
Abstract
Background
Type 1 Diabetes (T1D) is a T cell-mediated autoimmune disorder caused by the destruction of insulin-secreting cells. B7-H3 (CD276) plays a vital role in T cell response. However, B7-H3 expression and its clinical significance in T1D remain unclear. The aim of this study was to investigate the correlations between the expression of B7-H3 and clinical parameters in T1D patients. The possible role of B7-H3 gene variants with T1D was also discussed.
Methods
Four B7-H3 single nucleotide polymorphisms (SNPs) were genotyped in 121 T1D patients and 120 healthy controls by polymerase chain reaction (PCR) direct sequencing. Expression of membrane B7-H3 (mB7-H3) in peripheral blood lymphocytes was determined by flow cytometry. Levels of soluble B7-H3 (sB7-H3) in serum were analyzed by enzyme linked immunosorbent assay (ELISA).
Results
The B7-H3 haplotype T-A-C-T was less frequently observed in T1D patients compared to the controls (OR: 0.31, 95% CI: 0.16–0.61). B7-H3 expression on monocytes showed significant upregulation in T1D patients and was positively correlated with several clinical features including ALT, fast C-peptide 120 min, HbAlc, IFN-γ, IL-6 and TNF-α (P < 0.05). The concentration of sB7-H3 in serum increased in T1D patients (P < 0.0001). We also observed that B7-H3-T-A-C-T was associated with the decreased release of sB7-H3 but not the membrane form.
Conclusions
B7-H3 may act as a potential biomarker related to the pathogenesis of T1D. The B7-H3-T-A-C-T polymorphism variant is associated with the low risk of T1D as well as less release of sB7-H3.
Similar content being viewed by others

Background
Type 1 diabetes (T1D) is a chronic autoimmune disease that leads to the selective loss of insulin-producing β-cells by activating the T cells [1]. The co-stimulatory and co-inhibitory pathways modulate T cells activation and hence play a vital role in T1D. Scientists had made several advances in the past to understand how co-stimulatory and co-inhibitory pathways affect T1D, such as CD28/B7.1, CD40/CD40L, ICOS/ICOSL and PD1/PD-L1 [2,3,4,5,6]. B7-H3, as a co-stimulatory molecule, which belongs to the B7 immunoglobulin superfamily, is frequently increased in response to the autoantigens and pathogens during host T cell immunoregulation [7].
The biological effect of B7-H3 for its co-stimulatory and co-inhibitory properties remains controversial [8, 9]. There are studies revealing the capability of B7-H3 in promoting CD4+ and CD8+ T cell proliferation, cytotoxic T lymphocytes (CTLs) induction and interferon-gamma (IFN-γ) secretion in vitro [8, 10]. However, some studies speculated that B7-H3 may also have the inhibitory function. B7-H3 could inhibit the proliferation of CD4+ and CD8+ T cells and crucial transcription factors namely nuclear factor of activated T-cells (NFAT), nuclear factor kappa B (NF-κB) and activator protein 1 (AP-1) [10,11,12]. In neuroblastoma, B7-H3 exerted a protective role and thereby downregulated natural killer (NK) cell function [13]. Additionally, it was reported that the soluble and membrane form of B7-H3 could inhibit NK cell-mediated lysis in glioma [14]. Recently, we demonstrated the upregulated expression of sB7-H3 in T1D patients and found the correlation with gender as well as serum levels of creatinine (Cr), blood urea nitrogen (BUN), albumin to creatinine ratio (ACR) and high-density lipoprotein (HDL) [15]. Nevertheless, the expression and function of B7-H3 in T1D remains unknown.
In the present study, we analyzed the expression of B7-H3 and its clinical significance in T1D patients. We aimed to investigate four single nucleotide polymorphisms (SNPs) of B7-H3 genes to determine its associations with T1D risk in order to find the correlations among B7-H3 haplotype, secretion of soluble B7-H3 and biochemical parameters in T1D.
