
Abstract
Background
Observational studies have suggested that type 2 Diabetes Mellitus (DM2) is a potentially modifiable risk factor for lower BMD, but the causal relationship is unclear. This study aimed to examine whether the association of DM2 with lower BMD levels was causal by using Mendelian randomization (MR) analyses.
Methods
We collected genome-wide association study data for DM2 and BMD of total body and different skeletal sites from the IEU database. Subsequently, we performed a two-sample Mendelian randomization analysis using the Two Sample MR package.
Results
We identified a positive association between DM2 risk (61,714 DM2 cases and 596,424 controls) and total BMD, and other skeletal sites BMD, such as femoral neck BMD, ultra-distal forearm BMD and heel BMD. However, non-significant trends were observed for the effects of DM2 on lumbar-spine BMD.
Conclusion
In two-sample MR analyses, there was positive causal relationship between DM2 and BMD in both overall samples. In summary, while observational analyses consistently indicate a strong association between DM2 and low BMD, our MR analysis introduces a nuanced perspective. Contrary to the robust association observed in observational studies, our MR analysis suggests a significant link between DM2 and elevated BMD.
Similar content being viewed by others

Introduction
Human longevity has increased significantly as a result of societal advancement and evolution. As a result, the rise in age-related diseases has emerged as a significant concern for people worldwide. Among these conditions, osteoporosis stands out as a widespread geriatric malady, characterized by the gradual weakening of bones, often resulting in an elevated vulnerability to fractures [1, 2]. Furthermore, the outcomes in terms of both medical implications and economic burdens are substantial and untenable [3, 4]. Despite the significant prevalence of osteoporosis, lines of evidence for osteoporosis risk factors have yet to be thoroughly established. Prior studies have unveiled a multitude of factors that contribute to osteoporosis risk, encompassing age, sex, physique, ethnicity, familial predisposition to fractures, specific medication usage, tobacco consumption, insufficient peak bone density, limited physical engagement, and low blood levels of Vit D3 [3]. Lately, a growing body of evidence indicates a potential link between metabolic disruptions and the onset of osteoporosis, which shifts the focus of research to metabolic risk factors [5, 6].
Being the prevailing metabolic ailment, DM2 has a significant threat to aging populations due to its array of complications. Apart from being a strong risk factor for cardiovascular diseases, DM2 may also increase the risk of developing osteoporosis. To date, the potential relationship between diabetes and osteoporosis has been recognized in animal [7]. Nonetheless, contrary to the consistently favorable outcomes observed in laboratory investigations, clinical studies have yielded incongruent findings. A meta-analysis of 15 observational studies found that individuals with DM2 from both genders have higher BMD levels [8]. And some cross-sectional or case-control studies also have not effectively explored the autonomous correlation between diabetes mellitus and osteoporosis [9]. The inconsistency could stem from significant individual variations among patients in clinical research, or it might be attributed to the absence of a clinical correlation between diabetes and BMD levels. In order to thoroughly investigate the association between the DM2 and BMD levels, a research approach more effective than RCT is required.
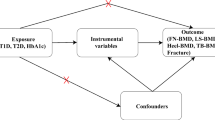
Mendelian randomization (MR) studies, which use an epidemiological approach that assesses the causal effect of a risk factor on an outcome, have been increasingly used to overcome the aforementioned limitations and explore causal relationships [10]. Since genetic variants are randomly assigned, the confounding factors are minimized by the MR method. Genetic variation significantly associated with exposure can therefore be used as instrumental variables (IVs). For instrumental variables to be valid, three conditions must be met: IV1, which is linked to the exposure; IV2, which remains unrelated to the outcome when considering the exposure; and IV3, which maintains independence from all known confounding factors up to the present. (Fig. 1) Currently, a dearth of substantial evidence exists concerning the causative factors underlying osteoporosis. Nevertheless, limited studies have concentrated on investigating the connection between DM2 status and BMD through the utilization of MR analysis.

The flowchart of A two-sample MR
Our aim is to investigate the potentially of the relationship between DM2 and BMD levels using a large-scale genome-wide association study (GWAS) data sets by two-sample MR study. We hypothesize that DM2 is a causal risk factor for increased BMD levels. This study may help to reveal the genetic characteristics and biological mechanisms of DM2 and BMD.
Methods
Study design and assumptions
We first performed two-sample MR to assess the causal relationship between DM2 and BMD. A two-sample MR analysis was applied to explore the causal effects of the DM2 on BMD as our experimental flow chart shows in Fig. 2. A two-sample MR method should conform to three fundamental assumptions: filtered IVs must be strongly related to the DM2; filtered IVs are not correlated with confounding factors; IVs can only influence BMD through the DM2. (Fig. 1) We used publicly available GWAS data with the informed consent and ethical approval previously obtained.

