
Abstract
Pulmonary hypertension (PH) refers to a group of deadly lung diseases characterized by vascular lesions in the microvasculature and a progressive increase in pulmonary vascular resistance. The prevalence of PH has increased over time. Currently, the treatment options available for PH patients have limited efficacy, and none of them can fundamentally reverse pulmonary vascular remodeling. Stem cells represent an ideal seed with proven efficacy in clinical studies focusing on liver, cardiovascular, and nerve diseases. Since the potential therapeutic effect of mesenchymal stem cells (MSCs) on PH was first reported in 2006, many studies have demonstrated the efficacy of stem cells in PH animal models and suggested that stem cells can help slow the deterioration of lung tissue. Existing PH treatment studies basically focus on the paracrine action of stem cells, including protein regulation, exosome pathway, and cell signaling; however, the specific mechanisms have not yet been clarified. Apoptotic and afunctional pulmonary microvascular endothelial cells (PMVECs) and alveolar epithelial cells (AECs) are two fundamental promoters of PH although they have not been extensively studied by researchers. This review mainly focuses on the supportive communication and interaction between PMVECs and AECs as well as the potential restorative effect of stem cells on their injury. In the future, more studies are needed to prove these effects and explore more radical cures for PH.
Similar content being viewed by others

Introduction
Pulmonary hypertension (PH) is a serious chronic progressive disease characterized by dysfunction and vascular endothelial injuries of pulmonary arterioles [1, 2], with abnormal obliterative vascular smooth muscle cell proliferation [3], which can further progress to pulmonary vascular remodeling and right ventricular hypertrophy. These lesions may eventually lead to right heart failure and even death. Consistently, PH is defined as a mean pulmonary artery pressure (mPAP) ≥ 25 mmHg observed at right-sided heart catheterization (RHC). The 6th World Symposium on Pulmonary Hypertension proposed a recent hemodynamic definition of PH, mPAP > 20 mmHg, as the upper limit of normal pulmonary artery pressure [4], and the value is associated with a greater risk of mortality, with pulmonary arterial wedge pressure (PAWP) ≤ 15 mm Hg and pulmonary vascular resistance (PVR) ≥ 3 Wood units [5]. Depending on different causes of disease and clinical pathological features, the World Health Organization divides PH into 5 types: pulmonary arterial hypertension, PH due to left heart disease, PH due to lung diseases and/or hypoxia, PH due to pulmonary artery obstruction, and PH with unclear and/or multifactorial mechanisms. The latest hemodynamic definition can help to aid in early PH diagnosis.
The mechanism of PH has not been completely clarified. This disease is closely related to the abnormal structure and function of pulmonary microvessels, mainly including the following points: (1) hypoxia-induced pulmonary arteriolar contraction and thickening of pulmonary arteriolar media smooth muscle; (2) pulmonary inflammatory reaction; and (3) pulmonary microvascular injury and dysfunction. Among these mechanisms, endothelial cells appear to play a particularly important and crucial role. These cells are believed to reverse PH [6]. However, multiple treatments that target a series of endothelial secretions (e.g., endothelin (ET), nitric oxide (NO) and the prostacyclin pathway) and aim to regulate pulmonary vascular endothelial cells and smooth muscle cells have recently been used for PH therapy, thereby controlling the progression of the disease to a certain extent. It is still difficult to fundamentally recover the functions of pulmonary microvasculature and to reverse vascular remodeling, future PH treatment should focus mainly on reversing remodeling of pulmonary vascular diseases.
In recent years, cell therapy has attracted considerable clinical attention for the treatment of various diseases. Increasing studies have shown that cell therapy may provide a novel therapeutic approach to PH. In the treatment of lung disease, there has been constant progress in preclinical animal experiments of stem cell therapy. Numerous studies have provided evidence to demonstrate a potential therapeutic effect of stem cells in acute lung injury (ALI), chronic obstructive pulmonary disease (COPD), acute respiratory distress syndrome (ARDS) and pulmonary fibrosis (PF) [7,8,9,10]. To date, PH remains an incurable disease, and cell therapy represents an important future direction for research. As an important platform for cell therapy, stem cells have recently become a widespread focus of study for an increasing number of researchers. This review examines the mechanism underlying the impact of stem cells on reversing vascular remodeling in the treatment of PH from the root.
PMVEC dysfunction causes PH
Pulmonary microvascular endothelial cells (PMVECs) are thin single cells arranged in a layer on the inside of blood vessels and are the most abundant cells in lung tissue (40%) [11]. PMVECs are essential in maintaining the barrier function of the pulmonary microvasculature [12]. The pulmonary alveolar capillary barrier is formed jointly by PMVECs and alveolar epithelial cells (AECs) [13], the damage of either of which will have an impact on pulmonary functions. As the primary barrier, PMVECs are also adversely affected first. For example, reactive oxygen species (ROS) and oxidative stress are important mechanisms underlying the development of many lung diseases, while microvascular endothelial cells are critical target cells for ROS, which may have a higher sensitivity to ROS in the lung than in other tissues [14]. Normally, pulmonary capillaries are poorly permeable. However, where there is damage or apoptosis to PMVECs, cell dysfunction or defects will occur. As a result, there will be a change in the permeability of pulmonary capillaries [15]. Increased pulmonary microvascular permeability is a common characteristic of many pulmonary diseases [16,17,18].
Apoptosis and injuries of PMVECs have been reported to play an essential role in the development of numerous lung diseases. Pulmonary edema is a hallmark pathological feature of ALI [19], and pulmonary microvascular permeability is an important cause of noncardiogenic pulmonary edema [20]. Evidence has shown that diffuse PMVEC injury causes destruction of the pulmonary microvascular endothelial barrier, and enhanced permeability is responsible for the interstitial edema associated with ALI [21]. In a study on sepsis, lipopolysaccharide (LPS) accelerated the rate of apoptotic cell death in PMVECs via activation of yes-associated protein, leading to microvascular hyperpermeability and ALI under sepsis [22]. Microvascular injuries have also been recognized as an important event in the evolution of PF [23]. A few subsequent empirical studies of PF have confirmed this conclusion through the specific marker detection of PMVEC lesions [24], cell morphology and activity [25]. In the natural course of early COPD, the pulmonary microvascular blood flow is overall reduced in any of the subjects accompanied by apoptosis of PMVECs [26]. A previous study demonstratedthat cigarette smoke extract induces apoptosis of PMVECs by decreasing mitochondrial respiration in cell models, which in turn induces the occurrence of COPD [27]. This demonstrates that changes in pulmonary microvascular permeability caused by pulmonary microvascular damage are a common feature of many pulmonary diseases and may be a very important inducing factor.
PH is a progressive disease of the pulmonary microvascular system, with the fundamental pathological features of progressive loss of pulmonary microvessels and remodeling of pulmonary microvessels [28, 29] as well as abnormal vasoconstriction [30]. Remodeling of distal pulmonary vasculature refers to the structural changes in the blood vessels of the pulmonary artery branch region, including the narrowing of pulmonary arterioles, thickening of the muscular layer, and proliferation of endothelial cells. The thickening of pulmonary arterioles and inappropriate pulmonary vasoconstriction restrict the normal flow of pulmonary blood, resulting in increased pulmonary arterial pressure and further exacerbating the pathological changes in pulmonary vasculature. The increase of pulmonary vascular resistance caused by distal pulmonary vascular remodeling is a key cause of increase of pulmonary artery pressure [31].
PMVEC dysfunction is believed to be at the root of PH [32]. During pregnancy, changes in certain proinflammatory cytokines or vascular signals in pregnant women may promote the development of alveolar septal fibrosis and impair the development of pulmonary microvasculature, thus leading to neonatal PH [33]. And models induced by the endothelial toxin monocrotaline (MCT) and hypoxia are commonly used in experiments established in animal models of PH, both of which have been shown to trigger initial apoptosis and damage to endothelial cells [34,35,36]. When the PMVECs show abnormal endothelial functions due to other factors, it may lead to pulmonary vasoconstriction and remodeling of the distal pulmonary vasculature through the following pathological alterations:
(1) Pulmonary microvascular permeability: Under pathological conditions such as a high glucose state, inflammatory infiltration, hypoxia or hyperoxia, PMVECs are subjected to damage, leading to apoptosis. This results in the release of injury markers such as CD31, as well as pro-inflammatory cytokines like tumor necrosis factor-alpha (TNF-α) and interleukin-6 (IL-6). Additionally, the expression of the cell adhesion protein VE-cadherin is decreased, thereby compromising the structural integrity and adhesive capacity of PMVECs; this phenomenon leads to a bare capillary basement membrane [37,38,39,40,41] and thus increases pulmonary microvascular permeability. In addition, inflammatory cells and platelets synthesize and release large amounts of vascular endothelial growth factor (VEGF) under inflammatory conditions [42], which can significantly enhance vessel permeability [43]. When the pulmonary microvascular permeability increases, proteins, inflammatory cells, and inflammatory factors can leak into the surrounding tissues [44,45,46]. Several inflammatory factors, including PDGF, TNF-α, IL-6, IL-8, and TGF-β, among others, can leak and stimulate excessive proliferation of smooth muscle cells. PDGF is a potent cell proliferation factor that can increase smooth muscle cells by a factor of 2.3 in 48 h by upregulating transient receptor potential channel protein 6 (TRPC6) [47]. In animal models of PH, it has been found that the expression level of platelet-derived growth factor receptor-beta (PDGFR-β) significantly increases and is an important factor in promoting smooth muscle cell proliferation through pulmonary microvascular leakage [48, 49].
(2) Imbalance in the release level of the regulator of vasomotor activity: Under normal conditions, PMVECs synthesize and secrete various vascular regulatory factors, playing a crucial role in the regulation of vascular dilation and constriction.
NO is a biologically active gas molecule synthesized by nitric oxide synthase (NOS) in endothelial cells, neurons, and other cells. It possesses diverse biological functions, including vasodilation, inhibition of platelet aggregation, and anti-inflammatory effects. NO can promote vasodilation by relaxing smooth muscle cells, thereby increasing blood flow and improving vascular function. In PH disease, it has been found that the production of endogenous NO in the lung is reduced [50, 51].The synthesis of NO is influenced by many factors. The stable expression of the mechanosensitive cation channel Piezo2 is essential for maintaining intracellular calcium influx and NO synthesis in PMVECs. When there is an imbalance between the TGF-β and bone morphogenetic proteins (BMP) signaling pathways, the expression of Piezo2 is reduced, leading to decreased phosphorylation of NOS and reduced NO synthesis. Consequently, this may weaken the inhibitory effect of cGMP-mediated smooth muscle cell proliferation, resulting in increased pulmonary muscularization [52].
Cav-1 is mainly expressed in PMVECs and plays an important role in maintaining pulmonary microvascular permeability and development [53, 54]; it can directly bind to the endothelial nitric oxide synthase (eNOS) and regulate its activity and stability by affecting the phosphorylation status of eNOS, further modulating its function [55]. The abnormal expression and dysfunction of Cav-1 protein is associated with PH [56, 57]. In the study of PMVECs, it was found that under the influence of inflammatory mediators such as IL-6, PMVECs were damaged, resulting in the loss of the expression of Cav-1 protein and the uncoupling of eNOS, failure to synthesize NO normally, and excessive peroxynitrite, which led to the loss of BMP receptor-2 (BMPR2), and activated TGF-β/pSMAD2/3 signaling pathway, thus forming pulmonary vascular remodeling [58]. BMPR2 is an anti-apoptotic factor of endothelial cells, and its mutation and loss is an important genetic basis for PH [59, 60].
Hyperconstriction of pulmonary blood vessels is another important pathological feature of PH. Current research indicates that ET the strongest arteriovenous vasoconstrictor peptide [61], which is produced by the breakdown of big endothelin by endothelin converting enzyme on endothelial cells [62]. ET-1, one of the three isomers of ET, is mainly synthesized by endothelial cells and is a vasoactive factor regulating pulmonary vascular tension [63]. Its combination with ET-1 receptor type A (ETA) in pulmonary blood vessels can lead to pulmonary vascular contraction, smooth muscle cell proliferation and pulmonary fibrosis [64]. Under physiological conditions, ET-1 levels were maintained at low levels due to NO inhibit the expression of the ET-1 and the scavenging effect of the ET-1 receptor type B (ETB) in the lungs [65, 66]. Elevated ET-1 levels have been observed in both clinical PH patients and experimental PH animal models in lung tissue and systemic plasma [67, 68]. Moreover, in most PH patients, the clearance effect of ETB did not change, so excessive synthesis is the main reason for the increase of ET-1 [69].The main source of increased ET-1 synthesis in PH is PMVECs [70]. In in vitro experiments, TGF-β and BMP9 can promote the synthesis of ET-1 by inducing the phosphorylation of Smad1/5 in human PMVECs, among which BMP9 is more effective and rapid, and can induce the phosphorylation of Smad2 at the same time [71, 72]. BMPR2 loss or knockdown can promotes the signaling effects of BMP9 and TGF-β in PH [73].
