
Abstract
Background
The exchangeable aluminum (Al), released from the acid soils, is another addition to the environmental stress factors in the form of Al toxicity stress. Al stress affects the normal crop development and reduces the overall yield of rapeseed (Brassica napus L.). The response mechanism of plants to Al toxicity is complicated and difficult to understand with few QTL related studies in rapeseed under Al toxicity stress.
Result
Using 200,510 SNPs developed by SLAF-seq (specific-locus amplified fragment sequencing) technology, we carried out the genome-wide association analysis (GWAS) in a population of 254 inbred lines of B. napus with large genetic variation and Al-tolerance differences. There were 43 SNPs significantly associated with eight Al-tolerance traits in the seedling stage were detected on 14 chromosomes, and 777 candidate genes were screened at the flanking 100 kb region of these SNPs. Moreover, RNA-seq detected 8291 and 5341 DEGs (the differentially expressed gene) in the Al -tolerant line (ATL) and -sensitive line (ASL), respectively. Based on integration of GWAS and RNA-seq analysis, 64 candidate genes from GWAS analysis differentially expressed at least once in 6 h vs 0 h or 24 h vs 0 h conditions in ATL or ASL. Moreover, four out of sixty-four candidate genes (BnaA03g30320D, BnaA10g11500D, BnaC03g38360D and BnaC06g30030D) were differentially expressed in both 6 h and 24 h compared to 0 h (control) conditions in both lines. The proposed model based on the candidate genes excavated in this study highlighted that Al stress disturb the oxidation-redox balance, causing abnormal synthesis and repair of cell wall and ABA signal transduction, ultimately resulting in inhibition of root elongation.
Conclusions
The integration of GWAS and transcriptome analysis provide an effective strategy to explore the SNPs and candidate genes, which has a potential to develop molecular markers for breeding Al tolerant rapeseed varieties along with theoretical basis of molecular mechanisms for Al toxicity response of Brassica napus plants.
Similar content being viewed by others


Introduction
Aluminum (Al), after oxygen and silicon, is the most abundant metal element in the earth’s crust. Al exists in the form of insoluble silicates or oxides which are less harmful to the growth and development of crops [1, 2]. However, soil pH value below 5.5 promotes exponential release of the exchangeable Al (mainly A13+, Al(OH)2+ and Al(OH)2+) from silicates or oxides, which has a strong toxic effect on crop roots growth [3,4,5]. The free Al3+ ions can bind to the plasma membrane and nucleus, inhibiting the elongation and division of tip cells of crop roots, which affects the uptake of water and nutrients [6,7,8]. At present, about 40% of the world’s potentially arable lands are acidic (pH < 5.5) [9, 10]. Al toxicity has become a prominent factor affecting the crop growth on these acidic soils.
Brassica napus is the second largest oilseed crop in the world, providing edible oil for humans along with other multiple usages in the form of vegetable, forage, ornamental flower, honey and fertilizer [11, 12]. In China, the Yangtze River region where was the high rapeseed producing area has major issues with acidic soils specifically Al toxicity stress limiting the growth and seed yield of B. napus [4, 13]. At present, Al tolerance related research mainly focus on model plants such as Arabidopsis, rice, wheat and barley [14,15,16]. Previous research has demonstrated Al tolerance in plants as a complex trait controlled by multiple genes and pathways [17, 18]. Various genetic loci and genes involved in Al exclusion and tolerance mechanisms have been identified in model plants [18]. Success linked to a few genes (BnALMT1, BnALMT2, CS and WMnSOD1) in improving Al tolerance in transgenic plants has been studied previously [19,20,21]. However, many crucial genes linked to Al resistance in other crops, such as RAL1 [22], OsFRDL2 [23], FeSTAR2 [24] and VuSTOP1 [25], have not been reported in B. napus. It limits our understanding about the genetic variation and molecular mechanism of Al tolerance in B. napus.
Genome-wide association study (GWAS) have proved as a powerful tool in identification of desired trait linked genes in plants [26,27,28], and been applied in mining gene loci and candidate genes related to Al tolerance in various crops such as rice, wheat, and barley [14,15,16]. Recently, for B. napus, Gao et al. [13] detected 13 SNPs associated with two traits of relative root length and relative dry weight during germination using GWAS analysis. In addition, some studies have reported more accuracy and efficacy in screening the candidate genes for agronomic and stress-related traits by integrating GWAS and RNA-seq [29,30,31,32]. Zhang et al. [31] identified 16 loci significantly associated with water stress response in Canola using GWAS, and then 79 candidate genes were identified by combining differentially expressed genes (DEGs) detected by RNA-seq with loci from GWAS. Later on same approach identified 24 stalk rot resistance-related candidate genes in 17 sites, and 33 functional candidate genes related to rapeseed harvest index [30, 32]. Therefore, a combined strategy of GWAS and RNA-seq analysis showed more reliable potential to identify the candidate genes related to complex traits of rapeseed.
In this study, a set of 254 inbred lines of B. napus with large genetic variation and Al-tolerance differences were selected. This study also had an advantage of the 200,510 high-quality SNPs developed by SLAF-seq (specific-locus amplified fragment sequencing) technology [33]. GWAS was carried out to detect SNPs linked to the Al-tolerance and loci of elite allelic variation. In addition, the roots of two highly tolerant and susceptible rapeseed lines treated with Al ion stress were used to identify DEGs related to Al tolerance using RNA sequencing. DEGs for Al tolerance within the LD intervals containing significant SNP markers were selected as Al tolerance candidate genes by combining the analysis of GWAS and RNA-seq. The objective of this study was to identify SNP markers and candidate genes linked with Al tolerance in rapeseed.
Results
Phenotypic data
Eight traits for each of 254 rapeseed inbred lines were investigated under the stress of Al toxicity, and descriptive statistical analysis was summarized in Table S1 for fresh weight above ground, root average diameter, root fresh weight, root elongation, total root surface area, total root tip, total root volume, total root length of 254 rapeseed under CK and Al stress (Treatment). The coefficients of variation (CV, %) in CK and Treatment were ranged from 17.3 to 105.3 and 17.2 to 117.8, respectively.
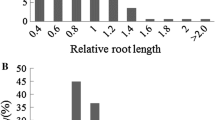
The CV of eight traits between CK and Treatment ranged from 11.3 for relative root elongation (RRE) to 30.8 for relative root fresh weight (RRFW) (Table 1). RRE varied from 0.537 to 0.947 with an average of 0.750, and RRFW varied from 0.490 to 2.143 with an average of 1.071. The genotypes showed extremely significant differences with normal distribution for all the phenotypic traits among 254 inbred lines (Table 1; Fig. 1). This pattern showed presence of a broad phenotypic mutation under Al stress, which could effectively be used to locate Al tolerance linked candidate genes by GWAS.

