
Abstract
Background
The transfer of chloroplast DNA into nuclear genome is a common process in plants. These transfers form nuclear integrants of plastid DNAs (NUPTs), which are thought to be driving forces in genome evolution, including sex chromosome evolution. In this study, NUPTs in the genome of a dioecious plant Asparagus officinalis L. were systematically analyzed, in order to investigate the characteristics of NUPTs in the nuclear genome and the relationship between NUPTs and sex chromosome evolution in this species.
Results
A total of 3155 NUPT insertions were detected, and they represented approximated 0.06% of the nuclear genome. About 45% of the NUPTs were organized in clusters. These clusters were derived from various evolutionary events. The Y chromosome contained the highest number and largest proportion of NUPTs, suggesting more accumulation of NUPTs on sex chromosomes. NUPTs were distributed widely in all of the chromosomes, and some regions preferred these insertions. The highest density of NUPTs was found in a 47 kb region in the Y chromosome; more than 75% of this region was occupied by NUPTs. Further cytogenetic and sequence alignment analysis revealed that this region was likely the centromeric region of the sex chromosomes. On the other hand, the male-specific region of the Y chromosome (MSY) and the adjacent regions did not have NUPT insertions.
Conclusions
These results indicated that NUPTs were involved in shaping the genome of A. officinalis through complicated process. NUPTs may play important roles in the centromere shaping of the sex chromosomes of A. officinalis, but were not implicated in MSY formation.
Similar content being viewed by others


Background
In addition to nuclear genomes, two organellar genomes, namely, mitochondrial and plastid genomes, are found in plant cells. These three genomes coexist within each individual cell, where DNA is horizontally transferred between these cell parts, especially from the organelles to the nucleus [1, 2]. The extensive sequence transfer of organellar DNA into the nucleus produces nuclear integrants of plastid DNA (NUPTs) or nuclear integrants of mitochondrial DNAs (NUMTs). NUPTs and NUMTs provide new raw materials for genome complexity and remarkably contribute to chromosome evolution [3,4,5,6]. NUMTs exist widely in animals, plants, fungi, and protists [7, 8], whereas NUPTs only exist in plants. NUMTs have been extensively studied [9, 10], but NUPTs are still under research even though they have been explored in a number of plants [11].
NUPTs are widely distributed in plant species [8, 12]. Various properties, such as size, abundance, age, structure, and chromosomal localization of NUPTs have been analyzed in several model plant species [13, 14]. In Arabidopsis, approximately 18% of the total protein-coding genes are of chloroplast origin [13]. In Oryza sativa, large fragments of NUPTs are mainly distributed in the vicinity of centromeres. In addition, after plastid DNAs were integrated, these NUPTs were rapidly fragmented, shuffled, and most of them are discarded from the nuclear genome within a million years. These data in O. sativa indicate that the plant nuclear genome is in dynamic process of frequent integration and rapid elimination of the chloroplast genome and the pericentromeric regions are hot spots for the turnover of NUPTs [15]. The chromosomal distributions, correlation with transposable elements (TEs), and evolution dynamism of NUPTs show species-specific patterns. Although the integration mechanism of NUPTs is still not completely elucidated, non-homologous end joining and microhomology-mediated end joining of DNA double strand breaks are likely the mechanisms by which the organellar DNA fragments are integrated into the nucleus [16, 17]. NUPTs have also been cytogenetically analyzed in several plant species. For example, NUPT sites have been detected in 10 maize inbred lines through fluorescence in situ hybridization (FISH) analysis by using 14 overlapping fragments of the chloroplast genome. The integrations of NUPTs are recent and occurred frequently, as the distribution patterns vary greatly among different lines [18]. However, systematic analysis involving bioinformatics analysis and experimental cytogenetic studies in most plant species has yet to be performed.
Similar to TEs, inserted NUPTs can lead to the instability of the genome, such as causing chromosome structure rearrangement, size expansion, and heterochromatization [6, 19, 20]. Thus, the integration of NUPTs into the nuclear genome adds genetic diversity and is of physiological and evolutionary importance [21, 22]. Several studies revealed that NUPTs preferentially accumulate in the sex chromosomes of dioecious plants [23,24,25]. For example, in Silene latifolia, a bacterial artificial chromosome (BAC) clone containing a part of the chloroplast genome strongly hybridized to the Y chromosome, but its hybridization to the X chromosome and the autosomes is remarkably weak. The accumulation of these NUPTs may be one of the main factors of the size expansion of the Y chromosome in S. latifolia [23]. In another dioecious model plant, Carica papaya, NUPTs accumulated in the male-specific region of the Y chromosome (MSY) and the hermaphrodite-specific regions of Yh (HSY) are 12 times more than those in the X chromosome and four times more than those of the genome-wide average. The difference is caused by the proliferation, usually co-amplifying with surrounding retrotransposons of NUPTs in the MSY and HSY [25]. These studies reveal that the accumulation of NUPTs is strongly related to the evolution of plant sex chromosomes.
