
Abstract
Background
Black shank disease caused by Phytophthora nicotianae is a serious threat to flue-cured tobacco production. Whole-plant resistance is characterized by the expression of a number of pathogenesis-related proteins, genes, and the activity of different defense-related enzymes. In this study, we investigated the activity of defense-related enzymes and expression of differentially expressed proteins through the iTRAQ technique across two flue-cured tobacco cultivars, i.e., K326 and Hongda, in response to the black shank pathogen.
Results
Results showed that the highest disease incidence was recorded in flue-cured tobacco cultivar Hongda compared with K326, which shows that Hongda is more susceptible to P. nicotianae than K326. A total of 4274 differentially expressed proteins were detected at 0 h and after 24 h, 72 h of post-inoculation with P. nicotianae. We found that 17 proteins induced after inoculation with P. nicotianae, including pathogenesis (5), photosynthesis (3), oxidative phosphorylation (6), tricarboxylic acid cycle (1), heat shock (1), and 14–3-3 (1) and were involved in the resistance of flue-cured tobacco against black shank disease. The expression of 5 pathogenesis-related proteins and the activities of defense-related enzymes (PPO, POD, SOD, and MDA) were significantly higher in the leaves of K326 than Hongda after inoculation with P. nicotianae.
Conclusion
These results provide new molecular insights into flue-cured tobacco responses to P. nicotianae. It is concluded that differences in protein expressions and defense-related enzymes play an important role in developing resistance in flue-cured tobacco cultivars against black shank disease.
Similar content being viewed by others


Background
Black shank caused by an oomycete Phytophthora nicotianae is a destructive soilborne disease of tobacco (Nicotiana tabacum L) worldwide, including China [1]. Black shank disease was first reported in Java (Indonesia) in 1896 by Van Breda de Haan, and since then, it has been widespread around the globe. Tobacco crops are badly affected by this pathogen, which results in substantial yield losses [2]. P. nicotianae has a wide host range, infects 255 plant genera from 90 plant families, and causes crown rot, root rot, fruit rot, damping-off, leaf, and stem infection [3]. It can survive in the soil for a long period (4–6 years) even in the absence of a host plant and reproduces both sexually and asexually, having a reproduction cycle of 72 h [4, 5].
Tobacco crops can get affected by P. nicotianae at any growth stage from seedling to harvest, and it is difficult to control the disease [1]. In China, it is commonly known as Hei Jing Bing "Black Shank Disease" by the local farmers based on the typical disease symptoms [6]. The infected plants generally show symptoms of root and crown rot, yellowing, wilting, and necrosis of leaves. On cut stems, the signs of P. nicotianae are observed as white mycelium on the necrotic plate-like disks [7]. The average disease incidence of tobacco black shank is recorded between 10 to 20%, but it can reach up to 100% [8]. In severe outbreaks and prolonged drought periods, it occurs in epidemic form, resulting in plant death and destruction of the whole crop [9].
Soil environmental conditions such as rhizospheric microbial diversity, availability of nutrients and organic matter, soil texture, and soil pH are the main factors responsible for soilborne diseases [10]. Over the past few decades, important integrated disease management methods such as resistant cultivars [11], chemical pesticides (synthetic fungicides) [12], crop rotation, and soil replacement [13] have been adopted to control this disease. However, these methods have many disadvantages, such as the long-term application of synthetic fungicides may lead to resistance in the pathogen, affect soil microbial diversity, and are environmentally unfriendly [1, 14]. Therefore, microorganism-mediated biocontrol is considered a practical approach to mitigate soilborne diseases [15]. These microorganisms reduced the incidence of soilborne disease through the mechanism of competitive inhibition, production of antimicrobial compounds, induction of host resistance, and reshaping the soil microbiome's community structure [15, 16].
Cultivation and selection of disease-resistance cultivars play an important role to control tobacco black shank disease [11]. Many studies have proven that the incidence of black shank disease was higher in susceptible cultivars than resistance cultivars [17]. Upon pathogen attack, the plant defense system activates, which helps the plant to limit the invasion. During the defense activation the expression of certain disease-resistance changes significantly [18]. Disease-resistance proteins identify the proteins secreted by pathogens and trigger the immune responses to control pathogen infection [19]. Defense-related enzymes and compounds such as catalase (CAT), peroxidase (POD), superoxide dismutase (SOD), polyphenol oxidase (PPO), and malondialdehyde (MDA) are involved in the defense function and metabolic balance of plants [20, 21]. Proteomics is a practical approach to decipher an organism's physiological and pathological mechanisms [19].
For instance, the activities of defense-related enzymes POD, SOD, PPO, and PAL significantly increased in the stem and leaves of the patchouli plant after inoculation of Ralstonia solanacearum [22]. Many studies have proven that the content of MDA in tobacco leaves and enzyme activity such as SOD, POD, PPO, and CAT in tobacco plants increased after inoculation of black shank pathogen [23, 24]. Singh et al. (2021) used the iTRAQ technique to investigate the physiological role of pathogenesis-related proteins in two sugarcane varieties GT29 (smut-resistant) and Yacheng71-374 (smut-susceptible) with contrasting resistance to sugarcane smut. Results showed that after inoculation of Sporisorium scitamineum, the number of differentially expressed proteins was higher in GT29 than Yacheng71-374 [25]. Under normal conditions, plant defense-related enzymes and protein expression remained stable but when plants were exposed to biotic and abiotic stresses the protein expression changed significantly. However, it was found that defense-related enzymes and protein expression levels in different cultivars (resistant and susceptible) also differ. Therefore, enzymatic activity and protein expression in a healthy plant can be used as a reference index for screening of resistant cultivars.
Tobacco (Nicotiana tabacum) has been widely used as a model plant for in-depth molecular and biological studies. Scarce information is present on the molecular-based proteomic analysis and activities of defense-related enzymes in different flue-cured tobacco cultivars with contrasting resistance to blank shank disease. Therefore, the present study aims to fill this knowledge gap with more advanced techniques to explore the important mechanisms involved in two flue-cured tobacco cultivars with contrasting resistance against P. nicotianae. Differential responses of defense-related proteins and enzymatic activity help us to decipher the molecular mechanisms involved in flue-cured tobacco resistance against black shank pathogen.
