
Abstract
Background
The rapidly increasing availability of complete plastomes has revealed more structural complexity in this genome under different taxonomic levels than expected, and this complexity provides important evidence for understanding the evolutionary history of angiosperms. To explore the dynamic history of plastome structure across the subclass Alismatidae, we sampled and compared 38 complete plastomes, including 17 newly assembled, representing all 12 recognized families of Alismatidae.
Result
We found that plastomes size, structure, repeat elements, and gene content were highly variable across the studied species. Phylogenomic relationships among families were reconstructed and six main patterns of variation in plastome structure were revealed. Among these, the inversion from rbcL to trnV-UAC (Type I) characterized a monophyletic lineage of six families, but independently occurred also in Caldesia grandis. Three independent ndh gene loss events were uncovered across the Alismatidae. In addition, we detected a positive correlation between the number of repeat elements and the size of plastomes and IR in Alismatidae.
Conclusion
In our study, ndh complex loss and repeat elements likely contributed to the size of plastomes in Alismatidae. Also, the ndh loss was more likely related to IR boundary changes than the adaptation of aquatic habits. Based on existing divergence time estimation, the Type I inversion may have occurred during the Cretaceous-Paleogene in response to the extreme paleoclimate changes. Overall, our findings will not only allow exploring the evolutionary history of Alismatidae plastome, but also provide an opportunity to test if similar environmental adaptations result in convergent restructuring in plastomes.
Similar content being viewed by others

Backgroud
The subclass Alismatidae including 12 families and 500 species in ca. 54 genera, exhibits the greatest adaptive radiation of aquatic angiosperm in the world [1, 2]. Compared with the other two families (Araceae and Tofieldiaceae) of the order Alismatales, the group contains all hydrophytic life forms (submerged, floating-leaved, and emergent) and has a high concentration of water-pollinated plants [2, 3]. All marine flowering plants distributed in five families (Hydrocharitaceae, Cymodoceaceae, Ruppiaceae, Posidoniaceae, and Zosteraceae) belong to Alismatidae. Thus, Alismatidae can be considered as a key group for understanding the adaptive evolution of aquatic plants [4, 5].
Plastomes have been proven effective in solving plant phylogeny and evolution at different taxonomic levels due to their highly conserved and simple structure, and maternal inheritance [6, 7]. The recent explosion of novel plastome data has, however, revealed a large number of structural variations (SVs) in higher taxa (e.g., Malpighiales [8]; Fabaceae [9]; Araceae [10]) or microstructural changes between/within genera (e.g., Myriophyllum [11]; Monochoria [12]). Typical SVs include genomic rearrangements, gene loss, inversion and expansion/contraction of the inverted repeats (IRs) [13, 14]. For example, in the legume family (Fabaceae), many rearrangements were revealed and SVs were demonstrated to represent independent events in plastome evolution [9]. At the order level, Du et al. [15] made a plastome phylogeny of ferns and uncovered a variety of SVs supporting the deep phylogeny. In addition, plastid SVs can be related to life forms (e.g., hemiparasite [16]; phytoparasite [17]) or habitat adaptation (e.g., submerged [18]; alpine [19]). In parasitic plants, such as in some orchids [17] and Triuridaceae [20], the loss of photosynthesis-related genes has caused an extreme reduction in the size of plastomes and led to the marked reconfiguration of plastome structure. Iles et al. [21] and Peredo et al. [18] suggested that the loss of the ndh complex in aquatic angiosperms might explain the mechanism of reducing photooxidative stresses in submerged habitats. As well, SVs in plastomes may be associated with IR expansion/contraction, which in turn affects the plastome size [9]. The studies of Fu et al. [22, 23] revealed frequent gene loss (i.e., ndh complex) and plastome degradation in the genus Gentiana, which was suggested to parallel the diversification in this group. In Alismatidae, Ross et al. [24] found that the independent loss of the ndh complex in three families (Hydrocharitaceae, Cymodoceaceae, and Posidoniaceae) of Alismatidae may be associated with changes in plastome sizes [e.g., Thalassia hemprichii (loss of ndh complex; 178,261 bp) vs. Zostera marina (keep ndh complex; 143,877 bp)].
Previous studies have suggested that abundant repetitive elements, such as simple sequence repeats (SSRs), tandem repeats (TRs), and dispersed repeats, are commonly observed in rearranged plastomes and may be important factors driving the variation in plastome size [25]. Several studies have confirmed a correlation between repeat content with plastome size, for example, in Geraniaceae [26], Fabaceae [27], and Gymnosperms [28], or found that repeats promote rearrangements [29].
