
Abstract
Background
Primary sclerosing cholangitis (PSC) is a complex disease with pathogenic mechanisms that remain to be elucidated. Previous observational studies with small sample sizes have reported associations between PSC, dyslipidemia, and gut microbiota dysbiosis. However, the causality of these associations is uncertain, and there has been no systematic analysis to date.
Methods
The datasets comprise data on PSC, 179 lipid species, and 412 gut microbiota species. PSC data (n = 14,890) were sourced from the International PSC Study Group, while the dataset pertaining to plasma lipidomics originated from a study involving 7174 Finnish individuals. Data on gut microbiota species were derived from the Dutch Microbiome Project study, which conducted a genome-wide association study involving 7738 participants. Furthermore, we employed a two-step Mendelian randomization (MR) analysis to quantify the proportion of the effect of gut microbiota-mediated lipidomics on PSC.
Results
Following a rigorous screening process, our MR analysis revealed a causal relationship between higher levels of gene-predicted Phosphatidylcholine (O-16:1_18:1) (PC O-16:1_18:1) and an increased risk of developing PSC (inverse variance-weighted method, odds ratio (OR) 1.30, 95% confidence interval (CI) 1.03–1.63). There is insufficient evidence to suggest that gene-predicted PSC impacts the levels of PC O-16:1_18:1 (OR 1.01, 95% CI 0.98–1.05). When incorporating gut microbiota data into the analysis, we found that Eubacterium rectale-mediated genetic prediction explains 17.59% of the variance in PC O-16:1_18:1 levels.
Conclusion
Our study revealed a causal association between PC O-16:1_18:1 levels and PSC, with a minor portion of the effect mediated by Eubacterium rectale. This study aims to further explore the pathogenesis of PSC and identify promising therapeutic targets. For patients with PSC who lack effective treatment options, the results are encouraging.
Similar content being viewed by others

Background
Primary sclerosing cholangitis (PSC) is a rare and chronic cholestatic liver disease characterized by intrahepatic or extrahepatic stricturing, or both, accompanied by bile duct fibrosis [1]. Its prevalence is highest in northern Europe and lower in Asia, with about 16.2 cases per 100,000 people [2]. The causes of PSC remain unknown, and there are limited treatment options, leading to a median survival time of 13.2 years post-diagnosis before liver transplantation or mortality [3].
Recent studies have increasingly focused on dyslipidemia in patients with PSC. Jorgensen et al. highlighted the prevalence of hypercholesterolemia in PSC, which worsens with disease severity [4]. Additionally, Gandelman et al. proposed a disease-specific mechanism for dyslipidemia in PSC, based on a case study linking lipid abnormalities to hepatic impairment [5]. In parallel, studies with relatively small sample sizes (n < 30) have also explored lipidomic alterations in patients with PSC compared to healthy individuals [6, 7]. Inevitably, the aforementioned observational studies have limitations such as small sample sizes, residual confounding, and reverse causality, which may hinder causal assessment of the relationship between plasma lipidomics and PSC. For instance, Stokkeland K et al. had shown that administering lipid-lowering medications resulted in beneficial outcomes for PSC patients [8]. Consequently, a contentious debate persists regarding the etiological role of lipid abnormalities in PSC, prompting inquiry into whether these perturbations are foundational to the disease pathology or consequential outcomes thereof.
In recent years, modern and efficient lipidomics technologies have significantly expanded our knowledge of the diversity and breadth of circulating lipids. Lipid species encompass a wide range, including glycerophospholipids, glycerolipids, sphingolipids, and sterols [9]. Genome-wide association studies (GWAS) have transformed our comprehension of the genetic variations influencing lipid levels [10, 11]. Consequently, a study design that minimizes biases is essential to more convincingly establish the causal relationship between lipidomics and PSC.
The intestinal microbiota consists of approximately 4 × 1013 commensal bacteria, also known as the “human second genome” [12]. Recently, with the emergence of the “leaky gut” hypothesis and the “PSC microbiome” hypothesis, the gut microbiota has attracted increasing attention as a potential pathogenic factor for PSC [13,14,15]. Not only that, but changes in blood lipids have been found to be closely related to the abundance of the gut microbiota [16,17,18,19]. Consequently, the gut microbiota composition might serve as potential mediators between lipidomics and PSC.
