a. Conversely, using a miR-181a inhibitor or antagomir results in an increase in GRP78 protein expression, implying that inhibiting miR-181a allows for higher levels of GRP78 protein production [45]. miR-30d, miR-181a, and miR-199a-5p collaborate to control GRP78 levels by destabilizing GRP78 mRNA, resulting in reduced levels at both the mRNA and protein levels. This cooperative action is a common phenomenon in gene regulation, where multiple miRNAs fine-tune the expression of target genes. Further research is needed to comprehend the precise mechanisms underlying the destabilization by these miRNAs and the functional significance of this regulatory network in various biological processes and disease conditions [46]. According to reports, overexpressing lncRNA RP11-115N4.1 dramatically reduces K562 cell proliferation and modifies the immunological response by triggering HSP70 production by binding to HNRNPH3 [47]. The results strongly suggest that the lncRNA Hotair functions as a positive regulator in laryngeal squamous cell carcinoma by influencing the stability of GRP78 through posttranscriptional modifications, coordinated by the regulation of hsa-miR-30a-5p. Moreover, Hotair facilitates immune evasion by upregulating the expression of PD-L1 through the hsa-miR-30a-5p/GRP78 pathway [48]. Overexpression of miR-204 results in a decrease in the activation or expression of specific genes responsive to ER stress, namely GRP78, GRP94, and CHOP. This reduction in gene induction is associated with the phenotypic characteristics typically observed in senescent cells [49]. There is a strong association between HOTAIR and HSPA1A, a member of the heat shock protein family A (Hsp70), in breast cancer (BRCA) tissues. HOTAIR was found to elevate HSPA1A levels in BRCA cells after radiation exposure, affecting both mRNA and protein levels. miR-449b-5p, which normally inhibits HSPA1A production by binding to the 3′-UTR region of HSPA1A mRNA, is responsible for this action [50]. ER stress in the cardiovascular system is induced by reduced expression of miR-30 family miRNAs, leading to increased production of GRP78. This disruption in the regulatory activity of miR-30 family miRNAs hinders the normal physiological response to ER stress, impacting the balance of protein folding and processing. Ultimately, this imbalance may contribute to the development or progression of cardiovascular disorders [51]. Another study showed that NEAT1 expression is positively correlated with GRP78, but there is no further evidence of their interaction [52]. Another factor in protein folding is PDIs, which are regulated by microRNAs. PDIs, a class of oxidoreductases primarily located in the ER, play crucial roles in protein folding and maturation by catalyzing disulfide bond formation, isomerization, and reduction of nascent proteins [53]. Protein disulfide isomerase family A member 6 (PDIA6), also known as P5, is an enzyme belonging to the PDI family. PDIA6 plays a critical role in catalyzing protein folding processes, displaying both isomerase and chaperone activities. Interestingly, PDIA6 is downregulated during cellular senescence in bone marrow-derived mesenchymal stem cells (BMSCs) [54]. In 2023, Huang et al. demonstrated that in H2O2-treated human foreskin fibroblasts (HFF), the expression of PDIA6 is reduced compared to the control group. Interestingly, inhibition of miR-181a alleviates oxidative stress induced by H2O2 and reduces cellular senescence in HFF. Moreover, when PDIA6 is knocked down in combination with the downregulation of miR-181a, the inhibitory effects on cellular senescence and oxidative stress are reversed. This suggests that silencing miR-181a can mitigate H2O2-induced cellular senescence and oxidative stress by targeting PDIA6. Additionally, the findings suggest that PDIA6 may play a protective role in HFF, shielding them from cellular senescence induced by H2O2 [55]. The UPR pathway responds to disturbances in calcium homeostasis and is regulated by the ER oxidoreductase PDIA6 and miR-322. PDIA6 interacts with IRE1α, enhancing its activity by increasing phosphorylation and splicing of XBP1 mRNA. PDIA6 has minimal effects on other pathways involved in the cellular response to ER stress. Depletion of ER Ca2+ levels result in decreased levels of miR-322, leading to increased stability of PDIA6 mRNA and enhanced IRE1α activity during the ER stress response [56]. Regardless of the factors involved, ncRNAs contribute to the intricate regulatory networks governing protein folding and modification. The complexity of these processes and the importance of ncRNAs in maintaining protein homeostasis and cellular function are underscored by their involvement in multiple pathways of action. Understanding the role of ncRNAs in protein homeostasis involves examining their functions in related pathways, particularly those involved in cellular stress responses such as the UPR. Here’s a closer look at how ncRNAs function in these pathways.
Non-coding RNAs in ER stress
Observations from mammalian cells suggest that ncRNAs play a role in determining ER stress, and, reciprocally, ER stress also controls ncRNA expression [19] (Fig. 2). Numerous studies have explored the impact of ncRNAs on ER stress in various disorders, and we have previously provided an overview of ncRNAs involved in protein folding. XBP1 regulates miR-346 and miR-153 during ER stress, while certain miRNAs such as miR-34c, miR-665, and miR-30c directly target XBP1 mRNA [57]. The study indicates that miRNAs play a role in hindering the transition from cellular survival to programmed cell death. Predominantly, miRNAs implicated in cell death induced by ER stress are located within the PERK–eIF2α–CHOP signaling pathway. miRNAs such as miR-17, miR-34a, miR-96, and miR-125b are crucial in orchestrating the transition of the PERK–eIF2α–CHOP axis towards cell death in response to ER stress. This underscores the significance of these miRNAs in the context of cancer-related cell death [58]. Several other miRNAs, including miR-24, miR-29, miR-195, miR-221, miR-222, miR-346, and miR-665, have been linked to cell death induced by ER stress through their targeting of various factors [59,60,61,62,63,64]. miR-410 plays a regulatory role in ER stress, facilitating tumor progression by suppressing cellular migration and invasion in breast cancer cells. This effect is achieved through the specific modulation of gene expression, including CHOP, GRP94, Bip, and p-PERK [65, 66]. miR-30b-5p and miR-30c-5p have been observed to target the eIF2α protein, promoting cell survival in hepatocellular carcinoma (HCC) and breast cancer when treated with PS-341 [67]. PS-341 is known to impede ERAD and result in the accumulation of misfolded proteins [68]. The expression of PD-L1 in macrophages, promoting immune evasion of cancer cells, is activated by cancer-derived exosomal microRNAs (exo-miRs) induced by ER stress. This discovery establishes a novel connection between cancer and ER stress, elucidating a previously unknown mechanism [69]. Other ncRNA types, such as snoRNAs and siRNAs, may play a role in regulating ER stress. However, the specific roles and mechanisms of action of these ncRNAs in ER stress are still being explored. In general, ncRNAs are crucial for controlling ER stress and preserving protein homeostasis. They play a vital role in the adaptive response to ER stress, preventing cellular malfunction and disease by regulating the expression of critical genes involved in ER stress signaling pathways.

A model to depict the interaction between non-coding RNAs and ER stress
Non-coding RNAs as biomarkers in UPR
ncRNA molecules has become crucial in pinpointing biomarkers indicative of disease development and progression. The field of RNA therapeutics is undergoing a transformative phase with notable progress in oligonucleotide techniques and pharmacological discoveries. Targeting RNA structures and RNA–protein interactions with small molecules presents a promising avenue for developing the next generation of drugs. These advancements enable precise modulation of gene expression, offering potential therapeutic interventions at the genetic level [70]. Recently emerging findings highlight the pivotal roles of ncRNAs, including miRNAs and lncRNAs, in regulating UPR signaling pathways and shaping cellular responses to ER stress. These ncRNAs act as highly sensitive indicators of UPR irregularities, providing insights into the intricate molecular mechanisms of stress-related conditions like neurodegenerative disorders and specific cancers. Investigating the distinct signatures of ncRNAs associated with UPR activation or malfunction allows researchers to establish these molecules as reliable biomarkers for monitoring cellular stress and tracking disease progression. Effectively communicating the significance of ncRNAs in the UPR framework not only enhances our understanding of cellular stress responses but also presents significant opportunities for advancing diagnostic and therapeutic approaches in disorders linked to UPR dysregulation. Table 1 compiles various ncRNAs that undergo alterations when subjected to ER stress induction and UPR activation, providing insights into their functional mechanisms.