Methods
Patients and samples
In this study, we included the blood samples of 121 patients affected by T1D during the period 2015 and 2018 at the Second Affiliated Hospital of Soochow University. ‘The T1D Exchange Clinic Registry’ was chosen as the diagnostic criteria for T1D [16]. We matched 120 healthy controls with the T1D patients based on their age, gender and race, and they did not suffer from conditions like infectious diseases, cancer, acute or chronic inflammatory diseases. From the selected patients, we enrolled 52 T1D patients and 55 healthy controls to detect the expression of membrane form of B7-H3 on monocytes, and 103 T1D patients and 73 healthy controls to examine the concentration of soluble B7-H3. We obtained approval for this study from the Ethics Commitee of the Second Affiliated Hospital of Soochow University.
DNA extraction and polymorphism genotyping
We followed the standard procedure to isolate the genomic DNA from peripheral blood leucocytes and stored the DNA samples at −20 °C. We selected four SNPs in the B7-H3 gene based on the report presented by our team in the past [17]. Primers were designed according to the gene sequence (accession number AF363458) in Genbank to amplify the B7-H3 fragments. B7-H3 mutations were screened by direct sequencing the PCR products. For 20 samples, PCR amplification and sequencing of genomic DNA were repeated up to 2 times to confirm complete concordance.
Flow cytometry
We analyzed the serum and plasma using the routine procedures at the Jiangsu Institute of Clinical Immunology. The expression of membrane B7-H3 on monocytes in peripheral blood from patients and controls was detected by flow cytometry as previously described [18]. The gating strategy for the CD14+B7-H3+ cell subset was described in Supplementary Fig. 1, Additional File 1. FlowJo software (Tree Star, Ashland, OR) was used to analyze the data.
Soluble B7-H3 measurement
Serum collected from the peripheral blood of the T1D patients and healthy controls were stored at −80 °C. We used the lab-developed ELISA kit to detect the levels of sB7-H3 [19]. The microplate reader (Bio-Rad Laboratories, Hercules, CA, USA) was used to analyze the OD value at 450 nm.
Cytokines production
Cytometric bead array system (CBA) (BD-Pharmingen, CA) was used to quantify the cytokines including IL-2, IL-4, IL-6, IL-10, IL-17A, IFN-γ and tumor necrotic factor-alpha (TNF-α). 50 μl sera were mixed with 50 μl beads in each tube to observe the reaction seperately. PE detection reagent was then added and incubated for 3 h at room temperature. Finally, flow cytometry and BDTM CBA Software were applied to get the acquired data.
Statistical analysis
The statistical analysis was made using SNPstats software (http://bioinfo.iconcologia.net/SNPstats) [20]. Similar to logistic regression (LR), the odds ratio (OR) and 95% confidence interval (CI) were revealed. The magnitude of linkage disequilibrium (LD) was evaluated using the D’ and r2 calculations. Statistical analysis was performed by SPSS 22.0 (IBM Corporation, USA). Student’s t test was included and the differences between the groups were analyzed. Spearman’s rank correlation analysis was used and the correlation between B7-H3 protein levels and clinical variables were analyzed. P < 0.05 was considered statistically significant. Finally, we used GraphPad Prism5.0 (Graph-Pad Software, La Jolla, CA) for drawing graphs.
Results
Single nucleotide polymorphism (SNP) analysis
Patients’ clinical characteristics were summarized in Table 1. We successfully genotyped all the four SNPs in 121 T1D patients and 120 healthy controls (See Supplementary Fig. 2, Additional File 2). Table 2 showed the allele and genotype distribution of these four SNPs. The genotypic distribution in controls for all four SNPs was conformed to Hardy-Weinberg equilibrium (HWE). However, the genotype and allele distributions of all the four SNPs differed significantly between the T1D patients and the healthy controls (P < 0.05). The dominant model showed the smallest Akaike’s information criterion value (See Supplementary Table 1, Additional File 3). Thus, it was enrolled as the best inheritance model.