The interaction between AECs and PMVECs
AECs, comprised of alveolar type I (ATI) and type II epithelial (ATII) cells, play an important role in maintaining lung integrity and homeostasis. These cells begin to appear during the tubule-forming stage of lung development; among them, ATI cells are flat and cover more than 95% of the gas exchange surface. The ATI cells not only facilitate the ejection of sodium and water from the alveoli but are also involved in removing the amino acids from the airway [74]. These cells cannot proliferate; they are replaced by ATII cells that transdifferentiate into ATI cells when they are injured [75]. ATII cells are also regarded as progenitors of the alveolus and play an important role in the lungs innate immune response [76]. ATII cells can secrete surfactant, which reduce alveolar surface tension and prevent alveolar collapse [77]. In the mature lung, the alveolar walls are covered by squamous ATI cells and round secretory ATII cells. Although ATII cells occupy only a small surface area, they far outnumber ATI cells [78].
The alveolar epithelium and its adjacent pulmonary microvascular endothelium, as well as the extracellular matrix between them, together constitute the alveolar-capillary barrier and maintain the normal fluid exchange and gas exchange of the alveoli. The relationship between alveolar epithelium and pulmonary microvascular endothelium is very close, and the two influence and maintain each other. Because of the complexity of the pulmonary alveolar system, we know little about the interactions between AECs and PMVECs, but many in vitro culture experiments on ATII cells and PMVECs have shown epithelial-endothelial interactions.
Stimulated by endotoxins, pro-inflammatory cytokines, and plasma of sepsis patients, the protein permeability of human PMVECs increases. However, ATII cell line A549 can counteract this damage by releasing soluble factors suspected to be lipids, thereby reducing the permeability of human PMVECs stimulated by endotoxins by 39 ± 4% to 100 ± 3% [79]. This repairing effect of AECs on PMVECs was further confirmed in the study. Studies have shown that under normal physiological conditions, A549 cells and ATI-like cells release lipid mediators S1P and maintain the barrier function of human PMVECs by binding to S1P1 receptors, while under the stimulation of endogenous toxins, A549 cells and ATI-like cells increase the release of lipid mediators S1P and PGE2. Binding to the S1P1 receptors and EP4 receptors mediates the effect of enhancing the barrier function of human PMVECs [80]. In addition, an interesting phenomenon was found in this study: dexamethasone could block the release of PGE2 in A549 under normal conditions but could not block the release of PGE2 in ATI-like cells, which may indicate that there are other pathways in ATI cells to synthesize PGE2. Alveolar epithelial cells are involved in repairing the barrier function of PMVECs by releasing S1P. In previous studies, PMVECs have been shown to be able to release S1P in the case of lung injury. By binding the S1P receptor S1PR2 on the alveoli and initiating nuclear translocation of the transformation regulator yes-associated protein (YAP), the transformation of ATII cells into ATI cells is promoted [81]. Therefore, whether ATII cells will promote their own transformation through SIP to complete repair is expected to be further proved by relevant studies. Since 2019, the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) has been spreading globally. ATII cells represent the primary cellular targets of SARS-CoV-2 infection, while PMVECs also incur damage. Nonetheless, this impairment is not direct in nature; SARS-CoV-2 potentially instigates disruption within the mitochondria of ATII cells, subsequently precipitating an overabundance in the generation of ROS within these mitochondria. This sequence of events consequently stimulates the production of pro-inflammatory cytokines IL-1α, which are subsequently released into the intercellular space, thereby exerting an influence upon adjacent PMVECs. As a result, PMVECs undergo mitochondrial fragmentation, experience diminished Golgi apparatus integrity, and sustain compromised expression of the barrier-associated protein VE-cadherin [82]. In hypoxic conditions, AECII similarly exhibit deleterious effects on PMVECs. The study reveals that the ISM1 gene is upregulated in a low-oxygen environment. In comparison to the monoculture model of PMVECs, the co-culture model of PMVECs and AECII results in a further 37% increase in PMVEC permeability. This can be attributed to the overexpression of the hypoxia-inducible factor HIF-1α in AECII, which, by binding to regulatory elements, upregulates the expression of the permeability-associated gene ISM1, thus contributing to the enhanced PMVEC permeability [83].
In summary, there are complex interactions between AECs and PMVECs (Fig. 1). AECs and PMVECs maintain and interact with each other by releasing substances such as cytokines and lipids. When the body is in pathological surroundings, if the substances that regulate the secretion of ATII can favorably affect PMVECs by the paracrine effect of AECII, the damage to PMVECs can be reduced and the endothelial cell function of PMVECs can be repaired. The PMVECs that function normally can in turn promote the transformation of ATII to ATI and then repair injury of the lungs. Therefore, intervening in the abovementioned secretion will be an effective practice for the treatment of certain lung diseases including PH.

AECs and PMVECs interact with each other. Under conditions of injury, AECs have damaged organelles and dysfunction and regulate the permeability of pulmonary microvessels by secreting a number of mediators. The result of this adjustment will have different effects depending on the mediators. Similarly, PMVECs secrete lipids to affect alveolar repair in the state of injury. By Figdraw (www.figdraw.com)
Some problems in the current treatment of PH
The drug treatment system for PH is very complex, with many side effects and lack of sufficient stability. Patients usually receive more than one drug to treat PH. By directly regulating endothelial cells or acting on pulmonary artery smooth muscle cells (PASMCs), the drugs regulate blood vessels, either inhibiting vasoconstriction or promoting vasodilation, but they are unable to reverse vascular remodeling or restore the function of endothelial cells and AECs aiming at improving right ventricular function, that is, treating PH by addressing the root cause. Some medications require chronic use, resulting in other issues, such as decreased patient compliance, unknown long-term efficacy, and impaired organ function. In addition, PH primarily affects the arteries in the lungs, which is characterized by diffusion impairment, making it difficult for the drug to reach the intended area [84].
The currently available surgical options for the treatment of PH mainly include balloon pulmonary angioplasty (BPA), pulmonary artery thromboendarterectomy (PTE), and lung transplantation. These options have significantly improved patients’ symptoms and quality of life. However, some challenges remain with these options. PTE is currently the most effective and the preferred option for the treatment of CTEPH. Some patients with CTEPH can be completely cured with this approach. However, it is highly demanding for the surgeon’s techniques and experience, and 40% of the patients would not be indicated for this procedure as the position of the thrombus is difficult to reach [85]. BPA can significantly improve the patient’s symptoms and hemodynamic indicators, yet is associated with the complications primarily including pulmonary vascular injury and reperfusion pulmonary edema [86]. The 5-year survival rate following lung transplantation for PH is 45–50%; however, it is limited by the small number of donors and a high risk.
Possible role of stem cell therapy in the repair of PH
In recent years, new research has been conducted on the role of stem cells in the treatment of lung diseases. Currently, more than 50 clinical trials involving stem cell therapies for pulmonary diseases have been registered on ClinicalTrials.gov. Over the last years, much progress has been made in the research field of stem cell therapy in PH. Currently, there are three major groups of commonly studied stem cells: embryonic stem cells (ESCs), induced pluripotent stem cells (iPSCs) and mesenchymal stem cells (MSCs).
Embryonic stem cells
ESCs originate from the germinal vesicle, the earliest phase of embryo development. Specifically, ESCs are undifferentiated cells in the blastemal in early embryonic development (5th to 7th days after fertilization) and are characterized by infinite proliferation, self-renewal and a multidirectional differentiation capacity [87].
As mentioned above, ATII cells can promote the repair of the barrier function of pulmonary microvascular endothelial cells and reduce the permeability of pulmonary microvascular endothelial cells through paracrine effect, so transplantation of ATII that can secrete specific substances can be a therapeutic approach. Therefore, cell purity is the key problem to be solved first. Regardless of whether the tissue is cultured in primary or differentiated from ESCs, it is difficult to obtain sufficient purity and quantity of ATII cells. The production of ATII cells isolated from primary lung tissues is often low, prone to phenotypic loss, and accompanied by a loss of proliferative capacity [88, 89]. The use of ESC differentiation to obtain ATII cells has been studied for a long time.
In 2002, Ali et al. cultured embryoid bodies (EBs) derived from the differentiation of mouse ESCs with serum-free small airway growth medium (SAGM). On the 3rd and 14th days of culture, surfactant-protein-C (SPC) mRNA, a specific marker of ATII cells, was detected in cells. The purity of cells obtained in the experiment was quite low, and it relied heavily on the formation of EBs in the early stage. Moreover, a long period of time is required for differentiation. Nevertheless, this study revealed the possibility of ESCs differentiating into lung tissue [90]. In the follow-up study, the team tried to raise the output and realize more accurate differentiation status by recombining the effective components in SAGM. After removal of retinoic acid and triiodothyronine, the expression level was increased threefold. However, there was a large deviation in the repeatability of the data [91]. Although the addition of activin A could increase the output, the positive rate was quite low, as shown by the flow results, and the complete culture process took 35 days [92]. In 2007, our research team used a disaggregated seeding method to shorten the culture time required for detection of SPC mRNA transcripts without EB formation and pioneered a genetic screening directed differentiation procedure for human embryonic stem cells (hESCs), screening ATII cells with 99.6% purity and validating their cellular functions, cell biological features and morphological characteristics, as well as the expression of surfactant-protein-A (SPA), surfactant-protein-B (SPB) and SPC, which was detected by immunofluorescence staining, and the synthesis and secretion of the complement proteins C3 and C5 were detected on the 15th day of cell differentiation, which provided a reliable source of support for the late application of ATII cells [93]; cells obtained in this way show intervention and protection in ALI while recovering weight and arterial blood oxygen saturation, along with improved survival [94]. The differentiation of ESCs in vivo or in vitro is also regulated by the microenvironment [95, 96]. SPC protein expression was detected after 5 days of coculture with mouse lung embryonic mesenchymal cells, but this positive reaction did not occur when cocultured with mouse intestinal embryonic mesenchymal cells, indicating that the lung microenvironment may be more suitable for promoting the differentiation of embryonic stem cells into ATII cells [97].
ESCs can also differentiate into vascular endothelial cells. FOXF1 is a transcription factor that plays a pivotal role during embryonic development [98]. FOXF1 and its transcriptional target protein c-KIT are expressed in endothelial progenitor cell populations within the human pulmonary microvasculature [99]. In a study published in June this year, researchers used electroporation to introduce the FOXF1 and c-KIT genes into a mouse ESC line. Subsequently, they directed the differentiation of these stem cells into endothelial cells (CD31 + CD45-). Notably, these c-KIT + FOXF1 + EPCs, when transplanted into a mouse model, integrated into the alveolar microvasculature, expressing the alveolar capillary endothelial specific markers CAP1 and CAP2. The study also revealed that, in the lung tissue of mice transplanted with c-KIT + FOXF1 + EPCs, donor cells in the distal lung regions co-expressed early lung epithelial cell markers such as Nkx2.1(TTF1), SRY-box transcription factor 2 (SOX2), and SRY-box transcription factor 9 (SOX9). This suggests that the donor cells contain not only endothelial progenitors but also early lung progenitors, holding the potential to generate multiple types of lung cells in the recipient lungs. In addition, it is important that c-KIT + FOXF1 + EPC obtained through directed induction differentiation can effectively implant pulmonary microvascular in mice with hyperoxy-induced injury, successfully increasing the number of endothelial cells and restoring the density of pulmonary microvessels [100]. However, the differentiation method used in this research achieved only a 4% yield of endothelial cells.
The differentiation of ESCs into endothelial cells is regulated by transcription factors such as HIF. As a critical transcriptional regulator of lung tissue sensing oxygen changes in a hypoxic environment, HIF can regulate the expression of a range of hypoxia-related target genes and affect the process of pulmonary vascular remodeling. IL-33, a key factor in multiple types of pulmonary inflammation and vascular remodeling, promotes pulmonary vascular remodeling by increasing the expression of HIF-1α in PH [101]. Interestingly, some studies have demonstrated that HIF-1α promotes the differentiation of ESCs into functional vascular endothelial cells by initiating the transcription of ets variant 2 (ETV2) and simultaneously activating the NOTCH1 signaling pathway [102].
Therefore, the differentiation of ESCs into ATII cells or PMVEC by directional induction comes with significant research prospects. In summary, the following two pathways may be available for treating PH with ESCs: ① ESCs can directionally differentiate into normal ATII cells or PMVEC and restore the signal transduction between endothelium and epithelium through their interaction, thus restoring normal barrier function and alveolar epithelial function; ② ESC-derived ATII cells or PMVECs can be modified to perform specific functions against different pathogenic factors using gene-modification techniques, such as restoring the sensitivity of endothelial progenitor cells to hyperoxic environment and stimulating neovascularization and alveolization under hyperoxic environment. In addition to their therapeutic potential, ESCs can also be used in combination with gene editing techniques to create different models of PH, including the hereditary form of the condition. This approach allows researchers to investigate the roles of various genes, which, in turn, provides significant assistance in the study of targeted treatments for PH.
ESCs have the unique properties of unlimited self-renewal and pluripotency, this ESCs’ primary advantage is also its primary disadvantage. As mentioned above, the differentiation of ESCs is affected by the microenvironment. Teratomas can form when ESCs are in an ectopic microenvironment [103]. Overall, tumorigenicity is a potential hurdle that could halt ESC research. ESC-expressed Ras is an identified oncogene also required for endothelial differentiation of mouse ESCs [104]. MicroRNA-mediated regulation of Sox2 is an embryonic stem cell-specific transcription factor that controls self-renewal, pluripotency, and proliferation [105] and has been shown to function as an oncogene [106]. These problems must be ruled out, and the development of tumors must be avoided by preparing highly pure, directionally differentiated cells. In addition, potential immunological rejection and chromosomal variation are critical limitations [107,108,109]. Although our research team has made cells resistant to CD8 + T-cell-mediated destruction by targeting blockade of human leukocyte antigen (HLA) light chain-β2 microglobulin expression without affecting the self-renewal capacity, genomic stability or pluripotency of hESCs, this has only minimized hESC immunogenicity. The production of completely immunogenic hESCs continues to be a major challenge. Furthermore, due to the particularity in defining and sourcing hESCs, their clinical application is surrounded by ethical controversy.