The flowchart of the study. The whole workflow of MR analysis. Note: MR, Mendelian randomization; BMD, bone mineral density; LD, linkage disequilibrium; SNPs, Single-nucleotide polymorphisms
Data source
We acquired the summary statistics of the DM2 from a meta-analysis with ~ 16 million genetic variants in 62,892 DM2 cases and 596,424 controls of European ancestry [11].
We used BMD data from a meta-analysis with five different skeletal sites as outcomes, including TB-BMD (n = 56,284), FN-BMD (n = 32,961), UF-BMD (n = 21,907), H-BMD (n = 426,824), LS-BMD (n = 28,498) [12]. TB-BMD, FN-BMD, FABMD and LS-BMD were measured by DXA, whereas H-BMD was measured by quantitative ultrasound (QUS), which are two completely different procedures.
Instrumental variable selection
For two-sample MR analysis, a genome-wide single-nucleotide polymorphisms (SNPs) significantly associated with DM2 (p < 5 × 10− 8). Then, we pooled all genome-wide significant SNPs that were significantly associated with DM2 and then clumped these SNPs with respect to the lowest p-value corresponding to any of the two using a 1,0000-kb window and pairwise LD r 2 < 0.01. We calculated the proportion of phenotypic variance explained by instrumental variable SNPs of DM2 and computed the F-statistic (> 10) to verify whether they were strong instruments.
MR analysis and sensitivity analysis
We used the inverse-variance weighted (IVW) method as the primary MR approach [13]. MR-Egger, weighted median, and weighted mode further conducted to MR analysis. However, the estimation accuracy produced by MR-Egger is very low. Weighted median gives an accurate estimate based on the assumption that at least 50% of IVs are valid [14]. Weighted mode is sensitive to the difficult bandwidth selection for mode estimation [15].
Sensitivity analysis has been pivotal in MR studies to detect underlying heterogeneity and pleiotropy for MR estimates can be severely violated. We used the Cochran Q statistic and leave-one-out analysis to test for the heterogeneity, and Egger-intercept to test for the pleiotropy [13, 16]. And if there is heterogeneity, random-effects IVW models are applied; otherwise, the fixed-effect IVW model is applied [17].
Based on the MR models and pleiotropy assessments mentions above, we considered a relatively robust inference meeting the following items: (i) MR analyses presented a consistent direction of casual estimates among different methods. (ii) Intercept term derived from MR-Egger regression suggested no directional pleiotropy detected (p > 0.05). (iii) Leave-one-out analysis suggested causal estimate was not strongly driven by a single SNP.
Software
All statistical analyses were conducted using the “Two Sample MR” (version 0.5.7, Stephen Burgess, Chicago, IL, USA) and “Mendelian Randomization” (version 0.8.0) in the statistical program R (version 4.3.1). p < 0.05 was considered as statistically significant. Reporting follows the STROBE-MR statement.
Result
Causal effect of increased DM2 on TB-BMD
The specific MR data for all selected SNPs in the exposure set (DM2) and outcome set (TB-BMD) are presented in Supplementary Table S1. IVW analysis showed that there was a positive causal association between DM2 and TB-BMD (p = 0.002, OR = 1.03) (Table 1). MR Egger analysis showed that there was not a causal association between DM2 and TB-BMD (p = 0.33, OR = 1.02) (Table 1). Weighted median analysis demonstrated that DM2 had a positive causal association with TB-BMD (p < 0.05, OR > 1) (Table 1). Weighted mode analysis demonstrated that DM2 had a positive causal association with TB-BMD (p < 0.05, OR > 1) (Table 1). The estimated effect sizes of the SNPs on both the DM2 (exposure) and TB-BMD (outcome) are presented in scatter plots (Figure S1A). Funnel plot presents symmetrical distribution. (Figure S1B) Plots of the leave-one-out analysis, as shown in Figure S1C, demonstrate that no potentially influential SNP that drive the causal effect.