Until now, approximately 50 Alismatidae plastomes, representing ca. eight families and 20 genera, have been assembled and are available from GenBank (accessed on 1 January 2022). However, some genera have been extensively sampled (e.g., Ottelia [30]; Hydrocharis [31]), certain families lack accessible plastome resources. Thus, a comprehensive comparison of plastomes across Alismatidae remains scarce in the existing investigations.
In this study, we newly assembled 17 plastomes belonging to eight families in Alismatidae. We combined them with a set of existing complete plastomes to investigate the plastome structural evolution that covers all Alismatidae families, focusing on 1) exploring the pattern of SVs across Alismatidae plastomes; 2) detecting the correlations between plastome size and repetitive elements.
Results
Characteristics of Alismatidae plastomes
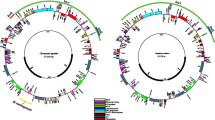
A total of 17 new plastomes representing six families were assembled and annotated here (Table S1). In combination with previously published data, the plastome size in Alismatidae displayed a size variation ranging from 143,877 bp (Zostera marina) to 179,007 bp (Sagittaria lichuanensis), with the normal quadripartite structure consisting of a large single copy (LSC, 80,881–99,125 bp), a small single copy (SSC, 2666–21,476 bp), and one pair of IRs (22,292–44,815 bp). Similarly, the GC content varied from 35.5% (Amphibolis antarctica and Z. marina) to 39.2% (T. hemprichii). All plastomes included 30 tRNA and four rRNA, whereas the number of PCGs varied from 68 to 79 (Table S1). This variation was explained by several gene loss events. The ndh complex was generally lost or pseudogenized in Najas and some marine species, including T. hemprichii, Halophila beccarii, A. antarctica, and Posidonia australis (Fig. 1). Also, two genes, comprised of rps19 and psbA, and one rps16 gene were lost in Z. marina and H. chevalieri, respectively.

Distribution of ndh gene loss, size of LSC and IR, and IR boundary across the phylogenetic tree in Alismatidae
IR extraction/expansion and structural variations
The junctions of SSC/IRa (JSA) and SSC/IRb (JSB) were compared to assess the IR extraction/expansion among the twelve families in Alismatidae. A total of 14 junction types were found. In the most common type I, the ycf1 was located on both the JSA and JSB. This type was found to be randomly distributed in various families (Fig. 1). The next most common types were II and III, where the JSA was located between ndhF and ndhH, and JSB was expanded into the ndhA with type II. In type II, JSA was placed in the ndhF-trnN intergenic spacer, and JSB was located in the ycf1. Nine more types were found independently distributed in nine genera (Fig. 1).
Six main types (I-VI) of SVs were detected, with A. gramineus as a reference (Fig. 2). In types I-IV, small-scale inversions occurred in LSC regions, three of which were anchored in the boundary of the accD gene. In type I, a small inversion from rbcL to trnV-UAC was located in the monophyletic lineage with six families and independently in Caldesia grandis. Type II was detected only in Triglochin maritima, which displayed ~ 8 kb inversion with nine PCGs, from accD to psbE. Moreover, accD gene inversion, named type III, was found in Aponogeton lakhonensis. Type IV was found in S. subulata, S. lichuanensis, and Limnophyton angolense in the family Alismataceae and the region trnQ-UUG + psbK + psbI + trnS-GCU was inverted. Type V was identified in the marine lineage of Hydrocharitaceae, where ycf2 with trnL-CAA was inverted in the IR region. The trnN-GUU gene inversion in type VI characterizes Caldesia grandis and H. dubia.