Among dioecious plants, Asparagus officinalis is an economically and nutritionally important vegetable crop that is cultivated worldwide. It has a haploid genome size of 1308 Mb with 2n = 2x = 20 chromosomes [26, 27]. The sex type of A. officinalis is determined by a pair of homomorphic sex chromosomes, namely, X and Y. A small MSY with size of 847 kb is detected recently [28]. In addition, the YY genotype (‘supermale’) is also viable [29]. These features indicate that the sex chromosomes of A. officinalis are in an early stage of sex chromosome evolution. The young sex chromosomes of A. officinalis can be used as a useful model to characterize the early stages of sex chromosome evolution. The recently sequenced A. officinalis genome provides a unique opportunity for the systematic investigation of NUPTs throughout the genome. In this study, we analyzed the amount, distribution pattern, and age of NUPTs by applying bioinformatics methods and investigated the cytogenetic chromosome localization of NUPTs through FISH analysis. The obtained data could provide a valuable basis for further studying the genome structure and sex chromosome evolution of A. officinalis.
Results
Identification of NUPTs in the nuclear genome of A. officinalis
We analyzed the chloroplast DNA sequences integrated in the nuclear genome of A. officinalis. A total of 3155 NUPT insertions were identified. The cumulative length of these NUPTs was 703,058 bp, constituting approximately 0.06% of the total nuclear genome. The identity of these matched nuclear sequences with the chloroplast DNA (cpDNA) sequence ranged from 71.81–100%. The NUPT sequence length varied from 33 to 8733 bp with a mean length of 223 bp (Additional file 1: Table S1). The distribution of NUPTs according to size is presented in Fig. 1. Generally, the number of NUPTs decreased sharply with increased size range. Approximately 72.8% of the NUPTs had a size of 33–199 bp, where 1120 small NUPTs are shorter than 100 bp and 1176 NUPTs had a size ranging from 100 to 199 bp. A total of 76 NUPTs (2.4%) had a large size of longer than 1000 bp. Among these long NUPTs, 19 are localized on chromosome 1, i.e. the Y chromosome [28].

Size distribution of NUPTs in the nuclear genome of A. officinalis
The distribution of NUPT sequences at different chromosomes and their positions were analyzed (Table 1, Fig. 2). Chromosome 1 (Y chromosome) presented the largest proportion of NUPTs and the largest number of detected NUPTs. A total of 440 NUPTs were localized on chromosome 1, accounting for 13.95% of the total NUPTs detected. The total length of NUPTs in chromosome 1 was 114,469 bp, accounting for approximately 0.09% of its size. Both the total and standard length of NUPTs (86,463 bp/100 Mb) of chromosome 1 were larger than those of other chromosomes. By contrast, chromosome 10 had the least NUPTs (169). However, considering the chromosome size, chromosome 4 was covered by the lowest proportion of NUPTs (~ 0.047%) (Table 1).

Distribution of NUPTs on 10 chromosomes of A. officinalis. The length of vertical lines indicates the length of the insertion, and each window size is 200 kb. Individual chromosomes are shown in separate panels
In the macroscopic scale, NUPTs were widely distributed throughout each chromosome (Fig. 2). However, the amount of NUPTs varied among different chromosomal regions. Some chromosomal regions clearly favor NUPT insertion. Within each sliding window of 200 kb, a total length of NUPTs > 5000 bp was found in 11 regions (Fig. 2) Chromosomes 3, 5, 6, and 7 each possessed two such regions, whereas chromosomes 1, 4, and 9 each had one such region. The highest NUPT coverage region was found within chromosome 1, where 35,406 bp sequences were detected as NUPTs within a 200 kb region. Detailed analysis of this region showed that NUPTs were mainly distributed in a region of less than 47 kb (locus site: 54519138–54,565,922). Within this 46,785-bp region, 35,328 bp sequences were detected as NUPTs.
Interestingly, although the Y chromosome (chromosome 1) possessed the most number of NUPTs, the MSY (locus site: 3411433–4,258,465) and the adjacent regions did not show NUPT insertion (Fig. 3), which implies that NUPTs may not be involved in the formation of MSY in A. officinalis.

Distribution of NUPTs on the first 8 Mb of the Y chromosome. The MSY region is indicated by grey background
The chromosome distribution analysis of NUPTs revealed that they were often clustered. We then conducted cluster analysis wherein two NUPTs are assigned to one cluster if they are separated by less than 5 kb. We found that 1396, or nearly 45% of the asparagus NUPTs were organized in such clusters (Additional file 1: Table S1). These NUPTs were assigned to 484 clusters, where the largest cluster (cluster 236) contained 26 NUPTs in chromosome 5. Another large cluster (cluster 27) had 25 NUPTs in chromosome 1 (Additional file 1: Table S1). Some typical clusters were visualized in dot-plots as shown in Fig. 4. NUPT clusters representing different insertion patterns were observed. These clusters were usually derived through unique insertional events, which underwent subsequent insertions, leading to separate, but tightly linked NUPTs identified as clusters (Fig. 4, cluster 339). The NUPT clusters were also frequently derived from several regions of the chloroplast DNA, each of which undergoes rearrangement with the evolution process (Fig. 4, clusters 27 and 294). We also identified some NUPT clusters organized by individual NUPT insertions, which originated from nearly identical chloroplast DNA region (Fig. 4, cluster 447). In addition, some NUPT clusters were derived from various regions of the chloroplast DNA, making these regions as chromosome hotspots of NUPT insertions (Fig.4, clusters 236 and 429).