Causal effect of increased DM2 on FN-BMD
The specific MR data for all selected SNPs in the exposure set (DM2) and outcome set (FN-BMD) are presented in Supplementary Table S2. IVW analysis showed that there was a positive causal association between DM2 and FN-BMD (p = 0.001, OR = 1.04) (Table 1). MR Egger analysis showed that there was not a causal association between DM2 and FN-BMD (p = 0.12, OR = 1.05) (Table 1). Weighted median analysis demonstrated that DM2 had no causal association with FN-BMD (p < 0.05, OR > 1) (Table 1). weighted mode analysis demonstrated that DM2 had a positive causal association with FN-BMD (p < 0.05, OR > 1) (Table 1). The estimated effect sizes of the SNPs on both the DM2 (exposure) and TB-BMD (outcome) are presented in scatter plots (Figure S2A). Funnel plot presents symmetrical distribution. (Figure S2B) Plots of the leave-one-out analysis, as shown in Figure S2C, demonstrate that no potentially influential SNP that drive the causal effect.
Causal effect of increased DM2 on UF -BMD
The specific MR data for all selected SNPs in the exposure set (DM2) and outcome set (UF-BMD) are presented in Supplementary Table S3. IVW analysis showed that there was a positive causal association between DM2 and UF-BMD (p = 7e− 4, OR = 1.05) (Table 1). MR Egger, weighted median, and weighted mode analysis showed that there was not a causal association between DM2 and UF-BMD (p > 0.05, OR > 1) (Table 1). The estimated effect sizes of the SNPs on both the DM2 (exposure) and UF-BMD (outcome) are presented in scatter plots (Figure S3A). Funnel plot presents symmetrical distribution. (Figure S3B) Plots of the leave-one-out analysis, as shown in Figure S3C, demonstrate that no potentially influential SNP that drive the causal effect.
Causal effect of increased DM2 on H -BMD
The specific MR data for all selected SNPs in the exposure set (DM2) and outcome set (H-BMD) are presented in Supplementary Table S4. IVW analysis showed that there was a positive causal association between DM2 and H-BMD (p = 4.3e− 4, OR = 1.03) (Table 1). MR Egger analysis showed that there was not a causal association between DM2 and H-BMD (p > 0.05) (Table 1). Weighted median analysis and weighted mode analysis demonstrated that DM2 had a positive causal association with H-BMD (p < 0.05, OR > 1) (Table 1). The estimated effect sizes of the SNPs on both the DM2 (exposure) and H-BMD (outcome) are presented in scatter plots (Figure S4A). Funnel plot presents symmetrical distribution. (Figure S4B) Plots of the leave-one-out analysis, as shown in Figure S4C, demonstrate that no potentially influential SNP that drive the causal effect.
Causal effect of increased DM2 on LS -BMD
The specific MR data for all selected SNPs in the exposure set (DM2) and outcome set (LS-BMD) are presented in Supplementary Table S5. IVW analysis showed that there was a positive causal association between DM2 and LS-BMD (p = 0.003, OR = 1.05) (Table 1). Weighted median analysis showed that there was not a causal association between DM2 and LS-BMD (p > 0.05, OR > 1) (Table 1). MR Egger and weighted mode showed that there was not a causal association between DM2 and LS-BMD (p > 0.05, OR < 1) (Table 1). The estimated effect sizes of the SNPs on both the DM2 (exposure) and LS-BMD (outcome) are presented in scatter plots (Figure S5A). Funnel plot presents symmetrical distribution. (Figure S5B) Plots of the leave-one-out analysis, as shown in Figure S5C, demonstrate that no potentially influential SNP that drive the causal effect.
Sensitivity analysis
Sensitivity analysis was conducted to verify the reliability of IVW results. IVW and MR-Egger test for heterogeneity showed that there was no heterogeneity in MR analysis results between DM2 and TB-BMD, FN-BMD, UF-BMD, H-BMD and LS-BMD (p > 0.05) (Table 2). And the funnel plots present symmetrical distribution. (Figure S1C-S5C)
Further validation of MR results
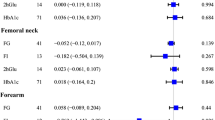
We further verified the IVW results. The results of IVW (fixed effects), weighted median and weighted mode revealed that DM2 had positive causal association with TB-BMD (p < 0.05, OR > 1) and H-BMD (p < 0.05, OR > 1) (Fig. 3). The results of IVW (fixed effects) and weighted mode demonstrated a positive causal association between DM2 and FN-BMD (p < 0.05, OR > 1) (Fig. 3). However, the other MR methods cannot verify the IVW results about DM2 and UF-BMD and DM2 and LS-BMD (p > 0.05) (Fig. 3), indicating that the results about DM2 having a positive causal connection with UF-BMD and LS-BMD are unstable.