Structure variation of plastomes across the phylogeny of Alismatidae. Six types of inversion were detected: I) from rbcL to trnV-UAC; II) from accD to psbE; III) accD; IV) from trnQ-UUG to trnS-GCU; V) from ycf2 to trnL-CAA; VI) trnN-GUU. Bootstrap supports (BS) and posterior probability (PP) are represented at nodes, except nodes with 100% BS and 1.0 PP. The first two values showed unpartitioned data, and the last two were estimated from partitioned data
Repeat sequences in Alismatidae plastomes
Similarly to other plastome characteristics, a variable number of repeats were uncovered within Alismatidae plastomes (Fig. 3; Table S2). For SSRs, the numbers ranged from 108 (N. flexilis) to 241 (A. antarctica). The highest number of SSRs were detected in the family Zosteraceae (205–223), whereas the most variable numbers of SSRs were found in Cymodoceaceae (160–241). Out of six SSR motifs, mononucleotide repeats were most abundant in all families (83–191), followed by dinucleotide repeats (13–44). TRs exhibited a broad range, from 26 (Elodea canadensis) to 288 (S. lichuanensis). Additionally, four direction types of repeats were searched, and Hydrocharitaceae showed the most variable repeats, ranging from 40 (Ottelia acuminata) to 961 (H. beccarii). Among these repeats, forward repeats always showed the highest number (13–479) compared to the other three types. Reverse and complement types were not detected in some species, such as E. canadensis (Table S2).

Distribution of repetitive elements and correlation analysis in Alismatidae. A SSR; B Dispersed repeats; C Tandem repeats; D correlation analysis among the repetitive elements and plastome/region size, and significant correlation (p < 0.05) was shown in the box without the Cross mark
The correlation analyses revealed a significant positive association between the size of IR and whole plastome (R = 0.77, p < 0.05; Fig. 3). In contrast, a negative correlation (R = -0.85, p < 0.05) was detected between the size of IR and SSC. The number of repeats and TRs showed a significant positive correlation with the length of IR (R = 0.56, p < 0.05; R = 0.44, p < 0.05) and the whole plastome (R = 0.55, p < 0.05; R = 0.67, p < 0.05).
Phylogenomic analysis
Phylogenomic analyses using two partitioning strategies and tree inference methods (ML and BI) resulted in identical and strongly supported topology ( Fig. 2; Table S3). The subclass Alismatidae was monophyletic with full support (BS = 100/100; PP = 1/1) and was further divided into two major clades. The first included Alismataceae, Hydrocharitaceae, and Butomaceae, while the rest of the families formed the second clade. In clade I, Hydrocharitaceae and Butomaceae were grouped together with full support (BS = 100/100; PP = 1/1) and were resolved as the sister of Alismataceae. In clade II, Aponogetonaceae was resolved as the sister to the remaining families. Marine species, which placed in Cymodoceaceae, Ruppiaceae, and Posidoniaceae, were resolved as a monophyletic sister of Potamogetonaceae and Zosteraceae with full support (BS = 100/100; PP = 1/1).
Discussion
Previous studies have revealed lineage-specific structural variations in plastomes of several flowering plant lineages and related them with the evolutionary history of the lineages under geological or climate change [24, 32]. For example, accD inversion was only found in the Asiatic species of Aponogeton (A. lakhonensis and A. undulatus) but not in the African species [33]. In Haloragaceae, Liao et al. [11] determined a 4-kb inversion in the lineage formed by Myriophyllum and its sister group and considered it might be associated with historical climate changes. In our study, ~ 8 kb inversion between rbcL and trnV-UAC (Type I) was found in addition to C. grandis in the monophyletic lineage of six families sister to the Juncaginaceae. Therefore, the inversion likely occurred after the divergence between Juncaginaceae and the other six families but before the initial diversification of these six families. According to the estimation of divergence times by Li et al. [34], this inversion took place between ca. 66 and 74 Ma, which coincides with the Cretaceous–Paleogene (K-Pg) extinction event [35]. Increasing genomic evidence has accumulated to support that angiosperms experienced a common paleopolyploidization during the K-Pg period in response to extreme climate change [36, 37]. It may be that chaotic genomic structure after polyploidization might simultaneously accelerate nucleoplasmic communication and cause non-random rearrangements in the plastome. However, until more data becomes available, this idea should be taken cautiously.
A novel finding here was that half of the inversion events (type I-III) in Alismatidae plastomes were located on or near the accD gene. The accD gene encodes the subunit of β-carboxyl transferase for Acetyl-CoA carboxylase, which plays an important role in regulating de novo fatty acid biosynthesis [38] and is essential for leaf growth and maintaining plastid compartment in tobacco [39]. Many studies have revealed a high variability of the accD gene across different plant lineages [40] and often anchored in rearrangement endpoints similar to what we observed here in Alismatidae (i.e., Cupressophytes [41]), which may be explained by continuous exchange with the nuclear genome.
