
Abstract
Background
B-box (BBX) proteins play important roles in regulating plant growth, development, and abiotic stress responses. BBX family genes have been identified and functionally characterized in many plant species, but little is known about the BBX family in blueberry (Vaccinium corymbosum).
Result
In this study, we identified 23 VcBBX genes from the Genome Database for Vaccinium (GDV). These VcBBXs can be divided into five clades based on gene structures and conserved domains in their encoded proteins. The prediction of cis-acting elements in the upstream sequences of VcBBX genes and protein–protein interactions indicated that VcBBX proteins are likely involved in phytohormone signaling pathways and abiotic stress responses. Analysis of transcriptome deep sequencing (RNA-seq) data showed that VcBBX genes exhibited organ-specific expression pattern and 11 VcBBX genes respond to ultraviolet B (UV-B) radiation. The co-expression analysis revealed that the encoded 11 VcBBX proteins act as bridges integrating UV-B and phytohormone signaling pathways in blueberry under UV-B radiation. Reverse-transcription quantitative PCR (RT-qPCR) analysis showed that most VcBBX genes respond to drought, salt, and cold stress. Among VcBBX proteins, VcBBX24 is highly expressed in all the organs, not only responds to abiotic stress, but it also interacts with proteins in UV-B and phytohormone signaling pathways, as revealed by computational analysis and co-expression analysis, and might be an important regulator integrating abiotic stress and phytohormone signaling networks.
Conclusions
Twenty-three VcBBX genes were identified in blueberry, in which, 11 VcBBX genes respond to UV-B radiation, and act as bridges integrating UV-B and phytohormone signaling pathways according to RNA-seq data. The expression patterns under abiotic stress suggested that the functional roles of most VcBBX genes respose to drought, salt, and cold stress. Our study provides a useful reference for functional analysis of VcBBX genes and for improving abiotic stress tolerance in blueberry.
Similar content being viewed by others


Introduction
Zinc-finger transcription factors are important regulatory factors that play essential roles in plant growth, development, and responses to environmental changes [1, 2]. Zinc-finger protein family members are divided into multiple subfamilies based on their structures and functions [3]. B-box (BBX) proteins, comprising a subfamily of zinc-finger transcription factors, contain one or two conserved B-box domains at their N termini. The B-box domain is involved in protein–protein interactions and transcriptional regulation during plant signaling [4]. Some BBX proteins contain a CCT (CONSTANS, CO-like and TOC1) domain at their C termini and are referred to as CO-like (COL); the CCT domain functions in transcriptional regulation and protein targeting to the nucleus [4, 5].
BBX genes have been identified in various plant species, and their functions have been demonstrated at the molecular level [6, 7]. BBX proteins are mainly involved in photomorphogenesis, plant hormone signaling networks, and abiotic stress responses [6]. In Arabidopsis, BBX4, BBX11, BBX20, BBX21, BBX22, and BBX23 promoter photomorphogenesis, while BBX18, BBX19, BBX24, BBX25, BBX28, BBX29, BBX30, BBX31 and BBX32 repress photomorphogenesis in response to a wide range of light signals [8,9,10]. BBX4 and BBX11 promote hypocotyl elongation in red light [9, 11]. BBX11, BBX20, BBX21, BBX22 and BBX23 interact with LONG HYPOCOTYL 5 (HY5) to promote its transcriptional activation activity and positively regulate light-mediated seedling development, whereas BBX24, BBX25, BBX28, BBX29, and BBX32 interact with HY5 to form heterodimers and repress photomorphogenesis in response to light signals [10, 12,13,14,15,16,17,18]. For BBX30 and BBX31, HY5 directly binds to their promoter to inhibit plant photomorphogenesis [17]. some studies have shown that BBX proteins also respond to UV-B radiation. For example, BBX24 from Arabidopsis represses the transcriptional activity of HY5 to negatively regulate UV-B-mediated photomorphogenic response [19]. In apple, MdCOL4/MdBBX54 (MDP0000232445) suppresses anthocyanin accumulation under UV-B and high temperature by interacting with MdHY5 to synergistically inhibit the expression of MdMYB1 [20, 21]. However, MdBBX22 (MDP0000298804) and MdBBX20 (MDP0000800387) promotes UV-B-induced anthocyanin biosynthesis through interacting with MdHY5 [22, 23]. At the same time, MdCOL11/MdBBX33 (MDP0000697407), a target of MdHY5, enhances UV-B- and temperature-induced anthocyanin biosynthesis in apple peel [24]. MdBBX37 (MDP0000157816), a negative regulator of light signaling, interacts with MdMYB1 and MdMYB9 to negatively regulate anthocyanin biosynthesis and directly binds to the promoter of MdHY5 to relieve MdHY5-mediated hypocotyl inhibition [25]. Thus, BBX proteins regulate photomorphogenesis by interacting with MdHY5.
Several studies have indicated that BBX proteins play integrated role in phytohormones signaling pathway-mediated cellular and developmental process. For instance, in high light conditions, BBX16 integrates light and auxin to regulate plant shoot branching by the activating expression of SUPERROOT 2 (SUR2), which is a suppressor of auxin biosynthesis [26]. In gibberellin (GA) signaling pathway, BBX18 promotes hypocotyl elongation under blue light condition by regulating the expression of GA biosynthesis or metabolic genes [27]. BBX24 is a positive regulator of GA signaling by interacting and inhibiting DELLA activity [28]. In brassinosteroid (BR) signaling pathway, BBX20/BZS1, BBX28 and BBX29 negatively regulate BR signaling in Arabidopsis seedlings, in which BBX20 interacts with COP1 in vitro to positively regulate light signaling, and BBX28 and BBX29 physically interact with BR-ENHANCED EXPRESSION1 (BEE1), BEE2, and BEE3 to orchestrate light and BR signaling. The BBX32 interacts with PIF3 and BZR1 to negatively regulate light signaling, promote BR signaling and inhibit cotyledon opening in Arabidopsis [29,30,31]. Therefore, BBX20, BBX28, BBX29 and BBX32 mediate the crosstalk between BR and light pathways in the different ways. In abscisic acid (ABA) signaling pathway, BBX19 directly binds to promoter of ABA INSENSITIVE5 (ABI5) to negatively regulate seed germination by promoting ABA signaling [32]. In addition, OsCOL9 interacts with OsRACK1 to enhance the rice blast resistance through SA and ET signaling pathways and BBX21 downregulates the gene expression of auxin, BR and ethylene signaling pathway components under shade [7, 33]. In conclusion, BBX proteins integrate phytohormones and light signals to affect light-mediated plant growth and development [8].