Methods
Experimental site and design description
A greenhouse experiment was conducted during the growing season from March-August in 2017 at Technology Extension Station of Tobacco Company in Xundian (25°46' N, 102°55' E) County, Kunming City, Yunnan Province, China. Seedlings (45 days old) of two flue-cured tobacco cultivars, K326 and Hongda were transplanted in pots (40 × 35 cm) containing 13 kg of soil. To overcome nutrient deficiency fertilizers were applied twice as base fertilizer (N-P2O5-K2O = 8- 16- 22) 45 g/pot and top fertilizer (20 days after transplantation) (N- P2O5- K2O = 15–0-33) 25 g/pot [26]. Each pot was irrigated with 1000 ml/plant of water thrice a week to maintain moisture level. The greenhouse conditions were maintained as a day/night temperature (30/20℃) with a 14 h light/10 h dark photoperiod [27]. The experiment was conducted under a completely random design, and each treatment was repeated thrice with 45 plants/pots of each cultivar (K326 and Hongda) in each treatment. All integrated filed management practices and sample collection protocols were performed according to the National Standards of Tobacco Industry in China as described by Tang et al. [28].
Pathogen strain, culture medium, and growth conditions
Phytophthora nicotianae used throughout this experiment was provided by Professor Guanghai Ji (State Key Laboratory for Conservation and Utilization of Bio-Resources in Yunnan), Yunnan Agricultural University, Kunming, China. Pure culture of P. nicotianae was prepared on oatmeal agar (OA) medium (Oatmeal 30 g/L; Agar 20 g/L; pH 7.0) and incubated at 27℃ for 7–10 days. Zoospores of P. nicotianae were prepared as described by Zhang et al. [29]; briefly, P. nicotianae was cultured in OA medium at 27℃ for three weeks. Then 10 ml/plate of 0.1% KNO3 solution was added, and plates were incubated at 27℃ for three days. After a sudden drop down to 4℃ for 30 min, the zoospore suspension was collected by filtering through gauze. The resulting zoospore suspension was centrifuged at 1200 rpm at 4℃ for 10 min. The supernatant was diluted with sterilized distilled water to a concentration of 1 × 106 zoospores/ml using a hemacytometer.
Inoculation of pathogen and sample collection
Zoospore suspension of P. nicotianae (1 × 106 zoospores/ml) was prepared as described above, and inoculation was done 30 days after seedling transplantation at the 5–6 leaf stage. Before inoculation, each pot was irrigated with 1000 ml of water, and about 30 ml/plant of zoospores suspension was sprayed on the leaves [1]. Leaf samples were collected at 0 h (before inoculation) and after 24 h, 72 h of post-inoculation with P. nicotianae. The collected samples were stored in an ultra-low volume refrigerator at -80℃ for further study (analysis of defense-related anti-oxidative and proteomics study). Here, K-0, K-24, K-72 and HD-0, HD-24, HD-72 represent the samples collected from K326 and Hongda, respectively, at 0 h (before inoculation) and after 24 h, 72 h of post-inoculation.
Analysis of defense-related enzymes
Polyphenol oxidase activity
Polyphenol oxidase activity was determined with the catechol colorimetric method by measuring the absorbance rate of quinone formation at OD525 nm using a spectrophotometer (PharmaSpec 1700) as described by Sikora et al. [30]. An increase in absorbance rate of 0.01 min−1 for quinone formation was taken as one unit of enzyme activity. The sample contained 0.3 ml of crude enzyme solution, two ml of phosphate buffer (0.05 mmol/ml and pH 7.0), and one ml of catechol (0.1 mol/ml).
Peroxidase activity
Oxidation of guaiacol is the rate-determining step for peroxidase activity. Peroxidase activity was determined by the guaiacol method using the methodology of Zhang and Shao [31]. Briefly, 3.9 ml of the reaction solution was added (28 µl of 30% H2O2, 50 ml of 0.1 mol/L phosphate buffer (pH 6.0), and 19 µl of guaiacol) to a 10-ml test tube and placed in a water bath at 25℃ for 5 min. Then, 0.1 ml of crude enzyme solution was added to the test tube, mixed thoroughly, and an increase in the absorbance rate of 0.01 min−1 at OD470 nm using a spectrophotometer (PharmaSpec 1700) was taken as one unit for enzyme activity.
Superoxide dismutase activity
Superoxide dismutase activity was evaluated by measuring its ability to inhibit photochemical reduction of nitro blue tetrazolium using the methodology of Kong et al. [32]. The reaction mixture (2.725 ml) contained (26 mM methionine, 1 μM EDTA, 20 μM riboflavin, 50 mM phosphate buffer (pH 7.8), and 750 μM nitro blue tetrazolium). After adding crude enzyme solution (25 μl) and distilled water (250 μl) into the reaction mixture, the reaction was allowed to run at 25–35 °C for 20 min under a 4000-lx fluorescent lamp. The absorbance of the reaction mixture was measured at OD560 nm using a spectrophotometer (PharmaSpec 1700). The 50% nitro blue tetrazolium photochemical reduction inhibition was taken as an enzyme activity unit.
Malondialdehyde activity
Malondialdehyde contents were determined using the thiobarbituric acid colorimetric method as described by Gao and Zhang [33]. Briefly, 0.2 g of samples were ground on ice in 10% trichloroacetic acid to make a homogenized mixture. The mixture was then added to a 3 ml solution (0.5% thiobarbituric acid + 10% trichloroacetic acid) and incubated at 90℃ for 15 min in a water bath. After this, the suspension was cooled down at room temperature and centrifuged at 10,000 rpm for 10 min at 4℃. The supernatant was collected and light absorbance was measured at OD450 nm, OD532 nm, and OD650 nm using a spectrophotometer (PharmaSpec 1700). Malondialdehyde contents were measured using following formula: MDA contents (nmol/g) = [6.45 × (A532-A650)—0.56 × A450] × VT/FW/VS. Here: VT is the volume of extraction solution; VS is the volume of test solution; FW is the weight of the sample.
Proteins extraction and iTRAQ assay
Proteins extraction and quantification
Proteins extraction, quantification, and digestion were done as previously described by Xie et al. [22]. Total protein was extracted from the frozen dried leaf samples, ground in liquid nitrogen, and extracted with lysis buffer (2 M thiourea, 7 M urea, 40 mM Tris–HCl, and 4% 3-[(3-Cholamidopropyl) dimethylammonio]-1-propanesulfonate (CHAPS), pH 8.5) supplemented with two mM ethylenediaminetetraacetic acid (EDTA) and one mM phenylmethane sulfonyl fluoride (PMSF). After 5 min of vigorous vortexing 10 mM dithiothreitol (DTT) was added to the suspension mixture. The suspension was sonicated at 200 W for 15 min and centrifuged at 15000 rpm for 20 min. The supernatant was collected and mixed with a 5 × volume of chilled acetone containing 10% (v/v) trichloroacetic acid (1:4, v/v) and incubated overnight at − 20℃. Proteins in the supernatant were collected and stored at − 80℃ for further analysis.
iTRAQ labeling and fractionation
Samples were labeled with iTRAQ 8-plex Multiplex Reagent Kit (AB Sciex U.K.) by following the manufacturer's instructions. All labeled samples were mixed in equal volume and fractionated using a high-performance liquid chromatography (HPLC) system (Thermo Dionex Ultimate 3000 BioRS).