Linkage disequilibrium (LD) and haplotypes association analysis
We calculated the pairwise LD between the four SNPs of the B7-H3 for both T1D patients and healthy controls in Chinese Han population. We detected strong LD (D’ > 0.97) between some pairs of the markers in the B7-H3 (See Supplementary Fig. 3, Additional File 4). We found that the association analysis of the haplotypes with T1D and the genotypes by LR remained similar (Table 3). Moreover, our results demonstrated the significant correlation between haplotype T-A-C-T and the disease (P < 0.0001); T-A-C-T haplotype was less frequently observed in cases when compared to the controls (OR: 0.31, 95% CI: 0.16–0.61). These results indicated that haplotypes might act as a protective phenotype in Chinese Han patients with T1D. We further adjusted the association analysis of the haplotypes by sex.
Increased membrane B7-H3 expression in T1D and its correlation with disease activity
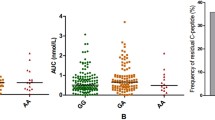
Of the 52 T1D patients and 55 healthy controls enrolled for flow cytometry analysis, the expression of B7-H3 on CD14+ monocytes was significantly higher in T1D patients (13.913 ± 8.640 vs 10.142 ± 4.596, P = 0.0295) (Fig. 1a). The mean fluorescence intensity (MFI) of mB7-H3 on CD14+ monocytes in PB samples from HC and T1D were also analyzed (See Supplementary Fig. 4, Additional File 5). Details about the statistical analysis of the correlation between clinical features and membrane levels of B7-H3 of T1D patients were summarized in Table 4. Our results revealed that B7-H3 had positive correlation with alanine aminotransferase (ALT) (r = 0.4135, P = 0.0080), fast C-peptide 120 min (r = 0.5515, P = 0.0117), hemoglobin A1c (HbAlc) (r = 0.3935, P = 0.0210), IFN-γ (r = 0.5943, P = 0.0003), IL-6 (r = 0.4057, P = 0.0212) and TNF-α (r = 0.5482, P = 0.0012). But the remaining parameters were not statistically significant.

Expression of B7-H3 in HC and T1D patients. a. Increased expression level of mB7-H3 on CD14+ monocytes in PB samples from T1D patients (n = 52) when compared with that from controls (n = 55, P < 0.05). b. The concentration of sB7-H3 is increased in T1D patients (n = 103) when compared with controls (n = 73, P < 0.0001)
Association between B7-H3 haplotypes and the expression of B7-H3 protein in T1D patients
We previously reported that the soluble form of B7-H3 was increased in T1D patients, and showed that titers of sB7-H3 were correlated with serum levels of Cr, BUN, ACR, HDL and gender [15]. In this study, the average concentration of sB7-H3 was (5.111 ± 2.821) ng/ml and (3.321 ± 1.729) ng/ml in T1D patients and healthy controls (P < 0.0001) respectively (Fig. 1b). The expression of membrane B7-H3 had no significant difference between the T1D patients carrying the different haplotypes (Fig. 2a). Concurrently we measured the concentration of sB7-H3 in the serum of T1D patients carrying different haplotypes and found that the T1D patients with T-A-C-T haplotype had lower concentrations of sB7-H3 when compared with the C-C-A-C haplotype (P = 0.0384; Fig. 2b).

Association of the C-C-A-C gene and T-A-C-T gene polymorphisms of the B7-H3 gene with different isoform of B7-H3. The T1D patients were divided into two groups according to their C-C-A-C or T-A-C-T haplotype. The levels of membrane form of B7-H3 (a) and soluble form of B7-H3(b) are shown
Discussion
Immune system and disease development are closely related and immune response can protect the body against different types of diseases. Therefore, it is critical to understand the co-stimulatory and co-inhibitory signals on multiple immune cell types in T1D. In this study, our research demonstrated that the expression of mB7-H3 on monocytes was significantly higher than healthy controls. The B7-H3-T-A-C-T polymorphism variant was associated with a decreased risk of T1D and furthermore was correlated with the secretion of sB7-H3 in serum.