In addition to photomorphogenesis and phytohormone signaling pathways, most BBX proteins also participate in plant responses to abiotic stress (cold, salt, and drought). For example, CmBBX19 from chrysanthemum (Chrysanthemum morifolium) interacts with CmABF3 to suppress drought tolerance, while CmBBX22 enhances drought tolerance possibly via regulating the ABA signaling pathway [34,35,36]. CmBBX24 improves tolerance to freezing and drought stress through influencing bioactive GA biosynthesis [37]. In Arabidopsis, BBX7 and BBX8 act downstream of CRYPTOCHROME2 (CRY2)-CONSTITUTIVELY PHOTOMORPHOGENIC 1 (COP1)-HY5 module to positively regulate blue light-dependent cold acclimation [38]. BBX24/STO binds to a Myb transcription factor homologue and enhances salt tolerance [39]. MdBBX10 from apple enhances tolerance to salt and drought stresses in Arabidopsis and involves in ABA-mediated response [40]. OsBBX11 from rice (Oryza sativa) regulates salt tolerance [41]. Thus, BBX proteins regulate abiotic stress response by interaction with transcript factors or phytohormone signaling pathway.
Blueberry (Vaccinium corymbosum) is an economically important small fruit crop that is often referred to as a “superfood” due to its high nutrient content and health benefits [42, 43]. Although BBX genes have been identified in various plant species via genome-wide studies based on complete plant genome sequences, a comprehensive study of BBX genes in blueberry has not yet been reported [21, 44,45,46,47]. A recent study showed that VcBBX21, VcBBX30, and VcBBX32 respond to UV-B radiation [48]. However, the roles of BBX family members in abiotic stress responses in blueberry remain unknown. The release of the Vaccinium genus genome database offers the possibility to systematically identify and investigate the putative functions of BBX genes in blueberry. For V. corymbosum, the W8520 draft genome sequence was originally performed on March 2014, and then genome sequence was annotated and made available [49, 50]. The sequencing and assembly of the V. corymbosum cv. Draper genome was completed and the function was annotated on March 2019, this genome database was widely used because of high quality [51, 52].
In this study, we identified 23 blueberry BBX genes from the Genome Database for Vaccinium (GDV) and predicted their gene structures, as well as the physicochemical properties, evolutionary relationships, and domains of the encoded proteins. We then analyzed the expression patterns of VcBBX genes in different organs and the responses of the VcBBX genes to UV-B radiation based on transcriptome deep sequencing (RNA-seq) data. At the same time we examined the relationships between VcBBXs and phytohormone pathway genes under UV-B radiation based on co-expression analysis. Finally, we analyzed the expression levels of VcBBXs under cold, salt, and drought stress. Our results provide a foundation for further functional characterization of VcBBX genes in blueberry.
Results
Identification and physicochemical properties of VcBBX family members in blueberry
To identify BBX genes in the blueberry genome, we used Arabidopsis and apple BBX genes as a query to search against the GDV. We also conducted a Hidden Markov Model (HMM) search using the B-box domain (pfam00643) against the GDV. After removing short and redundant sequences, we identified 23 putative VcBBX members; detailed information is provided in Table 1 and Table S1. The VcBBX proteins ranged from 125 (VcBBX31) to 480 (VcBBX12) amino acids in length, with molecular weights ranging from 14.2 kDa (VcBBX31) to 53.0 kDa (VcBBX12). The theoretical isoelectric point (pI) was highest for VcBBX31 (8.82) and lowest for VcBBX28 (4.38). The instability index (Ii) was the lowest for VcBBX12 (43.76) and the highest for VcBBX32 (59.72) in all the VcBBX proteins, indicating that these proteins are unstable. The aliphatic index (AI) ranged from 47.70 (VcBBX29) to 71.39 (VcBBX24). The GRAVY (grand average of hydropathicity) values were negative except for VcBBX23, indicating that most VcBBXs are hydrophilic proteins. Subcellular localization of VcBBXs (predicted using the WoLF PSORT program) indicated that most VcBBX proteins are located in the nucleus. However, VcBBX6, VcBBX18, and VcBBX19 were predicted to be in the cytosol, while VcBBX23 and VcBBX29 were predicted to localize to chloroplasts, suggesting that VcBBX proteins have diverse functions.
Phylogenetic analysis of VcBBX proteins
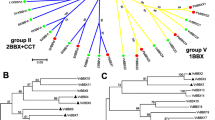
To investigate the evolutionary relationships of VcBBX family members, we reconstructed a phylogenetic tree based on the amino acid sequences of the 23 blueberry VcBBXs and 32 Arabidopsis AtBBXs (Fig. 1; Table S2). The VcBBXs were divided into five clades and named based on phylogenetic analysis and previous studies in Arabidopsis [5]. VcBBX1 and VcBBX6 were grouped into clade I with AtBBX1–6, and VcBBX7–13 were grouped into clade II with AtBBX7–13. Proteins in these two clades contain B-box1 and B-box2 domains, as well as a CCT domain. VcBBX18–25 were grouped into clade IV with AtBBX18–25; proteins in this clade contained B-box1 and B-box2 domains. Clade III proteins, comprising VcBBX15 and AtBBX15–17, contain B-box1 and CCT domains. Clade V consists of VcBBX28–32 and AtBBX26–32, which only contain a B-box1 domain (Figs. 1 and 2A).

Phylogenetic analysis of BBX proteins in blueberry (Vaccinium corymbosum) and Arabidopsis (Arabidopsis thaliana). White circles represent BBX proteins from Arabidopsis. Red, green, blue, and black dots represent blueberry BBX genes that are upregulated, downregulated, up- or downregulated, and not regulated by UV-B radiation, respectively

Phylogenetic relationships and structural analysis of blueberry BBXs. A Phylogenetic analysis of VcBBX proteins: different colors represent different groups and structures of VcBBX proteins. The pink, blue, and green boxes represent B-box1, B-box2, and CCT domains, respectively. B Structures of VcBBX genes. Dark green boxes, orange boxes, and black lines represent UTRs, exons, and introns, respectively
To explore the structures of the 23 VcBBX genes in blueberry, we obtained exon/intron information about them based on their coding and genomic sequences from the GDV. The number of exons ranged from one to five. VcBBX29 and VcBBX31 contain no introns or untranslated regions (UTRs), and VcBBX23 also lacks a UTR. Similar results were found in BBX families of tomatoes and peaches [45, 53]. Similarly, VcBBX genes with similar gene structures clustered into the same clade. For example, members of clade II contain four or five exons, and members of clade I harbor two exons (Fig. 2B).
Analysis of phytohormone- and abiotic stress–related cis-acting elements
To better understand the functions of VcBBX genes, we collected their promoter sequences 2000 bp upstream from the ATG start codon and predicted the cis-acting elements involved in phytohormone and abiotic stress responses using the PlantCARE online tool (Fig. 3). The VcBBX promoters contained various cis-acting elements related to phytohormones, such as ABA-, auxin-, SA-, GA-, and methyl jasmonate (MeJA)-responsive elements. These promoters also contained many abiotic stress-related elements, especially drought-inducible and low-temperature-responsive elements.