LC–MS/MS proteomic analysis
LC–MS/MS analysis was performed in buffer A (0.1% formic acid and 2% acetonitrile), and the final concentration of peptides was adjusted to 0.5 μg·μL−1. From each sample, ten μL supernatant was loaded into a two cm C18 trap column in an LC-20 CE nano HPLC (Shimadzu, Kyoto, Japan) via an auto-sampler. Samples were loaded at a rate of 8 μl/min for 4 min. The following 41 min gradient was then run at 300 nL/min: 5% to 35% B (0.1% formic acid 98% and acetonitrile), 5 min linear gradient to 80%, 80% B for 5 min, and a one min decrease to 5%. The peptides were subjected to nano-electrospray ionization followed by MS/MS via a Q-Exactive mass spectrometer (Thermo Fisher Scientific, San Jose, CA, USA).
Bioinformatics analysis
The whole experiment was performed in triplicate with 3–5 analytical replications in each treatment. Proteins identification and quantitative analysis (false-discovery rate (FDR) ≤ 0.05) were performed using the Mascot 2.3.02 search engine against the Nicotiana tabacum database (http://www.ncbi.nlm.nih.gov/protein?term) and Proteome Discoverer™ Software (Thermo), respectively. The relevant parameters used for screening of differentially expressed proteins are shown in Table 1. Proteins with fold changes ≥ 1.2 or ≤ 0.833 and Q-values < 0.05 in at least two replicate/treatment were considered as differentially abundant proteins [34]. To determine the biological process, molecular process, and cellular component, blast2go software (V4.5 pipeline) was used for GO annotation and compared with the gene ontology (GO) database (http://geneontology.org/) for the differences in protein components. Kyoto Encyclopedia of Genes and Genomes (KEGG) database (www.kegg.jp/kegg/kegg1.html) was used to classify and group the identified proteins [35]. Data were analyzed statistically using a t-test to determine differences at a 5% significance level with Microsoft Excel 2013 and SPSS version 22.0 (SPSS, Chicago, IL, USA) [36]. All figures were processed and analyzed using Adobe Illustrator CS5 (Adobe Systems Inc., San Francisco, CA, USA).
Disease incidence
Disease incidence was recorded once a week after post-inoculation till the end of the experiment. Disease incidence was graded according to GB/T 23222–2008 (National Standardization Management Committee of China, 2009) using a disease rating scale as follows: 0; healthy plants, 1; one-third of the leaves were wilted and diseased spot on stem does not exceed one-third of the stem, 3; half of the leaves were wilted and diseased spot exceeded half of the stem, 5; two-third of leaves were wilted and necrosis of older leaves, 7; all leaves were wilted and necrosis of more than half of leaves, 9; death of whole plant [37]. Disease incidence (DI) was calculated using the following formula: DI (%) = (Number of infected plants/Total investigated plants) × 100.
Results
Analysis of disease incidence
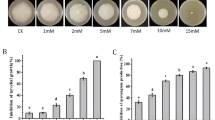
It was found that different flue-cured tobacco cultivars show different level of resistance against black shank disease. Severe environmental conditions like continuous high temperature and high humidity were provided through the experiment after the inoculation of P. nicotianae. In the early stage of infection, no significant difference was observed in disease incidence between K326 and Hongda. But as time passed, a significant difference was observed between K326 and Hongda in disease occurrence. In the case of Hongda, the disease incidence was reached up to 100% after the 8th week of post-inoculation, whereas K326 showed more resistance against black shank disease than Hongda (Fig. 1).

Disease incidence (%) in two flue-cured tobacco cultivars (K326 and Hongda). Significant difference among flue-cured tobacco cultivars is shown by different small letters on the bars, according to Duncan's multiple range test at p < 0.05
Effect of P. nicotianae on the activity of defense-related enzymes in different flue-cured tobacco cultivars
The activity of different defense-related enzymes (PPO, POD, and SOD) and MDA contents were examined in leaves of two flue-cured tobacco cultivars K326 and Hongda at 0 h, and after 24 h and 72 h of post-inoculation with P. nicotianae. Results revealed that the activity of PPO, POD, and SOD enzymes and MDA contents were significantly higher in K326 than Hongda (Fig. 2). However, the activity of PPO, POD, and SOD enzymes and MDA contents in both flue-cured cultivars K326 and Hongda initially increased and then decreased after 24 h and 72 h of post-inoculation with P. nicotianae, respectively.

Activities of defense-related enzymes across different flue-cured tobacco cultivars in response to black shank pathogen. PPO activity (A), POD activity (B), SOD activity (C), and MDA contents (D). Error bars represent the standard error of means (± SEM, n = 3). Different small letters on error bars indicate a significant difference according to Duncan's multiple range test at p < 0.05
Proteins identification and profiling
In this study, a total of 405,918 MS/MS spectra were confirmed using the iTRAQ LC–MS/MS technique; among them, 51,812 spectra matched with known protein spectra in the database. In addition, 14,146 peptides, 9,918 unique peptides, and 4,274 differentially expressed proteins were identified, of which 69.4% of proteins contained more than two peptides, as shown in Fig. 3. Proteins were screened based on a fold-change value ≥ 1.2 or ≤ 0.833 and a p-value < 0.05. Differential abundant protein screening was performed in three replicates and differential abundant proteins that were appeared at least twice in three replicates were selected for subsequent analysis.

Proteins identification and profiling for differently expressed proteins. Data are shown as the standard error of means (± SEM, n = 3) at p < 0.05
Analysis of differentially expressed proteins
The number of differentially expressed proteins obtained by statistical analysis is shown in Fig. 4. In this study, a total of 723 differentially expressed proteins (325 up-regulated and 398 down-regulated) were found in a group vise comparison between K-0 vs HD-0, indicating that selected cultivars K326 and Hongda are different from each other based on their genetics. Similarly, a total of 35 (15 up-regulated and 10 down-regulated), 271 (98 up-regulated and 173 down-regulated), 318 (183 up-regulated and 135 down-regulated), and 137 (59 up-regulated and 78 down-regulated) differentially expressed proteins were identified between K-24 vs K-0, HD-24 vs HD-0, K-72 vs K-0, and HD-72 vs HD-0, respectively.