Non-coding RNAs involved in ER stress in breast cancer
As mentioned, the UPR is initially activated in response to ER stress as a protective mechanism to safeguard cells from the adverse effects of misfolded proteins within the ER. It serves as a cellular strategy to restore normal homeostasis. However, if the UPR fails to restore the cell’s equilibrium, it can ultimately trigger cell death [71]. Therefore, it seems logical to consider the UPR as a potential tumor-suppressive system. Nevertheless, the role of ER stress and the UPR in cancer remains a subject of intense debate due to numerous pieces of evidence suggesting that, in ER-stressed cancer cells, the UPR may promote tumor development [72]. Also, some specific ncRNAs can influence the UPR signaling pathway; conversely, UPR components can boost the production of certain ncRNAs. This connection is particularly relevant in cancer studies, offering potential insights for effective tumor control strategies [6, 73] (Table 2). In 2019, Zhang et al. demonstrated that overexpression of the lncRNA MEG3, also known as GTL2, FP504, or LINC00023, significantly inhibits breast cancer cell growth both in vitro and in vivo by increasing apoptosis in breast cancer cells. MEG3 was found to enhance the expression of proteins associated with the ER stress response, including GRP78, IRE1, PERK, and ATF6. Additionally, MEG3 increased the levels of pro-apoptotic proteins, namely CHOP and caspase-3 [7]. This suggests that ER stress-triggered apoptosis mediated by MEG3 may be a consistent phenomenon observed across different cancer types [74, 75]. In another study, the impact of inducing ER stress on the expression of lncRNAs is observed. ER stress induction, through the activation of the GRP78/OCT4/lncRNA MIAT/AKT pathway, is shown to lead to drug resistance in breast cancer cells. The commonly used chemotherapy drug, 5-fluorouracil (5-FU), triggers ER stress, resulting in the upregulation of key ER stress proteins and the activation of the regulatory pathway mentioned above. This, in turn, promotes the proliferation of breast cancer cells, supporting tumor progression [76]. As highlighted in this review, XBP1s serve as a crucial transcription factor regulating various genes related to ER homeostasis, cell viability, angiogenesis, metastasis, and drug resistance. This factor is generated through the activation of the IRE1 pathway, leading to the excision of a 26-nucleotide intronic segment. The splicing rate of XBP1 (XBP1 spliced/unspliced) is significantly increased in malignant breast tissue compared to non-malignant tissue. Notably, the upregulation of lncRNAs NORAD, NEAT1, and LINC00299 is linked to an elevation in the splicing rate of XBP1. Conversely, the expression of the lncRNA CASC2 exhibits a negative correlation, indicating that an increase in CASC2 gene expression is associated with a reduction in the splicing rate of XBP1 [77]. Furthermore, induced ER stress in breast tumor tissue appears to significantly influence cancer progression through its impact on miRNA expression. This induction leads to the enhanced exosomal expression and secretion of miR-27a-3p, consequently promoting increased immune evasion in breast cancer. MiR-27a-3p achieves this effect by upregulating PD-L1 expression, targeting the MAGI2/PTEN/PI3K axis [78]. Moreover, the primary transducer of the UPR, IRE1, disrupts de novo miRNA biosynthesis and maturation through the RIDD process. The RNase activity of IRE1 results in the degradation of tumor suppressor miRNAs such as miR-3607-3p, miR-374a-5p, and miR-96 in breast cancer cells. This pathway amplifies the expression of the miR-3607-3p target, the oncogenic factor RAB3B, establishing a positive association with cancer progression, invasion, and metastasis [79]. Liang et al. demonstrated that the hypoxia-induced ER stress/IRE1α/XBP1 pathway upregulates miR-153 expression in breast cell lines. miR-153 inhibits tumor angiogenesis by suppressing the HIF1α/VEGFA axis through binding to the 3′-UTR of HIF1A mRNA in breast cancer cells [80]. UPR-induced miR-616 is another miRNA that acts as a tumor suppressor in breast cancer by downregulation of c-MYC [81]. As mentioned above, the interaction between ER stress, ncRNAs, and the prognosis of breast cancer is complex and controversial. In certain pathways, the association between ER stress-related proteins and ncRNA expression aligns with the development of breast cancer, and other pathways following the suppression of tumor.
Therapeutic strategies based on non-coding RNAs and the ER stress in breast cancer
Current methods for managing breast cancer are diverse and include radiation therapy, cytotoxic-based chemotherapy, surgical mastectomy, hormone therapy, and treatment with monoclonal antibodies [82]. While chemotherapy drugs like carboplatin, paclitaxel, and doxorubicin have been discovered for breast cancer treatment, they come with limitations such as poor oral bioavailability, drug resistance, inefficient delivery, and high toxicity. Therefore, the development of anti-cancer drugs without side effects is crucial [83,84,85]. Researchers have shifted their focus to studying alternative biological targets and novel signaling pathways that can effectively inhibit breast cancer growth and metastasis [86]. Numerous studies have indicated that ncRNAs exhibit different expressions in various stages of development and pathological conditions, including breast cancer [87,88,89,90]. This suggests that ncRNAs may serve as an alternative treatment for cancer prevention and management in the future.
ncRNAs and drug resistance in breast cancer
Despite the availability of several treatment methodologies, overcoming treatment tolerance remains a significant challenge in improving the clinical outcome of breast cancer [91]. Drug resistance poses a major obstacle to chemotherapy-based systemic treatment in metastatic and advanced breast cancer, contributing to lower recurrence-free survival rates in breast cancer patients [92, 93]. Previous studies have highlighted the involvement of ncRNAs, mainly long lncRNAs and miRNAs, in drug resistance in breast cancer [94]. For instance, miR-451 and miR-326 regulate drug resistance by controlling the expression of multidrug resistance 1 (MDR1) [95]. Other studies have shown that altered expression of miR-21 and miR-298 is associated with chemoresistance in breast cancer cells, targeting PTEN and MDR1, respectively [96, 97]. Additionally, miR-221/222 is linked to increased resistance to tamoxifen by targeting p27Kip1 in breast cancer [98, 99]. Investigations into trastuzumab-resistant breast cancer cells revealed that miR-200c downregulation leads to enhanced epithelial-mesenchymal transition (EMT) and tumorigenesis [100]. Moreover, miR-218 induces cisplatin resistance by targeting BRCA1 in breast cancer patients [101]. Long-term exposure to doxorubicin upregulates the intronic lncRNA Adriamycin Resistance-Associated (ARA) in doxorubicin-resistant MCF7 cells, contributing to inhibition of proliferation and increased drug sensitivity upon ARA deletion [102]. H19, a lncRNA, counteracts the downregulation of estrogen receptor α (ERα) protein induced by endocrine therapy, promoting treatment resistance in breast cancer cells [103]. Additionally, hypoxia-driven modulation of tumor suppressor ncRNAs and upregulation of carcinogenic ncRNAs play a pivotal role in creating resistance to various therapeutic agents in breast cancer cells. For instance, hypoxia inhibits miR-873-5p, increasing the expression of drug resistance-related targets such as MDR1 and pregnane X receptor (PXR) [104]. Hypoxia-induced inhibition of miR-326 expression, in a HIF1α-dependent manner, increases ITGA5 expression, resulting in chemotherapy resistance in triple-negative breast cancer (TNBC) cells [105]. Furthermore, hypoxia-induced miR-424 targets the apoptosis-related tumor suppressor PDCD4, conferring resistance to chemotherapies like doxorubicin and etoposide [106]. These examples illustrate the intricate involvement of ncRNAs in treatment resistance in breast cancer. Researchers continue to explore the molecular mechanisms of ncRNAs, their potential as biomarkers for predicting response, and as targets for overcoming drug resistance. Staying updated with the latest scientific findings is crucial for a comprehensive understanding of this complex issue as the field of ncRNA research evolves.