Diagram of the predicted regulatory cis-elements involved in phytohormone and abiotic stress responses in the VcBBX promoters. The black line below the diagram indicates the lengths of the VcBBX promoters. The different colored symbols represent cis-acting elements with different functions, as described below the diagram
VcBBX protein interactions
To predict the functions of the VcBBXs, we assembled a protein interaction network for each member using their orthologs from Arabidopsis (Fig. 4). The 23 VcBBX proteins in blueberry correspond to 15 BBX/COL proteins in Arabidopsis. The acronyms and full names of the interactors are provided in Table S3. These proteins interacted not only with BBX proteins (STH, BZS1, BBX32, BBX8, CO, BBX9, BBX31, and COL1), but also with other proteins such as transcription factors (MYB: MYB1, MYB10, MYB16; basic helix-loop-helix [bHLH]: HFR1; bZIP: HY5, HYH; zinc-finger: CDF3), an E3 ubiquitin-protein ligase (COP1), and proteins involved in the circadian clock (XCT, PCL1, LHY, ELF3, ELF4, ELF4-LIKE3, and ELF4-LIKE4). VcBBX29/30/32 (BBX29 ortholog), VcBBX11 (At2g47890 ortholog), VcBBX6 (COL5 ortholog), and VcBBX22/24/25 (COL2 ortholog) were predicted to interact with proteins in the ethylene signaling pathway (DEAR3, RAP2.7, TOE2, and RCD1), while VcBBX20/21 (BBX21 ortholog) and VcBBX22/24/25 (STO/BBX24 ortholog) likely interact with proteins in the ABA signaling pathway (ABI5 and ABA1). We also observed that VcBBX29/30/32 (BBX29 ortholog) and VcBBX7/8 (COL9 ortholog) are predicted to interact with COR27, which is involved in cold stress responses. These results indicate that VcBBX proteins may be participated in plant growth and development, as well as plant responses to abiotic stress and phytohormone signaling pathways.

Protein interaction networks predicted for 23 VcBBX proteins based on their orthologs in Arabidopsis using the online tool STRING. The red balls indicate VcBBX proteins; the names of the orthologs in blueberry are shown in brackets. The other colored balls represent individual VcBBX interactors. The blue and purple lines represent known interactions from curated databases and those that were experimentally determined, respectively. The green, red, and blue lines represent predicted interactions from gene neighborhood, gene fusions, and gene co-occurrence, respectively. The yellow-green, black, and blue lines represent text mining, co-expression, and protein homology, respectively. The acronyms define proteins that have been reported in Arabidopsis
The organ-specific expression pattern analysis of VcBBX genes in blueberry
To provide clues to the putative roles of the VcBBX genes in blueberry development, the expressions of VcBBX family genes were analyzed in flower bud, flower at anthesis, petal fall, green fruit, pink fruit, ripe fruit root, shoot, leaf at day, and leaf at night from transcriptome sequencing results and the heatmap of organic-specific expressions was drawn based on log10 (FPKM) values (Fig. 5). The VcBBXs were divided into five groups and same groups shared similar expression patterns. The VcBBX13, VcBBX9, VcBBX10, VcBBX20 and VcBBX23 were clustered in group A because they did not expressed or showed low expression in some organs. The members of group B (VcBBX6, VcBBX8 and VcBBX9) and group C (VcBBX28, VcBBX29, VcBBX12 and VcBBX7) showed low expression during fruit development, however, the expression levels of VcBBX members from group B are higher than that from group C in root, shoot, leaf, and flower. The VcBBX18, VcBBX24, VcBBX1, and VcBBX25 were clustered in the group D and showed high expression in all the organs and the expression of VcBBX31, VcBBX30, VcBBX21, and VcBBX32 from group E were higher in the flowers and fruits than in the root, shoot and leaf. These results indicated that most blueberry VcBBX genes showed organ-specific expression pattern, potentially suggesting the functional divergence of VcBBX genes. The VcBBX18, VcBBX24, VcBBX1, VcBBX25, VcBBX31, VcBBX30, VcBBX21, and VcBBX32 may play important role in regulation of flower and fruit development.

Transcript profiling of VcBBX genes in various organs based on log10 (FPKM) values from RNA-seq data. The color scale is shown on the right, block with green indicated low expression levels, while red indicated high expression levels. The gray block indicated that the gene was not expressed
VcBBX gene expression in response to UV-B radiation
To reveal the roles of VcBBXs in plant responses to abiotic stress, we downloaded RNA-seq data from the BioProject database, which were obtained from blueberry calli treated for 0, 1, 3, 6, 12, and 24 h with UV-B radiation [52]. Eleven VcBBXs were responsive to UV-B radiation, including VcBBX8, VcBBX15, and VcBBX21 (from cladesII, III, and IV, respectively), which were downregulated; and VcBBX1, VcBBX6, VcBBX11, VcBBX24, VcBBX28, VcBBX30, and VcBBX32 (from various clades), which were upregulated. VcBBX31 was upregulated at 3 and 6 h and downregulated at 12 and 24 h of UV-B treatment (Fig. 6A and Table S4). These results suggest that VcBBXs from different clades might participate in the same regulatory pathways. In all the differentially expressed VcBBX genes, VcBBX6 expression was upregulated during UV-B treatment and the value of log10 (FC) reached 2.09 (1 h), 4.01 (3 h), 3.61 (6 h), 2.84 (12 h), and 2.16 (24 h) under UV-B treatment. The expression of VcBBX30 also was upregulated from 3 h onwards, reaching the highest level at 24 h (5.16 for log10 (FC) value). However, VcBBX15 was downregulated from 1 h onwards, reaching the lowest level (-3.22 for log10 (FC) value) at 6 h in expression levels.

Expression analysis of VcBBX genes under UV-B radiation. A Transcript profiling of VcBBX genes under UV-B radiation based on log10 (FPKM) values from RNA-seq data. The color scale is shown on the right, block with green indicated low expression levels, while red indicated high expression levels. The gray block indicated that the gene was not expressed. B Gene expression patterns of VcBBX genes under UV-B radiation using qRT-PCR analysis. Values are means ± SD from three independent biological replicates, each with three technical replicates; Statistically significant differences were assessed using Student’s t-test (* p < 0.05)
To validate the accuracy and reliability of the RNA-seq data under UV-B radiation, expression levels of eleven differentially expressed VcBBX genes under UV-B radiation for 0, 1, 3, 6, 12, and 24 h were analyzed by the RT-qPCR (Fig. 6B). Consistent with RNA-Seq data, the expression of VcBBX1, VcBBX6, VcBBX11, VcBBX24, VcBBX28, VcBBX30, and VcBBX32 significantly upregulated and that of VcBBX8, VcBBX15, and VcBBX21 significantly downregulated under UV-B treatment compared to the 0-h control. Furthermore, the relative expression levels of VcBBX6 significantly increased 10.76-, 11.66-, 8.89-, 5.9–3, and 3.71-fold at 1, 3, 6, 12, and 24 h of UV-B treatment relative to the 0 h treatment, respectively. The expression of VcBBX30 also was significantly upregulated during UV-B treatment and reached the highest level at 3 h (9.69-fold compared to the 0-h control). We also found that VcBBX24 and VcBBX32 also was significantly upregulated and increased more than seven-fold during UV-B treatment compared to the 0-h control. RNA-seq data and RT-qPCR analysis showed that VcBBX6, VcBBX24, VcBBX30 and VcBBX32 maybe play important roles under UV-B radiation.