Identification of differentially expressed protein in group-wise comparison across different flue-cured tobacco cultivars after specific hours of post-inoculation with Phytophthora nicotianae. Data are shown as the standard error of means (± SEM, n = 3) at p < 0.05. Here: K; K326, HD; Hongda, 0; before inoculation at 0 h, and 24, 72; after 24 h and 72 h of post-inoculation
Bioinformatics analysis for differentially expressed proteins
GO classification analysis was performed for differentially expressed proteins between 5 different groups (K-0 vs HD-0, K-24 vs K-0, HD-24 vs HD-0, K-72 vs K-0, and HD-72 vs HD-0) as shown in Fig. 5 (A; biological processes, B; molecular processes, C; cellular components). Results showed that differential proteins were mainly involved in cellular processes, metabolic processes, single-cell processes, biological regulation, and response to a stimulus. Proteins related to molecular functions are mostly binding proteins and catalytic activities (enzymes). As cell components, proteins are mainly involved in cells, cell parts, organelle, and membrane.

GO classification analysis for differentially expressed proteins in group-wise comparison across different flue-cured tobacco cultivars after specific hours of post-inoculation with black shank pathogen. All identified proteins (4274 in total) were categorized according to Biological processes (A), Molecular processes (B), and Cellular components (C). Here: K; K326, HD; Hongda, 0; before inoculation at 0 h, and 24, 72; after 24 h and 72 h of post-inoculation
Screening of proteins related to induction of black shank pathogen
To further investigate the difference in response of K326 and Hongda to black shank infection, differentially expressed proteins of two flue-cured tobacco cultivars were analyzed after 24 h and 72 h of post-inoculation with P. nicotianae (Fig. 6). Results revealed that after 24 h of post-inoculation, K326 showed ten unique proteins, while Hongda showed 96 unique proteins. However, 11 proteins in K326 and 171 in Hongda were induced after 24 h of post-inoculation. Four different proteins were found in two flue-cured cultivars after 24 h of post-inoculation, among which one showed an opposite expression trend and three showed the same trend (two up-regulated and one down-regulated) (Fig. 6A).

Venn diagram for differentially expressed proteins in flue-cured tobacco cultivars (K326 and Hongda) after inoculation of black shank pathogen. Here: 24 h of post-inoculation (A) and 72 h of post-inoculation (B)
Unique proteins in K326 increased to 184, while specific proteins in Hongda decreased to 37 after 72 h of post-inoculation. However, 87 proteins in K326 and 53 in Hongda were induced after 72 h of post-inoculation. There were 47 differentially expressed proteins in two flue-cured cultivars, among which five showed the opposite expressions, and 42 showed the same expression trends (27 up-regulated and 15 down-regulated) (Fig. 6B). It was found that 17 proteins were involved in the resistance of flue-cured tobacco to black shank after 0 h (before inoculation) and 24 h, 72 h of post-inoculation (Table 2). By identifying and screening the above 17 differential proteins, five pathogenies-related proteins, three photosynthesis-related proteins, six proteins involved in oxidative phosphorylation, one protein involved in the tricarboxylic acid cycle, one heat shock protein, and one 14–3-3 protein were found. These specific proteins may be an important factor in determining flue-cured tobacco resistance to the black shank disease.
Discussion
Activities of defense-related enzymes across different flue-cured tobacco cultivars in response to black shank pathogen
Induced systemic resistance and systemic acquired resistance of plants play an important role in disease development after a pathogen attack. Studies have proven that the activities of defense-related enzymes in plants remain stable during normal growth and change when exposed to biotic and abiotic stresses [38]. An increase in the activity of PPO enzyme can enhance the plant's resistance to biotic and abiotic stresses [39]. Similarly, POD and SOD are known to be as important scavenging enzymes of reactive oxygen species (ROS) in plants [40]. Peroxidase can catalyze hydrogen peroxide (which is toxic to plants) into non-toxic water and oxygen. Simultaneously, SOD removes superoxide anion free radicals produced by plants under stress and protects cells from damage [41, 42]. We observed that PPO, POD, and SOD enzymes expression were different among different flue-cured tobacco cultivars. After being infected with black shank pathogen, the activities of PPO, POD, and SOD enzymes increased in both cultivars K326 and Hongda, but enzyme activity was found higher in K326.
Malondialdehyde is a final product of membrane lipid peroxidation in plants under stress and is an important biochemical index to reflect the degree of membrane damage. When plants are under stress, higher malondialdehyde contents in leaves indicate a higher degree of membrane and cell damage [42, 43]. In our study, we found that the contents of MDA in the leaves of K326 were increased initially and then decreased significantly after the inoculation of P. nicotianae. Although the varying trend in the MDA contents in Hongda remained consistent, and no significant difference was found compared to control (0 h before inoculation). Our study demonstrates that the activities of PPO, POD, and SOD enzymes and MDA contents in the leaves of K326 increased significantly compared with Hongda after the inoculation of P. nicotianae. This may be related to systemic acquired resistance of K326 to black shank pathogen, while Hongda is more susceptible to black shank disease than K326.
Differentially expressed proteins induced by flue-cured tobacco cultivars in response to P. nicotianae
In this study, a total of 4274 differentially expressed proteins were found in the leaves of K326 and Hongda via iTRAQ technique after the inoculation of P. nicotianae. Among them, 723, 25, 271, 318, and 137 differentially expressed proteins were identified in a group-wise comparison between K-0 vs HD-0, K-24 vs K-0, HD-24 vs HD-0, K-72 vs K-0, and HD-72 vs HD-0, respectively. Further analysis of differentially expressed proteins between K326 and Hongda revealed that protein composition and contents in Hongda changed earlier than K326, which is similar to the findings of Parker et al. [44]. The resistance of K326 to black shank disease mainly manifests in a degree of damage to the photosynthetic system, cell oxidation balance, lignin synthesis capacity, and the induction strength of potential resistant genes. Preliminary, we found that twelve proteins were induced by the black shank pathogen. The expression of one D1 protein and one PPO protein decreased in K326 after pathogen inoculation, and the rest of the proteins showed an upward trend. The expression of two proteins involved in photosynthesis and one protein involved in oxidative phosphorylation was down-regulated in Hongda after the inoculation of P. nicotianae. So, we conclude that changes in the expression of differential proteins may be closely related to the genetic resistance of flue-cured tobacco cultivars to the black shank pathogen.