Estimated causal effects between DM2 and BMD using different MR methods
Discussion
DM2 influences bone metabolism, but the relation of DM2 with BMD remains inconsistent across studies. Observational studies provide evidence for higher fracture risk for a given BMD in individuals with DM2 compared to those without this condition [18]. Despite this higher fracture risk, some observational studies found that individuals with DM2 tend to have a higher BMD than non-diabetic individuals [19, 20]. In a meta-analysis of 3,437 subjects with DM2 generally have higher BMD as compared with healthy controls, with significant differences of 0.04 (95%CI: 0.02, 0.05) at the femoral neck, 0.06 (95%CI: 0.04, 0.08) at the hip and 0.06 (95%CI: 0.04, 0.07) at the spine [8]. What is the association between DM2 and BMD? Because of confounding in observational studies between DM2 and other metabolic factors known to influence bone homeostasis [21], the precise effects of DM2 and BMD remain unclear. MR has great potential for analyzing the causal associations between diseases and traits. As far as we know, this study is the first to investigate the genetic causal associations between DM2 and different skeletal sites BMD, which provides insights into the inconsistently prior reported relationship. Our MR analysis concluded that overall individuals with DM2 have higher TB-BMD, FN-BMD, UF-BMD, and H-BMD. And the association between DM2 and LS-BMD did not reach statistical significance, raising the possibility that the effect of DM2 on BMD is site-specific. Sensitivity analyses did not essentially change our results or conclusions.
An array of mechanisms potentially underlies the connection between DM2 and rising BMD, though their exact nature remains largely obscure. In the ensuing discussion, we approach the subject from a clinical standpoint and highlight the pivotal factors that exert substantial influence on the intricate interrelationship between DM2 and BMD.
Obesity and hyperinsulinemia have been theorized to constitute two significant attributes of DM2 that exhibit a positive correlation with BMD [22]. However, it was observed that a substantial proportion of the studies included did not fundamentally alter the association even after accounting for BMI. Numerous intricate pathways exist through which obesity might affect the connection between diabetes and BMD. The influence of body fatness on the precision of DXA-based BMD assessments, as evidenced in obese individuals with diabetes, is noteworthy [23]. However, the potential measurement error should be of minimal concern, given that this phenomenon has been demonstrated to potentially result in either underestimation or overestimation of values. Moreover, it has been shown to exert a negligible impact on the precision of BMD measurements. Conversely, adipose tissue releases an extensive array of adipokines that have been suggested to play a role, whether directly or indirectly, in the modulation of bone remodeling processes [24]. Plasma leptin concentrations have been shown to be higher in diabetic men than in healthy controls. Elevated plasma leptin levels have been demonstrated in diabetic men compared to their healthy counterparts [25]. Furthermore, it has been revealed that leptin can hinder osteoclast formation by diminishing the production of RANK/RANKL while enhancing osteoprotegerin levels [26,27,28,29]. And other adipokines, such as adiponectin and resistin, are also found to be present in osteoblasts and osteoclasts [30] (Fig. 4). The impact of these adipokines on bone metabolism remains predominantly unclear, although their potential involvement in directing the differentiation of mesenchymal progenitor cells towards either osteogenic or adipogenic pathways is noteworthy [31, 32].

The possible mechanisms that an association between DM2 and increased BMD.
Several of the reviewed studies suggested that insulin levels might partially mediate the positive association between DM2 and increased BMD [33,34,35]. Individuals with DM2 typically exhibit an excess of insulin. From a physiological perspective, insulin exerts an anabolic influence on bone, primarily attributed to its structural resemblance to Insulin-like growth factor 1 (IGF-1). This resemblance allows insulin to engage with the IGF-1 receptor, which is found on osteoblasts [36]. The signaling pathway of IGF-1 holds paramount importance in bone acquisition [37], as evidenced by both human and mouse investigations revealing a noteworthy and affirmative correlation between IGF-1 levels and BMD [38, 39]. Considering this standpoint, a hypothesis arises that hyperinsulinemia might potentially trigger a mitogenic impact on osteoblasts, promoting their differentiation through the activation of the IGF-1 signaling pathway (Fig. 4). Furthermore, certain indirect effects of insulin on bone formation may conceivably be mediated by osteogenic factors such as amylin, osteoprotegerin, sex steroids, and sex hormone-binding globulin.