Dot-plot analysis of some typical NUPT clusters. NUPT cluster names are reported at the top of each plot. X axis reports the positions in the indicated chromosomes. Y axis reports the chloroplast sequence coordinates. Colored lines in the plots represent different NUPT sequences. Details about these regions are reported in Additional file 1: Table S1
Distribution of NUPTs according to their origin from the chloroplast genome
To understand the transfer pattern of chloroplast DNA into the nuclear genome of A. officinalis, we analyzed the origin of the NUPTs. Among the 3155 NUPTs, 571 were derived from inverted repeat (IR), 2166 from large single copy region (LSC), and 418 from small single copy region (SSC) (Table 2). The sizes of IR, LSC, and SSC were 52,942, 83,821, and 17,901 bp, respectively, while the densities of NUPTs originated from IR, LSC, and SSC were 11/kb, 26/kb, and 23/kb, respectively (Table 2). It is obvious that DNAs from the LSC region were more frequently transferred to the nuclear genome. Further analysis revealed that the high frequency of transfer from the LSC region was mainly due to a small region with a size of 286 bp (locus site: 50,003–50,288). A total of 443 NUPTs were derived from this region, accounting for 14% of the total number of NUPTs. The cumulative length of NUPTs originated from this region was 65,490 bp. Evidently, the sequences of the 286-bp region presented an unexpectedly high transfer frequency to nuclear genome (Fig. 5).

Distribution of NUPT sequences on the chloroplast genome. A sliding window of 500 bp was used to calculate for the total length of the inserted NUPTs. The two IR regions are marked by grey background
Insertion age of NUPTs
We used the p-distance to calculate for the relative insertion age of each NUPT. Both for the number and the cumulative length of NUPTs had a high peak in the p-distance interval of 0–0.01 (Fig. 6), which represented the youngest age. Then, with the increase in p-distance, the number and cumulative length of NUPTs decreased, except that a small peak was observed in the p-distance interval of 0.23–0.25 (Fig. 6). This peak is mainly due to the high transfer frequency of the 286-bp sequence within the LSC region, because the majority of the NUPTs derived from the 286-bp region showed large p-distance from 0.23–0.25. Generally, the evolution time of NUPTs increased with the decrease in the length of NUPTs. The mean p-distance of NUPTs was 0.10, whereas that of NUPTs with length between 500 and 1000 bp was 0.07, and that of NUPTs longer than 1000 bp was only 0.05. The mean insertion age and mean length of NUPTs slightly differed among the different chloroplast DNA origins. The NUPTs that originated from the LSC region showed a mean p-distance of 0.11 and mean length of 218 bp. The mean insertion age of NUPTs derived from the IR and SSC regions was slightly low with a mean p-distance of 0.09. The average sizes of NUPTs derived from the IR and SSC regions were 238 and 225 bp, respectively (Table 2).

Age distribution of NUPTs in the nuclear genome of A. officinalis. The numbers (a) and cumulative length (b) of NUPTs for each p-distance interval are shown
Chromosome localization of NUPTs
A total of 32 pairs of primers were designed for overlapping chloroplast fragments with an average desired fragment length of approximately 4051 bp. Finally, 28 chloroplast fragments were amplified successfully. Among these fragments, 17, 8, and 3 belonged to the LSC, IR, and SSC region, respectively.
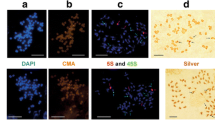
FISH analysis showed that 24 of the 28 probes could generate stable and clear signals on A. officinalis chromosomes. Among these, 12 probes were dispersed in almost all the chromosomes (Fig. 7). AoLSC5, AoLSC15, AoLSC18, AoLSC19, and AoIR7 showed evenly distributed signals on all the chromosomes, whereas AoLSC6, AoLSC7, AoLSC10, AoLSC11, AoLSC14, and AoSSC4 were more inclined to distribute in blocks along the chromosomes (Fig. 7). AoLSC17 showed weak signals mainly at the pericentromeric regions of all chromosomes. AoLSC1, AoLSC9, and AoIR1 mainly occupied the centromeric regions, whereas AoIR4 was distributed in the centromeric and adjacent pericentromeric regions of all the chromosomes (Fig. 7).

Chromosome localization of chloroplast DNA on the metaphase chromosomes of A. officinalis (signals widely distributed on all chromosomes). Each chloroplast DNA probe was labeled with Texas red (red signal), 45S rDNA was labeled with Chroma Tide Alexa Fluor 488 (green signal), and the chromosomes were counterstained with DAPI (blue). Bars = 10 μm
The other eight probes showed more intense signals on specific chromosomes, among which, two probes (AoSSC3 and AoLSC4) were mainly distributed in the centromeric regions of chromosome 2 (Fig. 8). AoSSC3 was exclusively localized in the centromeric regions of chromosome 2 (Fig. 8a, c), while AoLSC4 was distributed in the other chromosomes, in addition to chromosome 2 (Fig. 8b, d).

Chromosome localization of two chloroplast DNA fragments on metaphase chromosomes and karyotype analysis. a FISH result of AoLSC4; (b) FISH result of AoSSC3; (c) Karyotypic analysis of AoLSC4; (d) Karyotypic analysis of AoSSC3. Arrowheads indicate the sites of chloroplast DNA probe hybridization (red). Bars = 10 μm
More importantly, AoLSC2, AoIR2, AoIR3, and AoIR5 were all localized in the centromeric regions of one pair of chromosomes (Fig. 9a). Karyotype analysis shows that the four probes were localized on the same pair of chromosomes, i. e. the chromosome 5 (Fig. 9b). Previous studies reported that the pair of chromosome 5 in the karyotype analysis is the sex chromosome of A. officinalis [30, 31]. Genome sequence and analysis showed chromosome 1 of genome sequences is the sex chromosome. Thus, the chromosome 5 in the karyotype analysis is corresponding to chromosome 1 of the genome sequence. We also conducted BLAST analysis to investigate the distribution patterns of the NUPTs derived from the four chloroplast DNA fragments in the genome of A. officinalis. The results showed that NUPTs that originated from the four chloroplast DNA fragments were mainly distributed in adjacent regions in chromosome 1 of genome assembly (Fig. 9c). These sequences were concentrated on a region around 54–55 Mb of chromosome 1, which is within the region with maximum density of NUPTs, more than half of which is occupied by NUPTs (Fig. 2). Combined with the FISH results that these sequences were mainly located at the centromeric region of the sex chromosomes, the results suggested that the 54–55 Mb region should be within or around the centromeric region of the sex chromosome, and the centromeric regions of the sex chromosome of A. officinalis were occupied by plenty of NUPTs.