Some scholars argue that an embryo should be considered a human person from the moment of conception, and thus acquiring hESCs through embryo destruction is tantamount to murder. Therefore, regardless of the ultimate goal of hESC research, such disregard for life is unacceptable. On the other hand, supporters of hESC research believe that the definition of humanity should include social attributes. During the first 14 days of development, embryos have no nerves or brain, and at this stage, embryos are devoid of consciousness and the ability to engage in social relationships. Therefore, extracting ESCs during this period, before the embryo begins to develop its bodily plan, should be permitted. Embryos at this stage should not be defined as complete individuals but rather as “a potential human being,” symbolizing a form of “special respect.” However, this notion continues to face widespread opposition [110].
To balance technology and ethics and promote the development of hESC technology, many countries around the world have enacted their own regulations on hESC research, but the stringency varies greatly between different countries. Germany has always maintained a very conservative attitude towards the application and research of hESCs. It wasn’t until December 2002 that the German government first approved the entry of hESC lines into Germany. Before this, Germany had explicitly prohibited any hESC research by enacting the Embryo Protection Act [111]. Influenced by political factors, the United States has been cautious in the legal regulations of hESC research. The U.S. has long restricted funding for hESC research and prohibited the production of any hESC strains that require embryo destruction. Research using hESC strains is limited to those created before August 9, 2001, and regulation of hESC research projects funded by private or non-federal funds is relatively lenient [112]. The United Kingdom is one of the earliest countries in the world to legislate and regulate hESC research. After a prolonged period of ethical debate, the government has formulated relatively enlightened and comprehensive regulatory policies for hESC research. In 1990, the UK enacted the Human Fertilisation and Embryology Act, which, by reference to the “14-day” principle proposed by the US Department of Health, Education, and Welfare Ethics Advisory Board in 1979, permits the in vitro creation, preservation, and utilization of hESCs for research purposes. However, the ESCs used in research cannot develop in vitro for more than 14 days, and all hESC research conducted in the UK is only legal with approval from the Human Fertilisation and Embryology Authority (HFEA) [113]. Unlike Western religious beliefs, in China, influenced by millennia of doctrine, most people do not consider embryos sacred from the moment of conception. However, this does not mean that the Chinese do not respect embryos; rather, they believe that embryos have a special moral status. When they can be used to treat diseases, the Chinese can tolerate the destruction and use of human embryos. Therefore, research on hESCs is supported in China, and in recent years, this research direction has been designated as a key focus of national research and development plans. In terms of regulation, China primarily relies on the Measures for the Ethical Review of Biomedical Research Involving Human Subjects and the Ethical Guidelines for Human Embryonic Stem Cell Research to impose ethical restrictions on research involving hESCs. However, there is no comprehensive legal framework to regulate the research on hESCs or to penalize research activities that violate regulations. Since the “gene-edited babies” incident in 2018, China has been continuously revising its laws to improve its regulatory methods, but there is still a long way to go [114].
Currently, one of the unresolved issues may be how to reasonably obtain hESCs, ensuring that their sources comply with medical ethics and reach a unified consensus on religious beliefs. In addition, comprehensive regulations are essential to provide normative guidance for hESC research.
Induced pluripotent stem cells
First reported by Kyoto University in Japan in 2006, iPSCs are now widely used in the study of respiratory diseases. Transplantation of ATII cells formed by the differentiation of human iPSCs into a rat model of pulmonary fibrosis was shown to effectively inhibit the increase in α smooth muscle actin (α-SMA) and transforming growth factor-β (TGF-β) and reduce collagen content, thereby reducing pulmonary fibrosis and improving the alveolar structure [115]. In addition, a number of studies have confirmed the ability of iPSCs to battle ALI. In an experiment by Dongqi Xing et al., intravenous infusion of iPSC-derived pulmonary vascular endothelial cells expressing CXCR1/2 or CCR2/5 from rats significantly inhibited ALI induced by LPS [116]. Vincent Yi-Fong Su et al. showed that iPSCs reduced adhesion kinase activity via tissue inhibitors of metalloproteinase 1 (TIMP-1) and focal adhesion kinase (FAK)/Snail pathways and reduced LPS-induced histological signs, protein leakage and proinflammatory cytokine expression in ALI, as well as improved endothelial cell permeability [117].
The technique of creating iPSCs has good therapeutic potential, as it may enable the replacement of malfunctioning organs with newly differentiated cells. In addition to already being used to derive respiratory epithelial cells, vascular endothelial cells, and vascular smooth muscle cells [118, 119], iPSCs have also been applied in the study of lung development and vascular modeling [120].
In recent years, the role of IPS in PH has also begun to emerge in the field of pulmonary disease research. In an MCT-induced PH rat model, iPSCs and iPSC-conditioned medium could improve hemodynamics and right ventricular hypertrophy and significantly reduce the expression of the inflammatory markers CD68, MHC-II, IL-1β, IL-6, IL-12α, IL-12β, IL-23 and interferon γ [121].
In terms of differentiation, iPSCs also show potential for the treatment of PH. IPSC-derived ECs (iPSC-ECs) have many similarities with natural pulmonary arterial endothelial cells and can be used as substitutes for drug screening in PH. In animal experiments, IPSC-differentiated ECs were injected into the pulmonary vascular lumen of rats, restoring 85–95% of pulmonary microvascular barrier function and regenerating functional blood vessels [122]. ATII cells derived from iPSCs, which were obtained by Bernard Thébaud’s research team, with a 96% expression rate of SPB positivity at day 25, were screened and transplanted via the airway to prevent lung injury produced by high oxygen concentrations in mice [123]. The differentiation of iPSCs into ATII cells is not very efficient. IPSC-ATII overexpressing miR-22 was found to have increased SPC-positive cell expression (49.12% vs. 37.21%) compared to fibroblast factor-2-induced ATII cells at day 5 of culture by Fan Tao’s research team for comparison but was still inferior in cell function to primary cultured ATII cells [124]. As with embryonic stem cells, tumorigenicity represents a major obstacle to the clinical application of iPSCs [125, 126]. Our research team used genetic modification to precisely express Oct4, Sox2, cMyc and Klf4, followed by transgenic removal of the exogenous reprogramming factors. The resulting iPSC-differentiated ATII cells are homologous and have the structure and biological function of normal ATII cells, making this technique effective in protecting and regenerating damaged AECs without being tumorigenic [127].
Therefore, it demonstrates that lung injury repair through differentiation into endothelial cells or ATII cells is also an effective approach for the therapeutic use of iPSCs to treat PH. Additionally, iPSCs can also contribute to the treatment of PH in other ways as follows: ① Disease modeling: Studies have shown that ECs differentiated from heritable pulmonary arterial hypertension (HPAH) patients-derived iPSCs have similar cell morphology to the donor’s pulmonary artery endothelial cells (PAECs) and also exhibit functional abnormalities such as decreased cell migration ability and diminished ability to generate blood vessels. Among 8 donors, 2 patients with HPAH-derived iPSC-ECs and PAECs showed a significant increase in blood vessels after treatment with the BMPR2 signaling activators FK506 and Elafin [128], indicating the feasibility of using iPSC-ECs as a substitute for PAECs in establishing PH models. ② Drug screening: Based on the above findings, the Caspase detection of cell apoptosis was utilized to screen drugs for PH iPSC-ECs. Among six compounds, some tyrosine kinase inhibitors (TKIs) were identified as superior to others, including imatinib, in improving PH vascular function [129]. ③ Individualized treatment: Using sequencing technology to analyze PAECs derived from idiopathic pulmonary arterial hypertension (IPAH) patients, as well as endothelial cells differentiated from iPSCs originating from fibroblasts, evaluating differences in gene expression and methylation changes, elucidating the relationship between variant genes and the pathobiology of PH. Subsequently, correcting genes through zinc finger nucleases technology and comparing changes in vascular function after restoring normal gene expression. This study may offer a direction for personalized therapy for PH [130].
Meanwhile, iPSCs are also applied in the study of protective modifiers for HPAH. Mutations in BMPR2 can be found in about 70% of patients with HPAH and around 20% of patients with IPAH [131]. It has been shown that genetic deletion of endothelial BMPR2 activates endothelial apoptosis and causes PH in endothelial cells [132]. For familial PH, however, only 20% of carriers with BMPR2 gene mutations exhibit clinical symptoms, indicating that the cellular function can be protected from BMPR2 mutations in the presence of certain protective modifiers. Moreover, the comparative analysis found that FPH-derived iPSC-ECs showed significant decreases in cell adhesion and survival compared to unaffected carrier-derived iPSC-ECs. At the same time, the RNA analysis revealed two compensatory signals, i.e., pP38 and BIRC3 [133]. Furthermore, as iPSCs share the same genetic background as the donors, which largely avoids the issue of immune rejection and boasts strong genetic operability, investigators can attempt to repair these mutations using CRISPR-Cas9 or other gene editing techniques, thereby developing potential gene therapies that provide significant clinical value in the treatment of patients with HPAH.
Previous results showed that iPSC-derived MSCs (iPSC-MSCs) exhibited stronger proliferation than MSCs [134], and iPSC-MSCs also appeared to have better therapeutic efficacy in animal experiments [135]. IPSCs can avoid many of the ethical dilemmas and the risk of immune rejection but might induce tumor generation. Beyond this, obtaining high-quality patient-specific iPSCs requires considerable time and expense, and the technical aspect is also a major challenge. Previous studies have reported that it takes at least 3 weeks from the start of induction to the establishment of stable iPSCs, and the induction efficiency is less than 1/5000 [136,137,138].
Mesenchymal stem cells
MSCs are derived from the mesoderm and can undergo self-renewal and exhibit multipotency, along with multidirectional differentiation potential. In 2006, the International Society for Cellular Therapy (ISCT) listed three minimum standards for MSCs as follows: (1) show adherence and growth; (2) must express specific surface markers, including CD73, CD90 and CD105, without the expression of hematopoietic antigens, such as CD14, CD34, CD45 or CD11b and Human Leukocyte Antigen-antigen D Related (HLA-DR); and (3) have the capacity to differentiate into adipocytes, osteoblasts and chondrocytes [139]. MSCs are present in all tissues of mesenchymal origin and exist primarily in bone marrow. Currently, the categories of MSCs that are widely used in research primarily include bone marrow mesenchymal stem cells (BM-MSCs), adipose-derived mesenchymal stem cells (AD-MSCs), human umbilical cord mesenchymal stem cells (hUC-MSCs), human placenta mesenchymal stem cells (hPMSCs), dental pulp mesenchyme stem cells (DPSCs) and so on [140]. The above MSCs exhibit self-renewal, multilineage differentiation potential, low immunogenicity [141], and immunomodulatory properties. These features are regulated by a cascade of transcription factors [142,143,144,145]. In addition, growth factors and cytokines as well as RNA take part in the regulation of MSCs [146,147,148]. MSCs mainly exert their therapeutic effects through the secretion of paracrine and autocrine factors for tissue repair, tissue regeneration, anti-inflammation, inflammation reduction, and anti-apoptosis [149, 150], among others.
MSCs are an effective method for treating lung diseases, including ALI, COPD and PF. In an inflammatory environment, MSCs can secrete different factors, such as interleukin-2, which enhances regulatory T cells [151] (or through paracrine signaling by releasing anti-inflammatory mediators such as IL-10, PGE2, and ang-1) and repairs ALI, and MSCs can relieve inflammation, reduce the number of goblet cells, regenerate lung tissue, ameliorate the loss of alveolar septa, and reduce the levels of NF-κB and p65 in the lung tissue of rats with COPD [152]. In addition, MSCs can inhibit the synthesis of TGF-β1 in AECs in pulmonary fibrosis by releasing Stanniocalcin-1 (STC1), reducing the immune function of macrophages, and repairing the incorrect epithelial-mesenchymal transition relationship [153].
Similarly, many studies have demonstrated the therapeutic potential of MSCs for PH. MSCs are widely used in preclinical studies for the treatment of PH. In both rat and mouse disease models, MSCs have been observed to alleviate and improve PH symptoms [154]. Although the mechanism is unclear, there are three aspects that can be further studied.
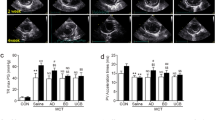
First, MSCs have immunoregulatory effects on PH. In the PH model induced by MCT, multiple related signaling pathways of inflammation showed abnormalities, such as prostaglandins, endothelin and the NO pathway. MSCs can restore the signaling pathways and protect against vascular remodeling induced by PH and then effectively reduce the thickness of the vessel wall and perivascular fibrosis [155]. The excessive proliferation of PASMCs associated with inflammation is one of the mechanisms of PH. MSCs significantly reduce the production of tumor necrosis factor-α (TNF-α), calcineurin (CaN) activity and nuclear factor of activated T cells (NFAT)-c2 activity, thereby inhibiting the excessive proliferation of PASMCs [156].