Insulin, as an anabolic hormone, holds a pivotal role in governing substrate metabolism across critical organs and tissues, including skeletal muscle, the liver, and adipose tissue [40]. The insulin receptor is expressed in both osteoblasts and osteoclasts. Insulin not only triggers the formation of osteoclasts but also fosters the proliferation, differentiation, and survival of osteoblasts. This collective effect tends to tip the scale in favor of bone formation [41]. Research involving insulin receptor knockout mice suggests that insulin signaling is essential for optimal bone acquisition, possibly owing to insulin’s involvement in governing bone energy metabolism [42, 43]. In fact, the administration of insulin leads to an augmentation in 18 F-fluorodeoxyglucose uptake within bone tissues in mice, a response noticeably diminished in mice wherein the insulin receptor is absent in osteoblasts [44]. Moreover, the activation of the insulin receptor within the growth plate of mice subjected to a hypercaloric diet stimulates skeletal growth and augments the process of growth plate chondrogenesis [45]. Osteoblasts also express the IGF-1 receptor [46]. IGF-1 binds to both the IGF-1 receptor and, with a slightly lower affinity, to the insulin receptor, thereby activating the insulin signaling pathway and eliciting osteoanabolic effects. Additionally, the utilization of Thiazide, which is anticipated to be more prevalent among individuals with diabetes, has been correlated with elevated BMD at various skeletal sites [47, 48]. Similarly, the usage of statins (likewise more common in diabetics) has also demonstrated an association with higher BMD [49, 50].
It is worth mentioning that DM2 affects BMD differently in different parts of the body. We speculated a possible mechanism for site-specific effects of DM2 on BMD could relate to the known disparate effects of DM2 on cortical and trabecular bone [51,52,53] and the significant regional variation in bone microstructure throughout the skeleton [54, 55]. Alternatively, measurement error for LS-BMD due to non-osteoporotic degenerative changes in the spine (such as osteophytes and degenerative disc disease) [56] or technical issues (such as positioning) [57] may have biased associations towards the null. Nonetheless, the precise rationale for potential measurement discrepancies having a greater impact on lumbar spine data in comparison to data from other anatomical sites remains uncertain. Additional investigations into the mechanisms that could potentially elucidate the site-specific effects of DM2 on BMD and other bone characteristics are imperative to shed light on these uncertainties.
Given that genetic instruments typically encapsulate lifelong exposures, the genetic associations examined in this study likely encompass the enduring impact of hyperglycemia on BMD over extended periods. Although limitations in available observational data preclude a reliable epidemiologic assessment of the specific contribution of disease duration to the effect of DM2 on BMD, our MR results are consistent with observational studies showing that individuals DM2 had higher BMD [8]. Moreover, our results also show that individuals DM2 higher total body BMD, femoral neck BMD, ultradistal forearm BMD and heel BMD. However, the presence of DM2 does not necessarily imply a reduction in the risk of fragility fractures, as the determinants of fractures extend regardless of BMD. The comprehensive exploration of the association between DM2 and an increased risk of vertebral fractures underscores the intricate nature of bone health in diabetic individuals. The correlation between vertebral fractures in DM2 and an elevated risk of non-vertebral fractures and mortality accentuates the systemic impact of this condition on skeletal integrity. The integration of Trabecular Bone Score emerges as a valuable adjunct for estimating vertebral fracture risk, offering a more nuanced assessment beyond conventional measures. Encouragingly, recent studies demonstrate the efficacy of interventions such as teriparatide and denosumab in reducing vertebral fracture risk in individuals with DM2 [58, 59]. Furthermore, the identification of increased bone marrow fat in DM2 highlights a potential contributor to bone fragility, reinforcing the necessity for a nuanced understanding of the intricate relationships between diabetes, bone health, and fracture risk. These cumulative findings underscore the importance of adopting a holistic approach to bone health management in individuals with DM2. Integrating advanced assessment tools and targeted interventions becomes imperative to address the multifaceted factors influencing fracture susceptibility in this population.
There are several limitations to this research. Among the MR statistical methods, although the causal effect of exposure on outcome was consistent in both IVW and WM (weighted median or weighted mode) test methods, the results of MR-Egger were less convincing. While this study is confined to the European population, further investigation is needed to determine whether this association exists in other groups. Moreover, the precise rationale for potential measurement discrepancies having a greater impact on lumbar spine data compared to data from other anatomical sites remains uncertain. It is imperative to conduct additional investigations into the mechanisms that could potentially elucidate the site-specific effects of DM2 on BMD and other bone characteristics, shedding light on these uncertainties.
Conclusions
In summary, our MR study provides evidence that genetic increases in DM2 risk have positive effects on TB-BMD, FN-BMD, UF-BMD, and H-BMD. Contrary to the robust association observed in observational studies, our MR analysis suggests a significant link between DM2 and elevated BMD.