Proteins associated with photosynthesis
It is well documented that photosynthetic system is very sensitive to external biotic and abiotic stresses. The changes in the photosynthetic system are the most fundamental characteristic of plant response to external stresses [45]. Studies have proven that whenever plants are subjected to external biotic or abiotic stresses, the expression of most of the genes related to photosynthesis pathway change significantly [45]. Among differentially expressed proteins, it was found that three proteins were involved in the photosynthesis of tobacco plants. As the core component of photosynthetic system PSII, rapid turnover of Photosystem II, protein D1 is the main prerequisite for PSII to function. However, light-driven electron transfer is necessary for the turnover of D1 protein function to repair PSII [46]. Our study demonstrates that D1 protein was down-regulated after the inoculation with P. nicotianae. The proteins related to Photosystem I reaction center subunit XI as a core component of chloroplasts were also down-regulated in Hongda after inoculation of black shank pathogen. Another photosynthesis-related protein, the ATP synthase-α subunit, was up-regulated in K326 after the inoculation of black shank pathogen. These results revealed that the black shank pathogen has a specific role in promoting photosynthesis in resistant cultivars.
Tricarboxylic acid (TCA) related proteins
The host's defense response to pathogens is an active and energy-consuming process. In the main catabolic pathway of organisms, TCA is related to the mutual conversion of three major substances and acts as an important metabolic pathway that provides a large amount of free energy to organisms [47]. Through analysis of differentially expressed proteins, we found that one protein is involved in the TCA cycle, and the expression of acetyltransaminase-pyruvate dehydrogenase complex was up-regulated in K326 after inoculation of P. nicotianae. Results showed that the TCA pathway is activated in K326 after inoculation of black shank pathogen, thereby producing enough intermediate substances and energy to overcome pathogen infection [48]. This change may be related to the higher disease resistance of K326 towards P. nicotianae than Hongda.
Oxidative phosphorylation and lignin synthesis related proteins
It has been found that ROS outbreak is one of the early disease-resistant responses of the host against pathogen and ROS production is a key signal in plant defense response [49]. Reactive oxygen species play an important role in plant defense response, but excessive accumulation of ROS can cause damage to normal cells. In order to reduce the contents of ROS in cells, plant cells have evolved a complex and refined elimination mechanism such as; antioxidants ascorbic acid, glutathione, and antioxidant enzymes [50]. Studies have proven that SOD, POD, and other enzyme activities in tobacco plants increase after black shank infection [23]. In this study, POD, SOD, and PPO contents were significantly increased in K326 after inoculation of P. nicotianae. It shows that POD, PPO, and SOD play a vital role in maintaining the balance of reactive oxygen species in cells and cell membranes' stability. Lignin is a complex phenolic resin polymer and is important for maintaining plant cell structure and resistance against pathogen infection [51]. Studies have proven that POD also plays a vital role in the synthesis of plant lignin [52, 53]. Two differentially expressed proteins screened were identified as POD, and their expression was found up-regulated in both tobacco cultivars (K326 and Hongda) after the inoculation of P. nicotianae. The contents of POD were found significantly higher in K326 than Hongda after inoculation of P. nicotianae. Similarly, the disease incidence was recorded minimum in K326 compared with Hongda, indicating that K326 activates its immune system against pathogen invasion.
Pathogenesis-related proteins
Pathogenesis-related proteins (PRPs) are a class of proteins produced by plants in a pathological or pathologically related environment. Pathogen infection, abiotic stress, allergic reactions, and system acquired resistance can stimulate the accumulation of PRPs [54]. Therefore, the production and activity of PRPs play an important role in plant's resistance to biotic and abiotic stress [54]. In this study, we found that the expression of PR-4 family protein was down-regulated in K326 and the expression of STH-2 family protein was up-regulated compared with Hongda. After 24 h of post-inoculation, expression levels of three PR-4 family proteins in K326 and Hongda were down-regulated. However, after 72 h of post-inoculation compared with 0 h of post-inoculation, the expression of three PR-4 family proteins and two STH-2 family proteins in the K326 were up-regulated. The expression of three PR-4 family proteins and two STH-2 family proteins in the Hongda was down-regulated and up-regulated, respectively. In addition, the expression of other resistance-related proteins also changed significantly in K326 and Hongda after inoculation of P. nicotianae.
14–3-3 protein is involved in the regulation of primary metabolism, ion transport, intracellular transport, enzyme activation, and gene expression [55]. Many studies have proven that the 14–3-3 protein interacts with chaperonin Hsp70 to form a guide complex that mediates thylakoid proteins import [56, 57]. Yan et al. (2004) transformed the Arabidopsis thaliana GF14λ gene encoding 14–3-3 protein into cotton. They found that the transgenic cotton wilting rate was low under drought conditions and photosynthesis efficiency was higher than wild-type cotton [58]. In K326, we found that the expression of 14–3-3a-1 protein was up-regulated after inoculation of P. nicotianae, which improves K326 resistance to P. nicotianae. Our results are in accordance with the study of Lee et al., who reported that 14–3-3 protein in the nucleus of sweet pepper is involved in transcriptional regulation against TMV infection [59].
Conclusion
In the light of obtained results, we conclude that the iTRAQ-based study expands our knowledge of defense-related enzymes and pathogenesis-related proteins in two flue-cured cultivars K326 and Hongda, after inoculation of P. nicotianae. A total of 4274 differentially abundant proteins were identified using the iTRAQ-based quantitative proteomic approach. Functional analysis of these differentially abundant proteins revealed that proteins were involved in photosynthesis, TCA cycle, oxidative phosphorylation, lignin synthesis, and PRPs. Enzymes such as PPO, POD, SOD, and MDA play an important role in the defense mechanism of flue-cured tobacco plants during black shank pathogen infection. Energy- and ROS-related biological processes and pathways were actively regulated in leave of K326 when exposed to P. nicotianae. These results advance our knowledge to understand the molecular changes involved in different flue-cured tobacco cultivars with contrasting resistance to black shank disease. Although proteomic analysis is a long-term goal, further studies should be done on protein signatures and transcriptomic analysis to screen the resistant cultivars against back shank pathogen.