Second, MSCs can repair PMVECs. In lung injury studies, MSC-derived microvesicles (MVs) in coculture systems were found to improve lung injury by promoting Ang1 mRNA and protein secretion and restoring impaired PMVEC permeability to glucose [157]. Likewise, the experimental results in ALI and ARDS studies show that MSCs support paracellular permeability recovery in PMVECs induced by LPS injury and reduce PMVEC apoptosis [158]. In PMVECs, the STAT3 signaling pathway helps upregulate the expression of certain induced proinflammatory cytokines associated with ALI and ARDS [159]. UC-MSC-derived exosomes inhibit STAT3 signal transduction and upregulate miR-204 expression in PMVECs under anaerobic PH conditions; miR-204 is a critical microRNA with a reduced expression level in human PH [160]. In addition, UC-MSC-derived exosomes can significantly inhibit hypoxia-induced PMVEC apoptosis and PASMC proliferation, thereby suppressing pulmonary vascular remodeling and improving PH [161]. PMVECs support NO expression and can mediate autovasodilation. Endothelial nitric oxide synthase (eNOS) in PMVECs is essential to maintain normal endothelial cell function and homeostasis and plays a modulatory role in PH [162]. ADSCs can increase eNOS expression in PMVECs and enhance NO production in cocultures to improve PMVECs [163]. Binding of caveolin-1 (CaV1) with eNOS inactivates eNOS. Intravenous injection of MSCs carrying CaV1 mutants into the rat model of PH (1) significantly increases the NO level in the animal serum, (2) reduces the expression of α-SMA, and (3) reduces the thickness of pulmonary vascular tunica media [164]. Moreover, exhaustion of extracellular superoxide dismutase (ecSOD) in pulmonary MSCs was shown to accelerate pulmonary microvascular remodeling in PH, which is regulated by the typical Wnt/β-catenin pathway [165].
The third point is the effect on AECs. In the rat model of ALI, MSCs and ATII showed the same therapeutic effect [166]. Under air-liquid culture conditions, MSCs showed similar morphology and function to AECs, which proved that MSCs have the potential to differentiate in this direction [167]. This differentiation process is regulated by signaling pathways. Through a study on the treatment of ARDS, researchers found that when the Wnt/β-catenin, Wnt/JNK, or Wnt/PKC signaling pathways are activated, this differentiation ability will be promoted [168, 169]. In contrast, once the Wnt5a pathway is inhibited, such a differentiation process will be blocked [170]. In addition, the action of positive regulatory genes not only promotes the differentiation of MSCs into AECs but also increases the retention of MSCs in the lungs. In addition to differentiation, MSCs can also reduce the expression of HIF-1 and ROS proteins, inhibit the apoptosis of AECs and play a protective role by interfering with hypoxia stress signals [171].
Extracellular vesicles (EVs) may play a crucial role in some of the therapeutic effects of the above MSCs. EVs are nanosized particles released by cells that are present in most tissues and biological fluids, such as serum, cerebrospinal fluid, urine and cerebrospinal fluid [172], and can also be obtained in cell culture medium. EVs were initially considered as garbage bags of cellular waste, which were used to dispose and collect cellular waste. Later, EVs were proved to stably carry many important signaling molecules, which are involved in important physiological processes such as angiogenesis, cell migration and cell growth [173,174,175]. Importantly, EVs also function as messengers between cells, such as drug carriers that carry important cellular signals. Extracellular vesicles mainly consist of four subgroups: exosomes (30–150 nm), MVs (50–1000 nm), apoptotic bodies (500–2000 nm) and oncosomes (1–10 μm) [176]. At present, the research focus is the exosome subgroup. They can transfer substances including DNA, RNA or proteins between cells and thereby influence the function of the recipient cells. For the above therapeutic effects of MSCs, paracrine effects contribute significantly, and these cells are able to express, synthesize, and secrete various bioactive molecules such as various growth factors, cytokines, regulators, and signal peptides, as well as regulate active factors involved in metabolism, immunity, cell differentiation, proliferation, migration, nutrition, and apoptosis, through which a balance has been achieved for the homeostasis of the body, thus providing a suitable environment for stem cell immune regulation and anti-apoptosis [177,178,179]. Afterward, we organized the results on MSC-derived EVs in the treatment of PH into Table 1.
MSCs are adult stem cells with poor specificity, and their ability to differentiate into AECs or PMVECs needs further confirmation and exploration. Fundamentally treating PH by intervening in AECs or PMVECs through paracrine effects is an optimal strategy in the case of MSCs. Compared with ESCs and iPSCs, MSCs have weaker differentiation ability despite their advantages in terms of low immunogenicity. Exosomes are the main treatment tool for MSCs. In addition, for treating PH, the dose and frequency of MSCs remain to be clarified, without a completely reliable basis for clinical treatment, which has limited their application. A better strategy is to facilitate MSC exosome expression of certain specific proteins by gene editing to promote the treatment of PH.
The advantage of MSCs in clinical practice also lies in the homing phenomenon, i.e., targeting the damaged tissue [189]. However, this strategy is not a completely “sharpened sword”. This homing phenomenon is not accurate and efficient [190, 191]; instead, it is subject to many influencing factors [192], such as inflammation status [193], the number of ligands [194] and organismic age [195]. Studies have found that MSCs are virtually nontumorigenic and display a stable phenotype in long-term culture [196]; moreover, they pose a low risk of immune rejection [197, 198]. However, such immune escape is not absolute [199]. The poor specificity of MSCs and the side effects caused by the downregulation of immune function are also issues that need to be considered in clinical application. In addition, MSCs may accelerate the risk of pulmonary fibrosis [200, 201], which needs to be further confirmed. In a study performed at Hannover Medical School in Germany this year, a 3-year-old girl with severe PH due to hereditary hemorrhagic telangiectasia was injected by clinical researchers with 5 consecutive intravascular infusions of allogeneic MSCs over 6 months. The results showed significantly improved hemodynamic parameters and reduced markers of vascular injury and vascular inflammation, with no adverse side effects observed [202]. Although there are still many unknowns about the efficacy of mesenchymal stem cells in treating PH, this is undoubtedly exciting news.
Conclusions
ESCs possess the broadest pluripotency, but their application is limited by ethical concerns and immunological rejection issues. IPSCs, generated from a patient’s own cells, have the potential for unlimited proliferation and hold possibilities for gene therapy to address HPAH. They also largely avoid ethical and immunological rejection issues. However, there are risks of genomic alterations during cell amplification in vitro, and the four reprogramming factors are associated with tumorigenicity. Therefore, complex techniques are required to fully address these safety concerns before using iPSCs. MSCs, due to their limited differentiation ability, are currently studied more for their paracrine effects. The specific mechanisms involved in treating PH are not fully understood, and treatment outcomes may vary among different patients. Moreover, it is necessary to further ascertain whether long-term application of MSCs is required and if any side effects may arise. Hence, better regulation and selection methods are needed. While there are numerous challenges to be resolved in using stem cell therapy for the treatment of PH, the therapeutic potential it holds is a compelling reason for further research and exploration.
Currently, more insight is needed into the underlying mechanisms in the study of PH. Early signs of PH are difficult to diagnose, and further research into stem cell-based PH therapy should focus on whether it can prevent or reverse the progression of persistent PH symptoms. PMVECs and AECs are two good entries, and more high-quality clinical research evidence is needed.
Data availability
All data is available on request.
Abbreviations
- PH:
-
Pulmonary hypertension
- mPAP:
-
Mean pulmonary artery pressure
- RHC:
-
Right-sided heart catheterization
- PAWP:
-
Pulmonary arterial wedge pressure
- PVR:
-
Pulmonary vascular resistance
- NO:
-
Nitric oxide
- ALI:
-
Acute lung injury
- COPD:
-
Chronic obstructive pulmonary disease
- ARDS:
-
Acute respiratory distress syndrome
- PF:
-
Pulmonary fibrosis
- PMVECs:
-
Pulmonary microvascular endothelial cells
- AECs:
-
Alveolar epithelial cells
- LPS:
-
Lipopolysaccharide
- BPD:
-
Bronchopulmonary dysplasia
- MCT:
-
Monocrotaline
- TRPC:
-
Transient receptor potential cation channel
- PKC:
-
Protein kinase C
- MAPK:
-
Mitogen-activated protein kinase
- cAMP:
-
Cyclic adenosine monophosphate
- VEGF:
-
Vascular endothelial growth factors
- VSMCs:
-
vascular smooth muscle cells
- PGI2:
-
Prostaglandin-I-2
- ET-1:
-
Endothelin
- 5-HT:
-
5-hydroxytryptamine
- S1P:
-
Sphingosine-1-phosphate
- S1PR2:
-
Type II S1P receptor
- BRCA1:
-
Breast cancer 1
- GREM1:
-
Gremlin1
- ang-1:
-
Angiopoietin-1
- BMPR-II:
-
Type II bone morphogenetic protein receptor
- HIF:
-
Hypoxia-inducible factor
- smad:
-
Drosophila mothers against decapentaplegic protein
- ATI:
-
Alveolar type I
- ATII:
-
Alveolar type II
- TD:
-
Trans-differentiation
- nCLD:
-
Neonatal chronic lung disease
- PDGF-Rα:
-
Platelet-derived growth factor receptor α
- PGE2:
-
Prostaglandin E2
- SARS-CoV-2:
-
Severe acute respiratory syndrome corona virus 2
- VE-cadherin:
-
Vascular endothelial cadherin
- ROS:
-
Reactive oxygen species
- IL:
-
interleukin
- IFN:
-
Interferon
- TER:
-
Transendothelial electrical resistance
- ISM1:
-
Isthmin1
- PDE-5:
-
Phosphodiesterase-5
- sGC:
-
Soluble guanylate cyclase
- ERAs:
-
Endothelin receptor antagonists
- PPHN:
-
Persistent pulmonary hypertension in children
- cGMP:
-
Cyclic guanosine monophosphate
- PASMCs:
-
Pulmonary artery smooth muscle cells
- PTE:
-
Pulmonary thromboendarterectomy
- ESCs:
-
embryonic stem cells
- iPSc:
-
Induced pluripotent stem cells
- MSCs:
-
Mesenchymal stem cells
- EB:
-
Embryoid bodies
- SAGM:
-
Small airway growth medium
- SPA:
-
Surfactant-protein-A
- SPB:
-
Surfactant-protein-B
- SPC:
-
Surfactant-protein-C
- C3:
-
Complement 3
- C5:
-
Complement 5
- ETV2:
-
Ets variant 2
- HLA:
-
Human leukocyte antigen
- α-SMA:
-
α smooth muscle actin
- TGF-β:
-
Transforming growth factor-β
- CXCR:
-
Chemokine receptor
- OCT4:
-
Octamer transcription factor 4
- SOX2:
-
SRY-box containing gene 2
- KLF4:
-
Kruppel-like factor 4
- ISCT:
-
International Society for Cellular Therapy
- HLA-DR:
-
Human Leukocyte Antigen-antigen D Related
- TIMP-1:
-
Tissue inhibitors of metalloproteinase 1
- FAK:
-
Focal adhesion kinase
- iPSC-ECs:
-
IPSC-derived ECs
- iPSC-MSCs:
-
IPSC-derived MSCs
- BM-MSCs:
-
Bone marrow mesenchymal stem cells
- AD-MSCs:
-
Adipose-derived mesenchymal stem cells
- hUC-MSCs:
-
Human umbilical cord mesenchymal stem cells
- hPMSCs:
-
Human placenta mesenchymal stem cells
- DPSCs:
-
Dental pulp mesenchyme stem cells
- STC1:
-
Stanniocalcin-1
- TNF-α:
-
Tumor necrosis factor-α
- CaN:
-
Calcineurin
- NFAT:
-
Nuclear factor of activated T cells
- MVs:
-
Microvesicles
- eNOS:
-
Endothelial nitric oxide synthase
- CaV1:
-
Caveolin-1
- ecSOD:
-
Extracellular superoxide dismutase
- Su/Hx:
-
Sugen-5416/Hyposia
- RVSP:
-
Right Ventricular Systolic Pressure
- RV:
-
Right Ventricle
- LV + S:
-
Left Ventricle + Septum
- MCT:
-
Monocrotaline
- HYRX:
-
Hyporoxia
References
Voelkel NF, Gomez-Arroyo J, Abbate A, Bogaard HJ, Nicolls MR. Pathobiology of pulmonary arterial hypertension and right ventricular failure. Eur Respir J. 2012;40:1555–65. https://doi.org/10.1183/09031936.00046612.
Guignabert C, Dorfmuller P. Pathology and pathobiology of pulmonary hypertension. Semin Respir Crit Care Med. 2017;38:571–84. https://doi.org/10.1055/s-0037-1606214.
Villagra J, Shiva S, Hunter LA, Machado RF, Gladwin MT, Kato GJ. Platelet activation in patients with sickle disease, hemolysis-associated pulmonary hypertension, and nitric oxide scavenging by cell-free hemoglobin. Blood. 2007;110:2166–72. https://doi.org/10.1182/blood-2006-12-061697.