Data availability
The article/Supplementary Material contains the original contributions presented in the study. Further questions should be directed to the corresponding author (Kaitan Yang) and first author (Jianbin Guan). The link for the publicly available database: DM2: https://gwas.mrcieu.ac.uk, Id: Ebi-a-GCST006867 PMID:30054458; BMD: , PMID:29304378.
References
Ensrud KE, Crandall CJ. Osteoporosis[J]. Ann Intern Med. 2017;167(3):C17–C32.
Compston JE, McClung MR, Leslie WD. Osteoporosis[J] Lancet. 2019;393(10169):364–76.
Clynes MA, Harvey NC, Curtis EM, et al. The epidemiology of osteoporosis[J]. Br Med Bull. 2020;133(1):105–17.
Burge R, Dawson-Hughes B, Solomon DH, et al. Incidence and economic burden of osteoporosis-related fractures in the United States, 2005–2025[J]. J Bone Miner Res. 2007;22(3):465–75.
Fischer V, Haffner-Luntzer M. Interaction between bone and immune cells: implications for postmenopausal osteoporosis[J]. Semin Cell Dev Biol. 2022;123:14–21.
Polyzos SA, Anastasilakis AD, Efstathiadou ZA, et al. Postmenopausal osteoporosis coexisting with other metabolic diseases: treatment considerations[J]. Maturitas. 2021;147:19–25.
Chau DL, Edelman SV, Chandran M. Osteoporosis and diabetes[J]. Curr Diab Rep. 2003;3(1):37–42.
Ma L, Oei L, Jiang L, et al. Association between bone mineral density and type 2 diabetes mellitus: a meta-analysis of observational studies[J]. Eur J Epidemiol. 2012;27(5):319–32.
Leidig-Bruckner G, Ziegler R. Diabetes mellitus a risk for osteoporosis? [J]. Exp Clin Endocrinol Diabetes. 2001;109(Suppl 2):493–S514.
Davies NM, Holmes MV, Davey SG. Reading mendelian randomisation studies: a guide, glossary, and checklist for clinicians[J]. BMJ. 2018;362:k601.
Xue A, Wu Y, Zhu Z, et al. Genome-wide association analyses identify 143 risk variants and putative regulatory mechanisms for type 2 diabetes[J]. Nat Commun. 2018;9(1):2941.
Medina-Gomez C, Kemp JP, Trajanoska K, et al. Life-course genome-wide association study meta-analysis of total body Bmd and assessment of age-specific effects[J]. Am J Hum Genet. 2018;102(1):88–102.
Burgess S, Davey SG, Davies NM, et al. Guidelines for performing mendelian randomization investigations: update for summer 2023[J]. Wellcome Open Res. 2019;4:186.
Bowden J, Davey SG, Haycock PC, et al. Consistent estimation in mendelian randomization with some invalid instruments using a weighted median estimator[J]. Genet Epidemiol. 2016;40(4):304–14.
Hartwig FP, Davey SG, Bowden J. Robust inference in summary data mendelian randomization via the zero modal pleiotropy assumption[J]. Int J Epidemiol. 2017;46(6):1985–98.
Burgess S, Thompson SG. Interpreting findings from mendelian randomization using the MR-Egger method[J]. Eur J Epidemiol. 2017;32(5):377–89.
Shi Q, Wang Q, Wang Z, et al. Systemic inflammatory regulators and proliferative diabetic retinopathy: a bidirectional mendelian randomization study[J]. Front Immunol. 2023;14:1088778.
Vestergaard P. Discrepancies in bone mineral density and fracture risk in patients with type 1 and type 2 diabetes–a meta-analysis[J]. Osteoporos Int. 2007;18(4):427–44.
Schwartz AV, Vittinghoff E, Bauer DC, et al. Association of BMD and FRAX score with risk of fracture in older adults with type 2 diabetes[J]. JAMA. 2011;305(21):2184–92.
Shan PF, Wu XP, Zhang H, et al. Age-related bone mineral density, osteoporosis rate and risk of vertebral fracture in mainland Chinese women with type 2 diabetes mellitus[J]. J Endocrinol Invest. 2011;34(3):190–6.
Karsenty G, Oury F. Biology without walls: the novel endocrinology of bone[J]. Annu Rev Physiol. 2012;74:87–105.
Javed F, Yu W, Thornton J, et al. Effect of fat on measurement of bone mineral density[J]. Int J Body Compos Res. 2009;7(1):37–40.
Kim MW, Lee DH, Huh JW, et al. The impact of obesity on the accuracy of DXA BMD for DXA-equivalent BMD estimation[J]. BMC Musculoskelet Disord. 2022;23(1):1130.