Cytogenetic and sequence alignment analysis of the four chloroplast DNA fragments localized on the centromeric regions of sex chromosomes. a FISH analysis of the four chloroplast DNA sequences. Arrowheads indicate the sites of chloroplast DNA probe hybridization (red). Bars = 10 μm; (b) Karyotypic analysis based on FISH results; (c) Enlarged sex chromosomes with FISH signals and BLAST analysis of these four chloroplast DNA sequences
Discussion
During evolution process, plant nuclear genomes have been shaped by the acquisition of new sequences of various origins. These sequences include TEs and the sequences derived from mitochondrial DNA (NUMTs) and cpDNA (NUPTs). TEs have been widely investigated in many plant genomes, and showed to play various roles in genome evolution, chromosome rearrangement, gene creation and regulation [32,33,34,35,36]. NUMTs have been studied extensively in animals [7, 37, 38]. By contrast, NUPTs only exist in plants and have been studied in a limited number of plant species [6, 31, 39]. In this study, systematic analyses and cytogenetic localization of NUPTs were conducted on a dioecious plant A. officinalis. The proportion of NUPTs in the asparagus genome (0.06%) is within the range observed in other plants. For example, the percentages of NUPTs in current analyzed nuclear genomes vary from 0.024% in Sorgum bicolor to 0.222% in O. sativa [40]. The NUPT proportion in the nuclear genome depends on the balance of the frequency of DNA migration from the plastids to the nuclear genome and the rate of NUPT loss in the nuclear genome [39]. It is also influenced by the assembly and annotation quality of the genome and the methods used in the analyses.
All the plastid DNA sequences can transfer to the nuclear genome of A. officinalis, indicating that all sequences are transferable. However, the transfer frequency from different parts of the plastid genome varied. The LSC region showed a high transfer rate, which is largely due to the 286-bp sequence with unexpectedly high transfer frequency. In most of the studied plant genomes, although the transfer rate varies among different regions of the chloroplast genome, no regions had an extremely low or high transfer frequency [40]. Thus, such high transfer frequency of a small size of region has not been reported in other species. Owing that nearly all the inserted NUPTs that originated from this 286-bp region showed similar and relatively large p-distance (ranging from 0.19–0.24 with only one exception), we speculate that these NUPT insertions were old-aged and may be formed through a sudden event caused by stress. In fact, experimental studies confirmed that stress can promote DNA migration from the chloroplasts to the nucleus. For example, DNA migration from chloroplasts to the nucleus is markedly increased by mild heat stress in two transplastomic tobacco lines [22].
Cluster analysis of NUPTs revealed that nearly 45% of the NUPTs were organized in clusters. Further dot-plot analysis indicated that these clusters might be formed through different evolutionary paths, implying complicated mechanisms shaping the genome of A. officinalis. A considerable number of NUPT sequences could be derived via a single or two insertional events that subsequently underwent mutations and chromosomal rearrangements. Mutations and chromosomal rearrangements can split the integrated cpDNA in separate, but co-linear regions. Such clusters derived from initially long cpDNA insertion and subsequent fragmentation have also been widely observed in organelle-derived insertions in other species [6, 41]. Long integrated NUPTs are assumed to be young and recent insertions, whereas short fragments are usually old insertions [42,43,44]. The relative age analyses results of NUPTs in A. officinalis are consistent with this assumption. In addition, the assumption is well supported by the co-linear cluster regions observed widely in the genome of A. officinalis. Long NUPTs might be degraded and fragmented by mutations and chromosomal rearrangements with the ongoing evolutionary process. NUPTs organized in clusters were also frequently derived from different regions of the chloroplast DNA. These chromosome regions recruited a large number of NUPT sequences, but no co-linear pattern was investigated. Such complicated regions might be formed through various direct NUPT insertions, implying that these regions could be more attractive for cpDNA integration. This phenomenon was probably due to the features of the chromosomal regions themselves, such as the condensation state of the region. For example, the pericentromeric regions which usually presenting dense heterochromatin state are hotspots for organelle-nucleus DNA transfer [15]. Alternatively, these regions might be formed by genome shuffling through chromosome rearrangement.