ncRNAs as possible biomarker candidates for diagnosis and prognosis in breast cancer
The utilization of ncRNAs as therapeutic agents can be characterized by two criteria: (i) dysregulation of ncRNA in cancer cells compared to normal cells and (ii) alteration in the phenotype of cancer cells through the targeting of ncRNA expression [107, 108]. Reports indicate that miRNAs in serum can serve as diagnostic markers in patients. For instance, the down-regulation of let-7 family miRNAs is associated with a poor prognosis [107]. High levels of the miR-106b-25 cluster are significantly associated with shorter time to recurrence in breast cancer [109]. miR-181a and miR-221/miR-222 clusters are directly related to tumor progression and have diagnostic and prognostic value [110, 111]. Furthermore, reduced levels of miR-1247-5p are associated with age, tumor size, and a poor disease prognosis. Frères et al. demonstrated that the levels of miR-34a and miR-122 in both plasma and tumor tissue of breast cancer patients increased following neoadjuvant chemotherapy (NAC), particularly after anthracycline treatment. Subsequently, they employed a diagnostic test based on eight circulating miRNAs, namely let-7d, miR-103, miR-16, let-7i, miR-19b, miR-107, miR-148a, and miR-22, as an alternative to mammography for the diagnosis of breast cancer [112]. On the other hand, circRNAs, being stable biomarkers with high stability in serum, saliva, urine, milk, and exosomes, exhibit great potential for cancer diagnosis and monitoring disease progression. However, studies investigating their role in breast cancer are currently very limited [82, 113,114,115]. Analysis of The Cancer Genome Atlas (TCGA) data on 1097 breast cancer patients revealed significant differential expression of lncRNAs in various subtypes of breast cancer. In particular, 1510 lncRNAs were differentially expressed in normal versus TNBC samples, while 672 lncRNAs showed differential expression in non-TNBC versus TNBC samples. Among these, three upregulated lncRNAs (AP000924.1, AC091043.1, and FOXCUT) demonstrated promising potential as biomarkers for the diagnosis of TNBC. On the other hand, three other lncRNAs (AL354793.1, AC010343.3, and FGF10-AS1) were associated with prognosis in breast cancer patients. These findings highlight the relevance of lncRNAs in breast cancer and their potential utility as diagnostic and prognostic markers in TNBC [116]. But beyond the role of ncRNAs as a biomarker, ncRNAs have emerged as promising therapeutic agents in cancer treatment, with ongoing clinical trials demonstrating their diverse applications. miRNAs and small interfering RNAs (siRNAs) have garnered particular attention for their ability to precisely regulate gene expression, offering targeted therapeutic opportunities. Clinical trials investigating miRNA-based therapies have shown their value as diagnostic and prognostic markers, with some yielding promising results in intervention studies (ClinicalTrials.gov identifier: NCT05477667, NCT02247453, NCT03776630, NCT05346757). For example, the therapeutic efficacy of miR-34a has been demonstrated in advanced solid tumors, although challenges have been encountered, highlighting the importance of understanding potential adverse events. Furthermore, the recent approval of siRNA-based drugs by the FDA for non-cancerous diseases represents a significant advancement, opening doors for progress in cancer therapy [117]. Collectively, these investigations underscore the evolving landscape of ncRNA-based therapies, showcasing their potential across various cancer types and positioning them as promising candidates for future cancer management strategies.
Novel therapeutic strategies based on ncRNAs and UPR in breast cancer
As previously mentioned, ER stress influences the expression of ncRNAs, and together, they synergistically impact the development of breast cancer. Within the realm of ncRNAs, miRNAs assume a crucial role in regulating the drug sensitivity of breast cancer cells by modulating the UPR signaling pathways [11]. Studies have indicated that ER stress can be regarded as a side effect induced by anticancer drugs like cisplatin, etoposide, doxorubicin, gemcitabine, cytarabine, and vinorelbine [118]. On the other hand, hypoxia created in tumor conditions can not only act as a factor to activate ER stress [37] but also act as an inducer of EMT in cancer [119, 120]. Furthermore, hypoxia may regulate the expression of various ncRNAs in both HIF signaling-dependent and independent ways [120]. Given the role of hypoxia in cancer progression in solid tumors, such as breast cancer, directly targeting HIFs provides an excellent therapeutic option in clinical settings [105, 121]. Numerous HIF-targeting inhibitors are currently undergoing preclinical trials for validation, either as standalone treatments or in combination with other regimens, to combat various advanced cancers. The production of HIF-1alpha in the tumor microenvironment induces the upregulation of PD-L1 in both myeloid-derived suppressor cells and tumor cells. Consequently, this process can suppress immune system responses against the tumor, contributing to immune evasion [122]. Yang et al. demonstrated the potential involvement of HIF-1a and miR-210 signaling pathways in breast cancer invasion and metastasis, suggesting that targeting these pathways could represent a novel therapeutic strategy for breast cancer [123]. Glycolysis has emerged as a primary metabolic process supporting anabolic growth and energy production in cancer cells. In breast cancer cells, inhibiting glycolysis is being explored as a novel therapeutic approach to address hypoxia and drug resistance associated with mitochondrial respiratory defects [124, 125]. Among the methods of treating tumor cells, chemotherapy usually fails because tumor cells acquire multi-drug resistance and lead to various outcomes, including ER stress tolerance (ERST) [126,127,128]. In addition, ER stress induces resistance to tyrosine kinase inhibitors by upregulating Bcl-xL [129]. Overall, UPR activation has been demonstrated to contribute to drug resistance. Hence, the exploration of small molecule inhibitors targeting UPR components holds promise as a strategy to overcome ER stress-induced drug resistance and sensitize cancer cells to apoptosis [130]. The overexpression of XBP1 has been observed in various human cancers, including breast cancer. Targeting XBP1, a crucial component of the UPR and a significant nuclear transcription factor, is being considered as a therapeutic strategy [131]. It is important to acknowledge that the diverse functions of the UPR and the targeting of this pathway in clinical settings can potentially lead to unintended side effects. Therefore, additional studies are necessary to thoroughly investigate and assess the possibility of unwanted side effects when specifically targeting the UPR. These studies are crucial for ensuring the safety and efficacy of UPR-targeted approaches in therapeutic interventions.
Conclusion
The exploration of ncRNAs in recent years has illuminated their pivotal role as potential therapeutics in the management and progression of breast cancer. Current evidence underscores the initiation of the UPR by tumor cells in response to environmental changes, portraying the UPR as a pro-oncogenic mechanism influencing diverse aspects of breast cancer. This intricate relationship orchestrates the regulation of ncRNA levels in tumors, suggesting the potential use of UPR modulators as biomarkers in cancer drug therapy and prognosis. However, the downstream components of the UPR, under specific circumstances, exhibit a dual role—they not only regulate ER stress-induced apoptosis but also foster breast cancer proliferation by modulating ncRNA expression. Conversely, ncRNAs reciprocally influence the expression of downstream UPR target genes. In this review, we discussed the mutual regulation of ER and ncRNAs, emphasizing the need for further research, particularly in clinical studies, to elucidate the UPR’s role in tumorigenic mechanisms. While the promise of ncRNA-based therapies in cancer management is evident, it is essential to recognize associated limitations. Challenges include efficient delivery to target tissues and cells, necessitating ongoing investigation into safety and effectiveness. Precision in targeting specific ncRNAs within complex regulatory networks poses a significant challenge, requiring meticulous modulation to avoid disrupting normal cellular functions. Potential unintended consequences, such as off-target effects and interference with unintended pathways, warrant thorough preclinical validation. Moreover, addressing the dynamic nature of RNA molecules, including stability and pharmacokinetics, remains a challenge, necessitating strategies to enhance their presence and stability in targeted tissues. Despite these challenges, the ongoing efforts in the field of ncRNA-based therapies hold substantial promise for advancing breast cancer management toward more effective and tailored interventions.