The ndh genes mainly code the NADH complex, which is necessary for the electron transfer from NADH to plastoquinone in photosystem I under temperature stress [42]. We verified the previously detected three independent loss events in Hydrocharitaceae, Posidoniaceae, and Cymodoceaceae [24]. Among these, all 11 plastid ndh genes were completely lost in the genera of Najas and Halophila. Generally, the ndh gene losses have been explained by the transference of the functional ndh to the nuclear genome [16, 43]. Peredo et al. [18] speculated that the loss of ndh genes in aquatic plants might be associated with adaptation to submerged environments. However, none of the ndh genes were lost in other closely related submerged species, such as O. acuminata and Ruppia brevipedunculata. This indicates that the plastome evolution in aquatic angiosperm may not be as straightforward and that the gene losses are not directly related to the life forms. Thus, we prefer the hypothesis of plastid-nuclear exchange while maintaining the function of ndh. Interestingly, nearly half of the genes (ndhA, F, G, H, I) in the junction of SC/IR belong to the ndh complex. We observed the lowest relative size of SSC (2% in N. marina) and the highest relative size of IR (25% in T. hemprichii and H. beccarii) in Hydrocharitaceae, a family with complete losses of ndh genes (Fig. 1). Therefore, it seems reasonable to assume that the loss/pseudogenization of ndh genes is associated with the IR extraction/expansion, as has been suggested in Gentianaceae [22], Orobanchaceae [16], and Polygonaceae [19].
Repetitive elements have been suggested to play a key role in stabilizing the structure and size of plastomes in flowering plants [25, 26]. We found abundant repeats in plastomes throughout Alismatidae, with a significant positive correlation between the size of the plastome, IR and the number of dispersed and tandem repeats (Fig. 3). Although we did not find any distribution pattern of repeats associated with the plastome size, as in some other studies [44], it was evident that large repeats (i.e., IR) contributed to the variation of plastome size. The IR size negatively correlated with the size of SSC but positively with the size of the full plastome. Furthermore, several studies have postulated an increase in the content of repeat elements. If that is true, a similar repeat pattern might be expected in the specific lineage with the inversion of type I in contrast to outgroups (e.g., Juncaginaceae, Scheuchzeriaceae). However, we failed to find such a pattern. Thus, our results suggested that a possible explanation of illegitimate repeats might be that they are randomly generated and lost in different evolutionary lineages. Nevertheless, the effect of repeat elements on the variation of plastome structure and size is complex. Due to the limited species sampling in this study, our data is not an ideal example for understanding the dynamic change process for plastome repeats. In addition, as we have previously noted, the extremely abundant repetitive sequences in the family Alismataceae hindered the successful assembly of complete plastomes [5]. Future studies of plastome evolution in Alismatidae might benefit from long-read sequencing technologies (e.g., PacBio, Nanopore).
Conclusion
Our study is the first to investigate the structural variation of plastomes in Alismatidae at the family level. Comparative analyses revealed high variation in plastome size, repetitive elements among all twelve families and the specific-lineage inversion from rbcL and trnV-UAC (Type I). Three independent ndh loss events were identified across the Alismatidae more likely in association with the IR extraction/expansion rather than as adaptation to certain habitats. However, we should note that before we can fully answer these questions, we need to know if these lost/pseudogenized genes are translocated in the nuclear genome instead. In addition, we detected a positive correlation between the number of repeat elements and the size of plastomes and IR in Alismatidae. In summary, our findings allowed exploration of the evolutionary history of Alismatidae plastome and also can provide an opportunity to test if similar environmental adaptations resulted in convergent restructuring in plastomes.
Materials and methods
Plant materials, DNA extraction, sequencing, and plastome assembly
A total of 38 species representing all 12 accepted families in Alismatidae were incorporated into this study. These included 1) 13 species belonging to five families newly sequenced in this study; 2) 21 plastomes from eight families downloaded from GenBank, 3) the sequenced data of the remaining four species, Amphibolis antarctica (SRR19106495), Najas marina (ERR5529706), Posidonia australis (SRR19106496) and Zannichellia palustris (ERR5554861), retrieved from the SRA database (Table S1). Three additional plastomes from Araceae, Tofieldiaceae, and Acoraceae were included as outgroups. The field sampling followed the ethics and legality of the local government and was permitted by the government.