NUPTs are distributed unevenly within the chromosomes of A. officinalis. Some regions prefer NUPT insertion. We discovered a 47 kb region, where more than 75% was occupied by NUPTs. This biased distribution pattern was also discovered in other analyzed plant genomes. For example, some chromosomal regions that are rich in NUPTs were found in Lotus japonicus, O. sativa, and Zea mays [40]. It has been reported that NUPTs are frequently distributed in pericentromeric regions, especially in species with a small genome size, whereas NUPTs have a wide distribution pattern in species with large genome sizes [6]. The location of the centromeres was unknown in the assembled chromosomes of A. officinalis. However, FISH analysis revealed that four chloroplast DNA sequences were preferentially distributed in the centromeric and adjacent regions of all the chromosomes. In addition, four fragments were exclusively located in the centromeric regions of the sex chromosomes, and two other sequences mainly occupied the centromeric regions of chromosome 2. These findings suggested that NUPTs likely more accumulated in centromeric or pericentromeric regions in A. officinalis. Centromeric and pericentromeric regions are hotspots of organelle–nucleus DNA transfer [15, 39], because of the low gene density and high heterochromatin content of these regions [43,44,45,46], which may provide a stable genomic environment for the integration of the organelle-derived DNA [6, 15]. However, Yoshida et al. showed that NUPTs also accumulate in the distal regions of chromosomes [38]. This finding might imply that in addition to pericentromeric regions, other regions also have high tolerance to NUPTs.
Both bioinformatic and cytogenetic analyses showed that the sex chromosomes of A. officinalis contained more NUPTs. However, the MSY and adjacent regions did not have NUPT insertion. These results indicated that NUPTs might facilitate the shaping of sex chromosome, but were not involved in the MSY evolution. Studies have shown that NUPTs preferentially accumulate on the sex chromosomes of R. acetosa, S. latifolia, and C. papaya [23,24,25]. These transferred organellar DNA sequences are possibly one of the driving forces in the sex chromosome evolution of dioecious plants. It is universally believed that sex chromosomes are derived from pairs of autosomes [47]. During the evolution process, sex chromosomes can recruit a large number of TEs and organellar DNAs [25, 48], and these sequences may facilitate the structural evolution of sex chromosomes. For example, the inserted TEs and organellar DNAs may lead to structural and morphological differentiation of sex chromosomes, and contribute to the degeneration of Y chromosomes [49]. However, the involvement of NUPTs in the MSY formation is poorly studied. In C. papaya, NUPTs that accumulate in HSY and MSY are much more than those in the X chromosome and the autosomes [25]. Furthermore, organellar DNA insertions are sparse between X and HSY, suggesting that the organellar DNA accumulates after the recombination of HSY is suppressed. The accumulation of NUPTs in papaya has contributed to the dramatic expansion and repeated accumulation in HSY [25]. In this study, NUPTs did not accumulate in the MSY of A. officinalis, indicating that NUPTs did not contribute to the formation and recombination suppression of MSY in this dioecious plant. The sex chromosomes of A. officinalis are very young. These results are consistent with the findings in papaya, whose NUPT insertions occur after the HSY is suppressed for recombination. However, the sex chromosomes do harbour more NUPTs than the autosomes. The mechanism and function of these NUPT insertions in the recombining regions of sex chromosomes are still not clear currently. One reasonable explanation is that although the sequences outside the MSY are usually not related to sex determination, they also suffered sexual selection because of linkage disequilibrium and may show different evolutionary rate compared with the sequences on the autosomes. In fact, the mechanism of sex determination and sex chromosome evolution may be far more complex than traditionally appreciated and usually unpredictable. For example, the recent discovered gene MpFGMYB, which encodes a key regulator of female sexual differentiation in liverwort, localized in autosomes, not sex chromosomes [50]. Full understanding of these processes involved in sex determination and sex chromosome evolution need further studies and accumulation of more data. Currently, the X chromosome of A. officinalis has not been sequenced. In the future, with the sequence of the X chromosome available, a comparison between the sequences of the X and Y chromosomes can be conducted. More information about the early stages of sex chromosome evolution can be obtained.
Conclusion
In conclusion, this study presented a comprehensive bioinformatics and cytogenetic analysis of NUPTs in the nuclear genome of A. officinalis, an important vegetable and model dioecious plant species for studying the early stages of sex chromosome evolution. More than 45% of the NUPTs were organized in tight clusters, mainly including separate but co-linear clusters and promiscuous insertional clusters with no co-linear pattern. The large differences among the NUPT origins suggest that the integrated regions might be formed through different evolutionary paths and NUPTs were involved in shaping the genome of A. officinalis through complicated mechanisms. NUPTs preferably accumulated on the sex chromosomes of A. officinalis but not at the MSY. Hence, NUPTs might play a role in the shaping of sex chromosome structure but were irrelevant to the MSY formation. Our analysis provided new insights into the genome structure and evolution of A. officinalis and established a basis for further studying the sex chromosome evolution of A. officinalis.
Methods
Plant materials
A. officinalis cultivar ‘UC309’ grown in the garden field of Henan Normal University was used for the experiments. The original seeds were bought from Henan Yinong Company in 2005. One male and one female plant generated from the seeds were crossed to produce F1. The F2 individuals generated from the F1 and the corresponding seeds were used in this study.
Data sources
The nuclear genome of A. officinalis was downloaded from https://www.ncbi.nlm.nih.gov/genome/?term=10978 [28], and the sequence of the chloroplast genome was provided by Dr. Pires [51]. The sizes of the nuclear and chloroplast genomes were 1187.54 Mbp and 154,664 bp, respectively.