Correlations between VcBBX genes and phytohormone pathway genes under UV-B radiation
The VcBBX promoter sequences contain various cis-acting elements related to phytohormones. Thus, we searched for differentially expressed genes (DEGs) from phytohormone pathways in RNA-seq data set of UV-B treatment and performed Pearson’s correlation coefficient (r) analysis between the VcBBXs and DEGs in various phytohormone pathways according to FPKM values (Table S5). Forty-eight genes from the auxin pathway showed a significant correlation in their expression levels with various VcBBX genes. Specifically, 27 auxin pathway genes showed a significant correlated in their expression with that of VcBBX21, 20 with VcBBX30, and 18 with VcBBX15; however, VcBBX28 expression levels were only significantly correlated with those of AUXIN RESPONSE FACTOR 18a (ARF18a). Forty-one genes in the ethylene pathway exhibited a significant correlation with VcBBX gene expression. Of these, VcBBX21 expression was significantly correlated with the largest number of genes (17) from the ethylene pathway, followed by VcBBX30 (16) and VcBBX15 (14), while VcBBX28 expression was only significantly correlated with that of ETHYLENE-RESPONSE FACTOR 3 (ERF3). The expression levels of 50 BR pathway genes were significantly correlated with those of VcBBXs. VcBBX21 was correlated with the most genes (25), followed by VcBBX30 (20), VcBBX32 (18), and VcBBX15 (15). Finally, the expression of 13, 16, and 7 genes was significantly correlated with that of VcBBX genes in the SA, GA, and ABA pathways, respectively, with VcBBX21, VcBBX30, and VcBBX32 expression being significantly correlated with the most number of genes from these three phytohormone pathways. At the same time, VcBBX24 also was significantly correlated with genes from auxin (7), GA (2), ABA (2), Ethylene (5), BR (6) and SA (2) under UV-B radiation (Fig. 7; Table S5).

Co-expression network analysis of DEGs between VcBBXs and plant hormone signaling–related genes under UV-B radiation. Red lines indicate positive correlations; blue lines indicate negative correlations. The thickness of the line represents the degree of correlation. The size of the circle represents the number of related genes
Expression analysis of VcBBX genes under abiotic stress
To further explore the roles of VcBBXs in plant responses to abiotic stress, we evaluated the expression patterns of the 23 VcBBX genes in the leaves of plants subjected to drought, salt, or cold stress by RT-qPCR (Fig. 8). The VcBBX genes exhibited different expression patterns in response to drought. The expression of most VcBBX genes (16/23) was significantly upregulated in response to drought treatment, especially VcBBX10, VcBBX13, VcBBX15, VcBBX19, VcBBX20, VcBBX24, and VcBBX31. However, the expression of VcBBX1, VcBBX6, VcBBX29, and VcBBX30 was significantly downregulated in response to this stress, and VcBBX8, VcBBX9, and VcBBX21 were not responsive to 6 or 12 h of drought stress (Fig. 8A). Fifteen VcBBX genes were responsive to salt stress, with twelve VcBBX genes being upregulated and three being downregulated. The relative expression levels of VcBBX8, VcBBX9, VcBBX11–13, VcBBX19, VcBBX20, VcBBX23, VcBBX24, and VcBBX29 increased more than two-fold following 6 or 12 h of salt stress treatment compared to the 0-h control (Fig. 8B).

Expression patterns of the 23 VcBBX genes in response to three abiotic stresses. A VcBBX expression under drought stress (20% PEG-6000). B VcBBX expression under salt stress (200 mM NaCl). C VcBBX expression under cold stress (5℃). All treatments were performed for 6 or12 h. Values are means ± SD from three independent biological replicates, each with three technical replicates; Statistically significant differences were assessed using Student’s t-test (* p < 0.05)
The expression of 21 VcBBX genes responded to cold treatment. The twelve VcBBX genes were upregulated and six were downregulated at 6 or 12 h of cold treatment. In which, the transcript levels of VcBBX9, VcBBX13, and VcBBX21 rapidly decreased at 6 h of cold treatment and increased at 12 h of treatment. The transcript levels of VcBBX1, VcBBX7–9, VcBBX19–21, VcBBX24, VcBBX30, and VcBBX31 increased more than two-fold at 6 or 12 h of treatment compared to the control (Fig. 8C). Many VcBBX genes also showed different responses to different stresses; for example, VcBBX22 was induced by drought, inhibited by cold, and did not respond to salt stress. Importantly, most VcBBX genes were induced by drought, cold, and salt stress, indicating that the roles of VcBBX genes in abiotic stress are conserved in plants. In summary, VcBBX19, VcBBX20, and VcBBX24 were signifcantly upregulated by drought, salt, and cold stress and increased more than two-fold at 6 or 12 h of treatment relative to the 0 h treatment, indicating that VcBBX19, VcBBX20, and VcBBX24 may be involved in a variety of abiotic stresses.
Discussion
Structure of VcBBX genes and evolutionary analyses
The BBX gene family has been identified in many plants, including fruit crops. However, the number of BBX family members differs among plant species. To date, 32 BBX genes have been identified in Arabidopsis, 64 in apple, 21 in strawberry (Fragaria vesca), 25 in pear (Pyrus communis), 24 in grapevine (Vitis vinifera) and peanut (Arachis duranensis), 22 in peach (Prunus persica) and 15 in sweet cherry (Prunus avium) [5, 21, 33, 46, 54,55,56,57]. In this study, after deleting short and redundant sequences, we identified 23 full length BBXs in blueberry. Thus, blueberry contains fewer BBX genes than most other plant species. These differences may be due to differences in genome size as well as tandem and segmental duplication events among plant species. However, the composition of conserved domains in the encoded BBX proteins of blueberry is similar to that of other species. We identified four types of blueberry BBX proteins: those with two tandem B-boxes and one CCT domain, those with one B-box domain and one CCT domain, those with two tandem B-boxes, and those with one B-box domain [5, 46, 53].