The B7 family, including the CD28/B7 axis, ICOS/ICOSL pathway, receptors like TIM3 and B7-H4, is important for the regulation of immune responses mediated by antigen-specific T cells and also has a significant impact on T1D [21,22,23,24,25]. As an important member of the B7 family, the role of B7-H3 (CD276) in T1D remains unclear. B7-H3 was identified in 2001 as a cell surface molecule in the B7 immunoglobulin superfamily, which played an important role in the initiation and termination of immune cell responses as well as cancer development [8]. However, its receptor has not been identified.
One study found the involvement of B7-H3 in the innate immune monocyte/macrophage-mediated inflammatory response and confirmed that B7-H3 was associated with human sepsis and could augment the inflammatory responses [26]. In line with the study above, our previous experiments showed that B7-H3 was not expressed on T cells [18]. In addition to the investigation of the expression of mB7-H3 on CD14+ monocytes, we also observed increased levels of sB7-H3 in T1D patients when compared with healthy controls. A variety of co-stimulatory molecules exist both in soluble and membrane forms. Soluble co-stimulatory molecules could be produced by proteolytic cleavage and/or splicing of mRNA [27, 28]. Therefore, soluble form of B7-H3 in T1D may due to the cleavage of its membrane form. High mB7-H3 levels on monocytes along with the upregulation of sB7-H3 in patients suggested its co-stimulatory function, which stimulated the self-tolerance breakdown and thus led to the autoimmunity. Our results revealed that B7-H3 played a stimulatory role in T1D. The basic mechanism of controlling and regulating its production in different types of cells remains to be elucidated.
Since monocytes are the key producers of inflammatory factors and several studies also demonstrated that the intermediate CD14+ monocytes produced TNF-α in abundance and monocytes may play an essential role in promoting the inflammatory response in T1D [29, 30]. Previous studies have shown the association of B7-H3 with inflammatory reactions [26]. B7-H3 could amplify LPS, NF-κB p65 and MAPK p38 signals and make them participate in monocyte/macrophage-mediated inflammatory responses [26, 31]. B7-H3 also participated in the progression of asthma and augmented the inflammatory response independent of the Toll-like receptor 2 (TLR2) pathways [32]. Consistent with the above results, our study indicated the proinflammatory role of B7-H3 in the progression of T1D.
In addition, we studied the relationship between clinical features and levels of mB7-H3 in T1D patients. The expression of mB7-H3 was significantly correlated with the secretion of IFN-γ and TNF-α. We accorded these results with initial studies where B7-H3 synergistically promoted the secretion of cytokine IFN-γ and moderately upregulated the TNF-α. Significant correlation of the expression B7-H3 in T1D and the remaining clinical features could be explained with the co-stimulatory effect. Compared with the above results, our former study reported that sB7-H3 and renal function (Cr, BUN, ACR) was positively correlated and assumed that the soluble form may engage in the progression of diabetic nephropathy [15]. With all these results discussed above, we hypothesized that B7-H3 might act as a co-stimulator of innate immunity, which led to pathological damage by activating inflammatory response as well as enhancing T cell- mediated immune response. The upregulation of B7-H3 in the pathogenesis of T1D resulted in the pancreatic islet suffering autoimmune destructions. Previously, our group reported that patients suffering from rheumatoid arthritis (RA) had a higher frequency of T-A-T-C haplotype of B7-H3, and confirmed that it was associated with RA risk and affected the release of the soluble form from the cell surface [17]. Herein, B7-H3-T-A-C-T was observed to associate with the protective role of T1D, and similarly, with soluble B7-H3 expression.
Despite the above findings, our research has some shortcomings due to the lack of human samples. In addition, we could not speculate possible answers when having less samples. More detailed study is recommended with larger samples to investigate the existence of a haplotype-dependent distribution pattern for B7-H3 expression in Chinese individuals and T1D patients.
Conclusions
In this study, we found a significant upregulation of mB7-H3 on monocytes and its correlation with several clinical features including ALT, fast C-peptide 120 min, HbAlc, IFN-γ, IL-6 and TNF-α in T1D patients. Moreover, the B7-H3 haplotype T-A-C-T was less frequently observed in T1D patients and was correlated with the decreased levels of soluble form but not the membrane form of B7-H3. Therefore, the B7-H3-T-A-C-T polymorphism variant was associated with less risk of T1D as well as the release of sB7-H3. The findings of this study indicated that B7-H3 might act as a potential biomarker related to the pathogenesis of T1D.