Mendelian randomization (MR) is a potential causal inference method that utilizes genetic variations as instrumental variables to infer the impact of exposure factors on outcomes from observational data [20]. MR can mitigate the effects of unmeasured confounders or biases while leveraging Mendelian inheritance patterns to avoid reverse causation [20]. In this study, we collected recently published summary statistics from GWAS for 149 lipid species, 412 gut microbiota species, and PSC. Through the application of the mediation MR analysis, our primary goal was to unveil the causal relationships among these variables, shed light on the pathogenesis of PSC, and identify promising therapeutic targets.
Methods
Study design
The data utilized in our analysis were publicly available and had been approved by the institutional review committees of the respective studies. Hence, no ethical committee review was necessary for this study. Furthermore, all findings generated are detailed in the article and its supplementary materials.
In this study, we explored the causal relationship between plasma lipid species and PSC by bidirectional MR. In addition, the mediating roles played by plasma metabolites were explored. In our study, single nucleotide polymorphisms (SNPs) were defined as instrumental variables (IVs) [21].
GWAS summary data sources
The data utilized in our study are publicly available, with detailed information provided in Table 1. Specifically, the PSC dataset was obtained from the International PSC Study Group by Sun-Gou Ji et al., focusing on the correlation between PSC quantity and SNPs. This is the largest GWAS dataset on PSC so far, with 2,871 PSC patients and 12,019 controls included [22]. The dataset concerning the plasma lipidomics originated from a study involving 7174 Finnish individuals. This investigation encompassed univariate and multivariate whole-genome analyses, encompassing 179 lipid species (Table S1). Through precise locus mapping and gene prioritization, associations with diseases were explored among 377,277 participants in FinnGen [9]. The data on 412 gut microbiota species came from the Dutch Microbiome Project study. In this research, a GWAS was conducted on a population of 7,738 participants, covering 207 microbial taxa and 205 pathways representing microbial composition and function [23]. It is noteworthy that all GWAS data were sourced from various consortia or organizations, ensuring no sample overlap.
Instrumental variable selection and data harmonization
In our analysis, we incorporated SNPs that exhibited genome-wide significance (P < 1 × 10− 5). Subsequently, these SNPs were grouped based on linkage disequilibrium (using a window size of 10,000 kb and r2 < 0.001). Moreover, palindromic and ambiguous SNPs were omitted from the IVs for the MR analysis [24]. The F statistic was computed by evaluating the variance explained by the SNPs for each exposure, calculated as [(N – K – 1)/K]/ [R2/ (1 – R2)], where K represents the number of genetic variants and N denotes the sample size. We excluded weak IVs (F-statistics < 10) from the analysis to ensure robustness and reliability of the results [25].
Statistical analysis
We conducted MR analysis using R software (version 4.2.0, http://www.r-project.org) in conjunction with the “Two-Sample MR” package (version 0.5.6) for precise and comprehensive analysis [26]. Moreover, the online tool PhenoScanner was used to assess all known phenotypes related to the considered genetic instruments in our analyses (http://www.phenoscanner.medschl.cam.ac.uk/).
Primary analysis
Figure 1A presents a schematic overview of our analysis, where we conducted a two-sample bidirectional MR study to explore the reciprocal causation between the plasma lipidomics and PSC. The Inverse variance weighting (IVW) method utilized meta-analysis to combine the Wald ratios of causal effects for each SNP [24, 27]. In addition to IVW, we employed supplementary methods such as Bayesian Weighted MR (BWMR) [28] and Weighted-Median [29] methods. These diverse methodologies, tailored to different validity assumptions, were employed to derive MR estimates. More specifically, IVW relies on the assumption that all SNPs function as valid instrumental variables, ensuring precise estimations. BWMR, designed for causal inference, addresses challenges by considering uncertainties stemming from weak effects due to polygenicity and by detecting outliers using Bayesian weighting to handle violations of the IV assumption caused by pleiotropy. The Weighted Median method demonstrates superior precision, as indicated by its smaller standard deviation compared to MR-Egger analysis. Even in the presence of horizontal pleiotropy, the Weighted Median method offers a consistent estimate, even when 50% of genetic variants are considered invalid instrumental variables [30].

