
Abstract
Background
Metabolic Syndrome (MetS) is characterized by risk factors such as abdominal obesity, hypertriglyceridemia, low high-density lipoprotein cholesterol (HDL-C), hypertension, and hyperglycemia, which contribute to the development of cardiovascular disease and type 2 diabetes. Here, we aim to identify candidate metabolite biomarkers of MetS and its associated risk factors to better understand the complex interplay of underlying signaling pathways.
Methods
We quantified serum samples of the KORA F4 study participants (N = 2815) and analyzed 121 metabolites. Multiple regression models adjusted for clinical and lifestyle covariates were used to identify metabolites that were Bonferroni significantly associated with MetS. These findings were replicated in the SHIP-TREND-0 study (N = 988) and further analyzed for the association of replicated metabolites with the five components of MetS. Database-driven networks of the identified metabolites and their interacting enzymes were also constructed.
Results
We identified and replicated 56 MetS-specific metabolites: 13 were positively associated (e.g., Val, Leu/Ile, Phe, and Tyr), and 43 were negatively associated (e.g., Gly, Ser, and 40 lipids). Moreover, the majority (89%) and minority (23%) of MetS-specific metabolites were associated with low HDL-C and hypertension, respectively. One lipid, lysoPC a C18:2, was negatively associated with MetS and all of its five components, indicating that individuals with MetS and each of the risk factors had lower concentrations of lysoPC a C18:2 compared to corresponding controls. Our metabolic networks elucidated these observations by revealing impaired catabolism of branched-chain and aromatic amino acids, as well as accelerated Gly catabolism.
Conclusion
Our identified candidate metabolite biomarkers are associated with the pathophysiology of MetS and its risk factors. They could facilitate the development of therapeutic strategies to prevent type 2 diabetes and cardiovascular disease. For instance, elevated levels of lysoPC a C18:2 may protect MetS and its five risk components. More in-depth studies are necessary to determine the mechanism of key metabolites in the MetS pathophysiology.
Similar content being viewed by others
Background
Metabolic Syndrome (MetS) is diagnosed by the presence of at least three out of five risk factors: abdominal obesity, hypertriglyceridemia, reduced high-density lipoprotein cholesterol (HDL-C), hypertension, and hyperglycemia [1]. This diagnosis carries a two-fold risk of developing cardiovascular disease (CVD) within 5–10 years and a five-fold greater risk of developing type 2 diabetes (T2D) [1]. Furthermore, individuals with MetS have a higher rate of developing serious complications from severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection [2]. Although global data on MetS is lacking, a study conducted in 2018 estimated that more than one billion people worldwide suffer from MetS [3], making it a significant public health issue due to its association with serious chronic complications such as CVD and T2D.
The etiology of MetS is attributed to multiple interacting factors such as genetic susceptibility, epigenetic factors, and environmental influences, as well as the five metabolic components [4]. Obesity, for instance, is largely influenced by socioeconomic and dietary factors, with excessive caloric intake possibly leading to inflammatory conditions and impaired energy metabolism [5]. Dyslipidemia, characterized by the imbalance of lipids such as raised triglycerides and lower HDL–C, is considered to be a risk factor for the development of atherosclerotic CVD [1]. Excessive food intake and abnormal lipid metabolism have been reported to be associated with the development of atherogenic dyslipidemia [6]. Hypertension, another significant component of MetS, arises from oxidative stress, endothelial dysfunction, and increased inflammatory mediators, posing a major risk for CVDs, including stroke [4]. In addition, hyperglycemia and insulin resistance exacerbate low-grade inflammation, enhancing the risk of CVDs [7]. Therefore, besides lifestyle modifications and psychological management, understanding the pathophysiology of MetS is vital for developing pharmaceutical interventions for personalized medicine [2].
Technological advancements enable the quantification and analysis of multi-omics data such as genomic, proteomic, and metabolomic data in clinical studies and human cohorts [8]. The comprehensive analysis of molecules (genes, proteins, and metabolites) may contribute to a systematic understanding of biological processes and to the development of personalized therapies [9]. Metabolome refers to the complete set of small molecule metabolites, which are the intermediates or end products of metabolism, and reflect the metabolic status of an individual or a population. By applying advanced analytical and statistical methods, metabolomics has the potential to reveal novel biomarkers that can improve the diagnosis, prognosis, and risk assessment of MetS and its related disorders [10, 11].
In particular, the utilization of the targeted metabolomics approach in previous studies provides a robust foundation for our current research, enabling comparisons and the identification of metabolites and potential biomarkers associated with the studied population. For instance, in the population-based KORA (Cooperative Health Research in the Region of Augsburg) study, targeted metabolomics has been intensively used to explore the association between metabolites and age, gender, smoking, alcohol intake, pre-diabetes, and T2D, as well as chronic kidney disease [12,13,14,15,16,17,18]. Similarly, in the SHIP (Health Study of Pomerania)-TREND-0 study, targeted metabolomics has been used to investigate the metabolites associated with the use of oral contraceptives [19].
Several metabolomic studies on MetS have employed various approaches, such as untargeted, targeted metabolomic, and lipidomic, revealing novel biomarkers and providing new insights into metabolic alterations in MetS [20,21,22,23]. However, previous studies have often utilized relatively smaller sample sizes (e.g., 30 individuals with MetS), or only 100 men [20, 22, 23]. To date, there remains a gap in research focusing on large population-based human cohorts. Therefore, to better understand the underlying mechanisms of MetS, it is necessary to conduct further research utilizing metabolomic approaches in larger sample sizes.
The present study, based on a targeted metabolomics approach, aims to identify MetS-associated metabolites in a population-based human cohort, namely the KORA F4 study [24], and replicate the findings in the SHIP-TREND-0 study [25]. The secondary objective is to investigate the association between the replicated MetS-associated metabolites and the five components of MetS, as well as to construct interaction networks among identified metabolites, enzymes, and biochemical processes to better understand the underlying mechanisms of MetS.
Methods
Description of human cohorts: discovery and replication studies
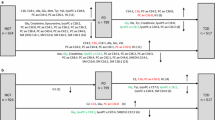
Discovery study The discovery study utilized data from the KORA F4, a follow-up to the KORA baseline survey number 4 (KORA S4) [24, 26]. The KORA F4 study, conducted between 2006 and 2008, examined 3080 individuals (aged 32–81 years). The clinical variables, demographics, and laboratory measurements of KORA F4 have been described in detail in our previous reports [27, 28]. Only participants with metabolite measurements and clinical variables for MetS were included. We excluded: (1) missing data on phenotypes and metabolites (N = 51); (2) the presence of metabolites’ extreme outliers detected by data points outside of the mean ± 5 × standard deviation range (N = 201); (3) non-fasting samples (N = 7); (4) and missing diagnosis of MetS (N = 6). In total, we included 2815 individuals in this discovery study (Fig. 1).