Availability of data and materials
The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- T1D:
-
Type 1 Diabetes
- SNP:
-
single nucleotide polymorphisms
- PCR:
-
polymerase chain reaction
- mB7-H3:
-
membrane form of B7-H3
- sB7-H3:
-
soluble form of B7-H3
- MFI:
-
mean fluorescence intensity
- Cr:
-
creatinine
- BUN:
-
blood urea nitrogen
- UA:
-
uric acid
- ALT:
-
alanine aminotransferase
- AST:
-
aspartate transferase
- TC:
-
cholesterol
- TG:
-
triglycerides
- LDL:
-
low-density lipoprotein
- HDL:
-
high-density lipoprotein
- IAA:
-
insulin autoantibody
References
Atkinson MA, Eisenbarth GS, Michels AW. Type 1 diabetes. Lancet. 2014;383(9911):69–82.
Salomon B, Lenschow DJ, Rhee L, Ashourian N, Singh B, Sharpe A, Bluestone JA. B7/CD28 costimulation is essential for the homeostasis of the CD4+CD25+ immunoregulatory T cells that control autoimmune diabetes. Immunity. 2000;12(4):431–40.
Fife BT, Pauken KE, Eagar TN, Obu T, Wu J, Tang Q, Azuma M, Krummel MF, Bluestone JA. Interactions between PD-1 and PD-L1 promote tolerance by blocking the TCR-induced stop signal. Nat Immunol. 2009;10(11):1185–92.
Kornete M, Mason ES, Girouard J, Lafferty EI, Qureshi S, Piccirillo CA. Th1-Like ICOS+ Foxp3+ Treg Cells Preferentially Express CXCR3 and Home to β-Islets during Pre-Diabetes in BDC2.5 NOD Mice. PloS one. 2015;10(5):e0126311.
Paterson AM, Brown KE, Keir ME, Vanguri VK, Riella LV, Chandraker A, Sayegh MH, Blazar BR, Freeman GJ, Sharpe AH. The programmed death-1 ligand 1:B7-1 pathway restrains diabetogenic effector T cells in vivo. J Immunol. 2011;187(3):1097–105.
Baker RL, Wagner DH, Haskins K. CD40 on NOD CD4 T cells contributes to their activation and pathogenicity. J Autoimmun. 2008;31(4):385–92.
Luo L, Zhu G, Xu H, Yao S, Zhou G, Zhu Y, Tamada K, Huang L, Flies AD, Broadwater M, et al. B7-H3 promotes pathogenesis of autoimmune disease and inflammation by regulating the activity of different T cell subsets. PLoS One. 2015;10(6):e0130126.
Chapoval AI, Ni J, Lau JS, Wilcox RA, Flies DB, Liu D, Dong H, Sica GL, Zhu G, Tamada K, et al. B7-H3: a costimulatory molecule for T cell activation and IFN-gamma production. Nat Immunol. 2001;2(3):269–74.
Suh WK, Gajewska BU, Okada H, Gronski MA, Bertram EM, Dawicki W, Duncan GS, Bukczynski J, Plyte S, Elia A, et al. The B7 family member B7-H3 preferentially down-regulates T helper type 1-mediated immune responses. Nat Immunol. 2003;4(9):899–906.
Hofmeyer KA, Ray A, Zang X. The contrasting role of B7-H3. Proc Natl Acad Sci U S A. 2008;105(30):10277–8.
Wang L, Fraser CC, Kikly K, Wells AD, Han R, Coyle AJ, Chen L, Hancock WW. B7-H3 promotes acute and chronic allograft rejection. Eur J Immunol. 2005;35(2):428–38.
Prasad DV, Nguyen T, Li Z, Yang Y, Duong J, Wang Y, Dong C. Murine B7-H3 is a negative regulator of T cells. J Immunol (Baltimore, Md : 1950). 2004;173(4):2500–6.