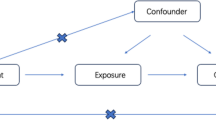
Diagrams illustrating associations examined in this study. (A) The total effect between plasma lipidomics and primary sclerosing cholangitis (PSC). c is the total effect using genetically predicted plasma lipidomics as exposure and PSC as outcome. d is the total effect using genetically predicted PSC as exposure and plasma lipidomics as outcome. (B) The total effect was decomposed into: (i) indirect effect using a two-step approach (where a is the total effect of plasma lipidomics on gut microbiota species, and b is the effect of gut microbiota species on PSC) and the product method (a × b) and (ii) direct effect (c′ = c – a × b). Proportion mediated was the indirect effect divided by the total effect
Mediation analysis
To investigate the potential role of gut microbiota species in mediating the causal relationship between the plasma lipidomics and PSC, we conducted mediation analyses employing a two-step MR design (Fig. 1B). The total effect (c in Fig. 1A) can be decomposed into an indirect effect (a × b in Fig. 1B) and a direct effect (c’ in Fig. 1B) [31]. We calculated the percentage mediated by the mediating effect by dividing the indirect effect by the total effect.
which was designated as the total effect.
Sensitivity analysis
Due to variations in experimental conditions, selected populations, and SNPs, heterogeneity may exist in two-sample MR analyses, potentially leading to biased estimates of causal effects.
Consequently, this study conducted heterogeneity tests for the primary IVW and MR-Egger methods. Cochrane’s Q value was utilized to assess the heterogeneity of the genetic instruments, with a P-value > 0.05 indicating no significant heterogeneity. Furthermore, an underlying assumption of MR analysis is that IVs solely influence the outcome through exposure, necessitating an examination for horizontal pleiotropy between exposure and outcome [32]. In this study, the MR-Egger intercept method was employed to assess the presence of pleiotropy. A P-value > 0.05 suggests a minimal or negligible likelihood of pleiotropy in the causal analysis, allowing for its exclusion. Finally, leave-one-out analysis was used to validate the consistency of the results [33].
Results
The workflow diagram for this study was shown in Fig. 2.

The workflow diagram for this study. PSC, Primary sclerosing cholangitis; IVW, Inverse variance weighting; MR, Mendelian randomization; Eubacterium rectale, k_Bacteria.p_Firmicutes.c_Clostridia.o_Clostridiales.f_Eubacteriaceae.g_Eubacterium.s_Eubacterium_rectale.
Association of 179 lipid species and PSC
To investigate the genetically predicted causal relationship between the plasma lipidomics and PSC, we treated 179 lipid species as exposures and PSC as the outcome. In this study, the IVW analysis method was used to identify 13 lipid species that may be causally related to PSC, with the exclusion of Phosphatidylcholine (O-16:1_18:2) levels and Triacylglycerol (49:1) levels due to the presence of horizontal pleiotropy (Table S3). Among these 13 lipid species, we only found one that has a positive genetically predicted causal relationship with PSC, which is Phosphatidylcholine (O-16:1_18:1) levels (PC O-16:1_18:1 levels) (IVW odds ratio [OR] per SD increase in PSC = 1.30 [95% CI, 1.03–1.63], P = 0.025; Weighted median OR per SD increase in PSC = 1.38 [95% CI, 1.01–1.88], P = 0.046; BWMR OR per SD increase in PSC = 1.31 [95% CI, 1.02–1.67], P = 0.032) (Fig. 3). Notably, a large number of Triacylglycerol-related lipidomics were found to be negatively correlated with PSC (Table S3).

Forest plot to visualize the causal effects of Eubacterium rectale with PC O-16:1_18:1 levels and PSC. PSC, Primary sclerosing cholangitis; PC O-16:1_18:1 levels, Phosphatidylcholine (O-16:1_18:1) levels; Eubacterium rectale, k_Bacteria.p_Firmicutes.c_Clostridia.o_Clostridiales.f_Eubacteriaceae.g_Eubacterium.s_Eubacterium_rectale.
In our MR analysis, no indication of reverse causality was detected between genetically predicted PC O-16:1_18:1 levels and PSC, with an odds ratio of 1.01 [95% CI, 0.98–1.05, P = 0.729] using the IVW method, as illustrated in Fig. 3. The characteristics of SNPs associated with PC O-16:1_18:1 levels and PSC in the bidirectional MR analysis are extensively documented in the supplementary files (Table S4, S5).