Frequency distribution of eight phenotypic traits related to Al tolerance in Brassica napus
Furthermore, Strong positive correlations (R = 0.5–0.9) were observed among most of the traits, such as relative fresh weight above ground (RFWAG) with RRFW and relative total root tip (RTRT), relative total root surface area (RTRSA) with relative total root volume (RTRV), relative total root length (RTRL) and relative root average diameter (RRD) (Table 2).
Genome-wide association study
A total of 200,510 SNPs were used in the GWAS analysis for Al tolerance using GLM and MLM. The distribution agrees of p-values have a high consistency with observations by the Quantile-quantile plots (Q-Q plots) analysis (Fig. 2). The GLM analysis detected a total of 43 SNPs significantly associated with eight Al tolerance traits. These SNPs explained the phenotypic variation from 8.22 to 14.36% and their distribution was detected on 14 of the 19 B. napus chromosomes (excluding A05, A07, C01, C07 and C08). Besides, the largest number of significant SNPs was on chromosome C02 (six SNPs) and RRFW trait had the most associated SNPs (eight SNPs) (Fig. 3; Table 3). MLM analysis detected a total of 12 significantly associated SNPs with RRD (1), RRFW (3), RRE (2), RTRSA (1), RTRT (1), RTRV (2), and RTRL (1) on six chromosomes (A01, A03, A04, C02, C03 and C05), respectively, explaining phenotypic variation of 9.49 to 14.52% (Fig. 4; Table 3). Totally, 43 significant SNPs associated with eight Al tolerance traits were identified by GLM and MLM analyses.

Quantile-quantile plots of estimated-lg (P) from association analysis of eight traits using GLM and MLM model

Manhattan plots for eight phenotypic traits related to Al tolerance in Brassica napus by GLM model. Note: The blue horizontal line represents that the extreme significance threshold -log10(P) value is approximately equal to 5.3

Manhattan plots for eight phenotypic traits related to Al tolerance in Brassica napus by MLM model. Note: The blue horizontal line represents that the extreme significance threshold -log10(P) value is approximately equal to 5.3
Among these significant SNPs (Table 3), two SNPs for RFWAG, detected by GLM analyses, were located on chromosomes A04. For RRD, three SNPs were identified on two chromosomes (A01 and C03), one SNP detected by GLM and two SNPs by both GLM and MLM models. Similarly, eight SNPs for RRFW were detected on chromosome A03, A06, A08, A10, C02 and C03, five of these were detected by GLM and three by both GLM and MLM analyses. GLM analysis was able to identify seven SNPs associated with RRE on six chromosomes (A03, A10, C02, C04, C06 and C09). GLM predictions were also confirmed by MLC identification of two SNPs on chromosome A03 associated with RRE. Six SNPs for RTRSA were detected on four chromosomes (A01, A02, A09 and C02) by only GLM analysis, however, one SNP was a shared result of both GLM and MLM analyses. MLM analysis detected six SNPs for RTRT, one SNP of which was consistent with GLM analysis. Five SNPs associated with RTRV were identified on chromosome A06, C04 and C05 by GLM analysis, two of which were consistent with MLM analysis. Five SNPs on four chromosomes (A01, A06, A09 and C02) linked to RTRL were detected by GLM, and two SNPs detected by both GLM and MLM models.
Furthermore, four SNP loci were linked to multiple traits such as locus Bn-A01-p8185115 (chromosome A01) was associated with three traits including RRD, RTRSA and RTRL. Similarly, Bn-A01-p11875598 was associated with RRD and RTRL, Bn-A04-p9422509 associated with RFWAG and RTRT, Bn-C02-p26059415 associated with RTRSA and RTRL (Table 3).
Identification of candidate genes
The candidate genes nearby the genome-wide significant SNPs were identified by using 100 kb flanking sequences of 43 SNPs (significantly associated with Al-tolerance) and ‘Darmor v4.1’ as the reference genome. This study resulted in the identification of 777 candidate genes, most of which genes were involved in various functions such as amino acid transport and metabolism, defense mechanisms, inorganic ion transport and metabolism by COG annotation (Fig. S1). Based on the functional annotations, some genes were known to be related with Al tolerance, such as MATE family proteins, ABC transporter family protein, aquaporin, sulfate transporter family protein, metal tolerance protein, glutathione S-transferase, xyloglucan endotransglucosylase/hydrolase protein and antioxidant proteins. Some candidate genes, such as BnaC04g06050D, BnaC04g06060D, BnaA03g43560D, BnaA03g30320D and BnaA07g29670D, mainly participated in the pathway related to transport and metabolism of inorganic ions, the transport and discharge of organic acids (citric acid), and oxidative stress response (Table S2).
Transcriptome sequencing analysis
The transcriptomes of two breeding lines under Al stress were analyzed. More than 10.15 billion clean reads from 18 libraries of two genotypes were generated and mapped to the reference genome. The alignment results showed that 669 million reads of the clean reads were successfully mapped to the reference genome (Table S3).
To determine genes correlated with Al stress response, DEGs for 0 h, 6 h and 24 h Al stress treatments for both lines were identified. In the Al-tolerant line, a total of 3053 genes showed up-regulation and 3644 genes showed down-regulation under 6 h treatment, whereas 655 genes showed up-regulation and 939 genes showed down-regulation under 24 h treatment compared to 0 h treatment (FDR ≤ 0.05 and Log2(FC) ≥ 1.0 or ≤ − 1.0). Among these DEGs, 270 genes were up-regulated and 508 genes down-regulated under both of the 6 h and 24 h Al stress durations. An up-regulation was observed for 33 genes under 6 h duration, but 24 h duration down-regulated the same genes compared to control. However, 10 genes were down-regulated under 6 h but up-regulated under 24 h compared to control (Fig. S2A).
In the Al-sensitive line, a total of 2248 and 2058 genes showed up-regulation and down-regulation, respectively, under 6 h compared to control. 220 genes showed up-regulation and 815 genes showed down-regulation under 24 h treatment duration compared to control. Among these DEGs, 99 genes were up-regulated and 490 genes down-regulated, both under 6 h and 24 h Al stress. Compared to control treatment, five genes were up-regulated under 6 h, but down-regulated under 24 h (Fig. S2B). We randomly selected six genes involved in the Al stress for expression validation by qRT-PCR, and the expression trends were similar with the RNA-seq data (Fig. S3).