Castriconi R, Dondero A, Augugliaro R, Cantoni C, Carnemolla B, Sementa AR, Negri F, Conte R, Corrias MV, Moretta L, et al. Identification of 4Ig-B7-H3 as a neuroblastoma-associated molecule that exerts a protective role from an NK cell-mediated lysis. Proc Natl Acad Sci U S A. 2004;101(34):12640–5.
Lemke D, Pfenning PN, Sahm F, Klein AC, Kempf T, Warnken U, Schnölzer M, Tudoran R, Weller M, Platten M, et al. Costimulatory protein 4IgB7H3 drives the malignant phenotype of glioblastoma by mediating immune escape and invasiveness. Clin Cancer Res. 2012;18(1):105–17.
Fang C, Li S, Xu R, Guo H, Jiang R, Ding S, Chen X, Huang Y, Hu J, Liu C, et al. Soluble B7-H3 (sB7-H3) is over-expressed in the serum of type 1 diabetes patients. Diabetes Res Clin Pract. 2018;143:332–6.
Beck RW, Tamborlane WV, Bergenstal RM, Miller KM, DuBose SN, Hall CA. The T1D exchange clinic registry. J Clin Endocrinol Metab. 2012;97(12):4383–9.
Sun J, Liu C, Gao L, Guo Y, Zhang Y, Wu P, Jiang J, Yan R, Zhang X. Correlation between B7-H3 expression and rheumatoid arthritis: A new polymorphism haplotype is associated with increased disease risk. Clin Immunol (Orlando, Fla). 2015;159(1):23–32.
Jiang J, Jiang J, Liu C, Zhang G, Gao L, Chen Y, Zhu R, Wang T, Wang F, Zhang X, et al. Enhancement of membrane B7-H3 costimulatory molecule but reduction of its soluble form in multiple sclerosis. J Clin Immunol. 2013;33(1):118–26.
Zhang G, Hou J, Shi J, Yu G, Lu B, Zhang X. Soluble CD276 (B7-H3) is released from monocytes, dendritic cells and activated T cells and is detectable in normal human serum. Immunology. 2008;123(4):538–46.
Solé X, Guinó E, Valls J, Iniesta R, Moreno V. SNPStats: a web tool for the analysis of association studies. Bioinformatics (Oxford, England). 2006;22(15):1928–9.
Principi N, Berioli MG, Bianchini S, Esposito S. Type 1 diabetes and viral infections: what is the relationship? J Clin Virol. 2017;96:26–31.
Ceeraz S, Nowak EC, Noelle RJ. B7 family checkpoint regulators in immune regulation and disease. Trends Immunol. 2013;34(11):556–63.
Herman AE, Freeman GJ, Mathis D, Benoist C. CD4+CD25+ T regulatory cells dependent on ICOS promote regulation of effector cells in the prediabetic lesion. J Exp Med. 2004;199(11):1479–89.
Wang X, Hao J, Metzger DL, Mui A, Ao Z, Akhoundsadegh N, Langermann S, Liu L, Chen L, Ou D, et al. Early treatment of NOD mice with B7-H4 reduces the incidence of autoimmune diabetes. Diabetes. 2011;60(12):3246–55.
Sanchez-Fueyo A, Tian J, Picarella D, Domenig C, Zheng XX, Sabatos CA, Manlongat N, Bender O, Kamradt T, Kuchroo VK, et al. Tim-3 inhibits T helper type 1-mediated auto- and alloimmune responses and promotes immunological tolerance. Nat Immunol. 2003;4(11):1093–101.
Zhang G, Wang J, Kelly J, Gu G, Hou J, Zhou Y, Redmond HP, Wang JH, Zhang X. B7-H3 augments the inflammatory response and is associated with human sepsis. J Immunol (Baltimore, Md : 1950). 2010;185(6):3677–84.
Nielsen C, Ohm-Laursen L, Barington T, Husby S, Lillevang ST. Alternative splice variants of the human PD-1 gene. Cell Immunol. 2005;235(2):109–16.