Association of 412 gut microbiota species with PSC
To analyze the data more effectively, we initially investigated the genetically predicted causal relationships between 142 gut microbiota species and PSC by IVW method. Following preliminary screening, a total of 17 gut microbiota species have been identified as potentially causally linked to PSC based on genetically predicted associations (Table S6), pending further scrutiny for heterogeneity and pleiotropy.
Association of PC O-16:1_18:1 levels with 17 gut microbiota species
In the aforementioned two-sample MR analysis, we have identified the genetically predicted causal relationship between PC O-16:1_18:1 levels and PSC, as well as preliminarily screened a total of 17 gut microbiota species that may potentially be causally related to PSC.
In the upcoming analysis, we will further explore the genetically predicated causal relationships between PC O-16:1_18:1 levels and the 17 gut microbiota species.
Following an initial screening using IVW, potential causal relationships were identified between PC O-16:1_18:1 levels and 3 gut microbiota species. After a comprehensive analysis, it was revealed that PC O-16:1_18:1 levels exhibit a negative causal relationship with FERMENTATION.PWY.mixed.acid.fermentation, while showing a positive causal association with LACTOSECAT.PWY.lactose.and.galactose.degradation.I and k_Bacteria.p_Firmicutes.c_Clostridia.o_Clostridiales.f_Eubacteriaceae.g_Eubacterium.s_Eubacterium_rectale (abbreviated as Eubacterium rectale), with no evidence of horizontal pleiotropy and heterogeneity (Table S7).
Association of 3 gut microbiota species with PSC
After rigorous screening, a total of 3 gut microbiota species have been included in the candidate list. In the subsequent work, we will focus on analyzing the genetically predicated causal relationships between these 3 gut microbiota species and PSC (Table S8).
The gut microbiota species FERMENTATION.PWY.mixed.acid.fermentation and LACTOSECAT.PWY.lactose.and.galactose.degradation.I, acting as mediators, exhibit opposing correlations between PC O-16:1_18:1 levels and PSC, hence excluded from the study.
Ultimately, we confirmed a positive genetically predicated causal relationship between Eubacterium rectale and PSC through MR analysis (IVW OR per SD increase in PSC = 1.46 [95% CI, 1.02–2.10], P = 3.91E-02; BWMR OR per SD increase in PSC = 1.48 [95% CI, 1.01–2.17], P = 4.67E-02), successfully passing tests for heterogeneity, horizontal pleiotropy, and sensitivity analysis (Fig. 3). Although Weighted median method failed to yield positive results, the overall direction was consistent.
Proportion of the association between PC O-16:1_18:1 levels and PSC mediated by Eubacterium rectale
In summary, we have ultimately discovered that Eubacterium rectale can act as a mediator in the pathway from PC O-16:1_18:1 levels to PSC. We observed an association between elevated PC O-16:1_18:1 levels and increased Eubacterium rectale (Table S9), which, in turn, were correlated with an elevated risk of PSC (Table S10). As illustrated in Fig. 1B, our study demonstrated that the indirect effect was 0.05, constituting 17.59% of the heightened risk of PSC associated with PC O-16:1_18:1 levels, while the direct effect was 0.22 (Fig. 4).

Schematic diagram of the Eubacterium rectale mediation effect. PSC, Primary sclerosing cholangitis; PC O-16:1_18:1 levels, Phosphatidylcholine (O-16:1_18:1) levels Eubacterium rectale, k_Bacteria.p_Firmicutes.c_Clostridia.o_Clostridiales.f_Eubacteriaceae.g_Eubacterium.s_Eubacterium_rectale.
Sensitivity analysis
In the above analysis, we utilized the online tool PhenoScanner to analyze each SNP and found no association between them and the outcome under investigation. Simultaneously, a “leave-one-out” approach was utilized for sensitivity analysis to explore whether specific SNPs influenced the causal relationships. The findings revealed that systematically excluding each SNP did not lead to significant changes in the model’s effect estimates or qualitative conclusions (Supplementary Figure S1, S2, S3, S4). In the MR analysis conducted, tests for heterogeneity and horizontal pleiotropy were performed, with parameters failing to meet the three key assumptions being excluded, ensuring the high credibility of the study results (Table S11).