Al-Qadi M, LeVarge B, Ford HJ. Epidemiology, pathogenesis, and clinical approach in group 5 pulmonary hypertension. Front Med (Lausanne). 2020;7:616720. https://doi.org/10.3389/fmed.2020.616720.
Simonneau G, Montani D, Celermajer DS, et al. Haemodynamic definitions and updated clinical classification of pulmonary hypertension. Eur Respir J. 2019;53. https://doi.org/10.1183/13993003.01913-2018.
Long L, Ormiston ML, Yang X, et al. Selective enhancement of endothelial bmpr-ii with bmp9 reverses pulmonary arterial hypertension. Nat Med. 2015;21:777–85. https://doi.org/10.1038/nm.3877.
Liu X, Gao C, Wang Y, Niu L, Jiang S, Pan S. Bmsc-derived exosomes ameliorate lps-induced acute lung injury by mir-384-5p-controlled alveolar macrophage autophagy. Oxid Med Cell Longev. 2021; 2021: 9973457. https://doi.org/10.1155/2021/9973457.
Islam D, Huang Y, Fanelli V, et al. Identification and modulation of microenvironment is crucial for effective mesenchymal stromal cell therapy in acute lung injury. Am J Respir Crit Care Med. 2019;199:1214–24. https://doi.org/10.1164/rccm.201802-0356OC.
Morrison TJ, Jackson MV, Cunningham EK, et al. Mesenchymal stromal cells modulate macrophages in clinically relevant lung injury models by extracellular vesicle mitochondrial transfer. Am J Respir Crit Care Med. 2017;196:1275–86. https://doi.org/10.1164/rccm.201701-0170OC.
Prockop DJ, Oh JY. Mesenchymal stem/stromal cells (mscs): role as guardians of inflammation. Mol Ther. 2012;20:14–20. https://doi.org/10.1038/mt.2011.211.
Winiarski BK, Acheson N, Gutowski NJ, McHarg S, Whatmore JL. An improved and reliable method for isolation of microvascular endothelial cells from human omentum. Microcirculation. 2011;18:635–45. https://doi.org/10.1111/j.1549-8719.2011.00128.x.
Xu J, Xu D, Yu Z, et al. Exosomal mir-150 partially attenuated acute lung injury by mediating microvascular endothelial cells and mapk pathway. Biosci Rep. 2022;42. https://doi.org/10.1042/BSR20203363.
Glynos C, Kotanidou A, Orfanos SE, et al. Soluble guanylyl cyclase expression is reduced in lps-induced lung injury. Am J Physiol Regul Integr Comp Physiol. 2007;292:R1448–55. https://doi.org/10.1152/ajpregu.00341.2006.
Grinnell K, Duong H, Newton J, Rounds S, Choudhary G, Harrington EO. Heterogeneity in apoptotic responses of microvascular endothelial cells to oxidative stress. J Cell Physiol. 2012;227:1899–910. https://doi.org/10.1002/jcp.22918.
Kerchberger VE, Bastarache JA, Shaver CM, et al. Haptoglobin-2 variant increases susceptibility to acute respiratory distress syndrome during sepsis. Jci Insight. 2019;4. https://doi.org/10.1172/jci.insight.131206.
Beckmann N, Cannet C, Karmouty-Quintana H, et al. Lung mri for experimental drug research. Eur J Radiol. 2007;64:381–96. https://doi.org/10.1016/j.ejrad.2007.08.012.
Jin Y, Qian J, Ju X et al. Osthole protects against acute lung injury by suppressing nf-kappab-dependent inflammation. Mediators Inflamm. 2018; 2018: 4934592. https://doi.org/10.1155/2018/4934592.
Qi D, Tang X, He J, et al. Omentin protects against lps-induced ards through suppressing pulmonary inflammation and promoting endothelial barrier via an akt/enos-dependent mechanism. Cell Death Dis. 2016;7:e2360. https://doi.org/10.1038/cddis.2016.265.
Kolliputi N, Galam L, Parthasarathy PT, Tipparaju SM, Lockey RF. Nalp-3 inflammasome silencing attenuates ceramide-induced transepithelial permeability. J Cell Physiol. 2012;227:3310–6. https://doi.org/10.1002/jcp.24026.
Kuklin VN, Kirov MY, Evgenov OV, et al. Novel endothelin receptor antagonist attenuates endotoxin-induced lung injury in sheep. Crit Care Med. 2004;32:766–73. https://doi.org/10.1097/01.ccm.0000114575.08269.f6.
Yang Y, Hu S, Xu X et al. The vascular endothelial growth factors-expressing character of mesenchymal stem cells plays a positive role in treatment of acute lung injury in vivo. Mediators Inflamm. 2016; 2016: 2347938. https://doi.org/10.1155/2016/2347938.
Yi L, Huang X, Guo F, et al. Lipopolysaccharide induces human pulmonary micro-vascular endothelial apoptosis via the yap signaling pathway. Front Cell Infect Microbiol. 2016;6:133. https://doi.org/10.3389/fcimb.2016.00133.
Magro CM, Allen J, Pope-Harman A, et al. The role of microvascular injury in the evolution of idiopathic pulmonary fibrosis. Am J Clin Pathol. 2003;119:556–67. https://doi.org/10.1309/0B06-Y93E-GE6T-Q36Y.
Takabatake N, Arao T, Sata M, et al. Involvement of pulmonary endothelial cell injury in the pathogenesis of pulmonary fibrosis: clinical assessment by 123i-mibg lung scintigraphy. Eur J Nucl Med Mol Imaging. 2005;32:221–8. https://doi.org/10.1007/s00259-004-1663-1.
Patel RB, Kotha SR, Sherwani SI, et al. Pulmonary fibrosis inducer, bleomycin, causes redox-sensitive activation of phospholipase d and cytotoxicity through formation of bioactive lipid signal mediator, phosphatidic acid, in lung microvascular endothelial cells. Int J Toxicol. 2011;30:69–90. https://doi.org/10.1177/1091581810388850.
Wouters E, Franssen FM. Chronic obstructive pulmonary disease: shifting the paradigm to the vasculature. Am J Respir Crit Care Med. 2019;199:258–9. https://doi.org/10.1164/rccm.201808-1542ED.
Gong J, Zhao H, Liu T, et al. Cigarette smoke reduces fatty acid catabolism, leading to apoptosis in lung endothelial cells: implication for pathogenesis of copd. Front Pharmacol. 2019;10:941. https://doi.org/10.3389/fphar.2019.00941.
de Jesus PV, Yuan K, Lyuksyutova MA, et al. Whole-exome sequencing reveals topbp1 as a novel gene in idiopathic pulmonary arterial hypertension. Am J Respir Crit Care Med. 2014;189:1260–72. https://doi.org/10.1164/rccm.201310-1749OC.
Yang J, Nies MK, Fu Z, et al. Hepatoma-derived growth factor predicts disease severity and survival in pulmonary arterial hypertension. Am J Respir Crit Care Med. 2016;194:1264–72. https://doi.org/10.1164/rccm.201512-2498OC.
Bauer PM, Bauer EM, Rogers NM, et al. Activated cd47 promotes pulmonary arterial hypertension through targeting caveolin-1. Cardiovasc Res. 2012;93:682–93. https://doi.org/10.1093/cvr/cvr356.
Scott TE, Qin CX, Drummond GR, Hobbs AJ, Kemp-Harper BK. Innovative anti-inflammatory and pro-resolving strategies for pulmonary hypertension: high blood pressure research council of Australia award 2019. Hypertension. 2021;78:1168–84. https://doi.org/10.1161/HYPERTENSIONAHA.120.14525.
Majka S, Hagen M, Blackwell T, et al. Physiologic and molecular consequences of endothelial bmpr2 mutation. Respir Res. 2011;12:84. https://doi.org/10.1186/1465-9921-12-84.
Chen Y, Zhang D, Li Y, et al. Risk factors and outcomes of pulmonary hypertension in infants with bronchopulmonary dysplasia: a meta-analysis. Front Pediatr. 2021;9:695610. https://doi.org/10.3389/fped.2021.695610.
Meyrick BO, Reid LM. Crotalaria-induced pulmonary hypertension. Uptake of 3 h-thymidine by the cells of the pulmonary circulation and alveolar walls. Am J Pathol. 1982;106:84–94.
Wang L, Luo H, Qin G, et al. The impact of sevoflurane on coupling of the left ventricular-to-systemic vasculature in rats with chronic pulmonary hypertension. J Cardiothorac Vasc Anesth. 2017;31:2027–34. https://doi.org/10.1053/j.jvca.2017.02.049.
Taraseviciene-Stewart L, Kasahara Y, Alger L, et al. Inhibition of the vegf receptor 2 combined with chronic hypoxia causes cell death-dependent pulmonary endothelial cell proliferation and severe pulmonary hypertension. Faseb J. 2001;15:427–38. https://doi.org/10.1096/fj.00-0343com.
Qi B, Chen HL, Shang D, Dong Y, Zhang GX, Yu L. Effects of hypoxia-inducible factor-1alpha and matrix metalloproteinase-9 on alveolar-capillary barrier disruption and lung edema in rat models of severe acute pancreatitis-associated lung injury. Exp Ther Med. 2014;8:899–906. https://doi.org/10.3892/etm.2014.1810.
Wang H, Chen Y, Li W, et al. Ligustrazine effect on lipopolysaccharide-induced pulmonary damage in rats. Burns. 2015;41:1235–41. https://doi.org/10.1016/j.burns.2015.02.010.
Yang L, Liu S, Han S, et al. The hdl from septic-ards patients with composition changes exacerbates pulmonary endothelial dysfunction and acute lung injury induced by cecal ligation and puncture (clp) in mice. Respir Res. 2020;21:293. https://doi.org/10.1186/s12931-020-01553-3.
Chen M, Chen C, Yuan X, et al. Angiotensin ii aggravates lipopolysaccharide induced human pulmonary microvascular endothelial cells permeability in high glucose status. Endocr J. 2018;65:717–25. https://doi.org/10.1507/endocrj.EJ17-0477.
Perkett EA, Klekamp JG. Vascular endothelial growth factor expression is decreased in rat lung following exposure to 24 or 48 hours of hyperoxia: implications for endothelial cell survival. Chest. 1998;114:S52–3. https://doi.org/10.1378/chest.114.1_supplement.52s.
Abadie Y, Bregeon F, Papazian L, et al. Decreased vegf concentration in lung tissue and vascular injury during ards. Eur Respir J. 2005;25:139–46. https://doi.org/10.1183/09031936.04.00065504.
Mura M, Dos SC, Stewart D, Liu M. Vascular endothelial growth factor and related molecules in acute lung injury. J Appl Physiol (1985) 2004; 97: 1605-17. https://doi.org/10.1152/japplphysiol.00202.2004.
Iwatani S, Mizobuchi M, Tanaka S, et al. Increased volume of tracheal aspirate fluid predicts the development of bronchopulmonary dysplasia. Early Hum Dev. 2013;89:113–7. https://doi.org/10.1016/j.earlhumdev.2012.08.007.
Hao Y, Wang Z, Frimpong F, Chen X. Calcium-permeable channels and endothelial dysfunction in acute lung injury. Curr Issues Mol Biol. 2022;44:2217–29. https://doi.org/10.3390/cimb44050150.
Herold S, Gabrielli NM, Vadasz I. Novel concepts of acute lung injury and alveolar-capillary barrier dysfunction. Am J Physiol Lung Cell Mol Physiol. 2013;305:L665–81. https://doi.org/10.1152/ajplung.00232.2013.
Yu Y, Sweeney M, Zhang S, et al. Pdgf stimulates pulmonary vascular smooth muscle cell proliferation by upregulating trpc6 expression. Am J Physiol Cell Physiol. 2003;284:C316–30. https://doi.org/10.1152/ajpcell.00125.2002.
Schermuly RT, Dony E, Ghofrani HA, et al. Reversal of experimental pulmonary hypertension by pdgf inhibition. J Clin Invest. 2005;115:2811–21. https://doi.org/10.1172/JCI24838.
Guignabert C, Alvira CM, Alastalo TP, et al. Tie2-mediated loss of peroxisome proliferator-activated receptor-gamma in mice causes pdgf receptor-beta-dependent pulmonary arterial muscularization. Am J Physiol Lung Cell Mol Physiol. 2009;297:L1082–90. https://doi.org/10.1152/ajplung.00199.2009.
Ghofrani HA, Humbert M, Langleben D, et al. Riociguat: mode of action and clinical development in pulmonary hypertension. Chest. 2017;151:468–80. https://doi.org/10.1016/j.chest.2016.05.024.
Cua CL, Rogers LK, Chicoine LG, et al. Down syndrome patients with pulmonary hypertension have elevated plasma levels of asymmetric dimethylarginine. Eur J Pediatr. 2011;170:859–63. https://doi.org/10.1007/s00431-010-1361-x.
Tian S, Cai Z, Sen P, et al. Loss of lung microvascular endothelial piezo2 expression impairs no synthesis, induces endmt, and is associated with pulmonary hypertension. Am J Physiol Heart Circ Physiol. 2022;323:H958–74. https://doi.org/10.1152/ajpheart.00220.2022.
Yan F, Su L, Chen X, Wang X, Gao H, Zeng Y. Molecular regulation and clinical significance of caveolin-1 methylation in chronic lung diseases. Clin Transl Med. 2020;10:151–60. https://doi.org/10.1002/ctm2.2.