Population description and study design. Abbreviations: SBP, systolic blood pressure; DBP, diastolic blood pressure
Replication study The replication study was based on the SHIP-TREND-0 study, a population-based study in northeastern Germany, which recruited 4,420 participants between 2008 and 2012 [25, 29, 30]. Details of the measurements performed in the SHIP-TREND-0 study were published [19, 25, 31]. The replication study included 988 individuals with data on MetS and metabolite measurements (Fig. 1).
KORA and SHIP studies were approved by the Ethics Committees of the Bavarian Medical Association in Munich, and the Institutional Review Board of the University of Greifswald, Germany, respectively. All study participants provided written informed consent.
The definition of MetS
The definition of MetS in KORA F4 and SHIP-TREND-0 studies was based on a joint scientific statement published in 2009 [1, 9]. MetS is diagnosed by the presence of any three of the following five components: (1) abdominal obesity with waist circumferences that measure ≥ 94 cm for men, or ≥ 80 cm for women; (2) hypertriglycerides with fasting serum triglycerides ≥ 150 mg/dL, or drug treatment for elevated triglycerides (fibrates); (3) low serum HDL–C < 40 mg/dL in men, or < 50 mg/dL in women, or drug treatment for reduced HDL–C (fibrates); (4) hypertension with systolic blood pressure ≥ 130 mmHg, or diastolic blood pressure ≥ 85 mmHg, or antihypertensive medication treatment; (5) hyperglycemia with fasting serum glucose level ≥ 100 mg/dL or antidiabetic drug treatment. According to the MetS diagnostic criteria, the KORA F4 and SHIP-TREND-0 participants were divided into MetS and non-MetS groups (Fig. 1).
Metabolite quantification and normalization
Serum samples from the KORA F4 study were measured with the AbsoluteIDQ™ p150 kit (BIOCRATES Life Sciences AG, Innsbruck, Austria) for the quantification of 163 metabolites [12]. Specifically, samples were randomly distributed on 38 kit plates, each plate also including three quality control (QC) samples provided by the manufacturer and one zero sample (PBS) in addition to the individual samples [18, 32]. Among the quantified metabolites, only those that met all of the following three criteria were used: (1) missing values < 10%; (2) median relative standard deviations (RSD, also called coefficient of variation (CV)) of three QC samples < 25%; (3) 50% of measured sample values equal or above the limits of detection (LOD). In total, 121 metabolites that met QC included 14 amino acids, 1 monosaccharide, 18 acylcarnitines, 67 phosphatidylcholines (PCs), 9 lysoPCs (LPCs), and 12 sphingomyelins (Additional file 1: Table S1).
To minimize the technical variations that metabolomics data inevitably contain, metabolite concentrations were adjusted by a non-parametric method TIGER, which is based on an adaptable ensemble learning architecture [32]. In addition, to ensure comparability between different metabolites, their values were natural-log transformed and standardized to have a mean value of 0 and a standard deviation of 1.
In the SHIP-TREND-0 study, metabolic profiling was measured using the AbsoluteIDQ™ p180 kit (BIOCRATES Life Sciences AG, Innsbruck, Austria). Metabolites from the SHIP-TREND-0 that passed quality control were used for replication. A detailed description of the metabolite measurements, as well as pre-processing, has been provided in published papers [19, 33]. The metabolomics measurements in the SHIP-TREND-0 study were performed for a subset of participants without self-reported diabetes. Nonetheless, the study included participants with prediabetes, along with a sufficient number of individuals making the analysis robust and generalizable (Fig. 1).
Statistical analysis
We first used a multiple linear regression model to identify MetS-associated metabolites with the metabolite concentration values as dependent variable and the grouping variable as independent variable. To include potential confounders, we adjusted for age, sex and smoking status, physical activity, and alcohol intake. In the KORA study, we used the multiple linear regression model: Y metabolite concentration ~ X grouping (non-MetS = 0, prevalent MetS = 1) + age (years) + sex (female = 0, male = 1) + smoking status (non-smoker = 0, former smoker = 1, and current smoker = 2) + physical activity (inactive = 0, active =1; regular exercise per week ≥ 1 h is considered active, irregular equal or less than 1 h is considered inactive) + alcohol intake (g/day). Each metabolite was analyzed separately. To account for multiple testing for the 121 utilized metabolites, a Bonferroni cutoff was applied and only metabolites with P-value < 0.05/121 = 4.13E-04 were considered to be statistically significantly associated with MetS in the KORA F4 study (Fig. 1).
To replicate MetS-associated metabolites, in the SHIP-TREND-0 study, we used similar multiple linear regression model: Y metabolite concentration ~ X grouping (non-MetS = 0, prevalent MetS = 1) + age (years) + sex (female = 2, male = 1) + smoking status (non-smoker = 0, former smoker = 1, and current smoker = 2) + physical activity (inactive = 0, active =1) + alcohol intake (g/day). Each metabolite was analyzed separately. A Bonferroni cutoff was applied for multiple testing corrections and only metabolites with a P-value < 0.05/88 = 5.68E-04 were considered significant in the SHIP-TREND-0 study.
A multiple logistic regression analysis was further employed to evaluate the association between identified and replicated MetS-associated metabolites and each of the five MetS components in the KORA F4 study. In the multiple logistic regression model, the grouping variable of each component of MetS was defined as outcome and metabolite concentration values as explanatory variables: Y grouping (non-component 1 = 0, prevalent component 1 = 1) ~ X metabolite concentration + age + sex + smoking status + physical activity + alcohol intake + other four components. For each of the components of MetS, each metabolite was assessed individually. For multiple testing correction, a Bonferroni cutoff was applied and only metabolites with P-value < 0.05/56 = 8.93E-04 were considered to be statistically significantly associated with the component of MetS in the KORA F4 study (Fig. 1).
All analyses were carried out using R, version 4.1.2, and STATA/MP 17.
Construction of metabolic networks
CIDeR is a manually curated multifactorial database that integrates interactions between heterogeneous factors associated with human diseases [34]. Data in CIDeR has been sourced from research articles and reviews, the annotations for all interactions were manually annotated by experienced biocurators.
Based on the CIDeR database, in-depth literature research was conducted on the identified metabolites to examine the underlying mechanisms, linked biological processes, clinical phenotypes, and enzymes (protein and encoding gene) that are specifically associated with the five components of MetS.