Discussion
Most lipids detected in human serum or plasma remain stable and are well correlated with the liver lipidome [34]. Thus, the circulating lipidome is an attractive source of biomarkers for hepatobiliary diseases [35,36,37]. Previously, studies with relatively limited sample sizes (n < 30) have investigated lipidomic changes in patients with PSC compared to healthy individuals [6, 7, 38], but the results were limited by the influence of various confounding factors. Based on our current understanding, this study represents the first comprehensive assessment of the causal relationship between plasma lipidomes and PSC. Previous research has already unveiled the close connections among gut microbiota, PSC, and plasma lipidomes [13,14,15,16,17,18,19]. However, existing evidence is confined to observational studies, susceptible to various confounding factors. Leveraging existing GWAS data, this study innovatively employs MR analysis to link these three elements, aiming to demonstrate the causal relationship between PSC and plasma lipidomes. By incorporating GWAS data on gut microbiota, the study explores their potential mediating role.
In this study, our findings suggest that genetically predicted PC O-16:1_18:1 levels were associated with an increased risk of PSC (30% increased risk of PSC for every 1 SD increase in PC O-16:1_18:1 levels), and 17.59% of this effect was mediated through Eubacterium rectale levels in the gut. Furthermore, the MR analyses carried out in this study revealed no indications of horizontal pleiotropy or heterogeneity, and sensitivity analyses also affirmed the robustness of the findings.
PSC is closely related to the intestine. With the emergence of the “leaky gut” hypothesis and the “PSC microbiome” hypothesis, the gut microbiota as a potential pathogenic factor for PSC has received increasing attention [13, 15, 16]. In this study, we identified that the gut microbe Eubacterium rectale can act as a mediator to facilitate the occurrence and progression of PSC by mediating PC O-16:1_18:1. Similarly, Alfonso et al. discovered a positive correlation between plasma phosphatidylcholine and Eubacterium rectale through multi-omics analysis, which aligns with our findings [39]. Furthermore, Wolfgang et al. proposed the discovery of a novel pathway for phosphatidylcholine to be transported from endogenous sources through mucosal cell gaps and across tight junction barrier to the intestinal lumen surface [40]. Phosphatidylcholine in intestinal mucus binds to membrane-bound mucin 3, which is then transferred to mucin 2 to establish a hydrophobic mucus barrier against the colonic luminal microbial community, playing a crucial role in maintaining intestinal microbial homeostasis [40]. However, in the case of tight junction disruption, plasma phosphatidylcholine cannot be transferred to intestinal mucus, leading to elevated levels in the blood and decreased levels in intestinal mucus. This imbalance results in gut dysbiosis, toxin absorption, and the induction of ulcerative colitis [41,42,43]. Approximately 5% of ulcerative colitis patients progress to PSC, explaining why the majority of PSC patients (70%) also have ulcerative colitis [1].
Choline and its derivatives are not only essential components of cell membranes but also play a crucial role in the transport and metabolism of lipid cholesterol [44]. Furthermore, abnormalities in phospholipid metabolism may impact various biological processes, such as inflammation [45]. In this study, we found that genetic predicted plasma PC O-16:1_18:1 levels have a causal relationship with PSC. In line with the serum metabolomics analysis of a small sample population conducted by Jesus M et al., elevated levels of phosphatidylcholine were observed in the serum of patients with PSC [6]. Interestingly, as phosphatidylcholine serves as a crucial component of bile, a separate study reported a decrease in phosphatidylcholine levels in the bile of PSC patients [46]. The biliary epithelium, akin to other mucosal surfaces, is regarded as a protective surface that secretes mucus to maintain its integrity [47]. It employs phosphatidylcholine bound to mucins produced locally to preserve its hydrophobic nature [48]. Likewise, when tight junction is compromised, the inability of plasma phosphatidylcholine to transfer to biliary mucus results in reduced phosphatidylcholine levels within the bile ducts, consequently triggering the onset of local chronic inflammation [40, 49]. This may be one of the reasons for the direct effect.
This study has several limitations. Firstly, the threshold for significant genome-wide SNPs is typically set at P < 5 × 10 − 8. However, in this study, a threshold of P < 1 × 10 − 5 was used, which may increase the likelihood of discovering statistically significant SNPs but also raise the risk of false positives. Secondly, while all the analytical results of this study meet the three major assumptions of MR analysis, the overall statistical significance is relatively weak. Additionally, differences in lipidomics and microbiomics between Finnish and Dutch populations may introduce confounding effects related to regional and dietary factors. Dietary habits vary across regions and can influence the composition of the gut microbiota, thereby affecting the associations between plasma lipids and gut bacteria [50, 51]. PSC, as a rare disease, also exhibits variations in incidence rates across different regions [52]. Biases resulting from population differences could potentially impact the results. Finally, the findings of this study warrant validation using larger sample sizes.