Total genomic DNA extraction, DNA fragmentation and preparation of sequencing libraries of all 13 newly sampled species followed the description by Li et al. [31]. Genome skimming per sample was conducted on the Illumina HiSeq 2000 platform in Novogene (Tianjin, China), with 150 bp paired-end reads. Newly sequenced reads of 13 species and four SRA sequenced reads were filtered with Fastp v.0.20.1 [45] using default settings. Complete plastomes were de novo assembled using the software GetOrganelle v.1.7.1 [46] with default parameters. Plastid Genome Annotator (PGA [47]) and the online tool Geseq [48] were utilized for the annotation of assembled plastomes. The annotations for protein-coding genes (PCGs) were checked and adjusted manually according to published plastomes of Alismatales. All newly generated plastomes were deposited in the China National GeneBank DataBase (CNGBdb, https://db.cngb.org/; Table S1).
Comparative plastome analysis
The plastome information, including the gene content, the proportion of coding and non-coding regions, and average gene density (genes/kb), were calculated using a custom python script. The IR extraction/expansion and structural variations (SVs) were detected using Geneious R v.11.0.5 (Biomatters, Auckland, New Zealand) with default settings.
Characterization of SSRs and repetitive sequences
The MISA script [49] was utilized to search the SSRs with the minimal parameters set to eight repeat motifs for mononucleotides, five for dinucleotides, four for trinucleotides and three for tetranucleotide, pentanucleotide and hexanucleotide repeats. Also, four types of dispersed repeats, including Forward (F), Reverse(R), Complement (C), and Palindromic (P), were identified using the online tool REPuter [50], with settings of a Hamming distance of three and minimal repeat size of 30 bp. Tandem repeats (TRs) within plastomes were further identified using the Tandem Repeats Finder program [51] with default parameters.
Phylogenetic reconstruction
A total of 41 plastomes, representing thirty-eight species from Alismatidae and three outgroup species (Tofieldia thibetica, Epipremnum amplissimum, and Acorus gramineus), was included in our phylogenetic analyses. All PCGs of each plastome were extracted and aligned using MAFFT v. 7.505 implemented in PhyloSuite v. 1.2.2 [52]. TrimAl v. 1.2 [53] was used to remove the ambiguous regions from the aligned sequence matrix with default settings before concatenating all clean alignments to a supermatrix. Phylogenetic analyses employing Maximum likelihood (ML) and Bayesian inference (BI) were conducted using unpartitioned data and data partitioned by genes. In the partitioned strategy, the best partitioning scheme was inferred using PartitionFinder v. 2.1.1 [54]. The program ModelFinder [55] was further used for searching the best substitution models. ML analysis was conducted in IQ-TREE v. 1.6.12 [56] with 5,000 rapid bootstrap replicates (BS). BI was carried on with MrBayes v. 3.2.7 [57] by running 2 × 107 generations and sampling every 1000 generations. The first 20% of trees were discarded as burn-in, and the remaining trees were summarized in a majority-rule consensus tree with the Bayesian posterior probabilities (PP).
Correlation analysis
Pearson’s correlation coefficients with significance level between the repeat (SSRs, TRs, repetitive sequences) and plastome sizes (whole genome, IR, and SSC) were conducted and visualized using the R package ‘ggcorrplot’ [58].
Availability of data and materials
All newly annotated plastomes in this study are available from the China.
National GeneBank DataBase with CNGB-Project ID: CNP0003742 and accession numbers: N_001484957, and N_001484959 to N_001484974 (See Additional file: Table S1).
Abbreviations
- BI:
-
Bayesian inference
- bp:
-
Base pair
- BS:
-
Bootstrap support
- IR:
-
Inverted repeat
- LSC:
-
Large single copy
- ML:
-
Maximum Likelihood
- PCGs:
-
Protein coding genes
- PP:
-
Posterior probability
- rRNA:
-
Ribosomal RNA
- SSC:
-
Small single copy
- SSR:
-
Simple sequence repeat
- SVs:
-
Structural variations
- tRNA:
-
Transfer RNA
- TRs:
-
Tandem repeats
References
Chen LY, Chen JM, Gituru RW, Wang QF. Eurasian origin of Alismatidae inferred from statistical dispersal–vicariance analysis. Mol Phylogenet Evol. 2013;67(1):38–42.
Les DH, Garvin DK, Wimpee CF. Phylogenetic studies in the monocot subclass Alismatidae: evidence for a reappraisal of the aquatic order Najadales. Mol Phylogenet Evol. 1993;2(4):304–14.
Cook CDK. Aquatic plant book. Hague: SPB Academic Publishing; 1990.
Chen LY, Lu B, Morales-Briones DF, Moody ML, Liu F, et al. Phylogenomic analyses of Alismatales shed light into adaptations to aquatic environments. Mol Biol Evol. 2022;39(5):msac079.