Results
Characteristics of discovery and replication study participants
Table 1 shows the characteristics of the KORA F4 study participants (N = 2815), which included 959 participants with MetS and 1,856 individuals without MetS (Fig. 1). Significant differences in age and sex were observed between the two groups. Participants with MetS were notably older and included a higher proportion of males. Consistent with the diagnostic criteria, individuals with MetS had higher BMI and waist circumferences. Regarding blood measurements, the levels of triglycerides, HDL–C, blood pressure, fasting glucose, and HbA1c were worse in the MetS group than in the non-MetS group. As can be seen from Table 1, smoking status differed significantly between the two groups, with a higher proportion of former smokers in the MetS group. Furthermore, the non-MetS group had a larger proportion of physically active participants, while alcohol intake did not demonstrate statistical differences between the groups.
The SHIP-TREND-0 study included 988 participants, comprising 211 individuals diagnosed with MetS and 777 non-MetS participants. Among the MetS individuals, just over half (56%) were male. Consistent with the KORA F4 study, a closer inspection of the table shows significant differences between MetS and non-MetS groups in all included variables except for alcohol intake (Table 1, Fig. 1).
Identified and replicated 56 MetS-associated metabolites
Of the 121 utilized metabolites, we identified 89 metabolites that were significantly (P-value < 0.05/121 = 4.13E-04) associated with MetS based on the multiple linear regression models when comparing 959 participants with MetS to 1856 individuals without MetS in the KORA F4 study (Fig. 2a and Table 2). We further performed replications of the 89 MetS-associated metabolites in the SHIP-TREND-0 study. Since metabolite C8:1 was not measured in the SHIP-TREND-0 study due to the different analytical measurement kits, we used the remaining 88 metabolites for replication. Among these, 85 metabolites had the same effect direction in both studies, and 56 remained significant after correction for multiple testing, thus considered successfully replicated (P-value < 0.05/88 = 5.68E-04, Table 2). For example, acyl-alkyl PC with 34 carbons and three double-bonds (PC ae C34:3) demonstrated a negative association with MetS in KORA F4 (ß = − 0.76, P = 4.06E-82) and SHIP-TREND-0 (ß = − 0.73, P = 1.08E-20). Among the successfully replicated metabolites, 13 metabolites (4 amino acids (Val, xLeu (Leu/Ile), Phe and Tyr), 1 monosaccharide (sum of hexoses, H1), 2 acylcarnitines, and 6 PCs) were positively associated with MetS, and 43 metabolites (3 amino acids (Gly, Ser, and Gln), 30 PCs, 2 LPCs, 8 sphingomyelins) were negatively associated with MetS (Table 2).

The metabolites associated with MetS and its five components. a Volcano plot of regression results of 121 metabolites. The horizontal dashed lines represent the Bonferroni cut-off value. Orange dots: positively associated, blue dots: negatively associated, gray dots: not significant. b Circle plot of the association between 56 replicated MetS-associated metabolites and 5 components of MetS. Metabolites with bold font indicate significant association with three or more components
Metabolites associated with the five components
We further investigated whether and how the identified and replicated 56 MetS-associated metabolites were associated with each of the five components of MetS. Using the multiple logistic regression analyses, we found that abdominal obesity, hypertriglyceridemia, reduced HDL–C, hypertension, and hyperglycemia were significantly (P-value < 0.05/56 = 8.93E-04) associated with 36, 37, 50, 13, 35 metabolites, respectively (Figs. 2b, 3a, Additional file 1: Table S2). In addition, lysoPC a C18:2 was the only metabolite significantly negatively associated with all five components: abdominal obesity (odds ratio, OR = 0.57, P = 3.96E-23), hypertriglyceridemia (OR = 0.69, P = 6.57E-10), low HDL–C (OR = 0.65, P = 1.34E-12), hypertension (OR = 0.82, P = 2.62E-04), and hyperglycemia (OR = 0.81, P = 1.24E-04) (Additional file 1: Table S2). The concentration of lysoPC a C18:2 was lower in individuals with MetS and in those with each of the five components compared to respective controls (Fig. 3b). Furthermore, BCAAs (Val, xLeu (Leu/Ile) and aromatic amino acids (Phe and Tyr) were positively associated with at least three of the five components (Fig. 2b). The concentrations of Val, xLeu, Phe and Tyr were higher in participants with abdominal obesity, hypertriglyceridemia, reduced HDL–C (except for Tyr), hypertension (except for BCAAs and Tyr), and hyperglycemia (except for Val and Phe), when compared to corresponding controls, respectively (Additional file 1: Table S2). Moreover, two acylcarnitines (C0 and C3) showed positive associations with abdominal obesity, hypertriglyceridemia, and reduced HDL–C (Fig. 2b).

The number of overlapped metabolites among the five components and comparison of the overlapped metabolite. a The Venn diagram displays the number of metabolites that were Bonferroni significantly associated with each component. b Box plots shows the log-transformed and scaled concentrations of lysoPC a C18:2 in the MetS group, the five components of MetS groups, and the corresponding non-case groups
Three created metabolic networks
We constructed three networks by systematically investigating the identified and replicated MetS-associated metabolites with MetS and its five components based on the CIDeR database. We found that the conversions between PCs and LPCs are mediated by multiple enzymes (e.g., LCAT, lecithin cholesterol acyltransferase; LIPG, endothelial lipase; LPCAT3, lysophosphatidylcholine acyltransferase—3) (Fig. 4 and Additional file 1: Table S3). All of these three enzymes are functionally interacting with at least one of the five components. For example, hypertension regulates LIPG's activity which can hydrolyze the fatty acid from both PCs and LPCs (Fig. 4). Additionally, LIPG’s concentration was reported positively associated with increased abdominal obesity (waist circumference), hypertriglyceridemia, hyperglycemia, and decreased HDL–C (Fig. 4). To further explore underlying molecular mechanisms, we created two networks of amino acids and acylcarnitines (Figs. 5, 6, see "Discussion").

Metabolic network of PCs and LPCs. LCAT lecithin cholesterol acyltransferase, APOA1 apolipoprotein A1, LIPG endothelial lipase, LPCAT3 lysophosphatidylcholine acyltransferase-3, PEMT phosphatidylethanolamine N-methyltransferase, ENPP2 ectonucleotide pyrophosphatase/phosphodiesterase 2, SLC2A4 solute carrier family 2, facilitated glucose transporter member 4

Metabolic network of BCAAs. BCKDK branched chain ketoacid dehydrogenase kinase, BCAT2 Branched Chain Amino Acid Transaminase 2, SLC7A5 Solute Carrier Family 7 Member 5, PPM1K Protein phosphatase 1 K, SLC2A4 solute carrier family 2, facilitated glucose transporter member 4, CRAT carnitine acetyltransferase

Metabolic network of Ser, Gly, Phe, and Tyr. TH tyrosine hydroxylase, PAH phenylalanine hydroxylase, TAT tyrosine aminotransferase, BH4 tetrahydrobiopterin, THF tetrahydrofolate, GLDC glycine decarboxylase, AMT aminomethyltransferase, SHMT serine hydroxymethyltransferase
Discussion
MetS is a complex syndrome derived to capture the interplay of risk factors for cardiometabolic diseases. To better understand the underlying molecular mechanisms, we analyzed more than 3800 study participants in two population-based human cohorts (KORA F4 and SHIP-TREND-0). Of 121 metabolites, 56 were identified and replicated as MetS-associated metabolites and further evaluated with each of the five components representing a criterion for the MetS. The creation of three metabolic networks facilitated the understanding of the interplay between metabolites, proteins, biomedical processes, phenotypes, and MetS components.