Availability of data and materials
The datasets used during the current study are available from the corresponding author upon reasonable request.
Abbreviations
- ATF4:
-
Activating transcription factor 4
- BMSCs:
-
Bone marrow-derived mesenchymal stem cells
- BRCA1:
-
Breast cancer type 1 susceptibility protein
- CALR:
-
Calreticulin
- CHOP:
-
C/EBP homologous protein
- circRNA:
-
Circular RNA
- c-MYC:
-
Cellular myelocytomatosis oncogene
- EMT:
-
Epithelial–mesenchymal transition
- ER:
-
Endoplasmic reticulum
- ERAD:
-
ER-associated degradation
- ERST:
-
Endoplasmic reticulum stress tolerance
- GRP78:
-
Glucose-regulated protein 78
- GTL2:
-
Gene trap locus 2
- HIF1α:
-
Hypoxia-inducible factor 1 alpha
- HOTAIR:
-
HOX antisense intergenic RNA
- ICD:
-
Immunogenic cell death
- IRE1α:
-
Inositol-requiring enzyme 1 alpha
- JNK:
-
C-Jun N-terminal kinase
- lncRNAs:
-
Long non-coding RNAs
- MAPK:
-
Mitogen-activated protein kinase
- MDR1:
-
Multidrug resistance 1
- MEG3:
-
Maternally expressed gene 3
- miRNAs:
-
MicroRNAs
- NAC:
-
Neoadjuvant chemotherapy
- ncRNAs:
-
Non-coding RNAs
- NEAT1:
-
Nuclear-enriched abundant transcript 1
- NORAD:
-
Non-coding RNA activated by DNA damage
- P53:
-
Tumor protein 53
- PDIA6:
-
Protein disulfide isomerase family a member 6
- PDIs:
-
Protein disulfide isomerases
- p-eIF2α:
-
Phosphorylated eukaryotic translation initiation factor 2 alpha
- PI3K:
-
Phosphoinositide 3-kinase
- PKR:
-
Protein kinase R
- PTEN:
-
Phosphatase and tensin homolog
- PXR:
-
Pregnane X receptor
- RIDD:
-
Regulated IRE1-dependent decay
- TNBC:
-
Triple-negative breast cancer
- TRAF2:
-
Tumor necrosis factor receptor-associated factor 2
- TRAIL-R1/DR4:
-
Tumor necrosis factor-related apoptosis-inducing ligand receptor 1/death receptor 4
- UPR:
-
Unfolded protein response
- VEGFA:
-
Vascular endothelial growth factor A
- XBP1:
-
X-box binding protein 1
References
Kaikkonen MU, Lam MT, Glass CK. Non-coding RNAs as regulators of gene expression and epigenetics. Cardiovasc Res. 2011;90(3):430–40.
Yan H, Bu P. Non-coding RNA in cancer. Essays Biochem. 2021;65(4):625–39.
Krol J, Loedige I, Filipowicz W. The widespread regulation of microRNA biogenesis, function and decay. Nat Rev Genet. 2010;11(9):597–610.
Mercer TR, Dinger ME, Mattick JS. Long non-coding RNAs: insights into functions. Nat Rev Genet. 2009;10(3):155–9.
Yadav RK, Chae SW, Kim HR, Chae HJ. Endoplasmic reticulum stress and cancer. J Cancer Prev. 2014;19(2):75–88.
Wu M-Z, Fu T, Chen J-X, Lin Y-Y, Yang J-E, Zhuang S-M. LncRNA GOLGA2P10 is induced by PERK/ATF4/CHOP signaling and protects tumor cells from ER stress-induced apoptosis by regulating Bcl-2 family members. Cell Death Dis. 2020;11(4):276.
Zhang Y, Wu J, Jing H, Huang G, Sun Z, Xu S. Long noncoding RNA MEG3 inhibits breast cancer growth via upregulating endoplasmic reticulum stress and activating NF-κB and p53. J Cell Biochem. 2019;120(4):6789–97.
Wang M, Kaufman RJ. Protein misfolding in the endoplasmic reticulum as a conduit to human disease. Nature. 2016;529(7586):326–35.
Chen B, Retzlaff M, Roos T, Frydman J. Cellular strategies of protein quality control. Cold Spring Harb Perspect Biol. 2011;3(8): a004374.
Yan P, Ren J, Zhang W, Qu J, Liu GH. Protein quality control of cell stemness. Cell Regen. 2020;9(1):22.
Zhao T, Du J, Zeng H. Interplay between endoplasmic reticulum stress and non-coding RNAs in cancer. J Hematol Oncol. 2020;13(1):163.
Vijayalalitha R, Archita T, Juanitaa GR, Jayasuriya R, Amin KN, Ramkumar KM. Role of long non-coding RNA in regulating ER stress response to the progression of diabetic complications. Curr Gene Ther. 2023;23(2):96–110.
Arnold M, Morgan E, Rumgay H, Mafra A, Singh D, Laversanne M, et al. Current and future burden of breast cancer: global statistics for 2020 and 2040. Breast. 2022;66:15–23.
Pfeiffer RM, Webb-Vargas Y, Wheeler W, Gail MH. Proportion of US trends in breast cancer incidence attributable to long-term changes in risk factor distributions. Cancer Epidemiol Biomark Prev. 2018;27(10):1214–22.
Abolghasemi M, Tehrani SS, Yousefi T, Karimian A, Mahmoodpoor A, Ghamari A, et al. Critical roles of long noncoding RNAs in breast cancer. J Cell Physiol. 2020;235(6):5059–71.
Marchese FP, Raimondi I, Huarte M. The multidimensional mechanisms of long noncoding RNA function. Genome Biol. 2017;18(1):206.
Abdulhussain MM, Hasan NA, Hussain AG. Interrelation of the circulating and tissue microRNA-21 with tissue PDCD4 expression and the invasiveness of Iraqi female breast tumors. Indian J Clin Biochem. 2019;34(1):26–38.
Huang Z, Zhen S, Jin L, Chen J, Han Y, Lei W, et al. miRNA-1260b promotes breast cancer cell migration and invasion by downregulating CCDC134. Curr Gene Ther. 2023;23(1):60–71.
Xing Z, Zhang Y, Liang K, Yan L, Xiang Y, Li C, et al. Expression of long noncoding RNA YIYA promotes glycolysis in breast cancer. Can Res. 2018;78(16):4524–32.
Zeng H, Hou Y, Zhou X, Lang L, Luo H, Sun Y, et al. Cancer-associated fibroblasts facilitate premetastatic niche formation through lncRNA SNHG5-mediated angiogenesis and vascular permeability in breast cancer. Theranostics. 2022;12(17):7351–70.
Du WW, Yang W, Li X, Fang L, Wu N, Li F, et al. The circular RNA circSKA3 binds integrin β1 to induce invadopodium formation enhancing breast cancer invasion. Mol Ther. 2020;28(5):1287–98.
Qi L, Sun B, Yang B, Lu S. circRNA RPPH1 facilitates the aggravation of breast cancer development by regulating miR-542-3p/ARHGAP1 pathway. Cancer Biother Radiopharm. 2022;37(8):708–19.
Schneider K, Bertolotti A. Surviving protein quality control catastrophes—from cells to organisms. J Cell Sci. 2015;128(21):3861–9.
Karamali N, Ebrahimnezhad S, Khaleghi Moghadam R, Daneshfar N, Rezaiemanesh A. HRD1 in human malignant neoplasms: molecular mechanisms and novel therapeutic strategy for cancer. Life Sci. 2022;301: 120620.