Most studies have divided BBX family members into five clades, and BBX proteins with the same types of conserved domains clustered together. For example, BBX proteins from pear, Arabidopsis, purple false brome (Brachypodium distachyon), rice, and black cottonwood (Populus trichocarpa) were divided into five groups: group I or II proteins contain B-box1, B-box2, and CCT domains; group III proteins contain B-box1 and CCT domains; group IV proteins contain B-box1 and B-box2 domains; and group V proteins contain B-box1 domains [55]. We obtained the same results for blueberry BBX family members. At the same time, the number of exons in BBX genes belonging to the same clade were similar in blueberry and other species [45, 55]. These results suggest that BBX families in different species might share a common ancestor and similar evolutionary patterns to adapt to the environment.
The function predication of VcBBX genes based on organ-specific expression analysis
Gene expression analysis in essential for providing clues for functional prediction [44, 45]. In this study, VcBBX genes showed distinct expression patterns among organs. In which, VcBBX1, 6, and 8 showed a high expression in flower bud, flower at anthesis, and petal fall and they also have one CCT domain, Some study showed that BBX proteins with a CCT domain play critical role in flowering, suggesting that VcBBX1, 6, and 8 play important roles in flowering [5, 58]. Most VcBBX genes come from groups IV and V, including VcBBX18, VcBBX21, VcBBX24, VcBBX25, and VcBBX30-32, have higher expression in petal fall, green fruit, pink fruit and ripe fruit than other VcBBX genes. The groups IV and V contain one or two B-box domains without CCT domain. At same time, VcBBX21, VcBBX24, and VcBBX30-32 response to UV-B radiation, and UV-B radiation involve in color of flower and fruit [48]. In apple, MdBBX20, MdBBX22, MdBBX33 and MdBBX54 of group IV with two B-box domains without CCT domain regulate UV-B-induced anthocyanin biosynthesis and MdBBX37 of group V with one B-box domain without CCT domain also regulates anthocyanin biosynthesis [20, 22,23,24]. In pear, PpBBX16, PpBBX18 and PpBBX21, with two B-box domains without CCT domain, regulate anthocyanin accumulation [59, 60]. The previous study showed that VcBBX30 and VcBBX32 co-expressed with the possible anthocyanin biosynthesis gene VcMYBA2 and VcMYB114 under UV-B radiation [48]. Thus, VcBBX21, VcBBX24, and VcBBX30-32, especially VcBBX30 and VcBBX32, may regulate UV-B-induced anthocyanin biosynthesis. Above all we speculated that B-box domains might play critical role in anthocyanin biosynthesis. In this study, we also found that VcBBX18, VcBBX24, VcBBX1, and VcBBX25 were high expression in all the organs indicating that they have various roles in development of blueberry different organs.
The function predication of VcBBX genes under UV-B radiation
Light is a major environmental factor that regulates physiology and development in plants, such as photomorphogenesis, flowering, and anthocyanin accumulation. As a component of natural light, UV-B also regulates photomorphogenic responses. Inhibition of hypocotyl growth and promotion of anthocyanin accumulation are the most obvious effects of excess UV-B exposure [61,62,63,64,65]. BBX proteins play essential roles in these UV-B-mediated responses, and most BBX genes are induced by UV-B light in Arabidopsis [66]. AtBBX24 interacts with COP1 and HY5 in UV-B-induced inhibition of hypocotyl elongation, and AtBBX31 regulates UV-B-mediated photomorphogenesis in a HY5-dependent manner [19, 67, 68]. In apple, MdBBX22 promotes UV-B-induced anthocyanin biosynthesis by interacting with MdHY5, and MdCOL4 interacts with MdHY5 to synergistically inhibit anthocyanin accumulation under UV-B radiation [22, 23].
In the current study, 11 of the 23 VcBBX genes from five different clades responded to UV-B radiation, consisting of seven VcBBX genes, three downregulated genes, and one that was upregulated and then downregulated during UV-B treatment. Thus, most VcBBX genes are likely involved in UV-B-induced photomorphogenic responses; however, their specific functions require further study. The predicted protein interaction networks based on orthologous BBXs of Arabidopsis showed that VcBBX18–25 interact with HY5, HY5 HOMOLOG (HYH), and COP1. The previous study showed that VcBBX30 and VcBBX32 co-expressed with the VcHY5, and VcHY5 co-expressed with COP1 under UV-B radiation [48]. Moreover, VcBBX21, VcBBX24, VcBBX30, and VcBBX32 were regulated by UV-B radiation. Thus, VcBBX21, VcBBX24, VcBBX30, and VcBBX32 might interact with HY5, HYH, or COP1 to regulate UV-B-induced photomorphogenesis or anthocyanin accumulation.
The VcBBX genes may involved in phytohormone signaling pathways under UV-B radiation
BBX proteins play important roles in phytohormone signaling pathways [33, 34]. We determined that all sequences upstream of VcBBX genes contain ABA-, auxin-, SA-, GA-, and MeJA-responsive elements. The expression levels of 11 VcBBXs were significantly correlated with those of genes from the auxin, ethylene, BR, SA, GA, and ABA signaling pathways during UV-B radiation, as revealed by co-expression analysis based on RNA-seq data (Figs. 3 and 4). Thus, most blueberry VcBBX proteins might function in phytohormone signaling pathways. Several studies have shown that BBX proteins are involved in the crosstalk between light and phytohormone signaling during plant development [27, 37, 69]. However, little is known about how BBX genes coordinate with genes from phytohormone signaling pathways to regulate UV-B-induced physiology and development in plants. AtBBX24 interacts with DELLA proteins in the GA pathway to regulate UV-B-induced photomorphogenesis in Arabidopsis [28, 70]. We determined that 11 VcBBXs respond to UV-B radiation, and their expression was significantly correlated with that of many genes involved in phytohormone signaling pathways, especially the auxin, ethylene, and BR pathways. The expression of VcBBX21, VcBBX30, VcBBX32, and VcBBX15 was significantly correlated with that of most genes involved in phytohormone signaling pathways under UV-B radiation. In addition, VcBBX6, VcBBX11, VcBBX20, VcBBX21, VcBBX22, VcBBX24, VcBBX25, VcBBX29, VcBBX30, and VcBBX32 maybe interact with proteins in the ethylene or ABA signaling pathway, however protein interaction tests are needed for further confirmation. Thus, we propose that VcBBXs may serve as bridges integrating UV-B signaling and phytohormone signaling pathways in blueberry.
The expression pattern of VcBBX genes under abiotic stresses
BBX genes are involved in various stress responses in many plant species [21, 34]. For example, overexpressing AtBBX24 (also named SALT TOLERANCE [AtSTO]) improved salt tolerance in Arabidopsis, and heterologous expression of CmBBX22 from chrysanthemum (Chrysanthemum morifolium) improved drought tolerance in this plant [35, 39]. Most BBX genes also respond to salt, drought, and cold stress [35, 39, 47, 71]. Our study showed that 20 VcBBX genes respond to drought stress, 15 to salt stress, and 21 to cold stress, demonstrating that most VcBBX genes generally respond to abiotic stress. VcBBX13, VcBBX19, VcBBX20, VcBBX23, and VcBBX24 were upregulated and VcBBX6 was downregulated in response to all three stresses, suggesting that these genes play major roles in abiotic stress responses (Fig. 8).