Conclusion
In conclusion, our study revealed a causal association between PC O-16:1_18:1 levels and PSC, with a minor portion of the effect mediated by Eubacterium rectale. The underlying mechanisms may involve the transport pathway of PC mediated by tight junctions in biliary and intestinal epithelial cells. This study aims to further explore the pathogenesis of PSC and identify promising therapeutic targets. For patients with PSC who lack effective treatment options, the results are encouraging.
Data availability
Data are available from the corresponding author upon reasonable request.
Abbreviations
- PSC:
-
Primary Sclerosing Cholangitis
- MR:
-
Mendelian Randomization
- PC O-16:1_18:1:
-
Phosphatidylcholine (O-16:1_18:1)
- OR:
-
Odds Ratio
- CI:
-
Confidence Interval
- GWAS:
-
Genome-Wide Association Studies
- SNPs:
-
Single Nucleotide Polymorphisms
- IVs:
-
Instrumental Variables
- IVW:
-
Inverse Variance Weighting
- BWMR:
-
Bayesian Weighted Mendelian Randomization
- k_Bacteria.p_Firmicutes.c_Clostridia.o_Clostridiales.f_Eubacteriaceae.g_Eubacterium.s_Eubacterium_rectale:
-
Eubacterium rectale
References
Dyson JK, Beuers U, Jones DEJ, Lohse AW. Hudson M. Primary sclerosing cholangitis. Lancet. 2018;391:2547–59.
Boonstra K, Beuers U, Ponsioen CY. Epidemiology of primary sclerosing cholangitis and primary biliary cirrhosis: a systematic review. J Hepatol. 2012;56:1181–88.
Boonstra K, Weersma RK, van Erpecum KJ, et al. Population-based epidemiology, malignancy risk, and outcome of primary sclerosing cholangitis. Hepatology. 2013;58:2045–55.
Jorgensen RA, Lindor KD, Sartin JS, LaRusso NF, Wiesner RH. Serum lipid and fat-soluble vitamin levels in primary sclerosing cholangitis. J Clin Gastroenterol. 1995;20:215–9.
Gandelman G, Aronow WS, Weiss MB. Resolving hyperlipidemia after liver transplantation in a patient with primary sclerosing cholangitis. Am J Ther. 2006;13:171–4.
Banales JM, Iñarrairaegui M, Arbelaiz A, et al. Serum metabolites as diagnostic biomarkers for Cholangiocarcinoma, Hepatocellular Carcinoma, and primary sclerosing Cholangitis. Hepatology. 2019;70:547–62.
Bell LN, Wulff J, Comerford M, Vuppalanchi R, Chalasani N. Serum metabolic signatures of primary biliary cirrhosis and primary sclerosing cholangitis. Liver Int. 2015;35:263–74.
Stokkeland K, Höijer J, Bottai M, Söderberg-Löfdal K, Bergquist A. Statin use is Associated with Improved outcomes of patients with primary sclerosing Cholangitis. Clin Gastroenterol Hepatol. 2019;17:1860–e661.
Ottensmann L, Tabassum R, Ruotsalainen SE, et al. Genome-wide association analysis of plasma lipidome identifies 495 genetic associations. Nat Commun. 2023;14:6934.
Graham SE, Clarke SL, Wu KH, et al. The power of genetic diversity in genome-wide association studies of lipids. Nature. 2021;600:675–79.
Surakka I, Horikoshi M, Mägi R, et al. The impact of low-frequency and rare variants on lipid levels. Nat Genet. 2015;47:589–97.
Grice EA, Segre JA. The human microbiome: our second genome. Annu Rev Genomics Hum Genet. 2012;13:151–70.
Tabibian JH, O’Hara SP, Lindor KD. Primary sclerosing cholangitis and the microbiota: current knowledge and perspectives on etiopathogenesis and emerging therapies. Scand J Gastroenterol. 2014;49:901–8.
Tabibian JH, Talwalkar JA, Lindor KD. Role of the microbiota and antibiotics in primary sclerosing cholangitis. Biomed Res Int. 2013;2013:389537.