Our findings highlight H1 as the metabolite with the most positive association with MetS (Fig. 2a, Table 2), and exclusively associated with the component of hyperglycemia (Fig. 2b, and Additional file 1: Table S2). Based on the targeted metabolomics technology, we were unable to distinguish between different hexoses. Therefore, it is likely that the level of H1 is mainly represented by glucose and thereby directly linked to the definition of hyperglycemia, as expected [16].
We also found that 36 PCs were associated with MetS. Biological membranes are composed primarily of phospholipids, and phospholipids additionally serve as substrates to produce bioactive compounds, and the most abundant phospholipids in mammalian cell membranes are PCs [35]. PCs play an important role in lipoprotein metabolism by regulating the expression of key factors in lipoprotein production [36]. Consistent with our observations, four long-chain acyl-alkyl PCs (PC ae C42:5, PC ae C44:5, PC ae C40:4, and PC ae C44:4) were inversely associated with MetS in a study of the Health Survey of Sao Paulo, which analyzed plasma samples from 130 participants [37]. Another study analyzed morning urine samples and observed a higher concentration of metabolite PC (34:2) in individuals with MetS (N = 30) when compared to controls (N = 20) [22]. However, no significant association between PC aa C34:2 and MetS was found in the serum samples of the KORA F4 study. This discrepancy could be attributed to a multitude of factors, including the limited sample size utilized in the analysis and the potential impact of using urine samples on the specificity of the results.
We observed that two LPCs were negatively associated with MetS, and especially, lysoPC a C18:2 was significantly associated with all five MetS components. There is an increasing awareness of the connection between LPCs and cardiovascular and neurodegenerative disorders [38]. Prior studies have shown that LPC was important for the mechanism connecting saturated fatty acids to insulin resistance [39]. Using the targeted metabolomics approaches, it is shown that lower levels of lysoPC a C18:2 are not only found in adults with impaired glucose tolerance [16], but also in obese children [40], and this lipid is independently associated with decreased risk of T2D [17] as well as myocardial infarction [41]. These findings exhibit consistency with our results. Besides, LIPG has the ability to hydrolyze fatty acids from both PCs and LPCs, and LIPG has a significant association with features of MetS, it may potentially serve as a pro-atherogenic factor in individuals with MetS (Fig. 4) [42]. Furthermore, LIPG has been shown to produce LPC18:2 from PCs [43]. This implies that LIPG has an important role in the pathogenesis of MetS, as it is not only associated with the five MetS components (Fig. 4 and Additional file 1: Table S3), but also with lysoPC a C18:2, which is also associated with all five components in our study. In addition, lysoPC a C18:2 may also be involved in the regulation of lipid metabolism, inflammation, and glucose homeostasis, which are key factors in the development and progression of MetS [16, 17, 41]. However, we are just beginning to understand how PCs and LPCs are metabolically regulated in health and disease. To understand the mechanisms that underlie PC’s and LPC’s potential roles in metabolic diseases, future functional studies will be necessary.
Among the 14 amino acids, we found seven (Val, xLeu, Phe, Tyr, Ser, Gly, and Gln) were significantly associated with MetS. Consistent with previous studies, Ser was reported to be inversely associated with MetS in the Health Survey of Sao Paulo study [37]. Additionally, a previous study of 56 MetS participants found a positive association between MetS and BCAAs and aromatic amino acids, which is in line with our findings [23]. Given that meats and meat products are rich sources of BCAAs and aromatic amino acids, these observations could be a sign of meat consumption [44].
Recently, an untargeted metabolomics approach based on nuclear magnetic resonance technology was used to analyze about 1000 plasma samples of a community-based study in a Chinese population [20]. Of 85 used metabolites, 13 (e.g., d-maltose, d-fucose, and l-ornithine) were selected as candidate biomarkers for MetS and it was concluded that arginine metabolism may play a role in the pathophysiological mechanism of MetS [20]. The observations of d-maltose and d-fucose are consistent with our H1 hexoses observations, however, both arginine (ß = − 0.11, P = 1.26E-02) and ornithine (ß = 0.01, P = 9.04E-01) were not significantly associated with MetS in the KORA F4 study.
We observed a positive association of BCAAs not only with MetS, but also with abdominal obesity, hypertriglyceridemia, and reduced HDL–C. The expression of SLC7A5 (solute carrier family 7 member 5, also known as LAT1, the transporter of BCAAs) was reported to be reduced in obese individuals, in addition, several enzymes associated with BCAA catabolism, such as propionyl-coA carboxylase (PCC), branched-chain ketoacid dehydrogenase (BCKD) complex and branched-chain amino acid transaminase 2 (BCAT2) were reported down-regulated (Fig. 5 and Additional file 1: Table S4). Specifically, mitochondrial branched-chain amino acid aminotransferase (BCATm) and BCKD E1α were found reduced in adipose tissues, but not in skeletal muscles [45, 46]. Down-regulated enzymes observed in obese individuals and adipose tissues impair BCAA catabolism, consequently leading to raised BCAA levels in the circulation, as we found in serum samples (Fig. 5 and Additional file 1: Table S4). Moreover, we observed that both carnitines (C0 and C3) were positively associated with MetS, abdominal obesity, hypertriglyceridemia, and reduced HDL–C, respectively. C3 is a product of BCAA catabolism, which is impaired in MetS, thus C3 produced from BCAA should have been reduced in individuals with MetS. Our observation of raised C3 levels may be attributable to alternative causes (e.g. ß-oxidation of fatty acids with odd chain length, or the tricarboxylic acid cycle) (Fig. 5 and Additional file 1: Table S4). Raised BCAA and raised C3 levels have been found in individuals with insulin resistance, and the authors suggested that the BCAA catabolism might be impaired in adipose tissue but increased in other tissues such as skeletal muscle [47]. It remains an area of ongoing controversy and research whether BCAAs are a contributor or the result of insulin resistance [48]. The causal relationship between elevated levels of BCAAs, C0, and C3, and metabolic disturbances remains unclear. The regulation of BCAAs seems to be tissue-dependent and influenced by nutrition and health status, highlighting their potential role in obesity, diabetes, and multifactorial diseases such as MetS [49].