Availability of data and material
The datasets generated and analyzed during the current study are available in the iProX (https://www.iprox.cn/page/HMV006.html) having Accession Number IPX0003938000.
References
Liu T, Xiao Y, Yin J, Yi T, Zhou Z, Hsiang T, Tang Q, Chen W. Effects of cultured root and soil microbial communities on the disease of Nicotiana tabacum caused by Phytophthora nicotianae. Front Microbiol. 2020;11:929. https://doi.org/10.3389/fmicb.2020.00929.
Gallup CA, McCorkle KL, Ivors KL, Shew D. Characterization of the black shank pathogen, Phytophthora nicotianae, across North Carolina tobacco production areas. Plant Dis. 2018;102(6):1108–14. https://doi.org/10.1094/PDIS-02-17-0295-RE.
Panabieres F, Ali GS, Allagui MB, Dalio RJ, Gudmestad NC, Kuhn M-L, Guha Roy S, Schena L, Zampounis A. Phytophthora nicotianae diseases worldwide: new knowledge of a long-recognised pathogen. Phytopathologia Mediterranea. 2016;55:20–40.
Biasi A, Martin FN, Cacciola SO, di San Lio GM, Grünwald NJ, Schena L. Genetic analysis of Phytophthora nicotianae populations from different hosts using microsatellite markers. Phytopathology. 2016;106(9):1006–14. https://doi.org/10.1094/PHYTO-11-15-0299-R.
Guo D, Yuan C, Luo Y, Chen Y, Lu M, Chen G, Ren G, Cui C, Zhang J, An D. Biocontrol of tobacco black shank disease (Phytophthora nicotianae) by Bacillus velezensis Ba168. Pestic Biochem Physiol. 2020;165:104523. https://doi.org/10.1016/j.pestbp.2020.01.004.
Xiang D, Yao Z, Liu Y, Gai X, Du Y, Zhang Z, Yan N, Wang A, Fu Q. Analysis on Solanesol Content and Genetic Diversity of Chinese Flue‐Cured Tobacco (Nicotiana tabacum L.). Crop Sci. 2017;57(2):847–55. https://doi.org/10.2135/cropsci2016.07.0568.
Kroumova AB, Artiouchine I, Wagner GJ. Use of several natural products from selected Nicotiana species to prevent black shank disease in tobacco. Beiträge zur Tabakforschung International/Contrib Tobacco Res. 2016;27(3):113–25. https://doi.org/10.1515/cttr-2016-0013.
Zhang Q, Feng R, Zheng Q, Li J, Liu Z, Zhao D, Meng Y, Tian Y, Li W, Ma X, Wang S. Population genetic analysis of Phytophthora parasitica from tobacco in Chongqing. Southwestern China Plant disease. 2019;103(10):2599–605. https://doi.org/10.1094/PDIS-05-18-0879-RE.
McCorkle KL, Drake-Stowe K, Lewis RS, Shew D. Characterization of Phytophthora nicotianae resistance conferred by the introgressed Nicotiana rustica region, Wz, in flue-cured tobacco. Plant Dis. 2018;102(2):309–17. https://doi.org/10.1094/PDIS-03-17-0339-RE.
Cai Q, Zhou G, Ahmed W, Cao Y, Zhao M, Li Z, Zhao Z. Study on the relationship between bacterial wilt and rhizospheric microbial diversity of flue-cured tobacco cultivars. Eur J Plant Pathol. 2021;160(2):265–76. https://doi.org/10.1007/s10658-021-02237-4.
Deb D, Shrestha A, Maiti IB, Dey N. Recombinant promoter (MUASCsV8CP) driven totiviral killer protein 4 (KP4) imparts resistance against fungal pathogens in transgenic tobacco. Front Plant Sci. 2018;9:278. https://doi.org/10.3389/fpls.2018.00278.
You C, Zhang C, Kong F, Feng C, Wang J. Comparison of the effects of biocontrol agent Bacillus subtilis and fungicide metalaxyl–mancozeb on bacterial communities in tobacco rhizospheric soil. Ecol Eng. 2016;91:119–25. https://doi.org/10.1016/j.ecoleng.2016.02.011.
Niu J, Chao J, Xiao Y, Chen W, Zhang C, Liu X, Rang Z, Yin H, Dai L. Insight into the effects of different cropping systems on soil bacterial community and tobacco bacterial wilt rate. J Basic Microbiol. 2017;57(1):3–11. https://doi.org/10.1002/jobm.201600222.
Stumbriene K, Gudiukaite R, Semaskiene R, Svegzda P, Jonaviciene A, Suproniene S. Screening of new bacterial isolates with antifungal activity and application of selected Bacillus sp. cultures for biocontrol of Fusarium graminearum under field conditions. Crop Prot. 2018;113:22–8. https://doi.org/10.1016/j.cropro.2018.07.006.
Ahmed W, Yang J, Tan Y, Munir S, Liu Q, Zhang J, Ji G, Zhao Z. Ralstonia solanacearum, a deadly pathogen: Revisiting the bacterial wilt biocontrol practices in tobacco and other Solanaceae. Rhizosphere. 2022;21:100479.
Zhang J, Wei L, Yang J, Ahmed W, Wang Y, Fu L, Ji G. Probiotic Consortia: Reshaping the Rhizospheric Microbiome and Its Role in Suppressing Root-Rot Disease of Panax notoginseng. Front Microbiol. 2020;11:701. https://doi.org/10.3389/fmicb.2020.00701.
Zhang Y, Guo X, Yan X, Ren M, Jiang C, Cheng Y, Wen L, Liu D, Zhang Y, Sun M. Identification of stably expressed QTL for resistance to black shank disease in tobacco (Nicotiana tabacum L.) line Beinhart 1000–1. Crop J. 2018;6(3):282–90. https://doi.org/10.1016/j.cj.2017.12.002.
Teixeira PJP, Colaianni NR, Fitzpatrick CR, Dangl JL. Beyond pathogens: microbiota interactions with the plant immune system. Curr Opin Microbiol. 2019;49:7–17. https://doi.org/10.1016/j.mib.2019.08.003.
Martin GB, Bogdanove AJ, Sessa G. Understanding the functions of plant disease resistance proteins. Annu Rev Plant Biol. 2003;54(1):23–61. https://doi.org/10.1146/annurev.arplant.54.031902.135035.