VcBBX10, VcBBX13, VcBBX15, VcBBX19, VcBBX20, VcBBX24, and VcBBX31 were most strongly upregulated by drought stress among all VcBBX genes, suggesting that they might play major roles in plant responses to drought stress. VcBBX1, VcBBX7–9, VcBBX19–21, VcBBX24, VcBBX30, and VcBBX31 were most strongly upregulated by cold treatment. Among their encoded proteins, VcBBX7, VcBBX8, and VcBBX30 were predicted to interact with COR27, suggesting that these proteins play major roles in regulating the cold stress response. Moreover, VcBBX8, VcBBX9, VcBBX11–13, VcBBX19, VcBBX20, VcBBX23, VcBBX24, and VcBBX29 were mainly regulated by salt stress. In conclusion, VcBBXs play important roles in plant responses to abiotic stress. VcBBX24 responds to UV-B radiation, drought stress, salt stress, and cold stress, and VcBBX24 interacts with UV-B signaling pathway proteins (COP1 and HY5), an ABA signaling pathway protein (ABA-INSENSITIVE 1 [ABA1]), and an ethylene signaling pathway protein (RADICAL-INDUCED CELL DEATH1 [RCD1]). Similarly, AtBBX24 (VcBBX24 ortholog) is involved in plant responses to salt stress and UV-B radiation in Arabidopsis [39]. Thus, we propose that VcBBX24 is an important component of the abiotic stress regulatory network in blueberry.
Conclusions
In this study, we identified 23 putative VcBBX genes in the blueberry genome and classified them into five clades based on phylogenetic analysis and the conserved motifs of their encoded proteins. Analysis of cis-acting elements and prediction of protein interactions suggested that VcBBX genes participate in diverse phytohormone pathways and abiotic stress responses. The organ-specific expression analysis showed that VcBBXs played various roles in blueberry development. Analysis of RNA-seq data and co-expression analysis indicated that VcBBX genes function in plant responses to UV-B radiation and act as bridges between UV-B signaling and phytohormone signaling pathways. Furthermore, most VcBBX genes respond to drought, salt, and cold stress. These results provide valuable information for further analysis of the functions of VcBBX genes to guide breeding efforts for improved abiotic stress resistance in blueberry.
Materials and methods
Identification of BBX genes in the blueberry genome
To identify BBX genes in blueberry, two different procedures were used. First, the sequences of BBX genes from Arabidopsis (Arabidopsis thaliana) and apple (Malus domestica) were downloaded from The Arabidopsis Information Resource database (TAIR, https://www.arabidopsis.org/) and the Genome Database for Rosaceae (GDR, https://www.rosaceae.org/), respectively, and used as a query against the Genome Database for Vaccinium corymbosum cv. Draper V1.0 genome sequence (GDV, https://www.vaccinium.org/). Second, Hidden Markov Model (HMM) searches were performed in the GDV using the B-box domain (pfam00643) from the Pfam database (http://pfam.xfam.org/). The potential VcBBX genes in blueberry were investigated using the online programs UniProt (https://ww.uniprot.org/), SMART (http://smart.embl-heidelberg.de/), and CDD (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi), and genes without BBX domains in their encoded proteins were removed. Finally, a list of VcBBX genes encoding complete BBX domains was obtained by deleting shorter and redundant sequences.
The VcBBX members were named based on clustering with Arabidopsis proteins and functional annotations from a manually annotated and reviewed protein sequence databaset (Swiss-Prot). The molecular weight (MW), theoretical isoelectric point (pI), instability index (Ii), aliphatic index (AI), and grand average of hydropathicity (GRAVY) of the VcBBX proteins were calculated using the online program ExPASy (https://web.expasy.org/protparam/), and the subcellular localization of each VcBBX protein was predicted using the online program WoLF PSORT (https://www.genscript.com/wolf-psort.html?src=leftbar).
Phylogenetic analysis
The phylogenetic trees were reconstructed in MEGA X using the neighbor-joining method. Bootstrap analysis was carried out with 1000 replicates [72]. Arabidopsis sequences were downloaded from NCBI (https://www.ncbi.nlm.nih.gov/). The phylogenetic trees were divided into different clades refered studies on Arabidopsis [5].
Analysis of gene structures and conserved domains
The cDNA sequences and the corresponding untranslated regions (UTRs), exons, and introns of VvBBX family members were downloaded from the GDV and gene structure was drawn with TBtools software (v1.120) [73]. Conserved domains including B-box and CCT domains were identified using the online programs UniProt, SMART, and CDD.
Analysis of cis-acting regulatory elements in the promoters of VvBBX genes
The 2000-bp upstream sequences of VcBBX genes were downloaded from the GDV and submitted to the online program PlantCARE (https://bioinformatics.psb.ugent.be/webtools/plantcare/html/) to predict cis-acting elements. The cis-acting elements related to phytohormone and abiotic stress responses were retained and mapped along the presumptive promoters.
Construction of a protein interaction network
The VcBBX protein sequences were submitted to the online program STRING (https://string-db.org/). The orthologous proteins from Arabidopsis with the highest bit scores were chosen to predict the interacting proteins and to construct the protein–protein interaction networks; Arabidopsis thaliana was used as the reference organism.
Analysis organ-specific expression pattern of VcBBX genes
The expression profiles of VcBBX gene from blueberry cultivar ‘Draper’ in different organs including flower bud, flower at anthesis, petal fall, green fruit, pink fruit, ripe fruit, root, shoot, leaf at day and leaf at night, were downloaded from the BioProject database in the NCBI repository (https://www.ncbi.nlm.nih.gov/bioproject/PRJNA494180).
The expression pattern of VcBBX genes under UV-B radiation and co-expression network construction
The expression profiles of VcBBX genes and phytohormone signaling-related pathway genes under UV-B radiation from blueberry calli were downloaded from the BioProject database in the NCBI repository (https://www.ncbi.nlm.nih.gov/bioproject/PRJNA831018). Pearson’s correlation coefficient (r) analysis was performed between differentially expressed VcBBXs and genes in phytohormone signaling-related pathways under UV-B radiation (for 0, 1, 3, 6, 12, or 24 h) according to fragments per kilobase of transcript per million fragments mapped (FPKM) values using SPSS 19.0 software. Genes with a p-value ≤ 0.05 were considered to be significantly correlated. The co-expression networks were visualized based on r value of significant correlation using Cytoscape v3.9.1 software. The heatmaps of blueberry VcBBX genes based on log10 (FPKM) values in different organs and under UV-B radiation were drawn with TBtools software (v1.120) [73].