Patra A, Adhikary A, Ghosh S. The unfolded protein response (UPR) pathway: the unsung hero in breast cancer management. Apoptosis. 2023;28(3):263–76.
Feng Y-X, Jin DX, Sokol ES, Reinhardt F, Miller DH, Gupta PB. Cancer-specific PERK signaling drives invasion and metastasis through CREB3L1. Nat Commun. 2017;8(1):1079.
Syu JP, Chi JT, Kung HN. Nrf2 is the key to chemotherapy resistance in MCF7 breast cancer cells under hypoxia. Oncotarget. 2016;7(12):14659–72.
Cullinan SB, Zhang D, Hannink M, Arvisais E, Kaufman RJ, Diehl JA. Nrf2 is a direct PERK substrate and effector of PERK-dependent cell survival. Mol Cell Biol. 2003;23(20):7198–209.
Notte A, Rebucci M, Fransolet M, Roegiers E, Genin M, Tellier C, et al. Taxol-induced unfolded protein response activation in breast cancer cells exposed to hypoxia: ATF4 activation regulates autophagy and inhibits apoptosis. Int J Biochem Cell Biol. 2015;62:1–14.
Wang S, Chen XA, Hu J, Jiang JK, Li Y, Chan-Salis KY, et al. ATF4 gene network mediates cellular response to the anticancer PAD inhibitor YW3-56 in triple-negative breast cancer cells. Mol Cancer Ther. 2015;14(4):877–88.
Lee AS. Glucose-regulated proteins in cancer: molecular mechanisms and therapeutic potential. Nat Rev Cancer. 2014;14(4):263–76.
Bertolotti A, Zhang Y, Hendershot LM, Harding HP, Ron D. Dynamic interaction of BiP and ER stress transducers in the unfolded-protein response. Nat Cell Biol. 2000;2(6):326–32.
Huber AL, Lebeau J, Guillaumot P, Pétrilli V, Malek M, Chilloux J, et al. p58(IPK)-mediated attenuation of the proapoptotic PERK-CHOP pathway allows malignant progression upon low glucose. Mol Cell. 2013;49(6):1049–59.
Bobrovnikova-Marjon E, Grigoriadou C, Pytel D, Zhang F, Ye J, Koumenis C, et al. PERK promotes cancer cell proliferation and tumor growth by limiting oxidative DNA damage. Oncogene. 2010;29(27):3881–95.
McGrath EP, Logue SE, Mnich K, Deegan S, Jäger R, Gorman AM, et al. The unfolded protein response in breast cancer. Cancers. 2018;10(10):344.
Radford SE. Protein folding: progress made and promises ahead. Trends Biochem Sci. 2000;25(12):611–8.
Hong J, Zhou W, Wang X. Involvement of miR-455 in the protective effect of H(2)S against chemical hypoxia-induced injury in BEAS-2B cells. Pathol Res Pract. 2018;214(11):1804–10.
Du B, Wang X, Wu D, Wang T, Yang X, Wang J, et al. MicroRNA expression profiles identify biomarkers for predicting the response to chemoradiotherapy in rectal cancer. Mol Med Rep. 2018;18(2):1909–16.
Bao Q, Chen L, Li J, Zhao M, Wu S, Wu W, et al. Role of microRNA-124 in cardiomyocyte hypertrophy induced by angiotensin II. Cell Mol Biol. 2017;63(4):23–7.
Belmont PJ, Chen WJ, Thuerauf DJ, Glembotski CC. Regulation of microRNA expression in the heart by the ATF6 branch of the ER stress response. J Mol Cell Cardiol. 2012;52(5):1176–82.
Xu X, Li H. Integrated microRNA-gene analysis of coronary artery disease based on miRNA and gene expression profiles. Mol Med Rep. 2016;13(4):3063–73.
Hisaoka M, Matsuyama A, Nakamoto M. Aberrant calreticulin expression is involved in the dedifferentiation of dedifferentiated liposarcoma. Am J Pathol. 2012;180(5):2076–83.
Pobre KFR, Poet GJ, Hendershot LM. The endoplasmic reticulum (ER) chaperone BiP is a master regulator of ER functions: getting by with a little help from ERdj friends. J Biol Chem. 2019;294(6):2098–108.
Zhang LH, Yang XL, Zhang X, Cheng JX, Zhang W. Association of elevated GRP78 expression with increased astrocytoma malignancy via Akt and ERK pathways. Brain Res. 2011;1371:23–31.
Ouyang YB, Lu Y, Yue S, Xu LJ, Xiong XX, White RE, et al. miR-181 regulates GRP78 and influences outcome from cerebral ischemia in vitro and in vivo. Neurobiol Dis. 2012;45(1):555–63.
Su SF, Chang YW, Andreu-Vieyra C, Fang JY, Yang Z, Han B, et al. miR-30d, miR-181a and miR-199a-5p cooperatively suppress the endoplasmic reticulum chaperone and signaling regulator GRP78 in cancer. Oncogene. 2013;32(39):4694–701.
Liu M, Sun X, Zhu L, Zhu M, Deng K, Nie X, et al. Long noncoding RNA RP11-115N4.1 promotes inflammatory responses by interacting with HNRNPH3 and enhancing the transcription of HSP70 in unexplained recurrent spontaneous abortion. Front Immunol. 2021;12: 717785.
Yuan X, Shen Q, Ma W. Long noncoding RNA hotair promotes the progression and immune escape in laryngeal squamous cell carcinoma through microRNA-30a/GRP78/PD-L1 axis. J Immunol Res. 2022;2022:5141426.
Li G, Luna C, Qiu J, Epstein DL, Gonzalez P. Role of miR-204 in the regulation of apoptosis, endoplasmic reticulum stress response, and inflammation in human trabecular meshwork cells. Invest Ophthalmol Vis Sci. 2011;52(6):2999–3007.
Zhang S, Wang B, Xiao H, Dong J, Li Y, Zhu C, et al. LncRNA HOTAIR enhances breast cancer radioresistance through facilitating HSPA1A expression via sequestering miR-449b-5p. Thorac Cancer. 2020;11(7):1801–16.
Chen M, Ma G, Yue Y, Wei Y, Li Q, Tong Z, et al. Downregulation of the miR-30 family microRNAs contributes to endoplasmic reticulum stress in cardiac muscle and vascular smooth muscle cells. Int J Cardiol. 2014;173(1):65–73.
Hassani SS, Karamali N, Rajabinejad M, Ashjari D, Afshar Hezarkhani L, Gorgin Karaji A, et al. Dysregulation of long noncoding RNA NEAT1/miR-199a-5/BiP axis in patients with diabetic neuropathy. Lab Med. 2023;54(2):160–5.
Ellgaard L, Ruddock LW. The human protein disulphide isomerase family: substrate interactions and functional properties. EMBO Rep. 2005;6(1):28–32.
Yoo JK, Choi SJ, Kim JK. Expression profiles of subtracted mRNAs during cellular senescence in human mesenchymal stem cells derived from bone marrow. Exp Gerontol. 2013;48(5):464–71.
Huang Y, Yan H, Yang Y, Zhou J, Xu Q, Meng H. Downregulated miR-181a alleviates H(2)O(2)-induced oxidative stress and cellular senescence by targeting PDIA6 in human foreskin fibroblasts. An Bras Dermatol. 2023;98(1):17–25.
Groenendyk J, Peng Z, Dudek E, Fan X, Mizianty MJ, Dufey E, et al. Interplay between the oxidoreductase PDIA6 and microRNA-322 controls the response to disrupted endoplasmic reticulum calcium homeostasis. Sci Signal. 2014;7(329):ra54.
Byrd AE, Aragon IV, Brewer JW. MicroRNA-30c-2* limits expression of proadaptive factor XBP1 in the unfolded protein response. J Cell Biol. 2012;196(6):689–98.
Upton J-P, Wang L, Han D, Wang ES, Huskey NE, Lim L, et al. IRE1α cleaves select microRNAs during ER stress to derepress translation of proapoptotic Caspase-2. Science. 2012;338(6108):818–22.