Wang N, Zhang D, Sun G, et al. Lipopolysaccharide-induced caveolin-1 phosphorylation-dependent increase in transcellular permeability precedes the increase in paracellular permeability. Drug Des Devel Ther. 2015;9:4965–77. https://doi.org/10.2147/DDDT.S77646.
Murata T, Lin MI, Huang Y, et al. Reexpression of caveolin-1 in endothelium rescues the vascular, cardiac, and pulmonary defects in global caveolin-1 knockout mice. J Exp Med. 2007;204:2373–82. https://doi.org/10.1084/jem.20062340.
Huang J, Frid M, Gewitz MH, et al. Hypoxia-induced pulmonary hypertension and chronic lung disease: caveolin-1 dysfunction an important underlying feature. Pulm Circ. 2019;9:2045894019837876. https://doi.org/10.1177/2045894019837876.
Oliveira S, Chen J, Castellon M, et al. Injury-induced shedding of extracellular vesicles depletes endothelial cells of cav-1 (caveolin-1) and enables tgf-beta (transforming growth factor-beta)-dependent pulmonary arterial hypertension. Arterioscler Thromb Vasc Biol. 2019;39:1191–202. https://doi.org/10.1161/ATVBAHA.118.312038.
Oliveira S, Castellon M, Chen J, et al. Inflammation-induced caveolin-1 and bmprii depletion promotes endothelial dysfunction and tgf-beta-driven pulmonary vascular remodeling. Am J Physiol Lung Cell Mol Physiol. 2017;312:L760–71. https://doi.org/10.1152/ajplung.00484.2016.
Teichert-Kuliszewska K, Kutryk MJ, Kuliszewski MA, et al. Bone morphogenetic protein receptor-2 signaling promotes pulmonary arterial endothelial cell survival: implications for loss-of-function mutations in the pathogenesis of pulmonary hypertension. Circ Res. 2006;98:209–17. https://doi.org/10.1161/01.RES.0000200180.01710.e6.
Archer SL, Weir EK, Wilkins MR. Basic science of pulmonary arterial hypertension for clinicians: new concepts and experimental therapies. Circulation. 2010;121:2045–66. https://doi.org/10.1161/CIRCULATIONAHA.108.847707.
Du H, Li X, Zhao W, Jiang N. The difference between sacubitril valsartan and valsartan on vascular endothelial function, apn, mmp-9, and bnp levels in patients with hypertension and chronic heart failure. J Healthc Eng. 2022; 2022: 9494981. https://doi.org/10.1155/2022/9494981.
Laporte S, Denault JB, D’Orleans-Juste P, Leduc R. Presence of furin mrna in cultured bovine endothelial cells and possible involvement of furin in the processing of the endothelin precursor. J Cardiovasc Pharmacol. 1993;22(Suppl 8):S7–10. https://doi.org/10.1097/00005344-199322008-00004.
Bohm F, Pernow J. The importance of endothelin-1 for vascular dysfunction in cardiovascular disease. Cardiovasc Res. 2007;76:8–18. https://doi.org/10.1016/j.cardiores.2007.06.004.
Ayoub T. Pulmonary hypertension in liver transplant. Curr Opin Organ Transpl. 2011;16:331–7. https://doi.org/10.1097/MOT.0b013e328346e138.
Vanhoutte PM. Say no to et. J Auton Nerv Syst. 2000;81:271–7. https://doi.org/10.1016/s0165-1838(00)00126-0.
Schneider MP, Boesen EI, Pollock DM. Contrasting actions of Endothelin et(a) and et(b) receptors in cardiovascular disease. Annu Rev Pharmacol Toxicol. 2007;47:731–59. https://doi.org/10.1146/annurev.pharmtox.47.120505.105134.
Porta NF, Steinhorn RH. Pulmonary vasodilator therapy in the Nicu: inhaled nitric oxide, sildenafil, and other pulmonary vasodilating agents. Clin Perinatol. 2012;39:149–64. https://doi.org/10.1016/j.clp.2011.12.006.
Lin CC, Lin WN, Hou WC, Hsiao LD, Yang CM. Endothelin-1 induces vcam-1 expression-mediated inflammation via receptor tyrosine kinases and elk/p300 in human tracheal smooth muscle cells. Am J Physiol Lung Cell Mol Physiol. 2015;309:L211–25. https://doi.org/10.1152/ajplung.00232.2014.
Langleben D, Dupuis J, Langleben I, et al. Etiology-specific endothelin-1 clearance in human precapillary pulmonary hypertension. Chest. 2006;129:689–95. https://doi.org/10.1378/chest.129.3.689.
Giaid A, Yanagisawa M, Langleben D, et al. Expression of endothelin-1 in the lungs of patients with pulmonary hypertension. N Engl J Med. 1993;328:1732–9. https://doi.org/10.1056/NEJM199306173282402.
Star GP, Giovinazzo M, Langleben D. Effects of bone morphogenic proteins and transforming growth factor-beta on in-vitro production of endothelin-1 by human pulmonary microvascular endothelial cells. Vascul Pharmacol. 2009;50:45–50. https://doi.org/10.1016/j.vph.2008.09.001.
Star GP, Giovinazzo M, Langleben D. Bone morphogenic protein-9 stimulates endothelin-1 release from human pulmonary microvascular endothelial cells: a potential mechanism for elevated et-1 levels in pulmonary arterial hypertension. Microvasc Res. 2010;80:349–54. https://doi.org/10.1016/j.mvr.2010.05.010.
Newman JH, Phillips JR, Loyd JE. Narrative review: the enigma of pulmonary arterial hypertension: new insights from genetic studies. Ann Intern Med. 2008;148:278–83. https://doi.org/10.7326/0003-4819-148-4-200802190-00006.
Heuer JF, Sauter P, Pelosi P, et al. Effects of pulmonary acid aspiration on the lungs and extra-pulmonary organs: a randomized study in pigs. Crit Care. 2012;16:R35. https://doi.org/10.1186/cc11214.
Yu Z, Jin J, Wang Y, Sun J. High density lipoprotein promoting proliferation and migration of type ii alveolar epithelial cells during inflammation state. Lipids Health Dis. 2017;16:91. https://doi.org/10.1186/s12944-017-0482-x.
Lu R, Popov V, Patel J, Eaves-Pyles T. Burkholderia mallei and burkholderia pseudomallei stimulate differential inflammatory responses from human alveolar type ii cells (atii) and macrophages. Front Cell Infect Microbiol. 2012;2:165. https://doi.org/10.3389/fcimb.2012.00165.
Branchfield K, Li R, Lungova V, Verheyden JM, McCulley D, Sun X. A three-dimensional study of alveologenesis in mouse lung. Dev Biol. 2016;409:429–41. https://doi.org/10.1016/j.ydbio.2015.11.017.
Willems CH, Zimmermann LJ, Sanders PJ, et al. Alveolocapillary model system to study alveolar re-epithelialization. Exp Cell Res. 2013;319:64–74. https://doi.org/10.1016/j.yexcr.2012.09.010.
Wang L, Taneja R, Wang W, et al. Human alveolar epithelial cells attenuate pulmonary microvascular endothelial cell permeability under septic conditions. PLoS ONE. 2013;8:e55311. https://doi.org/10.1371/journal.pone.0055311.
Barnthaler T, Maric J, Platzer W, et al. The role of pge2 in alveolar epithelial and lung microvascular endothelial crosstalk. Sci Rep. 2017;7:7923. https://doi.org/10.1038/s41598-017-08228-y.
Chen Q, Rehman J, Chan M, et al. Angiocrine sphingosine-1-phosphate activation of s1pr2-yap signaling axis in alveolar type ii cells is essential for lung repair. Cell Rep. 2020;31:107828. https://doi.org/10.1016/j.celrep.2020.107828.
Wang P, Luo R, Zhang M, et al. A cross-talk between epithelium and endothelium mediates human alveolar-capillary injury during SARS-cov-2 infection. Cell Death Dis. 2020;11:1042. https://doi.org/10.1038/s41419-020-03252-9.
Li J, Xia Y, Huang Z, et al. Novel hif-1-target gene isthmin1 contributes to hypoxia-induced hyperpermeability of pulmonary microvascular endothelial cells monolayers. Am J Physiol Cell Physiol. 2021;321:C671–80. https://doi.org/10.1152/ajpcell.00124.2021.
Farha S, Laskowski D, George D, et al. Loss of alveolar membrane diffusing capacity and pulmonary capillary blood volume in pulmonary arterial hypertension. Respir Res. 2013;14:6. https://doi.org/10.1186/1465-9921-14-6.
Pepke-Zaba J, Delcroix M, Lang I, et al. Chronic thromboembolic pulmonary hypertension (cteph): results from an international prospective registry. Circulation. 2011;124:1973–81. https://doi.org/10.1161/CIRCULATIONAHA.110.015008.
Ogo T, Fukuda T, Tsuji A, et al. Efficacy and safety of balloon pulmonary angioplasty for chronic thromboembolic pulmonary hypertension guided by cone-beam computed tomography and electrocardiogram-gated area detector computed tomography. Eur J Radiol. 2017;89:270–6. https://doi.org/10.1016/j.ejrad.2016.12.013.
Evans MJ, Kaufman MH. Establishment in culture of pluripotential cells from mouse embryos. Nature. 1981;292:154–6. https://doi.org/10.1038/292154a0.
Fuchs S, Hollins AJ, Laue M, et al. Differentiation of human alveolar epithelial cells in primary culture: morphological characterization and synthesis of caveolin-1 and surfactant protein-c. Cell Tissue Res. 2003;311:31–45. https://doi.org/10.1007/s00441-002-0653-5.
Mason RJ, Lewis MC, Edeen KE, McCormick-Shannon K, Nielsen LD, Shannon JM. Maintenance of surfactant protein a and d secretion by rat alveolar type ii cells in vitro. Am J Physiol Lung Cell Mol Physiol. 2002;282:L249–58. https://doi.org/10.1152/ajplung.00027.2001.
Ali NN, Edgar AJ, Samadikuchaksaraei A, et al. Derivation of type ii alveolar epithelial cells from murine embryonic stem cells. Tissue Eng. 2002;8:541–50. https://doi.org/10.1089/107632702760240463.
Rippon HJ, Ali NN, Polak JM, Bishop AE. Initial observations on the effect of medium composition on the differentiation of murine embryonic stem cells to alveolar type ii cells. Cloning Stem Cells. 2004;6:49–56. https://doi.org/10.1089/1536230041372328.
Rippon HJ, Polak JM, Qin M, Bishop AE. Derivation of distal lung epithelial progenitors from murine embryonic stem cells using a novel three-step differentiation protocol. Stem Cells. 2006;24:1389–98. https://doi.org/10.1634/stemcells.2005-0465.
Wang D, Haviland DL, Burns AR, Zsigmond E, Wetsel RA. A pure population of lung alveolar epithelial type ii cells derived from human embryonic stem cells. Proc Natl Acad Sci U S A. 2007;104:4449–54. https://doi.org/10.1073/pnas.0700052104.
Wang D, Morales JE, Calame DG, Alcorn JL, Wetsel RA. Transplantation of human embryonic stem cell-derived alveolar epithelial type ii cells abrogates acute lung injury in mice. Mol Ther. 2010;18:625–34. https://doi.org/10.1038/mt.2009.317.
Odorico JS, Kaufman DS, Thomson JA. Multilineage differentiation from human embryonic stem cell lines. Stem Cells. 2001;19:193–204. https://doi.org/10.1634/stemcells.19-3-193.
Watt FM, Hogan BL. Out of eden: stem cells and their niches. Science. 2000;287:1427–30. https://doi.org/10.1126/science.287.5457.1427.
Van Vranken BE, Romanska HM, Polak JM, Rippon HJ, Shannon JM, Bishop AE. Coculture of embryonic stem cells with pulmonary mesenchyme: a microenvironment that promotes differentiation of pulmonary epithelium. Tissue Eng. 2005;11:1177–87. https://doi.org/10.1089/ten.2005.11.1177.
Lo PK, Lee JS, Liang X, et al. Epigenetic inactivation of the potential tumor suppressor gene foxf1 in breast cancer. Cancer Res. 2010;70:6047–58. https://doi.org/10.1158/0008-5472.CAN-10-1576.
Ren X, Ustiyan V, Guo M, et al. Postnatal alveologenesis depends on foxf1 signaling in c-kit(+) endothelial progenitor cells. Am J Respir Crit Care Med. 2019;200:1164–76. https://doi.org/10.1164/rccm.201812-2312OC.
Kolesnichenko OA, Flood HM, Zhang Y, et al. Endothelial progenitor cells derived from embryonic stem cells prevent alveolar simplification in a murine model of bronchopulmonary dysplasia. Front Cell Dev Biol. 2023;11:1209518. https://doi.org/10.3389/fcell.2023.1209518.
Liu J, Wang W, Wang L, et al. Il-33 initiates vascular remodelling in hypoxic pulmonary hypertension by up-regulating hif-1alpha and vegf expression in vascular endothelial cells. Ebiomedicine. 2018;33:196–210. https://doi.org/10.1016/j.ebiom.2018.06.003.