DEGs of Al-tolerant and Al-sensitive lines
A total of 2569 DEGs under 6 h and 265 DEGs under 24 h treatment durations were compared to control for Al-tolerant and Al-sensitive lines, respectively. In 6 h treatment, 1255 genes were up-regulated and 1306 genes down-regulated both in ATL and ASL. Moreover, four genes were up-regulated in ATL and down-regulated in ASL; another group of four genes down-regulated in ATL but up-regulated in ASL in 6 h treatment (Fig. 5). The 24 h treatment duration compared to 0 h up-regulated 46 genes and down-regulated 217 genes in the ATL and ASL, two genes were up-regulated in ATL and down-regulated in ASL (Fig. 5). Interestingly, 151 common DEGs were identified in ATL and ASL under both treatment durations (Table S4). Among these DEGs, 25 DEGs and 124 DEGs were respectively up-regulated and down-regulated both in 6 h and 24 h of ATL and ASL, two DEGs were up-regulated in 6 h of ASL while down-regulated both in 6 h and 24 h of ATL and in 24 h of ASL.

The number of up and down –regulation genes between ATL and ASL under Al toxicity
Based on COG annotations, some genes of 151 DEGs were involved in carbohydrate transport and metabolism (20 DEGs), posttranslational modification (14 DEGs), Cell wall/membrane/envelope biogenesis (7 DEGs), signal transduction mechanisms (14 DEGs), defense mechanisms (13 DEGs), inorganic ion transport and metabolism (7 DEGs) (Fig. S4). Some of DEGs such as MATE family, ABC transporter family, zinc finger, glutathione S-transferase, xyloglucan endotransglucosylase/hydrolase protein and heavy-metal-associated domain were responsive to Al stress (Table S5).
Identification of candidate genes by integrating GWAS and RNA-seq analysis
The potential candidate genes were prioritized by integrating DEGs obtained by GWAS and RNA-seq analysis. Out of 777 candidate genes identified by GWAS, 64 (8.24%) genes distributed on 13 of the 19 B. napus chromosomes (excluding A05, A07, C01, C07 C08 and C09). These candidate genes were differentially expressed in at least one genotype under 6 h or 24 h compared with 0 h conditions (Fig. 6; Table S6). The largest number of candidate genes were on chromosome A03 (18 genes) in the flanking 100 kb region of four SNPs. Ten candidate genes were screened from three SNPs and seven genes from two SNPs on chromosome A06 and A10, respectively. Similarly, six candidate genes were screened from the flanking region of one SNP (Bn-A01-p8185115) on chromosome A01.

Distribution of candidate genes and their corresponding SNP loci associated with Al tolerance. Note: The blue as SNP loci, red as the gene differentially expressed both in 6 h vs 0 h and 24 h vs 0 h of ATL and ASL. The numeric values represent the relative distances in the genome, 1 = 1 kb
The candidate genes enabled us to identify several Al tolerance related genes in breeding lines (Fig. 6; Fig. 7). For example, two genes (BnaA03g30330D and BnaA03g30320D) were detected adjacent to SNP Bn-A03-p14798182 and another gene (BnaA09g14730D) adjacent to SNP Bn-A09-p8460525 belong to the MATE gene family. A gene BnaC04g06210D on chromosome C04 belonged to ABC transporter family protein in the vicinity of SNP Bn-C04-p4409586. One gene of BnaA03g12450D involved in abscisic acid (ABA) signal regulation was found in the vicinity of SNP Bn-A03-p5766579. These genes BnaA01g15810D, BnaA01g15880D, BnaA03g22360D, BnaA06g18630D, BnaA10g06710D, BnaA10g11500D, BnaC04g05980D were located adjacent to SNP Bn-A01-p8185115, Bn-A03-p10703126, Bn-A06-p10959923, Bn-A10-p5162750, Bn-A10-p9658437 and Bn-C04-p4409586 on their respective chromosomes and participated in cell wall development. Besides, two genes (BnaA03g22680D and BnaA03g33540D) on chromosome A03 involved in ion transport process.

The possible metabolic pathway for candidate gens under Al stress (A) and the possible model for rapeseed in response to Al stress (B). Note: 1-P-G, glucose 1-phosphate; 6-P-Fru, fructose 6-phosphate; GDP-G, GDP-glucose; UDP-G, UDP-glucose; GDP-L Ga, galactose GDP-L; L-1-P Ga, galactose L-1-phosphate; 3-PG, 3-phosphoglycerate; PEP, phosphoenolpyruvate; ICP, ion channel protein; STOP1, sensitive to proton rhizotoxicity 1; MATE, MATE efflux family protein; ABA, abscisic acid; TCA, tricarboxylic acid cycle; EMP pathway, glycolysis pathway
Among these 64 candidate genes, four were simultaneously detected by GWAS and the common DEGs between 6 h vs 0 h and 24 h vs 0 h in ATL and ASL. These four genes include those encoding a MATE family protein (BnaA03g30320D), xyloglucan endotransglucosylase/hydrolase (BnaA10g11500D), a DnaJ domain protein (BnaC03g38360D), and an unknown function protein (BnaC06g30030D) (Table 4).
Discussion
Detection of novel SNP loci significantly associated with Al tolerance in B. napus
Al toxicity stress limits the crop growth and yield by affecting root meristem, cell mitosis and root growth in crops [8, 34]. Al toxicity has been reported in various crops targeting root related traits such as longest and primary root growth, total root growth and relative root elongation for discovering genes involved in Al tolerance by GWAS approach [13, 35, 36]. For example, Famoso et al. [16] identified 48 regions associated with three root growth parameters in rice seedlings; later on this finding helped in identification of 23 and 43 significant loci associated with relative root elongation in rice [35, 36]. Previously, 13 SNPs significantly associated with relative root length and relative dry weight during germination period detected in 169 rapeseed cultivars (lines) using 60 K Brassica Illumina Infinium SNP array [13]. In this research, we investigated eight traits related to Al tolerance (RFWAG, RRD, RRFW, RRE, RTRSA, RTRT, RTRV and RTRL) for 254 rapeseed accessions and detected 43 associated SNP loci on 14 chromosomes by GWAS using SLAF-Seq as detailed in our previous studies [33, 37], which explained the phenotypic variation from 8.22 to 14.36% (Fig. 2; Table 3). Among these SNP loci, Bn-A04-p9422509 and Bn-A09-p8460525 were respectively in the range 1 Mb of Bn-A04-p7776319 and Bn-A09-p9030563 which were significantly associated with Al tolerance at germination stage of B. napus in previous study [13]. In addition, there were 41 novel SNP loci discovered on 12 chromosomes (Table 3), and four SNPs of which (Bn-A01-p8185115, Bn-A01-p11875598, Bn-A04-p9422509 and Bn-C02-p26059415) were significantly associated with more than one trait, which might be caused by linkage or pleiotropy [38]. In previous studies, the RRE were used to evaluate the Al resistance [35, 39, 40], in this study, the RTRL were positive correlations with RRE, and five SNP loci were significantly associated with RTRL, three of five SNP loci were significantly associated with multiple traits. Therefore, RRE and RTRL can be used to evaluate the Al resistance of B. napus. Our results provide insights into the significantly association of SNPs with Al tolerance traits, which could be a potential marker for improving the Al tolerance breeding in B. napus.