Zhang Z, Zhang Z, Pei L, et al. How high-fat diet affects bone in mice: a systematic review and meta-analysis[J]. Obes Rev. 2022;23(10):e13493.
Kanabrocki EL, Hermida RC, Wright M, et al. Circadian variation of serum leptin in healthy and diabetic men[J]. Chronobiol Int. 2001;18(2):273–83.
Hamrick MW, Della-Fera MA, Choi YH, et al. Leptin treatment induces loss of bone marrow adipocytes and increases bone formation in leptin-deficient ob/ob mice[J]. J Bone Miner Res. 2005;20(6):994–1001.
Steppan CM, Crawford DT, Chidsey-Frink KL, et al. Leptin is a potent stimulator of bone growth in ob/ob mice[J]. Regul Pept. 2000;92(1–3):73–8.
Gordeladze JO, Drevon CA, Syversen U, et al. Leptin stimulates human osteoblastic cell proliferation, de novo collagen synthesis, and mineralization: impact on differentiation markers, apoptosis, and osteoclastic signaling[J]. J Cell Biochem. 2002;85(4):825–36.
Cornish J, Callon KE, Bava U, et al. Leptin directly regulates bone cell function in vitro and reduces bone fragility in vivo[J]. J Endocrinol. 2002;175(2):405–15.
Thommesen L, Stunes AK, Monjo M, et al. Expression and regulation of resistin in osteoblasts and osteoclasts indicate a role in bone metabolism[J]. J Cell Biochem. 2006;99(3):824–34.
Luo XH, Guo LJ, Xie H, et al. Adiponectin stimulates RANKL and inhibits OPG expression in human osteoblasts through the MAPK signaling pathway[J]. J Bone Miner Res. 2006;21(10):1648–56.
Oshima K, Nampei A, Matsuda M, et al. Adiponectin increases bone mass by suppressing osteoclast and activating osteoblast[J]. Biochem Biophys Res Commun. 2005;331(2):520–6.
Weyer C, Funahashi T, Tanaka S, et al. Hypoadiponectinemia in obesity and type 2 diabetes: close association with insulin resistance and hyperinsulinemia[J]. J Clin Endocrinol Metab. 2001;86(5):1930–5.
Liu L, Shi Z, Ji X, et al. Adipokines, adiposity, and atherosclerosis[J]. Cell Mol Life Sci. 2022;79(5):272.
Tai TY, Chen CL, Tsai KS, et al. A longitudinal analysis of serum adiponectin levels and bone mineral density in postmenopausal women in Taiwan[J]. Sci Rep. 2022;12(1):8090.
Pun KK, Lau P, Ho PW. The characterization, regulation, and function of insulin receptors on osteoblast-like clonal osteosarcoma cell line[J]. J Bone Miner Res. 1989;4(6):853–62.
Mohan S, Kesavan C. Role of insulin-like growth factor-1 in the regulation of skeletal growth[J]. Curr Osteoporos Rep. 2012;10(2):178–86.
Langlois JA, Rosen CJ, Visser M, et al. Association between insulin-like growth factor I and bone mineral density in older women and men: the Framingham Heart Study[J]. J Clin Endocrinol Metab. 1998;83(12):4257–62.
Zhao G, Monier-Faugere MC, Langub MC, et al. Targeted overexpression of insulin-like growth factor I to osteoblasts of transgenic mice: increased trabecular bone volume without increased osteoblast proliferation[J]. Endocrinology. 2000;141(7):2674–82.
Petersen MC, Shulman GI. Mechanisms of insulin action and insulin resistance[J]. Physiol Rev. 2018;98(4):2133–223.
Pramojanee SN, Phimphilai M, Chattipakorn N, et al. Possible roles of insulin signaling in osteoblasts[J]. Endocr Res. 2014;39(4):144–51.
Fulzele K, Riddle RC, DiGirolamo DJ, et al. Insulin receptor signaling in osteoblasts regulates postnatal bone acquisition and body composition[J]. Cell. 2010;142(2):309–19.
Thrailkill K, Bunn RC, Lumpkin CJ, et al. Loss of insulin receptor in osteoprogenitor cells impairs structural strength of bone[J]. J Diabetes Res. 2014;2014:703589.
Zoch ML, Abou DS, Clemens TL, et al. In vivo radiometric analysis of glucose uptake and distribution in mouse bone[J]. Bone Res. 2016;4:16004.
Wu S, Zhang Y, De Luca F. The effect of a high-calorie diet on bone growth is mediated by the insulin receptor[J]. Bone. 2019;122:166–75.