Yuan T, Zhang L, Yao S, Deng SY, Liu JQ. miR-195 promotes LPS-mediated intestinal epithelial cell apoptosis via targeting SIRT1/eIF2a. Int J Mol Med. 2020;45(2):510–8.
Li M, Zhang S, Qiu Y, He Y, Chen B, Mao R, et al. Upregulation of miR-665 promotes apoptosis and colitis in inflammatory bowel disease by repressing the endoplasmic reticulum stress components XBP1 and ORMDL3. Cell Death Dis. 2017;8(3): e2699-e.
Zhu Y, Sun Y, Zhou Y, Zhang Y, Zhang T, Li Y, et al. MicroRNA-24 promotes pancreatic beta cells toward dedifferentiation to avoid endoplasmic reticulum stress-induced apoptosis. J Mol Cell Biol. 2019;11(9):747–60.
Nolan K, Walter F, Tuffy LP, Poeschel S, Gallagher R, Haunsberger S, et al. Endoplasmic reticulum stress-mediated upregulation of miR-29a enhances sensitivity to neuronal apoptosis. Eur J Neurosci. 2016;43(5):640–52.
Dai R, Li J, Liu Y, Yan D, Chen S, Duan C, et al. miR-221/222 suppression protects against endoplasmic reticulum stress-induced apoptosis via p27(Kip1)- and MEK/ERK-mediated cell cycle regulation. Biol Chem. 2010;391(7):791–801.
Guo J, Yang Z, Yang X, Li T, Liu M, Tang H. miR-346 functions as a pro-survival factor under ER stress by activating mitophagy. Cancer Lett. 2018;413:69–81.
Wu H, Li J, Luo S, Wang G. MiR-410 acts as a tumor suppressor in estrogen receptor-positive breast cancer cells by directly targeting ERLIN2 via the ERS pathway. Cell Physiol Biochem. 2018;48(2):461–74.
Zhao T, Du J, Zeng H. Interplay between endoplasmic reticulum stress and non-coding RNAs in cancer. J Hematol Oncol. 2020;13:1–20.
Jiang L, Zang D, Yi S, Li X, Yang C, Dong X, et al. A microRNA-mediated decrease in eukaryotic initiation factor 2α promotes cell survival during PS-341 treatment. Sci Rep. 2016;6(1):21565.
Fribley A, Zeng Q, Wang C-Y. Proteasome inhibitor PS-341 induces apoptosis through induction of endoplasmic reticulum stress-reactive oxygen species in head and neck squamous cell carcinoma cells. Mol Cell Biol. 2004;24(22):9695–704.
Kim T, Croce CM. MicroRNA and ER stress in cancer. Semin Cancer Biol. 2021;75:3–14.
Beylerli O, Gareev I, Sufianov A, Ilyasova T, Guang Y. Long noncoding RNAs as promising biomarkers in cancer. Non-coding RNA Res. 2022;7(2):66–70.
Urra H, Dufey E, Lisbona F, Rojas-Rivera D, Hetz C. When ER stress reaches a dead end. Biochem Biophys Acta. 2013;1833(12):3507–17.
Oakes SA. Endoplasmic reticulum stress signaling in cancer cells. Am J Pathol. 2020;190(5):934–46.
Tang Q, Yuan Q, Li H, Wang W, Xie G, Zhu K, et al. miR-223/Hsp70/JNK/JUN/miR-223 feedback loop modulates the chemoresistance of osteosarcoma to cisplatin. Biochem Biophys Res Commun. 2018;497(3):827–34.
Chen RP, Huang ZL, Liu LX, Xiang MQ, Li GP, Feng JL, et al. Involvement of endoplasmic reticulum stress and p53 in lncRNA MEG3-induced human hepatoma HepG2 cell apoptosis. Oncol Rep. 2016;36(3):1649–57.
Huang ZL, Chen RP, Zhou XT, Zhan HL, Hu MM, Liu B, et al. Long non-coding RNA MEG3 induces cell apoptosis in esophageal cancer through endoplasmic reticulum stress. Oncol Rep. 2017;37(5):3093–9.
Yao X, Tu Y, Xu Y, Guo Y, Yao F, Zhang X. Endoplasmic reticulum stress confers 5-fluorouracil resistance in breast cancer cell via the GRP78/OCT4/lncRNA MIAT/AKT pathway. Am J Cancer Res. 2020;10(3):838–55.
Orak G, Rezaei HB, Ameli F, Maghsoodi F, Cheraghzade M, Adelipour M. The expression of lncRNAs CASC2, NEAT1, LINC00299 in breast cancer tissues and their relationship with the XBP1 splicing rate in Iranian patients during 2014–2019: a cross-sectional study. Health Sci Rep. 2023;6(9): e1552.
Yao X, Tu Y, Xu Y, Guo Y, Yao F, Zhang X. Endoplasmic reticulum stress-induced exosomal miR-27a-3p promotes immune escape in breast cancer via regulating PD-L1 expression in macrophages. J Cell Mol Med. 2020;24(17):9560–73.
Zhang K, Liu H, Song Z, Jiang Y, Kim H, Samavati L, et al. The UPR transducer IRE1 promotes breast cancer malignancy by degrading tumor suppressor microRNAs. iScience. 2020;23(9): 101503.
Liang H, Xiao J, Zhou Z, Wu J, Ge F, Li Z, et al. Hypoxia induces miR-153 through the IRE1α-XBP1 pathway to fine tune the HIF1α/VEGFA axis in breast cancer angiogenesis. Oncogene. 2018;37(15):1961–75.
Arabkari V, Sultana A, Barua D, Webber M, Smith T, Gupta A, et al. UPR-induced miR-616 inhibits human breast cancer cell growth and migration by targeting c-MYC. Int J Mol Sci. 2023;24(17):13034.
Tomar D, Yadav AS, Kumar D, Bhadauriya G, Kundu GC. Non-coding RNAs as potential therapeutic targets in breast cancer. Biochim Biophys Acta. 2020;1863(4): 194378.
Nitheesh Y, Pradhan R, Hejmady S, Taliyan R, Singhvi G, Alexander A, et al. Surface engineered nanocarriers for the management of breast cancer. Mater Sci Eng C Mater Biol Appl. 2021;130: 112441.
Nastiuk KL, Krolewski JJ. Opportunities and challenges in combination gene cancer therapy. Adv Drug Deliv Rev. 2016;98:35–40.
McCormick F. Cancer gene therapy: fringe or cutting edge? Nat Rev Cancer. 2001;1(2):130–41.
Yardim-Akaydin S, Karahalil B, Baytas SN. New therapy strategies in the management of breast cancer. Drug Discov Today. 2022;27(6):1755–62.
Shi Y, Lu J, Zhou J, Tan X, He Y, Ding J, et al. Long non-coding RNA Loc554202 regulates proliferation and migration in breast cancer cells. Biochem Biophys Res Commun. 2014;446(2):448–53.
Yan L, Zheng M, Wang H. Circular RNA hsa_circ_0072309 inhibits proliferation and invasion of breast cancer cells via targeting miR-492. Cancer Manag Res. 2019;11:1033–41.
López-Jiménez E, Andrés-León E. The implications of ncRNAs in the development of human diseases. Non-coding RNA. 2021;7(1):17.
Bahramy A, Zafari N, Rajabi F, Aghakhani A, Jayedi A, Khaboushan AS, et al. Prognostic and diagnostic values of non-coding RNAs as biomarkers for breast cancer: an umbrella review and pan-cancer analysis. Front Mol Biosci. 2023;10:1096524.
Peng L, Jiang J, Tang B, Nice EC, Zhang YY, Xie N. Managing therapeutic resistance in breast cancer: from the lncRNAs perspective. Theranostics. 2020;10(23):10360–77.