Habibi F, Ramezanian A, Rahemi M, Eshghi S, Guillén F, Serrano M, Valero D. Postharvest treatments with γ-aminobutyric acid, methyl jasmonate, or methyl salicylate enhance chilling tolerance of blood orange fruit at prolonged cold storage. J Sci Food Agric. 2019;99(14):6408–17. https://doi.org/10.1002/jsfa.9920.
Latef AAHA, Dawood MF, Hassanpour H, Rezayian M, Younes NA. Impact of the Static Magnetic Field on Growth, Pigments, Osmolytes, Nitric Oxide, Hydrogen Sulfide Phenylalanine Ammonia-Lyase Activity, Antioxidant Defense System, and Yield in Lettuce. Biology. 2020;9(7):172. https://doi.org/10.3390/biology9070172.
Xie H, Yang D-H, Yao H, Bai G, Zhang Y-H, Xiao BG. iTRAQ-based quantitative proteomic analysis reveals proteomic changes in leaves of cultivated tobacco (Nicotiana tabacum) in response to drought stress. Biochem Biophys Res Commun. 2016;469(3):768–75. https://doi.org/10.1016/j.bbrc.2015.11.133.
Wang G, Yang H, Zhao Z, Li F, Yi J. Differences in the response of tobacco variety defense enzyme activity to sterilization bacteria. Journal of Yunnan Agricultural University (Natural Sciences). 2012;27(03):321–6 CNKI:SUN:YNDX.0.2012-03-005.
Zhang W, Wei S, Wang Y, Luo D, Yao X, Zeng H, Zeng J, Liu C, He J, Li Q, Ye P. Study on the biochemical mechanism of the resistance of flue-cured tobacco varieties to Black Shank. Southwest Journal of Agriculture. 2012;25(06).
Singh P, Singh RK, Song Q-Q, Li H-B, Guo D-J, Malviya MK, Verma KK, Song XP, Lakshmanan P, Yang LT. Comparative analysis of protein and differential responses of defense-related gene and enzyme activity reveals the long-term molecular responses of sugarcane inoculated with Sporisorium scitamineum. Journal of Plant Interactions. 2021;16(1):12–29. https://doi.org/10.1080/17429145.2020.1867770.
Li C, Ahmed W, Li D, Yu L, Xu L, Xu T, Zhao Z. Biochar suppresses bacterial wilt disease of flue-cured tobacco by improving soil health and functional diversity of rhizosphere microorganisms. Appl Soil Ecol. 2022;171: 104314. https://doi.org/10.1016/j.apsoil.2021.104314.
Knödler M, Rühl C, Emonts J, Buyel JF. Seasonal weather changes affect the yield and quality of recombinant proteins produced in transgenic tobacco plants in a greenhouse setting. Front Plant Sci. 2019;10:1245. https://doi.org/10.3389/fpls.2019.01245.
Tang Z, Chen L, Chen Z, Fu Y, Sun X, Wang B, Xia T. Climatic factors determine the yield and quality of Honghe flue-cured tobacco. Sci Rep. 2020;10:1–12. https://doi.org/10.1038/s41598-020-76919-0.
Zhang C, Feng C, Zheng Y, Wang J, Wang F. Root Exudates Metabolic Profiling Suggests Distinct Defense Mechanisms Between Resistant and Susceptible Tobacco Cultivars Against Black Shank Disease. Front Plant Sci. 2020;11:1352. https://doi.org/10.3389/fpls.2020.559775.
Sikora M, Świeca M, Franczyk M, Jakubczyk A, Bochnak J, Złotek U. Biochemical properties of polyphenol oxidases from ready-to-eat lentil (Lens culinaris Medik.) sprouts and factors affecting their activities: A search for potent tools limiting enzymatic browning. Foods. 2019;8(5):154. https://doi.org/10.3390/foods8050154.
Zhang X, Shao X. Characterisation of polyphenol oxidase and peroxidase and the role in browning of loquat fruit. Czech Journal of Food Sciences. 2015;33(2):109–17. https://doi.org/10.17221/384/2014-CJFS.
Kong W, Zhao Y, Liu F, He Y, Tian T, Zhou W. Fast analysis of superoxide dismutase (SOD) activity in barley leaves using visible and near infrared spectroscopy. Sensors. 2012;12(8):10871–80. https://doi.org/10.3390/s120810871.
Gao Q, Zhang L. Ultraviolet-B-induced oxidative stress and antioxidant defense system responses in ascorbate-deficient vtc1 mutants of Arabidopsis thaliana. J Plant Physiol. 2008;165(2):138–48. https://doi.org/10.1016/j.jplph.2007.04.002.
Wu S, Guo Y, Joan HI, Tu Y, Adil MF, Sehar S, Zhao D, Shamsi IH. iTRAQ-based comparative proteomic analysis reveals high temperature accelerated leaf senescence of tobacco (Nicotiana tabacum L.) during flue-curing. Genom. 2020;112(5):3075–88. https://doi.org/10.1016/j.ygeno.2020.05.019.
Kanehisa M, Goto S. KEGG: kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000;28(1):27–30. https://doi.org/10.1093/nar/28.1.27.
Wan X, Yang J, Ahmed W, Liu Q, Wang Y, Wei L, Ji G: Functional analysis of pde gene and its role in the pathogenesis of Xanthomonas oryzae pv. oryzicola. Infect Genet Evol. 2021;94:105008. https://doi.org/10.1016/j.meegid.2021.105008.
Duan F, Song W. Overexpression of SoCYP85A1 Increases the Accumulation of Castasterone and Confers Enhanced Black Shank Tolerance in Tobacco Through Modulation of the Antioxidant Enzymes’ Activities. Front Plant Sci. 2019;10:349. https://doi.org/10.3389/fpls.2019.00349.
Horváth E, Szalai G, Janda T. Induction of abiotic stress tolerance by salicylic acid signaling. J Plant Growth Regul. 2007;26(3):290–300. https://doi.org/10.1007/s00344-007-9017-4.
Thipyapong P, Stout MJ, Attajarusit J. Functional analysis of polyphenol oxidases by antisense/sense technology. Molecules. 2007;12(8):1569–95. https://doi.org/10.3390/12081569.
Wang Y, Wisniewski M, Meilan R, Uratsu SL, Cui M, Dandekar A, Fuchigami L. Ectopic expression of Mn-SOD in Lycopersicon esculentum leads to enhanced tolerance to salt and oxidative stress. J Appl Hortic. 2007.
Wang B, Wang J, Zhao H, Zhao H. Stress induced plant resistance and enzyme activity varying in cucumber. Colloids Surf, B. 2006;48:138–42. https://doi.org/10.1016/j.colsurfb.2006.01.018.