Stress treatments and reverse-transcription quantitative PCR
Six-month-old blueberry cultivar ‘Northland’ plants from tissue culture were grown in a growth chamber at 25 °C with 70% relative humidity under a 16-h-light/8-h-dark photoperiod. The first to third fully expanded leaves were collected from plants following treatment with salt (200 mM), cold (5 °C), or drought (20% [w/v] polyethylene-glycol) for 6 or 12 h, using samples collected at time 0 (0 h) as control. Total RNA was extracted from the leaves using a Plant RNA Extraction Kit (Sangon Biotech, Shanghai, China). The cDNAs were synthesized using PrimeScript™ RT Master Mix (TaKaRa, Japan). Reverse-transcription quantitative (RT-qPCR) analysis was performed using an ABI 7900HT Real-time PCR system. The relative expression levels of the VcBBXs were calculated using the 2–∆∆Ct method and RT-qPCR results were normalised to 0 h. The glyceraldehyde-3-phosphate dehydrogenase housekeeping gene (GAPDH; GenBank accession no. AY123769) was used as the reference gene. Primer sequences for qPCR are shown in Table S6. All experiments were carried out with three independent biological replicates, and three technical replicates were performed for each biological replicate. One-way ANOVA was used to assess the differences in expression levels of VcBBX genes in plants subjected to the same stress for different periods, and Tukey’s test was used to identify significant differences using SPSS 19.0 software.
Availability of data and materials
All data generated or analysed during this study are included in this article [and its supplementary information files]. The raw sequencing data from this study has been deposited in in the NCBI repository https://www.ncbi. nlm.nih.gov/bioproject/PRJNA892908 for UV-B treatment and https://www.ncbi.nlm.nih.gov/bioproject/PRJNA494180 for organ-specific expression. BioSample: SAMN31399479 (UV-B 0 h), BioSample: SAMN31399480 (UV-B 1 h), SAMN31399481 (UV-B 3 h), SAMN31399482 (UV-B 6 h), SAMN31399483 (UV-B 12 h), and SAMN31399484 (UV-B 24 h) for UV-B treatment; SAMN10438841 (flower_bud), SAMN10438842 (flower_at_anthesis), SAMN10438849 (petal_fall), SAMN10438843 (green_fruit), SAMN10438850 (pink_fruit), SAMN10438851 (ripe_fruit), SAMN10438853 (shoot), SAMN10438852 (root), SAMN10438845 (leaf_night), and SAMN10438844 (leaf_day) for organ-specific expression.
References
Riechmann JL, Heard J, Martin G, Reuber L, Jiang C, Keddie J, et al. Arabidopsis transcription factors: genome-wide comparative analysis among eukaryotes. Science. 2000;290(5499):2105–10.
Kiełbowicz-Matuk A. Involvement of plant C2H2-type zinc finger transcription factors in stress responses. Plant Sci. 2012;185–186:78–85.
Takatsuji H. Zinc-fifinger transcription factors in plants. Cell Mol Life Sci. 1998;54(6):582–96.
Gangappa SN, Botto JF. The BBX family of plant transcription factors. Trends Plant Sci. 2014;19(7):460–70.
Khanna R, Kronmiller B, Maszle DR, Coupland G, Holm M, Mizuno T, et al. The Arabidopsis B-box zinc finger family. Plant Cell. 2009;21(11):3416–20.
Yadav A, Ravindran N, Singh D, Rahul PV, Datta S. Role of Arabidopsis BBX proteins in light signaling. J Plant Biochem Biotechnol. 2020;29(4):623–35.
Crocco CD, Holm M, Yanovsky MJ, Botto JF. Function of B-BOX proteins under shade. Plant Signal Behav. 2011;6(1):101–4.
Cao J, Yuan J, Zhang Y, Chen C, Zhang B, Shi X, et al. Multi-layered roles of BBX proteins in plant growth and development. Stress Biol. 2023;3(1):1.
Heng Y, Jiang Y, Zhao X, Zhou H, Wang X, Deng XW, et al. BBX4, a phyB-interacting and modulated regulator, directly interacts with PIF3 to fine tune red light-mediated photomorphogenesis. Proc Natl Acad Sci USA. 2019;116:26049–56.
Zhang X, Huai J, Shang F, Xu G, Tang W, Jing Y, et al. A PIF1/PIF3-HY5-BBX23 transcription factor cascade affects photomorphogenesis. Plant Physiol. 2017;174:2487–500.
Song Z, Heng Y, Bian Y, Xiao Y, Liu J, Zhao X, et al. BBX11 promotes red light-mediated photomorphogenic development by modulating phyB-PIF4 signaling. Abiotech. 2021;2:117–30.
Zhao XH, Heng YQ, Wang XC, Deng XW, Xu DQ. A positive feedback loop of BBX11-BBX21-HY5 promotes photomorphogenic development in Arabidopsis. Plant Commun. 2020;1:100045.
Job N, Datta S. PIF3/HY5 module regulates BBX11 to suppress protochlorophyllide levels in dark and promote photomorphogenesis in light. New Phytol. 2021;230:190–204.
Bursch K, Toledo-Ortiz G, Pireyre M, Lohr M, Braatz C, Johansson H. Identification of BBX proteins as rate-limiting cofactors of HY5. Nat Plants. 2020;6:921–8.
Gangappa SN, Holm M, Botto JF. Molecular interactions of BBX24 and BBX25 with HYH, HY5 HOMOLOG, to modulate Arabidopsis seedling development. Plant Signal Behav. 2013;8:e25208.
Lin F, Jiang Y, Li J, Yan T, Fan L, Liang J, et al. B-BOX DOMAIN PROTEIN28 negatively regulates photomorphogenesis by repressing the activity of transcription factor HY5 and undergoes COP1-mediated degradation. Plant Cell. 2018;30:2006–19.
Song Z, Yan T, Liu J, Bian Y, Heng Y, Lin F, et al. BBX28/BBX29, HY5 and BBX30/31 form a feedback loop to fine-tune photomorphogenic development. Plant J. 2020;104:377–90.
Holtan HE, Bandong S, Marion CM, Adam L, Tiwari S, Shen Y, et al. BBX32, an Arabidopsis B-Box protein, functions in light signaling by suppressing HY5-Regulated gene expression and interacting with STH2/BBX21. Plant Physiol. 2011;156:2109–23.
Jiang L, Wang Y, Li QF, Bjorn LO, He JX, Li SS. Arabidopsis STO/BBX24 negatively regulates UV-B signaling by interacting with COP1 and repressing HY5 transcriptional activity. Cell Res. 2012;22(6):1046–57.