Mining of candidate genes to uncover the Al tolerance gene network by integrating GWAS and transcriptome
Two main strategies have been found to deal with Al toxicity in acidic soils, including external exclusion and internal tolerance [17, 18, 41]. The external exclusion prevents plant roots absorbing a large amount of Al (Al3+) to reduce toxicity [17, 42]. The internal tolerance mechanism detoxify internal Al in plant cells by chelating with organic acids and converting the absorbed ionic Al into combined Al [17, 43]. Among these strategies, secretion of Al-induced root organic acid to chelate Al for protecting cell wall from Al binding is the well-documented mechanism [17, 44, 45]. Previously, map-based cloning found MATE gene family to resist Al toxicity stress in barley (HvAACT) and sorghum (SbMATE) [46, 47]. It was followed by MATE homologs to promote citrate excretion into the rhizosphere to protect roots from Al toxicity in maize (ZmMATE1) [48], rice (OsFRDL4 and OsFRDL2) [23, 49] and soybean (GmMATE75, GmMATE79 and GmMATE87) [50]. In current study, five genes belonging to MATE gene family were detected by GWAS, three candidate genes (BnaA03g30320D and BnaA03g30330D adjacent to SNP Bn-A03-p14798182, BnaA09g14730D adjacent to SNP Bn-A09-p8460525) were differentially expressed by RNA-seq (Fig. 7A). The expression of the gene BnaA03g30320D was more down-regulated in ASL than ATL under both 6 h vs 0 h and 24 h vs 0 h conditions. Our results are consistent with a previous detection of MATE family genes at germination stage under Al toxicity stress [13], which contribute in Al tolerance in B. napus.
The root cell wall becomes the next site of Al interaction after traversing the organic acid barrier in the root rhizosphere [18]. Even with a complex structure of cell wall, the negatively charged carboxyl groups in pectin and uncharged hemicellulose binding, resulted in the distortion of cell wall extension under Al stress [51, 52]. In the present study, one ABC transporter gene on chromosome C04 (BnaC04g06210D) was identified, which is involved in abscisic acid (ABA) transport and responses [53]. ABC transporter could regulate the plant Al tolerance by transporting UDP-glucose, which affects hemicellulose metabolism by regulating xyloglucan endotransglucosylase/hydrolases activity [24, 54]. ABC transporters also play an important role in Al resistance mechanism [53, 55, 56], for instance, OsALS1, FeALS1.1 and FeALS1.2, all of the homologous gene AtALS1 encoding a half-size ABC transporter, were involved in the internal detoxification of Al in rice and buckwheat [57, 58]. Furthermore, seven candidate genes (BnaA01g15810D and BnaA01g15880D adjacent to Bn-A01-p8185115, BnaA03g22360D adjacent to Bn-A03-p10703126, BnaA06g18630D adjacent to Bn-A06-p10955523, BnaA10g06710D adjacent to Bn-A10-p5162750, BnaA10g11500D adjacent to Bn-A10-p9658437, and BnaC04g05980D adjacent to Bn-C04-p4409586) involved in cell wall components were also detected in this study (Fig. 7A). Two of seven genes encoding xyloglucan endotransglucosylase/hydrolase (BnaA06g18630D and BnaA10g11500D) were detected on chromosome A06 and A10, respectively. Previously, Zhu et al. [51] reported that XTH31 encoding a xyloglucan endotransglucosylase/hydrolases regulates Al sensitivity by modulating cell wall xyloglucan content and Al binding capacity. Both ASL and ATL showed down-regulation of BnaA10g11500D in 6 h vs 0 h and 24 h vs 0 h conditions. ATL also showed a down-regulation of BnaA10g06710D (probable pectinesterase/pectinesterase inhibitor) in 6 h vs 0 h and 24 h vs 0 h conditions, indicating the presence of gene specific expression pattern.
Plant hormones such as ABA, JA and SA play an important role in the stress related defense system [59]. ABA signal transduction pathways provide an additional layer of regulatory control over Al tolerance in plants [60,61,62]. Furthermore, the exogenous application of ABA could increase the activity of citrate synthase and decrease Al accumulation [60, 61]. In this study, three candidate genes (BnaA03g12450D in the vicinity of Bn-A03-p5766579, BnaA03g30670D in the vicinity of Bn-A03-p14798182, and BnaC04g06210D in the vicinity of Bn-C04-p4409586) related to ABA signal pathway were identified on chromosome A03 and C04, respectively (Fig. 7A). One candidate gene, BnaA03g12450D encodes ABA receptor PYL8 with up-regulation in 6 h vs 0 h condition in both ASL and ATL. In addition, various other defense related genes were detected including BnaA03g12780D in the vicinity of Bn-A03-p5766579, BnaA08g21350D in the vicinity of Bn-A08-p15773789 and BnaC03g28420D in the vicinity of Bn-C03-p16673270 which encode for F-box proteins, and BnaA06g27360D encodes zinc finger protein (Fig. 7A). A C2H2-type zinc finger protein STOP1 as the major factor regulating MATE1 expression plays a critical role in Al tolerance, and the F-box protein RAE1 regulates the stability of the Al-resistance transcription factor STOP1 [25, 63, 64]. Further research on these genes will reveal their roles under Al stress in B. napus.