We observed that aromatic amino acids (Phe and Tyr) were positively associated with MetS. Phe is one of the essential amino acids in the human body, while Tyr is a semi-essential amino acid that can be synthesized from Phe. Three enzymes (PAH, phenylalanine hydroxylase, TH, tyrosine hydroxylase, and TAT, tyrosine aminotransferase) are reported to be responsible for the catabolism of these aromatic amino acids (Fig. 6 and Additional file 1: Table S5). PAH catalyzes the hydroxylation of Phe to Tyr [50]. Tyr can be further catabolized by TH to dopamine or by TAT to 4-hydroxyphenylpyruvate [51, 52]. Reduced expression of PAH was found in pigs with MetS [53], TH expression was found lower in individuals with central obesity [54], and lowered expression of TAT in the liver was reported in insulin-resistant mice [51].
We also found that Gly and Ser showed significant negative associations with MetS and its associated conditions, including increased waist circumference, hypertriglyceridemia, hypertension, and hyperglycemia (not for Ser). Gly is the simplest stable proteinogenic amino acid and can be biosynthesized from Ser in the body. Intake of Gly and Ser was reported to boost the rate of fatty acid oxidation and reduce triglyceride production [55, 56]. (Fig. 6). Human serine hydroxymethyltransferases (SHMTs) catalyze the conversion of Ser and Gly and vice versa, a process that is linked to the folate cycle [57, 58]. Additionally, the breakdown of Gly by the glycine cleavage system (GCS) is also associated with the folate cycle [59] (Fig. 6). Interestingly, the expression of two enzymes in the GCS, glycine decarboxylase (GLDC), and aminomethyltransferase (AMT), were found to be increased in obese rats [60] (shown with red arrows in Fig. 6). Folate treatment in clinical trials has shown a potential to improve insulin resistance and endothelial dysfunction in MetS patients [61]. Therefore, the increased catabolism of Gly may contribute to the reduction in Gly levels. Both Ser and Gly play important roles in maintaining cellular oxidative homeostasis [62]. Studies have shown that oral supplementation with Gly protects against oxidative damage in individuals with MetS [63]. On the other hand, reductions in Gly and Ser may lead to increased levels of reactive oxygen species (ROS) [64, 65], which could further inhibit the catabolism of Tyr and Phe by limiting the availability of the cofactor tetrahydrobiopterin (BH4) required by TH and PAH [66]. Moreover, TAT activity can be inhibited under oxidative stress conditions [67]. Consequently, the catabolism of Tyr and Phe could also be further reduced by higher ROS concentrations due to low Gly and Ser levels. In light of these findings, it seems that the raised concentrations of Tyr and Phe, and lowered levels of Gly and Ser observed in our study, are a consequence of the down-regulated expression and activity of PAH, TH, and TAT, and up-regulated GLDC and AMT expression (Fig. 6 and Additional file 1: Table S5). Moreover, raised levels of Gly and Ser decreased hypertension, abdominal obesity, and hypertriglyceridemia. This may indicate a protective effect of these two metabolites against MetS (Fig. 6 and Additional file 1: Table S5).
A limitation of our study is that one metabolite identified in the discovery study could not be replicated in SHIP-TREND-0 due to the use of different analytical kits. In addition, contrary to the KORA F4 study, the metabolomics quantification in the SHIP-TREND-0 study was performed only on a subgroup of participants who did not report having diabetes. Thus, the study was designed to include a relatively healthy cohort of participants, with non-diabetic individuals and, consequently, no diabetes-related diseases. Despite these structural differences in the study design, the study included participants with prediabetes, along with a sufficient number of metabolites. Moreover, the successful replication of the majority of the results indicated that these results are very robust and generalizable. An additional limitation of the study is that metabolites such as PCs and LPCs represent a group of possible metabolites rather than a single molecule. For instance, the metabolite lysoPC a C18:2 is known for its carbon chain length of the fatty acid and number of double bonds, but the position of the double bonds remains undefined. Therefore, the fatty acid in this LPC might be linoleic acid (C18:2 9Z, 12Z) or conjugated linoleic acid (C18:2 9E, 11E or C18:2 10E, 12Z) with different positions of the double bonds. For the PCs, the range of possible molecules is even wider, since we know the sum of the chain lengths and the sum of the double bonds of the two fatty acids in PC, but not which fatty acid is found at the sn1 or sn2 position. The level of the PCs or LPCs is determined by biosynthesis or catabolism, and the enzymes involved in these processes often exhibit a preference for specific fatty acids at specific positions. However, the substrate spectrum for all the enzymes is not yet fully known. Consequently, we were unable to construct networks for specific PCs or LPCs in our metabolic network analysis.
Conclusion
MetS is a complex and multifaceted syndrome, characterized by overlapping and interacting mechanisms that are not identical across individual phenotypes. A notable finding from our study is the identification of lysoPC a C18:2, a metabolite negatively associated not only with MetS but also with its five components. This metabolite might have the potential to be a key molecule in the prevention and treatment of MetS. Consequently, a greater and deeper comprehension of the lysoPC a C18:2 metabolic process, and its effect on the development of MetS is required. Additionally, the biological processes and enzymes involved in the network analysis of specific replicated MetS-associated metabolites can also be explored for preventive and therapeutic purposes.
Availability of data and materials
Both KORA and SHIP data sets are not publicly available because of data protection agreements but can be provided upon request through the KORA-PASST (Project application self-service tool, https://helmholtz-muenchen.managed-otrs.com/external) and the SHIP transfer office (https://www.fvcm.med.uni--greifswald.de/cm_antrag/index.php).