Logue EC, Bakkour S, Murphy MM, Nolla H, Sha WC. ICOS-induced B7h shedding on B cells is inhibited by TLR7/8 and TLR9. J Immunol. 2006;177(4):2356–64.
Ren X, Mou W, Su C, Chen X, Zhang H, Cao B, Li X, Wu D, Ni X, Gui J, et al. Increase in peripheral blood intermediate monocytes is associated with the development of recent-onset type 1 diabetes mellitus in children. Int J Biol Sci. 2017;13(2):209–18.
Ryba-Stanisławowska M, Myśliwska J, Juhas U, Myśliwiec M. Elevated levels of peripheral blood CD14(bright) CD16+ and CD14(dim) CD16+ monocytes may contribute to the development of retinopathy in patients with juvenile onset type 1 diabetes. APMIS. 2015;123(9):793–9.
Chen X, Li Y, Blankson S, Liu M, Huang D, Redmond HP, Huang J, Wang JH, Wang J. B7-H3 augments inflammatory responses and exacerbates brain damage via amplifying NF-κB p65 and MAPK p38 activation during experimental pneumococcal meningitis. PLoS One. 2017;12(1):e0171146.
Gu W, Zhang X, Yan Y, Wang Y, Huang L, Wang M, Shao X, Chen Z, Ji W. B7-H3 participates in the development of asthma by augmentation of the inflammatory response independent of TLR2 pathway. Sci Rep. 2017;7:40398.
Acknowledgements
The authors express thanks to the staff members of Jiangsu Institute of Clinical Immunology& Jiangsu Key Laboratory of Clinical Immunology, First Affiliated Hospital of Soochow University (Suzhou, China). The authors also wish to thank all subjects who participated in this study.
Funding
This study was funded by National Natural Science Foundation of China (Grant No.81873876, 81801595, 31700778 and 81600607), which provided the funds that were used to purchase materials, reagents and DNA sequencing service. The cost of publication was supported by grant from Jiangsu Provincial Medical Youth Talent (No. QNRC2016755). The funders had no role in study design, collection, analysis and interpretation of data, or writing of the manuscript.
Author information
Authors and Affiliations
Contributions
SD and YS contributed to data analysis, manuscript drafting and critical discussion. LS, SL, RJ and XC contributed to the laboratory testing. ZH and JS collected the data. CL and CF were responsible for study design and execution. XZ was responsible for supervision and mentorship. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Written voluntary informed consent was obtained from all the study participants. All procedures performed in studies involving were reviewed and approved by the Ethics Review Board of the Second Affiliated Hospital of Soochow University in accordance with the Helsinki Declaration of 1975, as revised in 2008.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Additional file 1: Fig. S1.
Expression of mB7-H3 in HC and T1D patients. Flow cytometry detection of B7-H3 on CD14+ monocytes in PB samples from healthy controls and T1D patients.
Additional file 2: Fig. S2
. Polymorphisms of B7-H3 gene in this study. Raw data of electropherograms showed the mutant types of SNPs: rs7173448 (a), rs7173476(b), 1359 A → C (c), rs145827704 (d).
Additional file 3: Supplementary Table 1.
The association analysis of SNP1 with T1D (adjusted by sex) by using logistic regression.
Additional file 4: Fig. S3
. Linkage disequilibrium analysis in B7-H3 gene. D is the deviation between the expected haplotype frequency (under the assumption of no association) and the observed frequency; D’ is a proportion of the maximum value of D, which is scaled in [− 1,1] range; R is the correlation coefficient between alleles.
Additional file 5: Fig. S4
. Expression of mB7-H3 in HC and T1D patients. Bars show the mean and SD of mB7-H3 mean fluorescence intensity (MFI).
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cite this article
Ding, S., Shan, Y., Sun, L. et al. Evaluation of the role of B7-H3 haplotype in association with impaired B7-H3 expression and protection against type 1 diabetes in Chinese Han population. BMC Endocr Disord 20, 123 (2020). https://doi.org/10.1186/s12902-020-00592-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12902-020-00592-7