Conclusions
A total of 43 SNP loci significantly associated with 8 phenotypic traits related to Al toxicity stress were detected on 14 chromosomes of B. napus by GWAS. Further exploration of SNP flanking regions discovered 777 candidate genes. RNA-seq approach detected 8291 and 5341 DEGs in ATL and ASL, respectively. Integration of GWAS and RNA-seq results found 64 differentially expressed candidate genes under 6 h and/or 24 h compared to control conditions. Among candidate DEGs, BnaA03g30320D and BnaA10g11500D encode MATE family protein and xyloglucan endotransglucosylase/hydrolase, respectively, which are responsive to Al toxicity stress. In addition, the proposed model showed that the oxidation-redox balance was perturbed under Al stress, causing abnormal cell wall repair and ABA signal transduction, ultimately leading to inhibition of root elongation. These exploratory analyses of Al toxicity linked candidate genes by integrating GWAS and RNA-seq showed a great power in uncovering genetic variation in Al toxicity stress in rapeseed. This strategy would be useful in understanding the molecular mechanisms responding Al toxicity. Furthermore, knowledge on the level of Al tolerance in rapeseed along with the associated SNPs from this research, would be useful for breeding future Al tolerant varieties.
Materials and methods
Plant materials and growth conditions
In this study, 254 oilseed rapeseed inbred lines were collected and preserved in the Key Laboratory of Crop Physiology, Ecology and Genetic Breeding (Jiangxi Agricultural University), Ministry of Education/Jiangxi Province. Of these oilseed rapeseed inbred lines, 220 lines were Semi-winter types, 15 lines were Spring types and 19 lines were Winter types. In total, 237 lines were collected from China, seven lines from Europe, five lines from Japan and five lines from Canada. The pertinent information for all accessions is shown in Table S7. These lines were grown under controlled conditions using growth chambers with 14 h light at 25 °C/20 °C (day/night) temperature.
Phenotyping for Al stress
Seeds of uniform size were selected from 254 accessions and separately surface sterilized in 1% hydrogen peroxide for 30 min [65]. Then, seeds were washed with ultrapure water for three times before spreading on the gauze cloth. In order to adapt the seedlings to total nutrient solution environment betterly, the uniform and healthy rapeseed seedlings before the lateral roots differentiation from the main roots were sequentially transferred into 1/4, 1/2 and total Hoagland’s nutrient solutions without Al treatment gradully. Each nutrient solution was cultured for 3 days. After transplanted into 0.5 mmol·L− 1 CaCl2 solution (pH 4.5) for 12 h, the seedlings were exposed to the nutrient solution (pH 4.5) containing 100 μmol·L− 1 AlCl3 for 28 days for Al stress treatment. The seedlings exposed to the total Hoagland’s nutrient solution (pH 4.5) with 0 μmol·L− 1 AlCl3 were used as a control. The pH of the solution was adjusted every 2 days and was kept 4.5 with NaOH or HCl. Each treatment had four biological replicates.
After 28 days, the fresh weight (g) of the above and below ground, and main root length (cm) were measured. Then, the root system was scanned by RhizoScan (Regent, Canada). The total root length (cm), root surface area (cm2), average root diameter (mm), root volume (cm3) and number of root tips of each material were analyzed by root image analysis software WinRHIZO STD4800 LA2400. The relative values (ratio) of each trait between treatments and controls were statistically analyzed using Excel and DPS, including relative fresh weight above ground (RFWAG), relative root average diameter (RRD), relative root fresh weight (RRFW), relative root elongation (RRE), relative total root surface area (RTRSA), relative total root tip (RTRT), relative total root volume (RTRV) and relative total root length (RTRL). The relative value was calculated by following formula: (the data of each phenotyping under Al treatment) / (the data of each phenotyping in control).
GWAS analysis
Based on the 200,510 SNPs developed in our previous research [33, 37], GWAS analysis for eight traits was carried out using generalized linear models (GLM) and mixed linear models (MLM) in Tassel 5.0 software [66]. GLM was adjusted using the Q-matrix which was calculated by the Admixture software package [67], MLM using Q-matrix and K-matrix was predicted by the SPAGeDi software [68]. The Quantile-Quantile plot (Q-Q plot) and the Manhattan plot were drawn by the GGplot2 software [69] and QQman software [70], respectively. The threshold value of -log10(P), set as -log10(1/200,510 SNPs), is approximately equal to 5.3 for significantly correlating SNPs.
To screen the candidate genes related to the Al tolerance, significant SNPs which were closely linked to the eight traits were mapped to the reference rapeseed genome [71]. The 100 kb flanking regions on either side of these SNPs were used to identify candidate genes. All candidate genes were selected based on GO (http://geneontology.org/), COG (https://www.ncbi.nlm.nih.gov/research/cog-project/), NR (ftp://ftp.ncbi.nih.gov/blast/db/FASTA/), SwissProt (http://www.expasy.org/sprot/) and KEGG (https://www.genome.jp/kegg/) databases.
RNA-seq under Al stress and data analysis
For RNA-seq, the Al -tolerant (ATL, FDH188) and -sensitive (ASL, FDH152) lines screened from 254 accessions in our previous research [72] were as the materials (Fig. S5). Two varieties were treated with 150 μmol·L− 1 AlCl3 for 0 h (control), 6 h and 24 h, respectively. Then the roots were quickly frozen in liquid nitrogen. Each treatment had three biological replicates.
To detect the DEGs, the low-quality reads with an ‘N’ percentage over 5% and more than 20% bases with a Q-value < 20 were removed by Perl program. The retained high quality reads were mapped to the reference rapeseed genome by Tophat [73], and then assembled by Cufflinks [74]. The genes expression levels were normalized by The Fragments Per Kb per Million fragments (FPKM) values. The genes with FPKM values ≤ 0.5 of all libraries were removed. False discovery rate (FDR) < 0.05 and log2(fold change (FC)) ≥ 1.0 or ≤ − 1.0 were used to determine the significantly DEGs. Combining the analysis of GWAS and RNA-seq, DEGs for Al tolerance within in the 100 kb intervals containing significant SNP markers will be selected as Al tolerance candidate genes.
The raw read data reported in this study have been deposited in the Genome Sequence Archive (GSA) in the National Genomics Data Center, under submission ID CRA003428 (https://ngdc.cncb.ac.cn/gsa/browse/CRA003428).
Validation of RNA-seq by qRT-PCR
Total RNA of 18 samples under 150 μmol·L-1 AlCl3 for 0 h, 6 h and 24 h between ATL and ASL were extracted by MiniBEST Universal RNA Extraction Kit (TaKaRa) followed by construction of cDNA libraries using PrimeScript™ RT Master Mix (TaKaRa). Expression of six DEGs was determined with TB Green® Premix Ex Taq™ II (TaKaRa) by eppendorf realplex2 (Eppendorf, Germany). The primer sequences for qRT-PCR are listed in Table S8. The relative expression levels were calculated using the 2-△△Ct method based on the normalization to the reference genes ACT6. Three technical replicates were performed for DEGs and reference genes.