Detection of NUPTs
NUPT insertions were identified using the BLASTN local alignment tools in the BLAST program package (ver. 2.2.31) with chloroplast genomic DNA as the query sequence and nuclear genome data as the database. The parameters were as follows: -dust no, e-value threshold of 1e-4, mismatch penalty of − 2, and word size of 9. The BLAST hits for the NUPTs that originated from the IR region of the chloroplast genome were counted only once because these BLAST hits were obtained in both IR regions and could not be distinguished. The NUPTs distributed on the nuclear chromosomes and originating from the cpDNA were visualized based on slide window lengths of 200 kb and 500 bp, respectively.
Estimation of NUPT age distribution
The p-distance between each NUPT and the corresponding chloroplast DNA sequence was calculated to estimate the relative integration time. p = nd/n, where nd is the number of nucleotide differences, and n is the number of aligned nucleotides [10].
PCR amplification of cpDNA regions
The total genomic DNA was isolated from young leaves of Asparagus officinalis. To obtain overlapping fragments representing the entire chloroplast genome of A. officinalis, the primers were designed using OLIGO 7 program [52]. The primer sets are listed in Additional file 2: Table S2. A total of 32 chloroplast DNA PCR products were amplified from the total extracted genomic DNA, containing the chloroplast DNA. The PCR amplification fragments were verified by sequencing the 5′- and 3′-end sequences.
FISH procedure
Slide preparation using mitotic metaphase spreads from the root tips, probe-labeling, hybridization, and picture-processing were done as previously described [53]. The individual chloroplast regions were labeled with Texas Red-5-dCTP (shown in red) and hybridized at a final concentration of 20 ng/μL each. The 45S rDNA was labeled with fluorescein-488-dUTP (shown in green), and double-color FISH was performed for a better characterization of the chromosomes. After hybridization and washing, all slides were treated with Vectashield (Vector Laboratories, Burlingame, Calif.) containing 4′,6-diamidino-2-phenylindole (DAPI) prior to visualization.
Availability of data and materials
The nuclear genome of A. officinalis was downloaded from https://www.ncbi.nlm.nih.gov/genome/?term=10978. The data and plant material that support the findings of this study are available from the corresponding author on request. All data generated or analyzed during this study are included in this published article.
Abbreviations
- BAC:
-
Bacterial artificial chromosome
- cpDNA:
-
chloroplast DNA
- DAPI:
-
4′,6-diamidino-2-phenylindole
- FISH:
-
Fluorescence in situ hybridization
- HSY:
-
Hermaphrodite-specific regions of Yh
- IR:
-
Inverted repeat
- LSC:
-
Large single copy region
- MSY:
-
Male-specific region of the Y chromosome
- NUMTs:
-
Nuclear integrants of mitochondrial DNAs
- NUPTs:
-
Nuclear integrants of plastid DNAs
- SSC:
-
Small single copy region
- TEs:
-
Transposable elements
References
Lewin R. No genome barriers to promiscuous DNA; the movement of DNA sequences between mitochondrial, chloroplast and nuclear genomes is even more prolific than had been expected. Science. 1984;224:970–1.
Martin W, Herrmann RG. Gene transfers from organelles to the nucleus: how much, what happens, and why? Plant Physiol. 1998;118:9–17.
Chen H, Yu Y, Chen X, Zhang Z, Gong C, Li J, Wang A. Plastid DNA insertions in plant nuclear genomes: the sites, abundance and ages, and a predicted promoter analysis. Funct Integr Genomics. 2015;15:131–9.
Lough AN, Roark LM, Kato A, Ream TS, Lamb JC, Birchler JA, Newton KJ. Mitochondrial DNA transfer to the nucleus generates extensive insertion site variation in maize. Genetics. 2008;178:47–55.
Martis MM, Klemme S, Banaei-Moghaddam AM, Blattner FR, Macas J, Schmutzer T, Scholz U, Gundlach H, Wicker T, Šimková H, Novák P, Neumann P, Kubaláková M, Bauer E, Haseneyer G, Fuchs J, Doležel J, Stein N, Mayer KFX, Houben A. Selfish supernumerary chromosome reveals its origin as a mosaic of host genome and organellar sequences. P Natl Acad Sciences USA. 2012;109:13343–6.
Michalovova M, Vyskot B, Kejnovsky E. Analysis of plastid and mitochondrial DNA insertions in the nucleus (NUPTs and NUMTs) of six plant species: size, relative age and chromosomal localization. Heredity. 2013;111:314–20.
Bensasson D, Zhang DX, Hartl DL, Hewitt GM. Mitochondrial pseudogenes: evolution’s misplaced witnesses. Trends Ecol Evol. 2001;16:314–21.
Hazkani-Covo E, Zeller RM, Martin W. Molecular poltergeists: mitochondrial DNA copies (numts) in sequenced nuclear genomes. PLoS Genet. 2010;6:e1000834.
Ko YJ, Kim S. Analysis of nuclear mitochondrial DNA segments of nine plant species: size, distribution, and insertion loci. Genomics Inform. 2016;14:90–5.
Yoshida T, Furihata HY, Kawabe A. Analysis of nuclear mitochondrial DNAs and factors affecting patterns of integration in plant species. Genes Genet Syst. 2017;92:27–33.
Smith DR, Crosby K, Lee RW. Correlation between nuclear plastid DNA abundance and plastid number supports the limited transfer window hypothesis. Genome Biol Evol. 2011;3:365–71.
Ayliffe MA, Scott NS, Timmis JN. Analysis of plastid DNA-like sequences within the nuclear genomes of higher plants. Mol Biol Evol. 1998;15:738–45.