Tsang KM, Hyun JS, Cheng KT, et al. Embryonic stem cell differentiation to functional arterial endothelial cells through sequential activation of etv2 and notch1 signaling by hif1alpha. Stem Cell Rep. 2017;9:796–806. https://doi.org/10.1016/j.stemcr.2017.07.001.
Bulic-Jakus F, Ulamec M, Vlahovic M, et al. Of mice and men: teratomas and teratocarcinomas. Coll Antropol. 2006;30:921–4.
Tan M, Zhao Y, Kim SJ, et al. Sag/rbx2/roc2 e3 ubiquitin ligase is essential for vascular and neural development by targeting nf1 for degradation. Dev Cell. 2011;21:1062–76. https://doi.org/10.1016/j.devcel.2011.09.014.
Ellis P, Fagan BM, Magness ST, et al. Sox2, a persistent marker for multipotential neural stem cells derived from embryonic stem cells, the embryo or the adult. Dev Neurosci. 2004;26:148–65. https://doi.org/10.1159/000082134.
Liu XF, Yang WT, Xu R, Liu JT, Zheng PS. Cervical cancer cells with positive sox2 expression exhibit the properties of cancer stem cells. PLoS ONE. 2014;9:e87092. https://doi.org/10.1371/journal.pone.0087092.
Amariglio N, Hirshberg A, Scheithauer BW, et al. Donor-derived brain tumor following neural stem cell transplantation in an ataxia telangiectasia patient. Plos Med. 2009;6:e1000029. https://doi.org/10.1371/journal.pmed.1000029.
Herberts CA, Kwa MS, Hermsen HP. Risk factors in the development of stem cell therapy. J Transl Med. 2011;9:29. https://doi.org/10.1186/1479-5876-9-29.
Nussbaum J, Minami E, Laflamme MA, et al. Transplantation of undifferentiated murine embryonic stem cells in the heart: teratoma formation and immune response. Faseb J. 2007;21:1345–57. https://doi.org/10.1096/fj.06-6769com.
Robertson JA. Human embryonic stem cell research: ethical and legal issues. Nat Rev Genet. 2001;2:74–8. https://doi.org/10.1038/35047594.
Campbell A. Ethos and economics: examining the rationale underlying stem cell and cloning research policies in the United States, Germany, and Japan. Am J Law Med. 2005;31:47–86. https://doi.org/10.1177/009885880503100102.
Gottweis H. The endless hesc controversy in the United States: history, context, and prospects. Cell Stem Cell. 2010;7:555–8. https://doi.org/10.1016/j.stem.2010.10.008.
Winston R. Does government regulation inhibit embryonic stem cell research and can it be effective? Cell Stem Cell. 2007;1:27–34. https://doi.org/10.1016/j.stem.2007.05.006.
Peng YJ, Huang X, Zhou Q. Ethical and policy considerations for human embryo and stem cell research in China. Cell Stem Cell. 2020;27:511–4. https://doi.org/10.1016/j.stem.2020.09.010.
Alvarez-Palomo B, Sanchez-Lopez LI, Moodley Y, Edel MJ, Serrano-Mollar A. Induced pluripotent stem cell-derived lung alveolar epithelial type ii cells reduce damage in bleomycin-induced lung fibrosis. Stem Cell Res Ther. 2020;11:213. https://doi.org/10.1186/s13287-020-01726-3.
Xing D, Wells JM, Giordano SS, et al. Induced pluripotent stem cell-derived endothelial cells attenuate lipopolysaccharide-induced acute lung injury. J Appl Physiol (1985). 2019;127:444–56. https://doi.org/10.1152/japplphysiol.00587.2018.
Su VY, Chiou SH, Lin CS, Mo MH, Yang KY. Induced pluripotent stem cells attenuate endothelial leakage in acute lung injury via tissue inhibitor of metalloproteinases-1 to reduce focal adhesion kinase activity. Stem Cells. 2019;37:1516–27. https://doi.org/10.1002/stem.3093.
Nakayama KH, Joshi PA, Lai ES, et al. Bilayered vascular graft derived from human induced pluripotent stem cells with biomimetic structure and function. Regen Med. 2015;10:745–55. https://doi.org/10.2217/rme.15.45.
Huang SX, Islam MN, O’Neill J, et al. Efficient generation of lung and airway epithelial cells from human pluripotent stem cells. Nat Biotechnol. 2014;32:84–91. https://doi.org/10.1038/nbt.2754.
Snoeck HW. Modeling human lung development and disease using pluripotent stem cells. Development. 2015;142:13–6. https://doi.org/10.1242/dev.115469.
Huang WC, Ke MW, Cheng CC, et al. Therapeutic benefits of induced pluripotent stem cells in monocrotaline-induced pulmonary arterial hypertension. PLoS ONE. 2016;11:e0142476. https://doi.org/10.1371/journal.pone.0142476.
Ren X, Moser PT, Gilpin SE, et al. Engineering pulmonary vasculature in decellularized rat and human lungs. Nat Biotechnol. 2015;33:1097–102. https://doi.org/10.1038/nbt.3354.
Shafa M, Ionescu LI, Vadivel A, et al. Human induced pluripotent stem cell-derived lung progenitor and alveolar epithelial cells attenuate hyperoxia-induced lung injury. Cytotherapy. 2018;20:108–25. https://doi.org/10.1016/j.jcyt.2017.09.003.
Tao F, Wang F, Zhang W, Hao Y. Microrna-22 enhances the differentiation of mouse induced pluripotent stem cells into alveolar epithelial type ii cells. Eur J Histochem. 2020;64. https://doi.org/10.4081/ejh.2020.3170.
Lee AS, Tang C, Rao MS, Weissman IL, Wu JC. Tumorigenicity as a clinical hurdle for pluripotent stem cell therapies. Nat Med. 2013;19:998–1004. https://doi.org/10.1038/nm.3267.
Gutierrez-Aranda I, Ramos-Mejia V, Bueno C, et al. Human induced pluripotent stem cells develop teratoma more efficiently and faster than human embryonic stem cells regardless the site of injection. Stem Cells. 2010;28:1568–70. https://doi.org/10.1002/stem.471.
Yan Q, Quan Y, Sun H, et al. A site-specific genetic modification for induction of pluripotency and subsequent isolation of derived lung alveolar epithelial type ii cells. Stem Cells. 2014;32:402–13. https://doi.org/10.1002/stem.1570.
Sa S, Gu M, Chappell J, et al. Induced pluripotent stem cell model of pulmonary arterial hypertension reveals novel gene expression and patient specificity. Am J Respir Crit Care Med. 2017;195:930–41. https://doi.org/10.1164/rccm.201606-1200OC.
Gu M, Donato M, Guo M, et al. Ipsc-endothelial cell phenotypic drug screening and in silico analyses identify tyrphostin-ag1296 for pulmonary arterial hypertension. Sci Transl Med. 2021;13. https://doi.org/10.1126/scitranslmed.aba6480.
Rabinovitch M. Combining induced pluripotent stem cell with next generation sequencing technology to gain new insights into pathobiology and treatment of pulmonary arterial hypertension. Pulm Circ. 2013;3:153–5. https://doi.org/10.4103/2045-8932.109963.
Xue X, Zhang S, Jiang W, et al. Protective effect of baicalin against pulmonary arterial hypertension vascular remodeling through regulation of tnf-alpha signaling pathway. Pharmacol Res Perspect. 2021;9:e00703. https://doi.org/10.1002/prp2.703.
Sauler M, Bazan IS, Lee PJ. Cell death in the lung: the apoptosis-necroptosis axis. Annu Rev Physiol. 2019;81:375–402. https://doi.org/10.1146/annurev-physiol-020518-114320.
Gu M, Shao NY, Sa S, et al. Patient-specific ipsc-derived endothelial cells uncover pathways that protect against pulmonary hypertension in bmpr2 mutation carriers. Cell Stem Cell. 2017;20:490–e5045. https://doi.org/10.1016/j.stem.2016.08.019.
Lian Q, Zhang Y, Zhang J, et al. Functional mesenchymal stem cells derived from human induced pluripotent stem cells attenuate limb ischemia in mice. Circulation. 2010;121:1113–23. https://doi.org/10.1161/CIRCULATIONAHA.109.898312.
Sun YQ, Deng MX, He J, et al. Human pluripotent stem cell-derived mesenchymal stem cells prevent allergic airway inflammation in mice. Stem Cells. 2012;30:2692–9. https://doi.org/10.1002/stem.1241.
Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006;126:663–76. https://doi.org/10.1016/j.cell.2006.07.024.
Syed KM, Joseph S, Mukherjee A, et al. Histone chaperone aplf regulates induction of pluripotency in murine fibroblasts. J Cell Sci. 2016;129:4576–91. https://doi.org/10.1242/jcs.194035.
Park IH, Lerou PH, Zhao R, Huo H, Daley GQ. Generation of human-induced pluripotent stem cells. Nat Protoc. 2008;3:1180–6. https://doi.org/10.1038/nprot.2008.92.
Dominici M, Le Blanc K, Mueller I, et al. Minimal criteria for defining multipotent mesenchymal stromal cells. The international society for cellular therapy position statement. Cytotherapy. 2006;8:315–7. https://doi.org/10.1080/14653240600855905.
Aurrekoetxea M, Garcia-Gallastegui P, Irastorza I, et al. Dental pulp stem cells as a multifaceted tool for bioengineering and the regeneration of craniomaxillofacial tissues. Front Physiol. 2015;6:289. https://doi.org/10.3389/fphys.2015.00289.
Crop M, Baan C, Weimar W, Hoogduijn M. Potential of mesenchymal stem cells as immune therapy in solid-organ transplantation. Transpl Int. 2009;22:365–76. https://doi.org/10.1111/j.1432-2277.2008.00786.x.
Quintana AM, Zhou YE, Pena JJ, O’Rourke JP, Ness SA. Dramatic repositioning of c-myb to different promoters during the cell cycle observed by combining cell sorting with chromatin immunoprecipitation. PLoS ONE. 2011;6:e17362. https://doi.org/10.1371/journal.pone.0017362.
Loh YH, Wu Q, Chew JL, et al. The oct4 and nanog transcription network regulates pluripotency in mouse embryonic stem cells. Nat Genet. 2006;38:431–40. https://doi.org/10.1038/ng1760.
Solana J, Irimia M, Ayoub S, et al. Conserved functional antagonism of celf and mbnl proteins controls stem cell-specific alternative splicing in planarians. Elife. 2016;5. https://doi.org/10.7554/eLife.16797.
Yoshizaki K, Hu L, Nguyen T, et al. Ablation of coactivator med1 switches the cell fate of dental epithelia to that generating hair. PLoS ONE. 2014;9:e99991. https://doi.org/10.1371/journal.pone.0099991.
Lin Y, Fang ZP, Liu HJ, et al. Hgf/r-spondin1 rescues liver dysfunction through the induction of lgr5(+) liver stem cells. Nat Commun. 2017;8:1175. https://doi.org/10.1038/s41467-017-01341-6.
Abomaray FM, Al JM, Alsaad KO et al. Phenotypic and functional characterization of mesenchymal stem/multipotent stromal cells from decidua basalis of human term placenta. Stem Cells Int. 2016; 2016: 5184601. https://doi.org/10.1155/2016/5184601.
Li CX, Talele NP, Boo S, et al. Microrna-21 preserves the fibrotic mechanical memory of mesenchymal stem cells. Nat Mater. 2017;16:379–89. https://doi.org/10.1038/nmat4780.
Elshaer SL, Evans W, Pentecost M, et al. Adipose stem cells and their paracrine factors are therapeutic for early retinal complications of diabetes in the ins2(akita) mouse. Stem Cell Res Ther. 2018;9:322. https://doi.org/10.1186/s13287-018-1059-y.
Mazo M, Cemborain A, Gavira JJ, et al. Adipose stromal vascular fraction improves cardiac function in chronic myocardial infarction through differentiation and paracrine activity. Cell Transpl. 2012;21:1023–37. https://doi.org/10.3727/096368911X623862.
Eggenhofer E, Luk F, Dahlke MH, Hoogduijn MJ. The life and fate of mesenchymal stem cells. Front Immunol. 2014;5:148. https://doi.org/10.3389/fimmu.2014.00148.
Schweitzer KS, Johnstone BH, Garrison J, et al. Adipose stem cell treatment in mice attenuates lung and systemic injury induced by cigarette smoking. Am J Respir Crit Care Med. 2011;183:215–25. https://doi.org/10.1164/rccm.201001-0126OC.
Ono M, Ohkouchi S, Kanehira M, et al. Mesenchymal stem cells correct inappropriate epithelial-mesenchyme relation in pulmonary fibrosis using stanniocalcin-1. Mol Ther. 2015;23:549–60. https://doi.org/10.1038/mt.2014.217.
Aliotta JM, Pereira M, Wen S, et al. Exosomes induce and reverse monocrotaline-induced pulmonary hypertension in mice. Cardiovasc Res. 2016;110:319–30. https://doi.org/10.1093/cvr/cvw054.
Oh S, Jang AY, Chae S, et al. Comparative analysis on the anti-inflammatory/immune effect of mesenchymal stem cell therapy for the treatment of pulmonary arterial hypertension. Sci Rep. 2021;11:2012. https://doi.org/10.1038/s41598-021-81244-1.