Taïbi K, Taïbi F, Abderrahim LA, Ennajah A, Belkhodja M, Mulet JM. Effect of salt stress on growth, chlorophyll content, lipid peroxidation and antioxidant defence systems in Phaseolus vulgaris L. S Afr J Bot. 2016;105:306–12. https://doi.org/10.1016/j.sajb.2016.03.011.
Shetty NP, Jørgensen HJL, Jensen JD, Collinge DB, Shetty HS. Roles of reactive oxygen species in interactions between plants and pathogens. Eur J Plant Pathol. 2008;121(3):267–80. https://doi.org/10.1007/s10658-008-9302-5.
Parker J, Koh J, Yoo MJ, Zhu N, Feole M, Yi S, Chen S. Quantitative proteomics of tomato defense against Pseudomonas syringae infection. Proteomics. 2013;13(12–13):1934–46. https://doi.org/10.1002/pmic.201200402.
Cohen SP, Leach JE. Abiotic and biotic stresses induce a core transcriptome response in rice. Sci Rep. 2019;9(1):1–11. https://doi.org/10.1038/s41598-019-42731-8.
Järvi S, Suorsa M, Aro EM. Photosystem II repair in plant chloroplasts—Regulation, assisting proteins and shared components with photosystem II biogenesis. Biochimica et Biophysica Acta (BBA)-Bioenergetics. 2015;1847(9):900–9. https://doi.org/10.1016/j.bbabio.2015.01.006.
Livingstone PG, Millard AD, Swain MT, Whitworth DE. Transcriptional changes when Myxococcus xanthus preys on Escherichia coli suggest myxobacterial predators are constitutively toxic but regulate their feeding. Microb Genom. 2018;4(2). https://doi.org/10.1099/mgen.0.000152.
Fu Y, Yoon JM, Jarboe L, Shanks JV. Metabolic flux analysis of Escherichia coli MG1655 under octanoic acid (C8) stress. Appl Microbiol Biotechnol. 2015;99(10):4397–408. https://doi.org/10.1007/s00253-015-6387-6.
Baxter A, Mittler R, Suzuki N. ROS as key players in plant stress signalling. J Exp Bot. 2014;65(5):1229–40. https://doi.org/10.1093/jxb/ert375.
Bobrovskikh A, Zubairova U, Kolodkin A, Doroshkov A. Subcellular compartmentalization of the plant antioxidant system: an integrated overview. PeerJ. 2020;8: e9451. https://doi.org/10.7717/peerj.9451.
Jia X-L, Wang G-L, Xiong F, Yu X-R, Xu Z-S, Wang F, Xiong AS. De novo assembly, transcriptome characterization, lignin accumulation and anatomic characteristics: novel insights into lignin biosynthesis during celery leaf development. Sci Rep. 2015;5(1):1–14. https://doi.org/10.1038/srep08259.
Xu L, Zhu L, Tu L, Liu L, Yuan D, Jin L, Long L, Zhang X. Lignin metabolism has a central role in the resistance of cotton to the wilt fungus Verticillium dahliae as revealed by RNA-Seq-dependent transcriptional analysis and histochemistry. J Exp Bot. 2011;62(15):5607–21. https://doi.org/10.1093/jxb/err245.
Liu Q, Zheng L, He F, Zhao F-J, Shen Z, Zheng L. Transcriptional and physiological analyses identify a regulatory role for hydrogen peroxide in the lignin biosynthesis of copper-stressed rice roots. Plant Soil. 2015;387(1):323–36. https://doi.org/10.1007/s11104-014-2290-7.
Huang LF, Lin KH, He SL, Chen JL, Jiang JZ, Chen BH, Hou YS, Chen RS, Hong CY, Ho SL. Multiple patterns of regulation and overexpression of a ribonuclease-like pathogenesis-related protein gene, OsPR10a, conferring disease resistance in rice and Arabidopsis. PLoS ONE. 2016;11(6):e0156414. https://doi.org/10.1371/journal.pone.0156414.
Camoni L, Visconti S, Aducci P, Marra M. 14–3-3 proteins in plant hormone signaling: doing several things at once. Front Plant Sci. 2018;9:297. https://doi.org/10.3389/fpls.2018.00297.
Jarvis P. Targeting of nucleus-encoded proteins to chloroplasts in plants. New Phytol. 2008;179(2):257–85. https://doi.org/10.1111/j.1469-8137.2008.02452.x.
Flores-Pérez Ú, Jarvis P. Molecular chaperone involvement in chloroplast protein import. Molecular Cell Research. 2013;1833(2):332–40. https://doi.org/10.1016/j.bbamcr.2012.03.019.
Yan J, He C, Wang J, Mao Z, Holaday SA, Allen RD, Zhang HJ. Overexpression of the Arabidopsis 14–3-3 protein GF14λ in cotton leads to a “stay-green” phenotype and improves stress tolerance under moderate drought conditions. Plant Cell Physiol. 2004;45(8):1007–14. https://doi.org/10.1093/pcp/pch115.
Lee BJ, Kwon SJ, Kim SK, Kim KJ, Park CJ, Kim YJ, Park OK, Paek KH. Functional study of hot pepper 26S proteasome subunit RPN7 induced by Tobacco mosaic virus from nuclear proteome analysis. Biochem Biophys Res Commun. 2006;351(2):405–11. https://doi.org/10.1016/j.bbrc.2006.10.071.
Acknowledgements
We would like to thank Shanghai Applied Protein Biotechnology (APT Bio. Tech. Shanghai, China) (www.aptbiotech.com) for proteomics analysis.
Funding
This research work was financially supported by Yunnan Agricultural University Scientific Research Foundation grant number KX900187.
Author information
Authors and Affiliations
Contributions
Z.Z. conceived and designed the experiments. R.S. and Y.T. performed the experiments and collected the data. R.S., W.A., and G.Z. analyzed the data. R.S. and W.A. wrote the manuscript and illustrated the figures. All authors contributed to the final draft of the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This study complies with relevant institutional, national, and international guidelines and legislation for using plant material.
Consent for publication
Not applicable.
Competing interests
The authors declare they have no conflicts of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visithttp://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Song, R., Tan, Y., Ahmed, W. et al. Unraveling the expression of differentially expressed proteins and enzymatic activity in response to Phytophthora nicotianae across different flue-cured tobacco cultivars. BMC Microbiol 22, 112 (2022). https://doi.org/10.1186/s12866-022-02531-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12866-022-02531-z