Abbreviations
- AMT:
-
Aminomethyltransferase
- BCAAs:
-
Branch-chain amino acids
- BCAT2:
-
Branched chain amino acid transaminase 2
- BCATm:
-
Mitochondrial branched chain amino acid aminotransferase
- BCKD:
-
Branched-chain ketoacid dehydrogenase
- BH4:
-
Tetrahydrobiopterin
- CV:
-
Coefficient of variance
- CVD:
-
Cardiovascular disease
- F4:
-
The first follow-up study of the KORA survey number 4
- GLDC:
-
Glycine decarboxylase
- GCS:
-
Glycine cleavage system
- HDL–C:
-
High density lipoprotein cholesterol
- KORA:
-
Cooperative Health Research in the Region of Augsburg
- LCAT:
-
Lecithin cholesterol acyltransferase
- LIPG:
-
Endothelial lipase
- LOD:
-
Limits of detection
- LPC:
-
Lysophosphatidylcholine
- LPCAT3:
-
Lysophosphatidylcholine acyltransferase-3
- MetS:
-
Metabolic syndrome
- PAH:
-
Phenylalanine hydroxylase
- PC:
-
Phosphatidylcholines
- PCC:
-
Propionyl-coA carboxylase
- QC:
-
Quality control
- ROS:
-
Reactive oxygen species
- RSD:
-
Relative standard deviations
- SARS-CoV-2:
-
Severe acute respiratory syndrome coronavirus 2
- SHIP:
-
The Health Study of Pomerania
- SHMT:
-
Serine hydroxymethyltransferase
- SLC7A5:
-
Solute Carrier Family 7 Member 5
- TAT:
-
Tyrosine aminotransferase
- TH:
-
Tyrosine hydroxylase
References
Alberti KGMM, Eckel RH, Grundy SM, Zimmet PZ, Cleeman JI, Donato KA, et al. Harmonizing the metabolic syndrome: a joint interim statement of the International Diabetes Federation Task Force on Epidemiology and Prevention; National Heart, Lung, and Blood Institute; American Heart Association; World Heart Federation; International Atherosclerosis Society; and International Association for the Study of Obesity. Circulation. 2009;120:1640–5.
Kao T-W, Huang C-C. Recent Progress in Metabolic Syndrome Research and Therapeutics. Int J Mol Sci. 2021;22:6862.
Saklayen MG. The Global Epidemic of the Metabolic Syndrome. Curr Hypertens Rep. 2018;20:12.
Bovolini A, Garcia J, Andrade MA, Duarte JA. Metabolic syndrome pathophysiology and predisposing factors. Int J Sports Med. 2021;42:199–214.
Grundy SM. Overnutrition, ectopic lipid and the metabolic syndrome. J Investig Med. 2016;64:1082–6.
Okopień B, Buldak L, Bołdys A. Fibrates in the management of atherogenic dyslipidemia. Expert Rev Cardiovasc Ther. 2017;15:913–21.
Laakso M, Kuusisto J. Insulin resistance and hyperglycaemia in cardiovascular disease development. Nat Rev Endocrinol. 2014;10:293–302.
Subramanian I, Verma S, Kumar S, Jere A, Anamika K. Multi-omics Data Integration, Interpretation, and Its Application. Bioinforma Biol Insights. 2020;14:1177932219899051.
Elhadad MA, Wilson R, Zaghlool SB, Huth C, Gieger C, Grallert H, et al. Metabolic syndrome and the plasma proteome: from association to causation. Cardiovasc Diabetol. 2021;20:111.
Wishart DS, Guo A, Oler E, Wang F, Anjum A, Peters H, et al. HMDB 5.0: the Human Metabolome Database for 2022. Nucleic Acids Res. 2022;50:D622–31.
Payab M, Tayanloo-Beik A, Falahzadeh K, Mousavi M, Salehi S, Djalalinia S, et al. Metabolomics prospect of obesity and metabolic syndrome; a systematic review. J Diabetes Metab Disord. 2022;21:889–917.
Yu Z, Zhai G, Singmann P, He Y, Xu T, Prehn C, et al. Human serum metabolic profiles are age dependent. Aging Cell. 2012;11:960–7.
Mittelstrass K, Ried JS, Yu Z, Krumsiek J, Gieger C, Prehn C, et al. Discovery of sexual dimorphisms in metabolic and genetic biomarkers. PLoS Genet. 2011;7:e1002215.
Xu T, Holzapfel C, Dong X, Bader E, Yu Z, Prehn C, et al. Effects of smoking and smoking cessation on human serum metabolite profile: results from the KORA cohort study. BMC Med. 2013;11:60.
Jaremek M, Yu Z, Mangino M, Mittelstrass K, Prehn C, Singmann P, et al. Alcohol-induced metabolomic differences in humans. Transl Psychiatry. 2013;3: e276.
Wang-Sattler R, Yu Z, Herder C, Messias AC, Floegel A, He Y, et al. Novel biomarkers for pre-diabetes identified by metabolomics. Mol Syst Biol. 2012;8:615.
Floegel A, Stefan N, Yu Z, Mühlenbruch K, Drogan D, Joost H-G, et al. Identification of serum metabolites associated with risk of type 2 diabetes using a targeted metabolomic approach. Diabetes. 2013;62:639–48.
Huang J, Huth C, Covic M, Troll M, Adam J, Zukunft S, et al. Machine learning approaches reveal metabolic signatures of incident chronic kidney disease in individuals with prediabetes and type 2 diabetes. Diabetes. 2020;69:2756–65.
Eick C, Klinger-König J, Zylla S, Hannemann A, Budde K, Henning AK, et al. Broad metabolome alterations associated with the intake of oral contraceptives are mediated by cortisol in premenopausal women. Metabolites. 2021;11:193.
Wang H, Wang Y, Li X, Deng X, Kong Y, Wang W, et al. Machine learning of plasma metabolome identifies biomarker panels for metabolic syndrome: findings from the China Suboptimal Health Cohort. Cardiovasc Diabetol. 2022;21:288.
Warmbrunn MV, Koopen AM, de Clercq NC, de Groot PF, Kootte RS, Bouter KEC, et al. Metabolite profile of treatment-naive metabolic syndrome subjects in relation to cardiovascular disease risk. Metabolites. 2021;11:236.
Ramakrishanan N, Denna T, Devaraj S, Adams-Huet B, Jialal I. Exploratory lipidomics in patients with nascent Metabolic Syndrome. J Diabetes Complications. 2018;32:791–4.
Ntzouvani A, Nomikos T, Panagiotakos D, Fragopoulou E, Pitsavos C, McCann A, et al. Amino acid profile and metabolic syndrome in a male Mediterranean population: a cross-sectional study. Nutr Metab Cardiovasc Dis. 2017;27:1021–30.
Rathmann W, Kowall B, Heier M, Herder C, Holle R, Thorand B, et al. Prediction models for incident type 2 diabetes mellitus in the older population: KORA S4/F4 cohort study. Diabet Med J Br Diabet Assoc. 2010;27:1116–23.
Völzke H, Schössow J, Schmidt CO, Jürgens C, Richter A, Werner A, et al. Cohort profile update: the study of health in Pomerania (SHIP). Int J Epidemiol. 2022;dyac034.
Yao Y, Schneider A, Wolf K, Zhang S, Wang-Sattler R, Peters A, et al. Longitudinal associations between metabolites and long-term exposure to ambient air pollution: results from the KORA cohort study. Environ Int. 2022;170: 107632.