Baker EK, El-Osta A. The rise of DNA methylation and the importance of chromatin on multidrug resistance in cancer. Exp Cell Res. 2003;290(2):177–94.
Sharom FJ. ABC multidrug transporters: structure, function and role in chemoresistance. Pharmacogenomics. 2008;9(1):105–27.
Davis S, Lollo B, Freier S, Esau C. Improved targeting of miRNA with antisense oligonucleotides. Nucleic Acids Res. 2006;34(8):2294–304.
Liang Z, Wu H, Xia J, Li Y, Zhang Y, Huang K, et al. Involvement of miR-326 in chemotherapy resistance of breast cancer through modulating expression of multidrug resistance-associated protein 1. Biochem Pharmacol. 2010;79(6):817–24.
Bao L, Hazari S, Mehra S, Kaushal D, Moroz K, Dash S. Increased expression of P-glycoprotein and doxorubicin chemoresistance of metastatic breast cancer is regulated by miR-298. Am J Pathol. 2012;180(6):2490–503.
Wang ZX, Lu BB, Wang H, Cheng ZX, Yin YM. MicroRNA-21 modulates chemosensitivity of breast cancer cells to doxorubicin by targeting PTEN. Arch Med Res. 2011;42(4):281–90.
Miller TE, Ghoshal K, Ramaswamy B, Roy S, Datta J, Shapiro CL, et al. MicroRNA-221/222 confers tamoxifen resistance in breast cancer by targeting p27Kip1. J Biol Chem. 2008;283(44):29897–903.
Zhou M, Liu Z, Zhao Y, Ding Y, Liu H, Xi Y, et al. MicroRNA-125b confers the resistance of breast cancer cells to paclitaxel through suppression of pro-apoptotic Bcl-2 antagonist killer 1 (Bak1) expression. J Biol Chem. 2010;285(28):21496–507.
Bai WD, Ye XM, Zhang MY, Zhu HY, Xi WJ, Huang X, et al. MiR-200c suppresses TGF-β signaling and counteracts trastuzumab resistance and metastasis by targeting ZNF217 and ZEB1 in breast cancer. Int J Cancer. 2014;135(6):1356–68.
He X, Xiao X, Dong L, Wan N, Zhou Z, Deng H, et al. MiR-218 regulates cisplatin chemosensitivity in breast cancer by targeting BRCA1. Tumour Biol. 2015;36(3):2065–75.
Jiang M, Huang O, Xie Z, Wu S, Zhang X, Shen A, et al. A novel long non-coding RNA-ARA: adriamycin resistance-associated. Biochem Pharmacol. 2014;87(2):254–83.
Basak P, Chatterjee S, Bhat V, Su A, Jin H, Lee-Wing V, et al. Long non-coding RNA H19 acts as an estrogen receptor modulator that is required for endocrine therapy resistance in ER+ breast cancer cells. Cell Physiol Biochem. 2018;51(4):1518–32.
Duan Y, Bai X, Yang J, Zhou Y, Gu W, Liu G, et al. Exposure to high-altitude environment is associated with drug transporters change: microRNA-873-5p-mediated alteration of function and expression levels of drug transporters under hypoxia. Drug Metab Dispos Biol Fate Chem. 2022;50(2):174–86.
Assidicky R, Tokat UM, Tarman IO, Saatci O, Ersan PG, Raza U, et al. Targeting HIF1-alpha/miR-326/ITGA5 axis potentiates chemotherapy response in triple-negative breast cancer. Breast Cancer Res Treat. 2022;193(2):331–48.
Zhang D, Shi Z, Li M, Mi J. Hypoxia-induced miR-424 decreases tumor sensitivity to chemotherapy by inhibiting apoptosis. Cell Death Dis. 2014;5(6): e1301-e.
Farazi TA, Horlings HM, Ten Hoeve JJ, Mihailovic A, Halfwerk H, Morozov P, et al. MicroRNA sequence and expression analysis in breast tumors by deep sequencing. Can Res. 2011;71(13):4443–53.
Liu QL, Zhang Z, Wei X, Zhou ZG. Noncoding RNAs in tumor metastasis: molecular and clinical perspectives. Cell Mol Life Sci. 2021;78(21–22):6823–50.
Smith AL, Iwanaga R, Drasin DJ, Micalizzi DS, Vartuli RL, Tan AC, et al. The miR-106b-25 cluster targets Smad7, activates TGF-β signaling, and induces EMT and tumor initiating cell characteristics downstream of Six1 in human breast cancer. Oncogene. 2012;31(50):5162–71.
Chen WX, Hu Q, Qiu MT, Zhong SL, Xu JJ, Tang JH, et al. miR-221/222: promising biomarkers for breast cancer. Tumour Biol. 2013;34(3):1361–70.
Taylor MA, Sossey-Alaoui K, Thompson CL, Danielpour D, Schiemann WP. TGF-β upregulates miR-181a expression to promote breast cancer metastasis. J Clin Investig. 2013;123(1):150–63.
Frères P, Josse C, Bovy N, Boukerroucha M, Struman I, Bours V, et al. Neoadjuvant chemotherapy in breast cancer patients induces miR-34a and miR-122 expression. J Cell Physiol. 2015;230(2):473–81.
Bahn JH, Zhang Q, Li F, Chan TM, Lin X, Kim Y, et al. The landscape of microRNA, Piwi-interacting RNA, and circular RNA in human saliva. Clin Chem. 2015;61(1):221–30.
Chen X, Gao C, Li H, Huang L, Sun Q, Dong Y, et al. Identification and characterization of microRNAs in raw milk during different periods of lactation, commercial fluid, and powdered milk products. Cell Res. 2010;20(10):1128–37.
Zeng B, Li Y, Feng Y, Lu M, Yuan H, Yi Z, et al. Downregulated miR-1247-5p associates with poor prognosis and facilitates tumor cell growth via DVL1/Wnt/β-catenin signaling in breast cancer. Biochem Biophys Res Commun. 2018;505(1):302–8.
Fan CN, Ma L, Liu N. Comprehensive analysis of novel three-long noncoding RNA signatures as a diagnostic and prognostic biomarkers of human triple-negative breast cancer. J Cell Biochem. 2019;120(3):3185–96.
Ito M, Miyata Y, Okada M. Current clinical trials with non-coding RNA-based therapeutics in malignant diseases: a systematic review. Transl Oncol. 2023;31: 101634.
Shah PP, Dupre TV, Siskind LJ, Beverly LJ. Common cytotoxic chemotherapeutics induce epithelial–mesenchymal transition (EMT) downstream of ER stress. Oncotarget. 2017;8(14):22625–39.
Clarke HJ, Chambers JE, Liniker E, Marciniak SJ. Endoplasmic reticulum stress in malignancy. Cancer Cell. 2014;25(5):563–73.
Rankin EB, Giaccia AJ. Hypoxic control of metastasis. Science. 2016;352(6282):175–80.
Powis G, Kirkpatrick L. Hypoxia inducible factor-1alpha as a cancer drug target. Mol Cancer Ther. 2004;3(5):647–54.
Noman MZ, Desantis G, Janji B, Hasmim M, Karray S, Dessen P, et al. PD-L1 is a novel direct target of HIF-1α, and its blockade under hypoxia enhanced MDSC-mediated T cell activation. J Exp Med. 2014;211(5):781–90.
Zhang Y, Yan J, Wang L, Dai H, Li N, Hu W, et al. HIF-1α promotes breast cancer cell MCF-7 proliferation and invasion through regulating miR-210. Cancer Biother Radiopharm. 2017;32(8):297–301.
Galluzzi L, Baehrecke EH, Ballabio A, Boya P, Bravo-San Pedro JM, Cecconi F, et al. Molecular definitions of autophagy and related processes. EMBO J. 2017;36(13):1811–36.
Park SM, Ou J, Chamberlain L, Simone TM, Yang H, Virbasius CM, et al. U2AF35(S34F) promotes transformation by directing aberrant ATG7 pre-mRNA 3′ end formation. Mol Cell. 2016;62(4):479–90.