Fulzele K, DiGirolamo DJ, Liu Z, et al. Disruption of the insulin-like growth factor type 1 receptor in osteoblasts enhances insulin signaling and action[J]. J Biol Chem. 2007;282(35):25649–58.
Wasnich RD, Benfante RJ, Yano K, et al. Thiazide effect on the mineral content of bone[J]. N Engl J Med. 1983;309(6):344–7.
Aung K, Htay T. Thiazide diuretics and the risk of hip fracture[J]. Cochrane Database Syst Rev, 2011(10):D5185.
Boquist L, Fahraeus B. The parathyroid glands of the Mongolian gerbil (Meriones unguiculatus)[J]. Pathol Eur. 1975;10(2):134–5.
Schoofs MW, Sturkenboom MC, van der Klift M, et al. HMG-CoA reductase inhibitors and the risk of vertebral fracture[J]. J Bone Miner Res. 2004;19(9):1525–30.
Pritchard JM, Giangregorio LM, Atkinson SA, et al. Association of larger holes in the trabecular bone at the distal radius in postmenopausal women with type 2 diabetes mellitus compared to controls[J]. Arthritis Care Res (Hoboken). 2012;64(1):83–91.
Pritchard JM, Giangregorio LM, Atkinson SA, et al. Changes in trabecular bone microarchitecture in postmenopausal women with and without type 2 diabetes: a two-year longitudinal study[J]. BMC Musculoskelet Disord. 2013;14:114.
Napoli N, Chandran M, Pierroz DD, et al. Mechanisms of diabetes mellitus-induced bone fragility[J]. Nat Rev Endocrinol. 2017;13(4):208–19.
Chen H, Zhou X, Fujita H, et al. Age-related changes in trabecular and cortical bone microstructure[J]. Int J Endocrinol. 2013;2013:213234.
O’Gradaigh D, Debiram I, Love S, et al. A prospective study of discordance in diagnosis of osteoporosis using spine and proximal femur bone densitometry[J]. Osteoporos Int. 2003;14(1):13–8.
Aoki TT, Grecu EO, Srinivas PR, et al. Prevalence of osteoporosis in women: variation with skeletal site of measurement of bone mineral density[J]. Endocr Pract. 2000;6(2):127–31.
Blank RD, Malone DG, Christian RC, et al. Patient variables impact lumbar spine dual energy X-ray absorptiometry precision[J]. Osteoporos Int. 2006;17(5):768–74.
Koromani F, Ghatan S, van Hoek M, et al. Type 2 diabetes Mellitus and vertebral fracture risk. Curr Osteoporos Rep. 2021;19(1):50–7.
Sun A, Hu J, Wang S, et al. Association of the visceral adiposity index with femur bone mineral density and osteoporosis among the U.S. older adults from NHANES 2005–2020: a cross-sectional study. Front Endocrinol (Lausanne). 2023;14:1231527.
Acknowledgements
We express our deepest gratitude to the following individuals for their invaluable contributions to this project: Conceptualization: JB.G. and KT.Y. Methodology: JB.G. and T.L. Software: JB.G. and T.L. Validation: H.C., JB.G., and KT.Y. Formal Analysis: JB.G. and T.L. Investigation: JB.G. and H.C. Resources: H.C. Data Curation: JB.G. Writing—Original Draft Preparation: JB.G. and T.L. Writing—Review and Editing: JB.G. and T.L. Visualization: KT.Y. Supervision: KT.Y. Project Administration: JB.G. We would like to acknowledge the collaborative efforts of these individuals, whose expertise and dedication significantly contributed to the successful completion of this work. Their collective commitment to excellence has been instrumental in shaping the final manuscript.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
Conceptualization, JB.G.and KT.Y.; methodology, JB.G. and T.L.; software, JB.G.and T.L.; validation, H.C., JB.G. and KT.Y.; formal analysis, JB.G. and T.L.; investigation, JB.G.and H.C.; resources, H.C.; data curation, JB.G.; writing—original draft preparation, JB.G. and T.L.; writing—review and editing, JB.G. and T.L.; visualization, KT.Y.; supervision, KT.Y.; project administration, JB.G. JB.G. is the first author and KT.Y. is the corresponding author. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Ethical approval and consent to participate
Not applicable.
Consent for publication
The data were sourced from a publicly available database and no human participants involved; hence ethical parameters were not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Guan, J., Liu, T., Chen, H. et al. Association of type 2 Diabetes Mellitus and bone mineral density: a two-sample Mendelian randomization study. BMC Musculoskelet Disord 25, 130 (2024). https://doi.org/10.1186/s12891-024-07195-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12891-024-07195-6