Jourdan C, Petersen A-K, Gieger C, Döring A, Illig T, Wang-Sattler R, et al. Body fat free mass is associated with the serum metabolite profile in a population-based study. PLoS ONE. 2012;7: e40009.
Meisinger C, Strassburger K, Heier M, Thorand B, Baumeister SE, Giani G, et al. Prevalence of undiagnosed diabetes and impaired glucose regulation in 35–59-year-old individuals in Southern Germany: the KORA F4 Study. Diabet Med J Br Diabet Assoc. 2010;27:360–2.
Schauer B, Grabe HJ, Ittermann T, Lerch MM, Weiss FU, Mönnikes H, et al. Irritable bowel syndrome, mental health, and quality of life: Data from a population-based survey in Germany (SHIP-Trend-0). Neurogastroenterol Motil. 2019;31: e13511.
Lüdtke L, Jürgens C, Ittermann T, Völzke H, Tost F. Age-related macular degeneration and associated risk factors in the population-based study of health in Pomerania (SHIP-Trend). Med Sci Monit Int Med J Exp Clin Res. 2019;25:6383–90.
Völzke H, Alte D, Schmidt CO, Radke D, Lorbeer R, Friedrich N, et al. Cohort profile: the study of health in Pomerania. Int J Epidemiol. 2011;40:294–307.
Han S, Huang J, Foppiano F, Prehn C, Adamski J, Suhre K, et al. TIGER: technical variation elimination for metabolomics data using ensemble learning architecture. Brief Bioinform. 2022;23:bbab535.
Zacharias HU, Hertel J, Johar H, Pietzner M, Lukaschek K, Atasoy S, et al. A metabolome-wide association study in the general population reveals decreased levels of serum laurylcarnitine in people with depression. Mol Psychiatry. 2021;26:7372–83.
Lechner M, Höhn V, Brauner B, Dunger I, Fobo G, Frishman G, et al. CIDeR: multifactorial interaction networks in human diseases. Genome Biol. 2012;13:R62.
Wang B, Tontonoz P. Phospholipid remodeling in physiology and disease. Annu Rev Physiol. 2019;81:165–88.
Cole LK, Vance JE, Vance DE. Phosphatidylcholine biosynthesis and lipoprotein metabolism. Biochim Biophys Acta. 2012;1821:754–61.
Carioca AAF, Steluti J, de Carvalho AM, Silva AM, da Silva IDCG, Fisberg RM, et al. Plasma metabolomics are associated with metabolic syndrome: a targeted approach. Nutr Burbank Los Angel Cty Calif. 2021;83: 111082.
Law S-H, Chan M-L, Marathe GK, Parveen F, Chen C-H, Ke L-Y. An updated review of lysophosphatidylcholine metabolism in human diseases. Int J Mol Sci. 2019;20:1149.
Han MS, Lim Y-M, Quan W, Kim JR, Chung KW, Kang M, et al. Lysophosphatidylcholine as an effector of fatty acid-induced insulin resistance. J Lipid Res. 2011;52:1234–46.
Wahl S, Yu Z, Kleber M, Singmann P, Holzapfel C, He Y, et al. Childhood obesity is associated with changes in the serum metabolite profile. Obes Facts. 2012;5:660–70.
Ward-Caviness CK, Xu T, Aspelund T, Thorand B, Montrone C, Meisinger C, et al. Improvement of myocardial infarction risk prediction via inflammation-associated metabolite biomarkers. Heart. 2017;103:1278–85.
Badellino KO, Wolfe ML, Reilly MP, Rader DJ. Endothelial lipase concentrations are increased in metabolic syndrome and associated with coronary atherosclerosis. PLoS Med. 2006;3: e22.
Gauster M, Rechberger G, Sovic A, Hörl G, Steyrer E, Sattler W, et al. Endothelial lipase releases saturated and unsaturated fatty acids of high density lipoprotein phosphatidylcholine. J Lipid Res. 2005;46:1517–25.
Rohrmann S, Overvad K, Bueno-de-Mesquita HB, Jakobsen MU, Egeberg R, Tjønneland A, et al. Meat consumption and mortality—results from the European Prospective Investigation into Cancer and Nutrition. BMC Med. 2013;11:63.
She P, Van Horn C, Reid T, Hutson SM, Cooney RN, Lynch CJ. Obesity-related elevations in plasma leucine are associated with alterations in enzymes involved in branched-chain amino acid metabolism. Am J Physiol Endocrinol Metab. 2007;293:E1552-1563.
Newgard CB. Metabolomics and metabolic diseases: where do we stand? Cell Metab. 2017;25:43–56.
Newgard CB. Interplay between lipids and branched-chain amino acids in development of insulin resistance. Cell Metab Elsevier. 2012;15:606–14.
Libert DM, Nowacki AS, Natowicz MR. Metabolomic analysis of obesity, metabolic syndrome, and type 2 diabetes: amino acid and acylcarnitine levels change along a spectrum of metabolic wellness. PeerJ. 2018;6: e5410.
Reddy P, Leong J, Jialal I. Amino acid levels in nascent metabolic syndrome: a contributor to the pro-inflammatory burden. J Diabetes Complications. 2018;32:465–9.
Schuck PF, Malgarin F, Cararo JH, Cardoso F, Streck EL, Ferreira GC. Phenylketonuria pathophysiology: on the role of metabolic alterations. Aging Dis. 2015;6:390–9.
Valera A, Rodriguez-Gil JE, Yun JS, McGrane MM, Hanson RW, Bosch F. Glucose metabolism in transgenic mice containing a chimeric P-enolpyruvate carboxykinase/bovine growth hormone gene. FASEB J. 1993;7:791–800.
Fitzpatrick PF. Structural insights into the regulation of aromatic amino acid hydroxylation. Curr Opin Struct Biol. 2015;35:1–6.
Karimi M, Petkova V, Asara JM, Griffin MJ, Sellke FW, Bishop AR, et al. Metabolomics and the pig model reveal aberrant cardiac energy metabolism in metabolic syndrome. Sci Rep. 2020;10:3483.
Leite F, Lima M, Marino F, Cosentino M, Ribeiro L. Dopaminergic receptors and tyrosine hydroxylase expression in peripheral blood mononuclear cells: a distinct pattern in central obesity. PLoS ONE. 2016;11: e0147483.
Imenshahidi M, Hossenzadeh H. Effects of glycine on metabolic syndrome components: a review. J Endocrinol Invest. 2022;45:927–39.
Sim W-C, Kim DG, Lee W, Sim H, Choi Y-J, Lee B-H. Activation of SIRT1 by L-serine increases fatty acid oxidation and reverses insulin resistance in C2C12 myotubes. Cell Biol Toxicol. 2019;35:457–70.