Yang J, Ma G, Wang K, et al. Causal associations between gut microbiota and cholestatic liver diseases: a mendelian randomization study. Front Med (Lausanne). 2024;11:1342119.
Miyajima Y, Karashima S, Ogai K, et al. Impact of gut microbiome on dyslipidemia in Japanese adults: Assessment of the Shika-Machi super preventive health examination results for causal inference. Front Cell Infect Microbiol. 2022;12:908997.
Wan X, Li T, Liu D et al. Effect of Marine Microalga Chlorella pyrenoidosa ethanol extract on lipid metabolism and gut microbiota composition in High-Fat Diet-Fed rats. Mar Drugs. 2018; 16.
Panyod S, Wu WK, Hu MY, et al. Healthy diet intervention reverses the progression of NASH through gut microbiota modulation. Microbiol Spectr. 2024;12:e0186823.
He X, Chen D, Guo Y, et al. Walnut Meal extracts Rich in polyphenols mitigate insulin resistance and modulate gut microbiota in high Fat Diet-Fed rats. J Med Food. 2022;25:618–29.
Emdin CA, Khera AV, Kathiresan S. Mendelian Randomization Jama. 2017;318:1925–26.
Davey Smith G, Hemani G. Mendelian randomization: genetic anchors for causal inference in epidemiological studies. Hum Mol Genet. 2014;23:R89–98.
Ji SG, Juran BD, Mucha S, et al. Genome-wide association study of primary sclerosing cholangitis identifies new risk loci and quantifies the genetic relationship with inflammatory bowel disease. Nat Genet. 2017;49:269–73.
Lopera-Maya EA, Kurilshikov A, van der Graaf A, et al. Effect of host genetics on the gut microbiome in 7,738 participants of the Dutch Microbiome Project. Nat Genet. 2022;54:143–51.
Hemani G, Zheng J, Elsworth B et al. The MR-Base platform supports systematic causal inference across the human phenome. Elife. 2018; 7.
Burgess S, Thompson SG. Avoiding bias from weak instruments in mendelian randomization studies. Int J Epidemiol. 2011;40:755–64.
Broadbent JR, Foley CN, Grant AJ, et al. MendelianRandomization v0.5.0: updates to an R package for performing mendelian randomization analyses using summarized data. Wellcome Open Res. 2020;5:252.
Burgess S, Butterworth A, Thompson SG. Mendelian randomization analysis with multiple genetic variants using summarized data. Genet Epidemiol. 2013;37:658–65.
Zhao J, Ming J, Hu X, et al. Bayesian weighted mendelian randomization for causal inference based on summary statistics. Bioinformatics. 2020;36:1501–08.
Bowden J, Davey Smith G, Haycock PC, Burgess S. Consistent estimation in mendelian randomization with some Invalid instruments using a weighted median estimator. Genet Epidemiol. 2016;40:304–14.
Zhang Y, Liu Z, Choudhury T, Cornelis MC, Liu W. Habitual coffee intake and risk for nonalcoholic fatty liver disease: a two-sample mendelian randomization study. Eur J Nutr. 2021;60:1761–67.
Carter AR, Sanderson E, Hammerton G, et al. Mendelian randomisation for mediation analysis: current methods and challenges for implementation. Eur J Epidemiol. 2021;36:465–78.
van Kippersluis H, Rietveld CA. Pleiotropy-robust mendelian randomization. Int J Epidemiol. 2018;47:1279–88.
Yin KJ, Huang JX, Wang P, et al. No Genetic Causal Association between Periodontitis and Arthritis: a bidirectional two-sample mendelian randomization analysis. Front Immunol. 2022;13:808832.
Ooi GJ, Meikle PJ, Huynh K, et al. Hepatic lipidomic remodeling in severe obesity manifests with steatosis and does not evolve with non-alcoholic steatohepatitis. J Hepatol. 2021;75:524–35.
Satriano L, Lewinska M, Rodrigues PM, Banales JM, Andersen JB. Metabolic rearrangements in primary liver cancers: cause and consequences. Nat Rev Gastroenterol Hepatol. 2019;16:748–66.
Masoodi M, Gastaldelli A, Hyötyläinen T, et al. Metabolomics and lipidomics in NAFLD: biomarkers and non-invasive diagnostic tests. Nat Rev Gastroenterol Hepatol. 2021;18:835–56.