Li ZZ, Lehtonen S, Martins K, Wang QF, Chen JM. Complete genus-level plastid phylogenomics of Alismataceae with revisited historical biogeography. Mol Phylogenet Evol. 2022;166:107334.
Jansen RK, Cai Z, Raubeson LA, Daniell H, Depamphilis CW, et al. Analysis of 81 genes from 64 plastid genomes resolves relationships in angiosperms and identifies genome-scale evolutionary patterns. Proc Natl Acad Sci USA. 2007;104(49):19369–74.
Wicke S, Schneeweiss GM, Depamphilis CW, Müller KF, Quandt D. The evolution of the plastid chromosome in land plants: gene content, gene order, gene function. Plant Mol Biol. 2011;76(3):273–97.
Jin DM, Jin JJ, Yi TS. Plastome structural conservation and evolution in the clusioid clade of Malpighiales. Sci Rep. 2020;10(1):1–6.
Lee C, Choi IS, Cardoso D, de Lima HC, de Queiroz LP, et al. The chicken or the egg? Plastome evolution and an independent loss of the inverted repeat in papilionoid legumes. Plant J. 2021;107(3):861–75.
Henriquez CL, Ahmed I, Carlsen MM, Zuluaga A, Croat TB, et al. Evolutionary dynamics of chloroplast genomes in subfamily Aroideae (Araceae). Genomics. 2020;112(3):2349–60.
Liao YY, Liu Y, Liu X, Lü TF, Mbichi RW, et al. The complete chloroplast genome of Myriophyllum spicatum reveals a 4-kb inversion and new insights regarding plastome evolution in Haloragaceae. Ecol Evol. 2020;10(6):3090–102.
Li ZZ, Gichira AW, Muchuku JK, Li W, Wang GX, et al. Plastid phylogenomics and biogeography of the genus Monochoria (Pontederiaceae). J Syst Evol. 2021;59(5):1027–39.
Bedoya AM, Ruhfel BR, Philbrick CT, Madriñán S, Bove CP, et al. Plastid genomes of five species of riverweeds (Podostemaceae): structural organization and comparative analysis in Malpighiales. Front Plant Sci. 2019;10:1035.
Choi IS, Jansen R, Ruhlman T. Lost and found: return of the inverted repeat in the legume clade defined by its absence. Genome Biol Evol. 2019;11(4):1321–33.
Du XY, Kuo LY, Zuo ZY, Li DZ, Lu JM. Structural variation of plastomes provides key insight into the deep phylogeny of ferns. Front Plant Sci. 2022;13:862772.
Li X, Yang JB, Wang H, Song Y, Corlett RT, et al. Plastid NDH pseudogenization and gene loss in a recently derived lineage from the largest hemiparasitic plant genus Pedicularis (Orobanchaceae). Plant Cell Physiol. 2021;62(6):971–84.
Wen Y, Qin Y, Shao B, Li J, Ma C, et al. The extremely reduced, diverged and reconfigured plastomes of the largest mycoheterotrophic orchid lineage. BMC Plant Biol. 2022;22(1):1–11.
Peredo EL, King UM, Les DH. The plastid genome of Najas flexilis: adaptation to submersed environments is accompanied by the complete loss of the NDH complex in an aquatic angiosperm. PLoS ONE. 2013;8(7):e68591.
Qu XJ, Zhang XJ, Cao DL, Guo XX, Mower JP, et al. Plastid and mitochondrial phylogenomics reveal correlated substitution rate variation in Koenigia (Polygonoideae, Polygonaceae) and a reduced plastome for Koenigia delicatula including loss of all ndh genes. Mol Phylogenet Evol. 2022;174:107544.
Lam VKY, Soto Gomez M, Graham SW. The highly reduced plastome of mycoheterotrophic Sciaphila (Triuridaceae) is colinear with its green relatives and is under strong purifying selection. Genome Biol Evol. 2015;7(8):2220–36.
Iles WJD, Smith SY, Graham SW. A well-supported phylogenetic framework for the monocot order Alismatales reveals multiple losses of the plastid NADH dehydrogenase complex and a strong long-branch effect. In: Wilkin P, Mayo SJ, editors. Early Events in Monocot Evolution. Cambridge: Cambridge University Press Systematics Association Special Series; 2013. p. 1–28.