Availability of data and materials
The datasets of RNA-seq during the current study are available in the Genome Sequence Archive (GSA) in the National Genomics Data Center repository (submission ID CRA003428, https://ngdc.cncb.ac.cn/gsa/browse/CRA003428). The data sets supporting the results of this article are included within the article and its additional files. The rapeseed cultivars and breeding materials are included within the article and its additional files (Table S7).
Abbreviations
- GWAS:
-
genome-wide association study
- ATL:
-
the Al-tolerant line
- ASL:
-
the Al-sensitive line
- B. napus :
-
Brassica napus
- DEGs:
-
the differentially expressed genes
- SLAF-seq:
-
specific-locus amplified fragment sequencing
- RFWAG:
-
relative fresh weight above ground
- RRD:
-
relative root average diameter
- RRFW:
-
relative root fresh weight
- RRE:
-
relative root elongation
- RTRSA:
-
relative total root surface area
- RTRT:
-
relative total root tip
- RTRV:
-
relative total root volume
- RTRL:
-
relative total root length
- FPKM:
-
The Fragments Per Kb per Million fragments
- FDR:
-
False discovery rate
- FC:
-
fold change
- GSA:
-
the Genome Sequence Archive
References
Lou HQ, Fan W, Jin JF, Xu JM, Chen WW, Yang JL, et al. A NAC-type transcription factor confers aluminum resistance by regulating cell wall-associated receptor kinase 1 and cell wall pectin. Plant Cell Environ. 2020;43(2):463–78.
Delhaize E, Ryan PR. Aluminum toxicity and tolerance in plants. Plant Physiol. 1995;107(2):315–21.
Ryan PR, Tyerman SD, Sasaki T, Furuichi T, Yamamoto Y, Zhang WH, et al. The identification of aluminium-resistance genes provides opportunities for enhancing crop production on acid soils. J Exp Bot. 2010;62(1):9–20.
Guo JH, Liu XJ, Zhang Y, Shen JL, Han WX, Zhang WF, et al. Significant acidification in major Chinese croplands. Science. 2010;327(5968):1008–10.
Kochian LV, Hoekenga OA, Piñeros MA. How do crop plants tolerate acid soils? Mechanisms of aluminum tolerance and phosphorous efficiency. Annu Rev Plant Biol. 2004;55(1):459–93.
Li M, Qin R, Jiang W, Liu D. Cytogenetical effects of aluminum on root meristem cells of helianthus annuus L. Bot Sci. 2015;93(1):15–22.
Da Rocha Oliveiros Marciano DP, Ramos FT, Alvim MN, Magalhaes JR, Costa Franca MG. Nitric oxide reduces the stress effects of aluminum on the process of germination and early root growth of rice. J Plant Nutr Soil Sc. 2010;173(6):885–91.
Doncheva S, Amenós M, Poschenrieder C, Barceló J. Root cell patterning: a primary target for aluminium toxicity in maize. J Exp Bot. 2005;56(414):1213–20.
Wood S, Sebastian K, Scherr S. Pilot analysis of global ecosystems: agroecosystems. Washington: World Resources Institute (WRI) and International Food Policy Reseach Institute (IFPRI); 2000. http://www.wri.org/wr2000.
von Uexkull H, Mutert E. Global extent, development and economic-impact of acid soils. Plant Soil. 1995;171:1–15.
Saeidnia S, Gohari AR. Importance of Brassica napus as a medicinal food plant. J Med Plants Res. 2012;6(14):2700–3.
Friedt W, Snowdon R. Oilseed rape. Oil Crops. 2009:91–126.
Gao H, Ye S, Wu J, Wang L, Wang R, Lei W, et al. Genome-wide association analysis of aluminum tolerance related traits in rapeseed (Brassica napus L.) during germination. Genet Resour Crop Evol. 2021;68(1):335–57.
Zhou G, Broughton S, Zhang XQ, Ma Y, Zhou M, Li C. Genome-wide association mapping of acid soil resistance in barley (Hordeum vulgare L.). front. Plant Sci. 2016;7:406.
Froese PS, Carter AH. Single nucleotide polymorphisms in the wheat genome associated with tolerance of acidic soils and aluminum toxicity. Crop Sci. 2016;56(4):1662–77.
Famoso AN, Zhao K, Clark RT, Tung CW, Wright MH, Bustamante C, et al. Genetic architecture of aluminum tolerance in rice (Oryza sativa) determined through genome-wide association analysis and QTL mapping. PLoS Genet. 2011;7(8):e1002221.
Yang JL, Fan W, Zheng SJ. Mechanisms and regulation of aluminum-induced secretion of organic acid anions from plant roots. J Zhejiang Univ Sci B. 2019;20(6):513–27.
Kochian LV, Piñeros MA, Liu J, Magalhaes JV. Plant adaptation to acid soils: the molecular basis for crop aluminum resistance. Annu Rev Plant Biol. 2015;66(1):571–98.
Wang Y, Xu H, Kou J, Shi L, Zhang C, Xu F. Dual effects of transgenic Brassica napus overexpressing CS gene on tolerances to aluminum toxicity and phosphorus deficiency. Plant Soil. 2013;362(1–2):231–46.
Ligaba A, Katsuhara M, Ryan PR, Shibasaka M, Matsumoto H. The BnALMT1 and BnALMT2 genes from rape encode aluminum-activated malate transporters that enhance the aluminum resistance of plant cells. Plant Physiol. 2006;142(3):1294–303.
Basu U, Good AG, Taylor GJ. Transgenic Brassica napus plants overexpressing aluminium-induced mitochondrial manganese superoxide dismutase cDNA are resistant to aluminium. Plant Cell Environ. 2001;24(12):1269–78.
Liu S, Zhao L, Liao Y, Luo Z, Wang H, Wang P, et al. Dysfunction of the 4-Coumarate:coenzyme a ligase 4CL4 impacts aluminum resistance and lignin accumulation in rice. Plant J. 2020;104(5):1233–50.
Yokosho K, Yamaji N, Fujii-Kashino M, Ma JF. Functional analysis of a MATE gene OsFRDL2 revealed its involvement in Al-induced secretion of citrate, but a lower contribution to Al tolerance in rice. Plant Cell Physiol. 2016;57(5):976–85.
Xu JM, Wang ZQ, Jin JF, Chen WW, Fan W, Zheng SJ, et al. FeSTAR2 interacted by FeSTAR1 alters its subcellular location and regulates Al tolerance in buckwheat. Plant Soil. 2019;436(1–2):489–501.