Paul B, Lewinska M, Andersen JB. Lipid alterations in chronic liver disease and liver cancer. JHEP Rep. 2022;4:100479.
Trottier J, Białek A, Caron P, et al. Metabolomic profiling of 17 bile acids in serum from patients with primary biliary cirrhosis and primary sclerosing cholangitis: a pilot study. Dig Liver Dis. 2012;44:303–10.
Benítez-Páez A, Kjølbæk L, Del Gómez EM et al. A multi-omics Approach to unraveling the microbiome-mediated effects of Arabinoxylan oligosaccharides in overweight humans. mSystems. 2019; 4.
Stremmel W, Staffer S, Gan-Schreier H, et al. Phosphatidylcholine passes through lateral tight junctions for paracellular transport to the apical side of the polarized intestinal tumor cell-line CaCo2. Biochim Biophys Acta. 2016;1861:1161–69.
Stremmel W, Staffer S, Schneider MJ, et al. Genetic mouse models with intestinal-specific tight Junction deletion resemble an Ulcerative Colitis phenotype. J Crohns Colitis. 2017;11:1247–57.
Ehehalt R, Wagenblast J, Erben G, et al. Phosphatidylcholine and lysophosphatidylcholine in intestinal mucus of ulcerative colitis patients. A quantitative approach by nanoelectrospray-tandem mass spectrometry. Scand J Gastroenterol. 2004;39:737–42.
Braun A, Treede I, Gotthardt D, et al. Alterations of phospholipid concentration and species composition of the intestinal mucus barrier in ulcerative colitis: a clue to pathogenesis. Inflamm Bowel Dis. 2009;15:1705–20.
Zeisel SH, da Costa KA. Choline: an essential nutrient for public health. Nutr Rev. 2009;67:615–23.
Feingold KR, Grunfeld C. The effect of inflammation and infection on lipids and lipoproteins. In: Feingold KR, Anawalt B, Blackman MR, Boyce A, Chrousos G, Corpas E, et al. editors. Endotext. South Dartmouth (MA): MDText.com, Inc. Copyright © 2000–2024. MDText.com, Inc.; 2000.
Gauss A, Ehehalt R, Lehmann WD, et al. Biliary phosphatidylcholine and lysophosphatidylcholine profiles in sclerosing cholangitis. World J Gastroenterol. 2013;19:5454–63.
Johansson ME, Sjövall H, Hansson GC. The gastrointestinal mucus system in health and disease. Nat Rev Gastroenterol Hepatol. 2013;10:352–61.
Lichtenberger LM. The hydrophobic barrier properties of gastrointestinal mucus. Annu Rev Physiol. 1995;57:565–83.
Stremmel W, Staffer S, Weiskirchen R. Phosphatidylcholine passes by Paracellular Transport to the apical side of the polarized biliary tumor cell line Mz-ChA-1. Int J Mol Sci. 2019; 20.
Gentile CL, Weir TL. The gut microbiota at the intersection of diet and human health. Science. 2018;362:776–80.
Qiao F, Tan F, Li LY, et al. Alteration and the function of Intestinal Microbiota in High-Fat-Diet- or Genetics-Induced lipid Accumulation. Front Microbiol. 2021;12:741616.
Mehta TI, Weissman S, Fung BM, et al. Global incidence, prevalence and features of primary sclerosing cholangitis: a systematic review and meta-analysis. Liver Int. 2021;41:2418–26.
Funding
This study was funded by the Jiangsu University Medical Education Collaborative Innovation Fund Project (No. JDY2023018) and the Medical Research Project of Jiangsu Health Commission (No. Z2021010).
Author information
Authors and Affiliations
Contributions
J. Z and D. Z were involved in the study concept and design; K. W, Y. X and C. C collected data and conducted analyses; J. Z wrote the draft of the article; D. Z revised the manuscript and had primary responsibility for final content. All authors reviewed and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This study was based on publicly available summary data and required no ethics approval or participant consent.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zhou, J., Zhu, D., Xu, Y. et al. Genetically predicted gut microbiota mediate the association between plasma lipidomics and primary sclerosing cholangitis. BMC Gastroenterol 24, 158 (2024). https://doi.org/10.1186/s12876-024-03246-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12876-024-03246-3