Fu PC, Chen SL, Sun SS, Favre A. Strong plastid degradation is consistent within section Chondrophyllae, the most speciose lineage of Gentiana. Ecol Evol. 2022;12(8):e9205.
Fu PC, Sun SS, Twyford AD, Li BB. Lineage-specific plastid degradation in subtribe Gentianinae (Gentianaceae). Ecol Evol. 2021;11(7):3286–99.
Ross TG, Barrett CF, Soto Gomez M, Lam VK. Plastid phylogenomics and molecular evolution of Alismatales. Cladistics. 2016;32(2):160–78.
Wu S, Chen J, Li Y, Liu A. Extensive genomic rearrangements mediated by repetitive sequences in plastomes of Medicago and its relatives. BMC Plant Biol. 2021;21(1):1–16.
Guisinger MM, Kuehl JV, Boore JL, Jansen RK. Extreme reconfiguration of plastid genomes in the angiosperm family Geraniaceae: rearrangements, repeats, and codon usage. Mol Biol Evol. 2011;28(1):583–600.
Cai Z, Guisinger M, Kim HG, Ruck E, Blazier JC, et al. Extensive reorganization of the plastid genome of Trifolium subterraneum (Fabaceae) is associated with numerous repeated sequences and novel DNA insertions. J Mol Evol. 2008;67(6):696–704.
Asaf S, Khan AL, Jan R, Khan A, Khan A, et al. The dynamic history of gymnosperm plastomes: Insights from structural characterization, comparative analysis, phylogenomics, and time divergence. Plant Genome. 2021;14(3):e20130.
Maréchal A, Parent JS, Véronneau-Lafortune F, Joyeux A, Lang BF, et al. Whirly proteins maintain plastid genome stability in Arabidopsis. Proc Natl Acad Sci USA. 2009;106(34):14693–8.
Li ZZ, Lehtonen S, Martins K, Gichira AW. Phylogenomics of the aquatic plant genus Ottelia (Hydrocharitaceae): implications for historical biogeography. Mol Phylogenet Evol. 2020;152:106939.
Li ZZ, Lehtonen S, Gichira AW, Martins K, Efremov A, et al. Plastome phylogenomics and historical biogeography of aquatic plant genus Hydrocharis (Hydrocharitaceae). BMC Plant Biol. 2022;22(1):1–11.
Kim KJ, Choi KS, Jansen RK. Two chloroplast DNA inversions originated simultaneously during the early evolution of the sunflower family (Asteraceae). Mol Biol Evol. 2005;22(9):1783–92.
Mwanzia VM, He DX, Gichira AW, Li Y, Ngarega BK, et al. The complete plastome sequences of five Aponogeton species (Aponogetonaceae): insights into the structural organization and mutational hotspots. Plant Divers. 2020;42(5):334–42.
Li HT, Yi TS, Gao LM, Ma PF, Zhang T, et al. Origin of angiosperms and the puzzle of the Jurassic gap. Nat Plants. 2019;5(5):461–70.
Vanneste K, Baele G, Maere S, Van de Peer Y. Analysis of 41 plant genomes supports a wave of successful genome duplications in association with the Cretaceous-Paleogene boundary. Genome Res. 2014;24(8):1334–47.
Vellekoop J, Smit J, van de Schootbrugge B, Weijers JW, Galeotti S, et al. Palynological evidence for prolonged cooling along the Tunisian continental shelf following the K-Pg boundary impact. Palaeogeogr Palaeoclimatol Palaeoecol. 2015;426:216–28.
Wu S, Han B, Jiao Y. Genetic contribution of paleopolyploidy to adaptive evolution in angiosperms. Mol Plant. 2020;13(1):59–71.
Bock R. Structure, function, and inheritance of plastid genomes. In: Bock R, editor. Cell and molecular biology of plastids. Berlin: Springer; 2007. p. 29–63.
Kode V, Mudd EA, Iamtham S, Day A. The tobacco plastid accD gene is essential and is required for leaf development. Plant J. 2005;44(2):237–44.
Sudianto E, Chaw SM. Two independent plastid accD transfers to the nuclear genome of Gnetum and other insights on Acetyl-CoA Carboxylase evolution in gymnosperms. Genome Biol Evol. 2019;11(6):1691–705.
Li J, Su Y, Wang T. The repeat sequences and elevated substitution rates of the chloroplast accD gene in cupressophytes. Front Plant Sci. 2018;9:533.