Fan W, Lou HQ, Gong YL, Liu MY, Cao MJ, Liu Y, et al. Characterization of an inducible C2H2-type zinc finger transcription factor VuSTOP1 in rice bean (Vigna umbellata) reveals differential regulation between low pH and aluminum tolerance mechanisms. New Phytol. 2015;208(2):456–68.
Liu Y, Hu G, Zhang A, Loladze A, Hu Y, Wang H, et al. Genome-wide association study and genomic prediction of Fusarium ear rot resistance in tropical maize germplasm. Crop J. 2021;9(2):325–41.
Ali A, Yoon-Hyun D, Noh T, Choi Y, Lee S, Oh S, et al. Preliminary genome-wide association mapping of rice bacterial leaf blight resistance loci using major Korean races of Xoo (Xanthomonas oryzae). Plant Omics. 2017;10(02):97–106.
Cai S, Wu D, Jabeen Z, Huang Y, Huang Y, Zhang G. Genome-wide association analysis of aluminum tolerance in cultivated and Tibetan wild barley. PLoS One. 2013;8(7):e69776.
Li Y, Ye H, Song L, Vuong TD, Song Q, Zhao L, et al. Identification and characterization of novel QTL conferring internal detoxification of aluminium in soybean. J Exp Bot. 2021;72(13):4993–5009.
Lu K, Xiao Z, Jian H, Peng L, Qu C, Fu M, et al. A combination of genome-wide association and transcriptome analysis reveals candidate genes controlling harvest index-related traits in Brassica napus. Sci Rep. 2016;6(1):36452.
Zhang J, Mason AS, Wu J, Liu S, Zhang X, Luo T, et al. Identification of putative candidate genes for water stress tolerance in canola (Brassica napus). Front Plant Sci. 2015;6:1058.
Wei L, Jian H, Lu K, Filardo F, Yin N, Liu L, et al. Genome-wide association analysis and differential expression analysis of resistance to Sclerotinia stem rot in Brassica napus. Plant Biotechnol J. 2016;14(6):1368–80.
Zhou Q, Zhou C, Zheng W, Mason AS, Fan S, Wu C, Fu D, Huang Y. Genome-wide SNP markers based on SLAF-seq uncover breeding traces in rapeseed (Brassica napus L.). Front Plant Sci. 2017;8:648.
Tan H, Xiang X, Tang J, Wang X. Nutritional functions of the funiculus in Brassica napus seed maturation revealed by transcriptome and dynamic metabolite profile analyses. Plant Mol Biol. 2016;92(4–5):539–53.
Zhang P, Zhong K, Zhong Z, Tong H. Mining candidate gene for rice aluminum tolerance through genome wide association study and transcriptomic analysis. BMC Plant Biol. 2019;19(1):490.
Zhang P, Zhong K, Tong H, Shahid MQ, Li J. Association mapping for aluminum tolerance in a core collection of rice landraces. Front Plant Sci. 2016;7:1415.
Zhou Q, Han D, Mason AS, Zhou C, Zheng W, Li Y, et al. Earliness traits in rapeseed (Brassica napus): SNP loci and candidate genes identified by genome-wide association analysis. DNA Res. 2018;25(3):229–44.
Wei L, Jian H, Lu K, Yin N, Wang J, Duan X, et al. Genetic and transcriptomic analyses of lignin- and lodging-related traits in Brassica napus. Theoretical Appl Genet Vol. 2017;130:1961–73.
Liu Y, Shi Q, Cao H, Ma Q, Nian H, Zhang X. Heterologous expression of a Glycine soja C2H2 zinc finger gene improves aluminum tolerance in Arabidopsis. Int J Mol Sci. 2020;21(8):2754.
Zhao L, Cui J, Cai Y, Yang S, Liu J, Wang W, et al. Comparative Transcriptome analysis of two contrasting soybean varieties in response to aluminum toxicity. Int J Mol Sci. 2020;21:4316.
Singh S, Tripathi DK, Singh S, Sharma S, Dubey NK, Chauhan DK, et al. Toxicity of aluminium on various levels of plant cells and organism: a review. Environ Exp Bot. 2017;137:177–93.
Degenhardt J, Larsen PB, Howell SH, Kochian LV. Aluminum resistance in the Arabidopsis mutant alr-104 is caused by an aluminum-induced increase in rhizosphere ph1. Plant Physiol (Bethesda). 1998;117(1):19–27.
Shen R, Ma JF. Distribution and mobility of aluminium in an Al-accumulating plant, Fagopyrum esculentum Moench. J Exp Bot. 2001;52(361):1683–7.
Li YY, Zhang YJ, Zhou Y, Yang JL, Zheng SJ. Protecting cell walls from binding aluminum by organic acids contributes to aluminum resistance. J Integr Plant Biol. 2009;51(6):574–80.
Ryan P, Delhaize E, Jones D. Function and mechanism of organic anion exudation from plant roots. Annu Rev Plant Physiol Plant Mol Biol. 2001;52:527–60.
Magalhaes JV, Liu J, Guimaraes CT, Lana UG, Alves VM, Wang YH, et al. A gene in the multidrug and toxic compound extrusion (MATE) family confers aluminum tolerance in sorghum. Nat Genet. 2007;39(9):1156–61.
Furukawa J, Yamaji N, Wang H, Mitani N, Murata Y, Sato K, et al. An aluminum-activated citrate transporter in barley. Plant Cell Physiol. 2007;48(8):1081–91.
Maron LG, Pineros MA, Guimaraes CT, Magalhaes JV, Pleiman JK, Mao C, et al. Two functionally distinct members of the MATE (multi-drug and toxic compound extrusion) family of transporters potentially underlie two major aluminum tolerance QTLs in maize. Plant J. 2010;61(5):728–40.
Yokosho K, Yamaji N, Ma JF. An Al-inducible MATE gene is involved in external detoxification of Al in rice. Plant J. 2011;68(6):1061–9.
Zhou Y, Wang Z, Gong L, Chen A, Liu N, Li S, et al. Functional characterization of three MATE genes in relation to aluminum-induced citrate efflux from soybean root. Plant Soil. 2019;443(1–2):121–38.
Zhu XF, Shi YZ, Lei GJ, Fry SC, Zhang BC, Zhou YH, et al. XTH31, encoding an in vitro xeh/xet-active enzyme, regulates aluminum sensitivity by modulating in vivo xet action, cell wall xyloglucan content, and aluminum binding capacity in Arabidopsis. Plant Cell. 2012;24(11):4731–47.