Monti M, Guiducci G, Paone A, Rinaldo S, Giardina G, Liberati FR, et al. Modelling of SHMT1 riboregulation predicts dynamic changes of serine and glycine levels across cellular compartments. Comput Struct Biotechnol J. 2021;19:3034–41.
Luciano-Mateo F, Hernández-Aguilera A, Cabre N, Camps J, Fernández-Arroyo S, Lopez-Miranda J, et al. Nutrients in energy and one-carbon metabolism: learning from metformin users. Nutrients. 2017;9:121.
Leung K-Y, De Castro SCP, Galea GL, Copp AJ, Greene NDE. Glycine cleavage system H protein is essential for embryonic viability, implying additional function beyond the glycine cleavage system. Front Genet. 2021;12: 625120.
Simmons RM, McKnight SM, Edwards AK, Wu G, Satterfield MC. Obesity increases hepatic glycine dehydrogenase and aminomethyltransferase expression while dietary glycine supplementation reduces white adipose tissue in Zucker diabetic fatty rats. Amino Acids. 2020;52:1413–23.
Setola E, Monti LD, Galluccio E, Palloshi A, Fragasso G, Paroni R, et al. Insulin resistance and endothelial function are improved after folate and vitamin B12 therapy in patients with metabolic syndrome: relationship between homocysteine levels and hyperinsulinemia. Eur J Endocrinol. 2004;151:483–9.
Mahmood K, Emadi A. 1-C metabolism-serine, glycine, folates-in acute myeloid leukemia. Pharm Basel Switz. 2021;14:190.
Díaz-Flores M, Cruz M, Duran-Reyes G, Munguia-Miranda C, Loza-Rodríguez H, Pulido-Casas E, et al. Oral supplementation with glycine reduces oxidative stress in patients with metabolic syndrome, improving their systolic blood pressure. Can J Physiol Pharmacol. 2013;91:855–60.
Zhou X, He L, Zuo S, Zhang Y, Wan D, Long C, et al. Serine prevented high-fat diet-induced oxidative stress by activating AMPK and epigenetically modulating the expression of glutathione synthesis-related genes. Biochim Biophys Acta Mol Basis Dis. 2018;1864:488–98.
Ruiz-Ramírez A, Ortiz-Balderas E, Cardozo-Saldaña G, Diaz-Diaz E, El-Hafidi M. Glycine restores glutathione and protects against oxidative stress in vascular tissue from sucrose-fed rats. Clin Sci Lond Engl. 1979;2014(126):19–29.
Cheng C-W, Liu M-H, Tang H-Y, Cheng M-L, Wang C-H. Factors associated with elevated plasma phenylalanine in patients with heart failure. Amino Acids. 2021;53:149–57.
Adams SH. Emerging perspectives on essential amino acid metabolism in obesity and the insulin-resistant state. Adv Nutr. 2011;2:445–56.
Acknowledgements
The authors thank all the KORA and SHIP study participants for donating their time and blood. We thank all participants for their long-term commitment to the KORA study, the staff for data collection and research data management and the members of the KORA Study Group (https://www.helmholtz-munich.de/en/epi/cohort/kora) who are responsible for the design and conduct of the KORA study.
Funding
Open Access funding enabled and organized by Projekt DEAL. The KORA study was initiated and financed by the Helmholtz Zentrum München—German Research Center for Environmental Health, which is funded by the German Federal Ministry of Education and Research (BMBF) and by the State of Bavaria. Data collection in the KORA study is done in cooperation with the University Hospital of Augsburg. The German Diabetes Center is supported by the German Federal Ministry of Health (Berlin, Germany). This study was supported in part by a grant from the German Federal Ministry of Education and Research to the German Center for Diabetes Research (DZD). This project has received funding from the Innovative Medicines Initiative 2 Joint Undertaking (JU) under grant agreement No. 821508 (CARDIATEAM). The JU receives support from the European Union’s Horizon 2020 research and innovation programme and EFPIA. SHIP is part of the Community Medicine Research net of the University of Greifswald, Germany, which is funded by the Federal Ministry of Education and Research (Grants no. 01ZZ9603, 01ZZ0103, and 01ZZ0403), the Ministry of Cultural Affairs and the Social Ministry of the Federal State of Mecklenburg-West Pomerania.
Author information
Authors and Affiliations
Contributions
MS designed the study, analyzed the data, interpreted the results, and wrote the manuscript. SH contributed to analyze the data, interpretation of the results, and manuscript edition. KK and JH performed replication analysis, contributed to interpretation of data, and wrote part of the manuscript. GF, CM, and AR constructed networks in the CIDeR database, interpreted the results and wrote part of the manuscript. SY and MH contributed to the interpretation of the results and manuscript edition. AH, NF, MB, MD, MN, HV, GH, HJG, CP, JA, KS, WR, and AP participated in the acquisition of the KORA or SHIP studies data. MB, MD, MN, HJG, JA, AP contributed to manuscript edition. RWS designed the study, contributed to analyze the data, interpreted the results, and wrote the manuscript. All authors read and approved the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
KORA and SHIP studies were approved by the Ethics Committees of the Bavarian Medical Association in Munich and the Institutional Review Board of the University of Greifswald, Germany, respectively. All study participants provided written informed consent.
Consent for publication
Not applicable.
Competing interests
HJG has received travel grants and speakers honoraria from Fresenius Medical Care, Neuraxpharm, Servier and Janssen Cilag as well as research funding from Fresenius Medical Care. The other authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Table S1.
Metabolite quantification and normalization in KORA F4. The table presents metabolite abbreviations, analyte classes, and biochemical names of 163 metabolites in columns first to third. Detecting rate of each metabolite in 3,061 KORA F4 individuals is shown in the fourth column. Column fifth displays the median coefficient of variation of 114 quality control samples. The sixth column displays the percentage of 3061 KORA F4 study individuals with values at or above the limit of detection. The final column displays each metabolite’s used/excluded status. Table S2. Logistic regression results of MetS-associated metabolites with each of the 5 components in the KORA study. Odds ratioswith 95% confidence interval. Significant P-value < 0.05/56 = 8.93E-04 are shown in bold. Table S3. Metabolic network of PCs and LPCs. Table S4. Metabolic network of BCAAs. Table S5. Metabolic network of Ser, Gly, Phe, and Tyr.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Shi, M., Han, S., Klier, K. et al. Identification of candidate metabolite biomarkers for metabolic syndrome and its five components in population-based human cohorts. Cardiovasc Diabetol 22, 141 (2023). https://doi.org/10.1186/s12933-023-01862-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12933-023-01862-z