Wang P, Duan W, Takabayashi A, Endo T, Shikanai T, et al. Chloroplastic NAD (P) H dehydrogenase in tobacco leaves functions in alleviation of oxidative damage caused by temperature stress. Plant Physiol. 2006;141(2):465–74.
Kim YK, Jo S, Cheon SH, Joo MJ, Hong JR, et al. Extensive losses of photosynthesis genes in the plastome of a mycoheterotrophic orchid, Cyrtosia septentrionalis (Vanilloideae: Orchidaceae). Genome Biol Evol. 2019;11(2):565–71.
Weng ML, Blazier JC, Govindu M, Jansen RK. Reconstruction of the ancestral plastid genome in Geraniaceae reveals a correlation between genome rearrangements, repeats, and nucleotide substitution rates. Mol Biol Evol. 2014;31(3):645–59.
Chen S, Zhou Y, Chen Y, Gu J. fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics. 2018;34:i884–90.
Jin JJ, Yu WB, Yang JB, Song Y. GetOrganelle: a fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020;21(1):1–31.
Qu XJ, Moore MJ, Li DZ, Yi TS. PGA: a software package for rapid, accurate, and flexible batch annotation of plastomes. Plant Methods. 2019;15(1):1–12.
Tillich M, Lehwark P, Pellizzer T, Ulbricht-Jones ES, Fischer A, et al. GeSeq–versatile and accurate annotation of organelle genomes. Nucleic Acids Res. 2017;45(W1):W6–11.
Beier S, Thiel T, Münch T, Scholz U, Mascher M. MISA-web: a web server for microsatellite prediction. Bioinformatics. 2017;33(16):2583–5.
Kurtz S, Choudhuri JV, Ohlebusch E, Schleiermacher C, Stoye J, et al. REPuter: the manifold applications of repeat analysis on a genomic scale. Nucleic Acids Res. 2001;29(22):4633–42.
Benson G. Tandem repeats finder: a program to analyze DNA sequences. Nucleic Acids Res. 1999;27(2):573–80.
Zhang D, Gao F, Jakovlić I, Zou H, Zhang J, et al. PhyloSuite: an integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol Ecol Resour. 2020;20(1):348–55.
Capella-Gutiérrez S, Silla-Martínez JM, Gabaldón T. trimAl: a tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics. 2009;25(15):1972–3.
Lanfear R, Frandsen PB, Wright AM, Senfeld T, Calcott B. PartitionFinder 2: new methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol Biol Evol. 2017;34(3):772–3.
Kalyaanamoorthy S, Minh BQ, Wong TKF, Von Haeseler A, Jermiin LS. ModelFinder: fast model selection for accurate phylogenetic estimates. Nat Methods. 2017;14(6):587–9.
Nguyen LT, Schmidt HA, Von Haeseler A, Minh BQ. IQ-TREE: a fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol Biol Evol. 2015;32(1):268–74.
Ronquist F, Teslenko M, Van Der Mark P, Ayres DL, Darling A, et al. MrBayes 3,2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst Biol. 2012;61(3):539–42.
Kassambara A. ggcorrplot: Visualization of a Correlation Matrix using 'ggplot2'. R package version 0.1.3. https://CRAN.R-project.org/package=ggcorrplot. Accessed 1 Oct 2022.
Acknowledgements
We are grateful to Rosie Woods for help in the DNA collection from Kew DNA bank. This Research was performed under Permit No. DNPW/8/27/1 from the department of National Parks and Wildlife, Zambia.
Funding
This study was supported by grants from the Strategic Priority Research Program of the Chinese Academy of Sciences (XDB31000000) and the National Natural Science Foundation China (32100186).
Author information
Authors and Affiliations
Contributions
SL, and J‑M C conceived and designed the study. Z‑Z L performed all analyses. Z‑Z L drafted the manuscript. Z‑Z L, SL, J‑MC collected the leaf materials. Z‑Z L, SL and J-M C revised the manuscript. All authors approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1:
Table S1. The genome content of all plastome sequences used in this study.
Additional file 2: Table S2.
Detailed information on repetitive elements across Alismatidae plastomes in this study.
Additional file 3: Table S3.
The best partition scheme and models were estimated based on PartitionFinder analysis.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Li, ZZ., Lehtonen, S. & Chen, JM. The dynamic history of plastome structure across aquatic subclass Alismatidae. BMC Plant Biol 23, 125 (2023). https://doi.org/10.1186/s12870-023-04125-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-023-04125-x