Lin Y, Wang Z, Liu L, Chen L. Akt is the downstream target of GRP78 in mediating cisplatin resistance in ER stress-tolerant human lung cancer cells. Lung Cancer. 2011;71(3):291–7.
Aguirre-Ghiso JA. Models, mechanisms and clinical evidence for cancer dormancy. Nat Rev Cancer. 2007;7(11):834–46.
Brewer JW, Diehl JA. PERK mediates cell-cycle exit during the mammalian unfolded protein response. Proc Natl Acad Sci USA. 2000;97(23):12625–30.
Terai H, Kitajima S, Potter DS, Matsui Y, Quiceno LG, Chen T, et al. ER stress signaling promotes the survival of cancer “Persister cells” tolerant to EGFR tyrosine kinase inhibitors. Can Res. 2018;78(4):1044–57.
Ojha R, Amaravadi RK. Targeting the unfolded protein response in cancer. Pharmacol Res. 2017;120:258–66.
Carrasco DR, Sukhdeo K, Protopopova M, Sinha R, Enos M, Carrasco DE, et al. The differentiation and stress response factor XBP-1 drives multiple myeloma pathogenesis. Cancer Cell. 2007;11(4):349–60.
Liang X, Liu J, Liu X, Jin Y, Xu M, Han Z, et al. LINP1 represses unfolded protein response by directly inhibiting eIF2α phosphorylation to promote cutaneous squamous cell carcinoma. Exp Hematol Oncol. 2023;12(1):31.
Tang G-H, Chen X, Ding J-C, Du J, Lin X-T, Xia L, et al. LncRNA LUCRC regulates colorectal cancer cell growth and tumorigenesis by targeting endoplasmic reticulum stress response. Front Genet. 2020;10:1409.
Jiang L, Wang C, Shen X. LncRNA GAS5 suppresses ER stress-induced apoptosis and inflammation by regulating SERCA2b in HG-treated retinal epithelial cell. Mol Med Rep. 2020;22(2):1072–80.
Wang S, Duan J, Liao J, Wang Y, Xiao X, Li L, et al. LncRNA H19 inhibits ER stress induced apoptosis and improves diabetic cardiomyopathy by regulating PI3K/AKT/mTOR axis. Aging. 2022;14(16):6809–28.
Maurel M, Dejeans N, Taouji S, Chevet E, Grosset CF. MicroRNA-1291-mediated silencing of IRE1α enhances glypican-3 expression. RNA. 2013;19(6):778–88.
Gupta A, Hossain MM, Read DE, Hetz C, Samali A, Gupta S. PERK regulated miR-424(322)-503 cluster fine-tunes activation of IRE1 and ATF6 during unfolded protein response. Sci Rep. 2015;5(1):18304.
Yu Y, Zhou H, Xiong Y, Liu J. Exosomal miR-199a-5p derived from endothelial cells attenuates apoptosis and inflammation in neural cells by inhibiting endoplasmic reticulum stress. Brain Res. 2020;1726: 146515.
Lin S, Que Y, Que C, Li F, Deng M, Xu D. Exosome miR-3184-5p inhibits gastric cancer growth by targeting XBP1 to regulate the AKT, STAT3, and IRE1 signalling pathways. Asia Pac J Clin Oncol. 2023;19(2):e27–38.
Rezghi Barez S, Movahedian Attar A, Aghaei M. MicroRNA-30c-2-3p regulates ER stress and induces apoptosis in ovarian cancer cells underlying ER stress. EXCLI J. 2021;20:922–34.
Tu L, Long X, Song W, Lv Z, Zeng H, Wang T, et al. MiR-34c acts as a tumor suppressor in non-small cell lung cancer by inducing endoplasmic reticulum stress through targeting HMGB1. Onco Targets Ther. 2019;12:5729–39.
Chatterjee N, Fraile-Bethencourt E, Baris A, Espinosa-Diez C, Anand S. MicroRNA-494 regulates endoplasmic reticulum stress in endothelial cells. Front Cell Dev Biol. 2021;9: 671461.
Xu G, Chen J, Jing G, Grayson TB, Shalev A. miR-204 targets PERK and regulates UPR signaling and β-cell apoptosis. Mol Endocrinol. 2016;30(8):917–24.
He L, Yuan J, Xu Q, Chen R, Chen L, Fang M. miRNA-1283 regulates the PERK/ATF4 pathway in vascular injury by targeting ATF4. PLoS ONE. 2016;11(8): e0159171.
Kong F, Zou H, Liu X, He J, Zheng Y, Xiong L, et al. miR-7112-3p targets PERK to regulate the endoplasmic reticulum stress pathway and apoptosis induced by photodynamic therapy in colorectal cancer CX-1 cells. Photodiagn Photodyn Ther. 2020;29: 101663.
Chitnis NS, Pytel D, Bobrovnikova-Marjon E, Pant D, Zheng H, Maas NL, et al. miR-211 is a prosurvival microRNA that regulates chop expression in a PERK-dependent manner. Mol Cell. 2012;48(3):353–64.
Miyamoto Y, Mauer AS, Kumar S, Mott JL, Malhi H. Mmu-miR-615-3p regulates lipoapoptosis by inhibiting C/EBP homologous protein. PLoS ONE. 2014;9(10): e109637.
Arabkari V, Barua D, Hossain MM, Webber M, Smith T, Gupta A, et al. miRNA-378 is downregulated by XBP1 and inhibits growth and migration of luminal breast cancer cells. Int J Mol Sci. 2024;25(1):186.
Ahmadi A, Khansarinejad B, Hosseinkhani S, Ghanei M, Mowla SJ. miR-199a-5p and miR-495 target GRP78 within UPR pathway of lung cancer. Gene. 2017;620:15–22.
Quan Y, Song Q, Wang J, Zhao L, Lv J, Gong S. MiR-1202 functions as a tumor suppressor in glioma cells by targeting Rab1A. Tumor Biol. 2017;39(4):1010428317697565.
Zhang Z, Wu S, Muhammad S, Ren Q, Sun C. miR-103/107 promote ER stress-mediated apoptosis via targeting the Wnt3a/β-catenin/ATF6 pathway in preadipocytes. J Lipid Res. 2018;59(5):843–53.
Chen Z, Liu Y, Yang L, Liu P, Zhang Y, Wang X. MiR-149 attenuates endoplasmic reticulum stress-induced inflammation and apoptosis in nonalcoholic fatty liver disease by negatively targeting ATF6 pathway. Immunol Lett. 2020;222:40–8.
Kim JO, Kwon EJ, Song DW, Lee JS, Kim DH. miR-185 inhibits endoplasmic reticulum stress-induced apoptosis by targeting Na+/H+ exchanger-1 in the heart. BMB Rep. 2016;49(4):208–13.
Wang X, Wang P, Zhu Y, Zhang Z, Zhang J, Wang H. MicroRNA-520a attenuates proliferation of Raji cells through inhibition of AKT1/NF-κB and PERK/eIF2α signaling pathway. Oncol Rep. 2016;36(3):1702–8. Retraction in https://doi.org/10.3892/or.2024.8706.
Acknowledgements
Not applicable.
Funding
No funding was received.
Author information
Authors and Affiliations
Contributions
N. K., M. J., N. Gh., R. M., Z. A., A. R., A. D., A. M. and Z. M.: conceptualization, writing—original draft, writing—review and editing A. R., A. D. and S. S.: preparing figures A. R., A. D. B. B.: supervision, review, edit, and finalize the manuscript. All authors read and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Karamali, N., Daraei, A., Rostamlou, A. et al. Decoding contextual crosstalk: revealing distinct interactions between non-coding RNAs and unfolded protein response in breast cancer. Cancer Cell Int 24, 104 (2024). https://doi.org/10.1186/s12935-024-03296-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12935-024-03296-3