Martin W, Rujan T, Richly E, Hansen A, Cornelsen S, Lins T, Leister D, Stoebe B, Hasegawa M, Penny D. Evolutionary analysis of Arabidopsis, cyanobacterial, and chloroplast genomes reveals plastid phylogeny and thousands of cyanobacterial genes in the nucleus. P Natl Acad Sciences USA. 2002;99:12246–51.
Shahmuradov IA, Akbarova YY, Solovyev VV, Aliyev JA. Abundance of plastid DNA insertions in nuclear genomes of rice and Arabidopsis. Plant Mol Biol. 2003;52:923–34.
Matsuo M, Ito Y, Yamauchi R, Obokata J. The rice nuclear genome continuously integrates, shuffles, and eliminates the chloroplast genome to cause chloroplast-nuclear DNA flux. Plant Cell. 2005;17:665–75.
Yu X, Gabriel A. Patching broken chromosomes with extranuclear cellular DNA. Mol Cell. 1999;4:873–81.
Guo X, Ruan S, Hu W, Cai D, Fan L. Chloroplast DNA insertions into the nuclear genome of rice: the genes, sites and ages of insertion involved. Funct Integr Genomics. 2008;8:101–8.
Roark LM, Hui AY, Donnelly L, Birchler JA, Newton KJ. Recent and frequent insertions of chloroplast DNA into maize nuclear chromosomes. Cytogenet Genome Res. 2010;129:7–23.
Noutsos C, Richly E, Leister D. Generation and evolutionary fate of insertions of organelle DNA in the nuclear genomes of flowering plants. Genome Res. 2005;15:616–28.
Sheppard AE, Timmis JN. Instability of plastid DNA in the nuclear genome. PLoS Genet. 2009;5:e1000323.
Huang CY, Ayliffe MA, Timmis JN. Simple and complex nuclear loci created by newly transferred chloroplast DNA in tobacco. P Natl Acad Sciences USA. 2004;101:9710–5.
Wang D, Lloyd AH, Timmis JN. Environmental stress increases the entry of cytoplasmic organellar DNA into the nucleus in plants. P Natl Acad Sciences USA. 2012;109:2444–8.
Kejnovsky E, Kubat Z, Hobza R, Lengerova M, Sato S, Tabata S, Fukui K, Matsunaga S, Vyskot B. Accumulation of chloroplast DNA sequences on the Y chromosome of Silene latifolia. Genetica. 2006;128:167–75.
Steflova P, Hobza R, Vyskot B, Kejnovsky E. Strong accumulation of chloroplast DNA in the Y chromosomes of Rumex acetosa and Silene latifolia. Cytogenet Genome Res. 2014;142:59–65.
VanBuren R, Ming R. Organelle DNA accumulation in the recently evolved papaya sex chromosomes. Mol Gen Genomics. 2013;288:277–84.
Arumuganathan K, Earle ED. Nuclear DNA content of some important plant species. Plant Mol Biol Rep. 1991;9:208–18.
Ellison JH. Asparagus breeding. In: Bassett MJ, editor. Breeding vegetable crops. Westport: AVI; 1986. p. 523–68.
Harkess A, Zhou J, Xu C, Bowers JE, der Hulst RV, Ayyampalayam S, Mercati F, Riccardi P, MR MK, Kakrana A, Tang H, Ray J, Groenendijk J, Arikit S, Mathioni SM, Nakano M, Shan H, Telgmann-Rauber A, Kanno A, Yue Z, Chen H, Li W, Chen Y, Xu X, Zhang Y, Luo S, Chen H, Gao J, Mao Z, Pires JC, Luo M, Kudrna D, Wing RA, Meyers BC, Yi K, Kong H, Lavrijsen P, Sunseri F, Falavigna A, Ye Y, Leebens-Mack JH, Chen G. The asparagus genome sheds light on the origin and evolution of a young Y chromosome. Nat Commun. 2017;8:1279.
Vyskot B, Hobza R. Gender in plants: sex chromosomes are emerging from the fog. Trends Genet. 2004;20:432–8.
Löptien H. Identification of the sex chromosome pair in asparagus (Asparagus officinalis L.). Z Pflanzenzuechtung. 1979;82:162–73.
Uno Y. Haploid production from polyembryonic seeds of Asparagus officinalis L. Acta Hort. 2002;589:217–21.
Cho J, Paszkowski J. Regulation of rice root development by a retrotransposon acting as a microRNA sponge. Elife. 2017;6:e30038.
Bennetzen JL, Wang H. The contributions of transposable elements to the structure, function, and evolution of plant genomes. Annu Rev Plant Biol. 2014;65:505–30.
Harkess A, Mercati F, Abbate L, McKain M, Pires JC, Sala T, Sunseri F, Falavigna A, Leebens-Mack J. Retrotransposon proliferation coincident with the evolution of dioecy in Asparagus. G3–Genes Genom Genet. 2016;6:2679–85.
Li SF, Su T, Cheng GQ, Wang BX, Li X, Deng CL, Gao WJ. Chromosome evolution in connection with repetitive sequences and epigenetics in plant. Genes. 2017;8:290.
Ustyantsev K, Blinov A, Smyshlyaev G. Convergence of retrotransposons in oomycetes and plants. Mob DNA. 2017;8:4.
Hazkani-Covo E, Graur D. A comparative analysis of numt evolution in human and chimpanzee. Mol Biol Evol. 2007;24:13–8.