Yang JL, Li YY, Zhang YJ, Zhang SS, Wu YR, Wu P, et al. Cell wall polysaccharides are specifically involved in the exclusion of aluminum from the rice root apex1. Plant Physiol. 2008;146(2):602–11.
Kuromori T, Miyaji T, Yabuuchi H, Shimizu H, Sugimoto E, Kamiya A, et al. ABC transporter AtABCG25 is involved in abscisic acid transport and responses. Proc Natl Acad Sci U S A. 2010;107(5):2361–6.
Huang CF, Yamaji N, Mitani N, Yano M, Nagamura Y, Ma JF. A bacterial-type ABC transporter is involved in aluminum tolerance in rice. Plant Cell. 2009;21(2):655–67.
Kar D, Pradhan AA, Datta S. The role of solute transporters in aluminum toxicity and tolerance. Physiol Plant. 2020;171(4):638–52.
Larsen PB, Geisler MJB, Jones CA, Williams KM, Cancel JD. ALS3 encodes a phloem-localized ABC transporter-like protein that is required for aluminum tolerance in Arabidopsis. Plant J. 2005;41(3):353–63.
Lei GJ, Yokosho K, Yamaji N, Fujii-Kashino M, Ma JF. Functional characterization of two half-size ABC transporter genes in aluminium-accumulating buckwheat. New Phytol. 2017;215(3):1080–9.
Huang C, Yamaji N, Chen Z, Ma JF. A tonoplast-localized half-size ABC transporter is required for internal detoxification of aluminum in rice. Plant J. 2012;69(5):857–67.
Kumar M, Kesawat MS, Ali A, Lee SC, Gill SS, Kim A. Integration of abscisic acid signaling with other signaling pathways in plant stress responses and development. Plants (Basel). 2019;8(12):592.
Fan W, Xu JM, Wu P, Yang ZX, Lou HQ, Chen WW, et al. Alleviation by abscisic acid of Al toxicity in rice bean is not associated with citrate efflux but depends on ABI5-mediated signal transduction pathways. J Integr Plant Biol. 2019;61(2):140–54.
Hou N, You J, Pang J, Xu M, Chen G, Yang Z. The accumulation and transport of abscisic acid insoybean (Glycine max L.) under aluminum stress. Plant Soil. 2010;330(1–2):127–37.
Shen H, Ligaba A, Yamaguchi M, Osawa H, Shibata K. X Yan, Matsumoto H. effect of K-252a and abscisic acid on the efflux of citrate from soybean roots. J Exp Bot. 2004;55(397):663–71.
Zhang Y, Zhang J, Guo J, Zhou F, Singh S, Xu X, et al. F-box protein RAE1 regulates the stability of the aluminum-resistance transcription factor STOP1 in Arabidopsis. Proc Natl Acad Sci U S A. 2019;116(1):319–27.
Iuchi S, Koyama H, Iuchi A, Kobayashi Y, Kitabayashi S, Kobayashi Y, et al. Zinc finger protein STOP1 is critical for proton tolerance in Arabidopsis and coregulates a key gene in aluminum tolerance. Proc Natl Acad Sci U S A. 2007;104(23):9900–5.
Qin S, Sun X, Hu C, Tan Q, Zhao X, Xin J, Wen X. Effect of NO3-:NH4+ ratios on growth, root morphology and leaf metabolism of oilseed rape (Brassica napus L.) seedlings. Acta Physiol Plant. 2017;39:198.
Bradbury PJ, Zhang Z, Kroon DE, Casstevens TM, Ramdoss Y, Buckler ES. TASSEL: software for association mapping of complex traits in diverse samples. Bioinformatics. 2007;23(19):2633–5.
Alexander DH, Novembre J, Lange K. Fast model-based estimation of ancestry in unrelated individuals. Genome Res. 2009;19(9):1655–64.
HARDY OJ, VEKEMANS X. Spagedi: a versatile computer program to analyse spatial genetic structure at the individual or population levels. Mol Ecol Notes. 2002;2(4):618–20.
Ginestet C. ggplot2: Elegant graphics for data analysis. Journal of the Royal Statistical Society: Series A (Statistics in Society). 2011;174(1):245–246.
Turner SD. qqman: an R package for visualizing GWAS results using Q-Q and manhattan plots. bioRxiv. 2014:005165.
Chalhoub B, Denoeud F, Liu S, Parkin IA, Tang H, Wang X, et al. Plant genetics. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science. 2014;345(6199):950–3.
Han D, Liu X, Wang X, Luo S, Fu D, Zhou Q. Effects of aluminum stress on morphology parameters of roots and physiological indexes in Brassica napus L. J Nuclear Agricultural Sci. 2019;33(9):1824–32.
Trapnell C, Pachter L, Salzberg SL. TopHat: discovering splice junctions with RNA-Seq. Bioinformatics. 2009;25(9):1105–11.
Trapnell C, Williams BA, Pertea G, Mortazavi A, Kwan G, van Baren MJ, et al. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat Biotechnol. 2010;28(5):511–5.
Acknowledgments
The authors thank all contributors for their work and would like to thank the reviewers for their valuable comments and suggestions.
Funding
This work was supported by the National Natural Science Foundation of China project (31860392, 32160463), Jiangxi Provincial Key Research and Development Project (20181ACF60015).
Author information
Authors and Affiliations
Contributions
Qinghong Zhou and Yingjin Huang designed the research plan; Huiwen Zhou, Xiaojun Xiao, and Depeng Han performed the research work; Qinghong Zhou, Asjad Ali, Wei Zheng and Guobin Xiao analyzed the data; Huiwen Zhou, Asjad Ali and Qinghong Zhou wrote the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Prior to conducting the research, the permission from Key Laboratory of Crop Physiology, Ecology and Genetic Breeding (Jiangxi Agricultural University), Ministry of Education/Jiangxi Province to use the the rapeseed cultivars and breeding materials in this work was obtained. The current study complies with relevant institutional, national, and international guidelines and legislation for experimental research and field studies on plants (either cultivated or wild), including the collection of plant materials.
Consent for publication
Not applicable.
Competing interests
The authors declare that there are no conflicts of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zhou, H., Xiao, X., Asjad, A. et al. Integration of GWAS and transcriptome analyses to identify SNPs and candidate genes for aluminum tolerance in rapeseed (Brassica napus L.). BMC Plant Biol 22, 130 (2022). https://doi.org/10.1186/s12870-022-03508-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-022-03508-w