Liu J, Han Z, Han Z, He Z. Mesenchymal stem cell-conditioned media suppresses inflammation-associated overproliferation of pulmonary artery smooth muscle cells in a rat model of pulmonary hypertension. Exp Ther Med. 2016;11:467–75. https://doi.org/10.3892/etm.2015.2953.
Hu S, Park J, Liu A, et al. Mesenchymal stem cell microvesicles restore protein permeability across primary cultures of injured human lung microvascular endothelial cells. Stem Cells Transl Med. 2018;7:615–24. https://doi.org/10.1002/sctm.17-0278.
Li JZ, Meng SS, Xu XP, et al. Mechanically stretched mesenchymal stem cells can reduce the effects of lps-induced injury on the pulmonary microvascular endothelium barrier. Stem Cells Int. 2020;2020:8861407. https://doi.org/10.1155/2020/8861407.
Wolfson RK, Mapes B, Garcia J. Excessive mechanical stress increases hmgb1 expression in human lung microvascular endothelial cells via stat3. Microvasc Res. 2014;92:50–5. https://doi.org/10.1016/j.mvr.2013.12.005.
Lee C, Mitsialis SA, Aslam M, et al. Exosomes mediate the cytoprotective action of mesenchymal stromal cells on hypoxia-induced pulmonary hypertension. Circulation. 2012;126:2601–11. https://doi.org/10.1161/CIRCULATIONAHA.112.114173.
Zhang S, Liu X, Ge LL, et al. Mesenchymal stromal cell-derived exosomes improve pulmonary hypertension through inhibition of pulmonary vascular remodeling. Respir Res. 2020;21:71. https://doi.org/10.1186/s12931-020-1331-4.
Maron BA, Zhang YY, White K, et al. Aldosterone inactivates the endothelin-b receptor via a cysteinyl thiol redox switch to decrease pulmonary endothelial nitric oxide levels and modulate pulmonary arterial hypertension. Circulation. 2012;126:963–74. https://doi.org/10.1161/CIRCULATIONAHA.112.094722.
Gao P, Yang X, Mungur L, Kampo S, Wen Q. Adipose tissue-derived stem cells attenuate acute lung injury through enos and enos-derived no. Int J Mol Med. 2013;31:1313–8. https://doi.org/10.3892/ijmm.2013.1328.
Chen H, Yang H, Yue H, et al. Mesenchymal stem cells expressing enos and a cav1 mutant inhibit vascular smooth muscle cell proliferation in a rat model of pulmonary hypertension. Heart Lung Circ. 2017;26:509–18. https://doi.org/10.1016/j.hlc.2016.08.002.
Chow K, Fessel JP, Kaoriihida-Stansbury, et al. Dysfunctional resident lung mesenchymal stem cells contribute to pulmonary microvascular remodeling. Pulm Circ. 2013;3:31–49. https://doi.org/10.4103/2045-8932.109912.
Guillamat-Prats R, Puig F, Camprubi-Rimblas M, et al. Intratracheal instillation of alveolar type ii cells enhances recovery from acute lung injury in rats. J Heart Lung Transpl. 2018;37:782–91. https://doi.org/10.1016/j.healun.2017.10.025.
Wang G, Bunnell BA, Painter RG, et al. Adult stem cells from bone marrow stroma differentiate into airway epithelial cells: potential therapy for cystic fibrosis. Proc Natl Acad Sci U S A. 2005;102:186–91. https://doi.org/10.1073/pnas.0406266102.
Zeng M, Chen Q, Ge S, et al. Overexpression of foxm1 promotes differentiation of bone marrow mesenchymal stem cells into alveolar type ii cells through activating wnt/beta-catenin signalling. Biochem Biophys Res Commun. 2020;528:311–7. https://doi.org/10.1016/j.bbrc.2020.05.042.
Liu A, Chen S, Cai S, et al. Wnt5a through noncanonical wnt/jnk or wnt/pkc signaling contributes to the differentiation of mesenchymal stem cells into type ii alveolar epithelial cells in vitro. PLoS ONE. 2014;9:e90229. https://doi.org/10.1371/journal.pone.0090229.
Jiao X, Lv Q, Cao SN. Microrna-26b-5p promotes development of neonatal respiratory distress syndrome by inhibiting differentiation of mesenchymal stem cells to type ii of alveolar epithelial cells via regulating wnt5a. Eur Rev Med Pharmacol Sci. 2019;23:1681–7. https://doi.org/10.26355/eurrev_201902_17130.
Bernard O, Jeny F, Uzunhan Y, et al. Mesenchymal stem cells reduce hypoxia-induced apoptosis in alveolar epithelial cells by modulating hif and ros hypoxic signaling. Am J Physiol Lung Cell Mol Physiol. 2018;314:L360–71. https://doi.org/10.1152/ajplung.00153.2017.
Thery C, Zitvogel L, Amigorena S. Exosomes: composition, biogenesis and function. Nat Rev Immunol. 2002;2:569–79. https://doi.org/10.1038/nri855.
Sung BH, Parent CA, Weaver AM. Extracellular vesicles: critical players during cell migration. Dev Cell. 2021;56:1861–74. https://doi.org/10.1016/j.devcel.2021.03.020.
Huang G, Garikipati V, Zhou Y, et al. Identification and comparison of hyperglycemia-induced extracellular vesicle transcriptome in different mouse stem cells. Cells. 2020;9. https://doi.org/10.3390/cells9092098.
Johnstone RM, Adam M, Hammond JR, Orr L, Turbide C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J Biol Chem. 1987;262:9412–20.
Taha EA, Ono K, Eguchi T. Roles of extracellular hsps as biomarkers in immune surveillance and immune evasion. Int J Mol Sci. 2019;20. https://doi.org/10.3390/ijms20184588.
Xiang MX, He AN, Wang JA, Gui C. Protective paracrine effect of mesenchymal stem cells on cardiomyocytes. J Zhejiang Univ Sci B. 2009;10:619–24. https://doi.org/10.1631/jzus.B0920153.
Okada H, Suzuki J, Futamatsu H, Maejima Y, Hirao K, Isobe M. Attenuation of autoimmune myocarditis in rats by mesenchymal stem cell transplantation through enhanced expression of hepatocyte growth factor. Int Heart J. 2007;48:649–61. https://doi.org/10.1536/ihj.48.649.
Miller LW. Trial of embryonic stem cell-derived cardiac progenitor cells: an encouraging start. J Am Coll Cardiol. 2018;71:439–42. https://doi.org/10.1016/j.jacc.2017.11.042.
Klinger JR, Pereira M, Del TM, et al. Mesenchymal stem cell extracellular vesicles reverse sugen/hypoxia pulmonary hypertension in rats. Am J Respir Cell Mol Biol. 2020;62:577–87. https://doi.org/10.1165/rcmb.2019-0154OC.
Willis GR, Fernandez-Gonzalez A, Anastas J, et al. Mesenchymal stromal cell exosomes ameliorate experimental bronchopulmonary dysplasia and restore lung function through macrophage immunomodulation. Am J Respir Crit Care Med. 2018;197:104–16. https://doi.org/10.1164/rccm.201705-0925OC.
Monroe MN, Zhaorigetu S, Gupta VS, et al. Extracellular vesicles influence the pulmonary arterial extracellular matrix in congenital diaphragmatic hernia. Pediatr Pulmonol. 2020;55:2402–11. https://doi.org/10.1002/ppul.24914.
Sharma M, Bellio MA, Benny M, et al. Mesenchymal stem cell-derived extracellular vesicles prevent experimental bronchopulmonary dysplasia complicated by pulmonary hypertension. Stem Cells Transl Med. 2022;11:828–40. https://doi.org/10.1093/stcltm/szac041.
Willis GR, Fernandez-Gonzalez A, Reis M, et al. Mesenchymal stromal cell-derived small extracellular vesicles restore lung architecture and improve exercise capacity in a model of neonatal hyperoxia-induced lung injury. J Extracell Vesicles. 2020;9:1790874. https://doi.org/10.1080/20013078.2020.1790874.
Ge L, Jiang W, Zhang S, et al. Mesenchymal stromal cell-derived exosomes attenuate experimental pulmonary arterial hypertension. Curr Pharm Biotechnol. 2021;22:1654–62. https://doi.org/10.2174/1389201022666201231113127.
Chen JY, An R, Liu ZJ, et al. Therapeutic effects of mesenchymal stem cell-derived microvesicles on pulmonary arterial hypertension in rats. Acta Pharmacol Sin. 2014;35:1121–8. https://doi.org/10.1038/aps.2014.61.
Liu Z, Liu J, Xiao M, et al. Mesenchymal stem cell-derived microvesicles alleviate pulmonary arterial hypertension by regulating renin-angiotensin system. J Am Soc Hypertens. 2018;12:470–8. https://doi.org/10.1016/j.jash.2018.02.006.
Klinger JR, Pereira M, Tatto MD, et al. Effect of dose, dosing intervals, and hypoxic stress on the reversal of pulmonary hypertension by mesenchymal stem cell extracellular vesicles. Pulm Circ. 2021;11:20458940211046137. https://doi.org/10.1177/20458940211046137.
Devine SM, Cobbs C, Jennings M, Bartholomew A, Hoffman R. Mesenchymal stem cells distribute to a wide range of tissues following systemic infusion into nonhuman primates. Blood. 2003;101:2999–3001. https://doi.org/10.1182/blood-2002-06-1830.
Sarkar D, Spencer JA, Phillips JA, et al. Engineered cell homing. Blood. 2011;118:e184–91. https://doi.org/10.1182/blood-2010-10-311464.
Zhang S, Lv C, Yang X, et al. Corticosterone mediates the inhibitory effect of restraint stress on the migration of mesenchymal stem cell to carbon tetrachloride-induced fibrotic liver by downregulating cxcr4/7 expression. Stem Cells Dev. 2015;24:587–96. https://doi.org/10.1089/scd.2014.0243.
Smith AN, Muffley LA, Bell AN, Numhom S, Hocking AM. Unsaturated fatty acids induce mesenchymal stem cells to increase secretion of angiogenic mediators. J Cell Physiol. 2012;227:3225–33. https://doi.org/10.1002/jcp.24013.
Bi L, Wang G, Yang D, Li S, Liang B, Han Z. Effects of autologous bone marrow-derived stem cell mobilization on acute tubular necrosis and cell apoptosis in rats. Exp Ther Med. 2015;10:851–6. https://doi.org/10.3892/etm.2015.2592.
Karp JM, Leng TG. Mesenchymal stem cell homing: the devil is in the details. Cell Stem Cell. 2009;4:206–16. https://doi.org/10.1016/j.stem.2009.02.001.
Zhang X, Sarkar K, Rey S, et al. Aging impairs the mobilization and homing of bone marrow-derived angiogenic cells to burn wounds. J Mol Med (Berl). 2011;89:985–95. https://doi.org/10.1007/s00109-011-0754-2.
Tomar GB, Srivastava RK, Gupta N, et al. Human gingiva-derived mesenchymal stem cells are superior to bone marrow-derived mesenchymal stem cells for cell therapy in regenerative medicine. Biochem Biophys Res Commun. 2010;393:377–83. https://doi.org/10.1016/j.bbrc.2010.01.126.
Lin L, Lin H, Bai S, Zheng L, Zhang X. Bone marrow mesenchymal stem cells (bmscs) improved functional recovery of spinal cord injury partly by promoting axonal regeneration. Neurochem Int. 2018;115:80–4. https://doi.org/10.1016/j.neuint.2018.02.007.
Melief SM, Zwaginga JJ, Fibbe WE, Roelofs H. Adipose tissue-derived multipotent stromal cells have a higher immunomodulatory capacity than their bone marrow-derived counterparts. Stem Cells Transl Med. 2013;2:455–63. https://doi.org/10.5966/sctm.2012-0184.
Ankrum JA, Ong JF, Karp JM. Mesenchymal stem cells: immune evasive, not immune privileged. Nat Biotechnol. 2014;32:252–60. https://doi.org/10.1038/nbt.2816.
Epperly MW, Guo H, Gretton JE, Greenberger JS. Bone marrow origin of myofibroblasts in irradiation pulmonary fibrosis. Am J Respir Cell Mol Biol. 2003;29:213–24. https://doi.org/10.1165/rcmb.2002-0069OC.
Yan X, Liu Y, Han Q, et al. Injured microenvironment directly guides the differentiation of engrafted flk-1(+) mesenchymal stem cell in lung. Exp Hematol. 2007;35:1466–75. https://doi.org/10.1016/j.exphem.2007.05.012.
Hansmann G, Chouvarine P, Diekmann F, et al. Human umbilical cord mesenchymal stem cell-derived treatment of severe pulmonary arterial hypertension. Nat Cardiovasc Res. 2022;1:568–76. https://doi.org/10.1038/s44161-022-00083-z.
Acknowledgements
Not applicable.
Funding
This work was supported by the grants from Quanzhou City Science& Technology Program of China (2020N033s), the Health Science and Technology Plan Project of Fujian Province, China (2020QNB029).
Author information
Authors and Affiliations
Contributions
SG wrote the manuscript, and designed the figures; SG and DW (deceased) edited the manuscript; All authors reviewed the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
^Deceased March 29, 2024.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Guo, S., Wang^, D. Novel insights into the potential applications of stem cells in pulmonary hypertension therapy. Respir Res 25, 237 (2024). https://doi.org/10.1186/s12931-024-02865-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12931-024-02865-4