Schiavo G, Strillacci MG, Ribani A, Bovo S, Roman-Ponce SI, Cerolini S, Bertolini F, Bagnato A, Fontanesi L. Few mitochondrial DNA sequences are inserted into the Turkey (Meleagris gallopavo) nuclear genome: evolutionary analyses and informativity in the domestic lineage. Anim Genet. 2018;49:259–64.
Richly E, Leister D. NUPTs in sequenced eukaryotes and their genomic organization in relation to NUMTs. Mol Biol Evol. 2004;21:1972–80.
Yoshida T, Furihata H, Kawabe A. Patterns of genomic integration of nuclear chloroplast DNA fragments in plant species. DNA Res. 2014;21:127–40.
Schiavo G, Hoffmann OI, Ribani A, Utzeri VJ, Ghionda MC, Bertolini F, Geraci C, Bovo S, Fontanesi L. A genomic landscape of mitochondrial DNA insertions in the pig nuclear genome provides evolutionary signatures of interspecies admixture. DNA Res. 2017;24:487–98.
Huang CY, Grünheit N, Ahmadinejad N, Timmis JN, Martin W. Mutational decay and age of chloroplast and mitochondrial genomes transferred recently to angiosperm nuclear chromosomes. Plant Physiol. 2005;138:1723–33.
Yuan Q, Hill J, Hsiao J, Moffat K, Ouyang S, Cheng Z, Jiang J, Buell C. Genome sequencing of a 239-kb region of rice chromosome 10L reveals a high frequency of gene duplication and a large chloroplast DNA insertion. Mol Gen Genomics. 2002;267:713–20.
Arabidopsis Genome Initiative. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature. 2000;408:796–815.
Neumann P, Navrátilová A, Koblízkováž A, Kejnovský E, Hřibová E, Hobza R, Widmer A, Doležel J, Macas J. Plant centromeric retrotransposons: a structural and cytogenetic perspective. Mob DNA. 2011;2:4.
Liao Y, Zhang X, Li B, Liu T, Chen J, Bai Z, Wang M, Shi J, Walling JG, Wing RA, Jiang J, Chen M. Comparison of Oryza sativa and Oryza brachyantha genomes reveals selection-driven gene escape from the centromeric regions. Plant Cell. 2018;30:1729–44.
Ming R, Bendahmane A, Renner SS. Sex chromosomes in land plants. Annu Rev Plant Biol. 2011;62:485–514.
Sousa A, Bellot S, Fuchs J, Houben A, Renner SS. Analysis of transposable elements and organellar DNA in male and female genomes of a species with a huge Y chromosome reveals distinct Y centromeres. Plant J. 2016;88:387–96.
Li SF, Zhang GJ, Yuan JH, Deng CL, Gao WJ. Repetitive sequences and epigenetic modification: inseparable partners play important roles in the evolution of plant sex chromosomes. Planta. 2016;243:1083–95.
Hisanaga T, Okahashi K, Yamaoka S, Kajiwara T, Nishihama R, Shimamura M, Yamato KT, Bowman JL, Kohchi T, Nakajima K. A cis-acting bidirectional transcription switch controls sexual dimorphism in the liverwort. EMBO J. 2019;38:e100240.
Steele PR, Hertweck KL, Mayfield D, McKain MR, Leebens-Mack J, Pires JC. Quality and quantity of data recovered from massively parallel sequence: examples in Asparagus and Poaceae. Am J Bot. 2012;99:330–48.
Rychlik W. OLIGO 7 primer analysis software. Methods Mol Biol. 2007;402:35–60.
Li SF, Guo YJ, Li JR, Zhang DX, Wang BX, Li N, Deng CL, Gao WJ. The landscape of transposable elements and satellite DNAs in the genome of a dioecious plant spinach (Spinacia oleracea L.). Mobile DNA. Chromosom Res. 2019;10:3.
Acknowledgements
The authors thank Dr. Pires and Dr. Hertweck (National Evolutionary Synthesis Center, Durham, North Carolina, USA) for providing the chloroplast DNA sequence of A. officinalis.
Funding
This work was financially supported by grants from the National Natural Science foundation of China (31470334 and 31770346), Program for Innovative Research Team (in Science and Technology) in University of Henan Province (17IRTSTHN017), and the Foundation for Young Teachers in Colleges and Universities of Henan Province (2016GGJS-051). Each fund supports the experimental material cost, paper revision and paper publication fee, etc. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
SL analyzed the data and wrote the manuscript, HZ and JY participated in the analyses of the data, JL, JW, RD and KJ conducted the experiment, CD and NL provided helpful suggestion in data analysis and substantively revised the manuscript, WG designed the project and revised the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The plant materials used in this study are widely used all over the world and no permits are required for the collection of plant samples. The plant materials are maintained in accordance with the institutional guidelines of the College of Life Sciences, Henan Normal University, China. This article did not contain any studies with human participants or animals and did not involve any endangered or protected species.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional files
Additional file 1:
Table S1. The detailed information of NUPTs in the nuclear genome of A. officinalis. (DOCX 44 kb)
Additional file 2:
Table S2. The primers used for amplification of chloroplast DNA fragments of A. officinalis. (XLSX 278 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Li, SF., Li, JR., Wang, J. et al. Cytogenetic and genomic organization analyses of chloroplast DNA invasions in the nuclear genome of Asparagus officinalis L. provides signatures of evolutionary complexity and informativity in sex chromosome evolution. BMC Plant Biol 19, 361 (2019). https://doi.org/10.1186/s12870-019-1975-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-019-1975-8