Fang H, Dong Y, Yue X, Che X, He N, Hu J, et al. MdCOL4 interaction mediates crosstalk between UV-B and high temperature to control fruit coloration in apple. Plant Cell Physiol. 2019;60(5):1055–66.
Liu X, Li R, Dai Y, Chen X, Wang X. Genome-wide identifcation and expression analysis of the B-box gene family in the apple (Malus domestica Borkh.) genome. Mol Genet Genomics. 2018;293(2):303–15.
An JP, Wang XF, Zhang XW, Bi SQ, You CX, Hao YJ. MdBBX22 regulates UV-B-induced anthocyanin biosynthesis through regulating the function of MdHY5 and is targeted by MdBT2 for 26s proteasome-mediated degradation. Plant Biotechnol J. 2019;17(12):2231–3.
Fang H, Dong Y, Yue X, Hu J, Jiang S, Xu H, et al. The B-box zinc finger protein MdBBX20 integrates anthocyanin accumulation in response to ultraviolet radiation and low temperature. Plant Cell Environ. 2019;42(7):2090–104.
Bai S, Saito T, Honda C, Hatsuyama Y, Ito A, Moriguchi T. An apple B-box protein, MdCOL11, is involved in UV-B- and temperature-induced anthocyanin biosynthesis. Planta. 2014;240(5):1051–62.
An J, Wang X, Espley R, Lin-Wang K, Bi S, You C, et al. An apple B-Box protein MdBBX37 modulates anthocyanin biosynthesis and hypocotyl elongation synergistically with MdMYBs and MdHY5. Plant Cell Physiol. 2020;61:130–43.
Zhang ZL, Ji RH, Li HY, Zhao T, Liu J, Lin CT, et al. CONSTANS-LIKE 7 (COL7) is involved in phytochrome B (phyB)-mediated light-quality regulation of auxin homeostasis. Mol Plant. 2014;7:1429–40.
Wang Q, Zeng J, Deng K, Tu X, Zhao X, Tang D, et al. DBB1a, involved in gibberellin homeostasis, functions as a negative regulator of blue light-mediated hypocotyl elongation in Arabidopsis. Planta. 2011;233(1):13–23.
Crocco CD, Locascio A, Escudero CM, Alabadí D, Blázquez MA, Botto JF. The transcriptional regulator BBX24 impairs DELLA activity to promote shade avoidance in Arabidopsis thaliana. Nat Commun. 2015;6:6202.
Fan XY, Sun Y, Cao DM, Bai MY, Luo XM, Yang HJ, et al. BZS1, a B-box protein, promotes photomorphogenesis downstream of both brassinosteroid and light signaling pathways. Mol Plant. 2012;5(3):591–600.
Cao J, Liang Y, Yan T, Wang X, Zhou H, Chen C, et al. The photomorphogenic repressors BBX28 and BBX29 integrate light and brassinosteroid signaling to inhibit seedling development in Arabidopsis. Plant Cell. 2022;34(6):2266–85.
Ravindran N, Ramachandran H, Job N, Yadav A, Vaishak KP, Datta S. B-box protein BBX32 integrates light and brassinosteroid signals to inhibit cotyledon opening. Plant Physiol. 2021;187:446–61.
Bai MJ, Sun JJ, Liu JY, Ren HR, Wang K, Wang YL, Wang CQ, Dehesh K. The B-box protein BBX19 suppresses seed germination via induction of ABI5. Plant J. 2019;99:1192–202.
Liu H, Dong S, Sun D, Liu W, Gu F, Liu Y, et al. CONSTANS-Like 9 (OsCOL9) interacts with receptor for activated C-Kinase 1 (OsRACK1) to regulate blast resistance through salicylic acid and ethylene signaling pathways. PLoS One. 2016;11(11):e0166249.
Xu Y, Zhao X, Aiwaili P, Mu X, Zhao M, Zhao J, et al. A zinc finger protein BBX19 interacts with ABF3 to affect drought tolerance negatively in chrysanthemum. Plant J. 2020;103:1783–95.
Liu YN, Chen H, Ping Q, Zhang ZX, Guan ZY, Fang WM, et al. The heterologous expression of CmBBX22 delays leaf senescence and improves drought tolerance in Arabidopsis. Plant Cell Rep. 2019;38:15–24.
Liu Y, Cheng H, Cheng P, Wang C, Li J, Liu Y, et al. The BBX gene CmBBX22 negatively regulates drought stress tolerance in chrysanthemum. Hortic Res. 2022;9:uhac181.
Yang Y, Ma C, Xu Y, Wei Q, Imtiaz M, Lan H, et al. A zinc finger protein regulates flowering time and abiotic stress tolerance in Chrysanthemum by modulating gibberellin biosynthesis. Plant Cell. 2014;26(5):2038–54.
Li Y, Shi Y, Li M, Fu D, Wu S, Li J, et al. The CRY2-COP1-HY5-BBX7/8 module regulates blue light-dependent cold acclimation in Arabidopsis. Plant Cell. 2021;33:3555–73.
Nagaoka S, Takano T. Salt tolerance-related protein STO binds to a Myb transcription factor homologue and confers salt tolerance in Arabidopsis. J Exp Bot. 2003;54(391):2231–7.
Liu X, Li R, Dai Y, Yuan L, Sun Q, Zhang S, et al. A B-box zinc finger protein, MdBBX10, enhanced salt and drought stresses tolerance in Arabidopsis. Plant Mol Biol. 2019;99:437–47.
Lei L, Cao L, Ding G, Zhou J, Luo Y, Bai L, et al. OsBBX11 on qSTS4 links to salt tolerance at the seeding stage in Oryza sativa L. ssp. Japonica. Front Plant Sci. 2023;14:1139961.
Norberto S, Silva S, Meireles M, Faria A, Pintado M, Calhau C. Blueberry anthocyanins in health promotion: a metabolic overview. J Funct Foods. 2013;5(4):1518–28.
Ribera AE, Reyes-Díaz M, Alberdi M, Zuniga GE, Mora ML. Antioxidant compounds in skin and pulp of fruits change among genotypes and maturity stages in highbush blueberry (Vaccinium corymbosum L.) grown in southern Chile. J Soil Sci Plant Nutr. 2010;10(4):509–36.
Huang J, Zhao X, Weng X, Wang L, Xie W. The rice B-Box zinc finger gene family: genomic identification, characterization, expression profiling and diurnal analysis. PLoS One. 2012;7(10):e48242.
Chun Z, Wang X, Li Y, Yu H, Li J, Lu Y, et al. Genomic organization, phylogenetic and expression analysis of the B-BOX gene family in tomato. Front Plant Sci. 2016;7:1552.
Wei H, Wang P, Chen J, Li C, Wang Y, Yuan Y, et al. Genome-wide identification and analysis of B-BOX gene family in grapevine reveal its potential functions in berry development. BMC Plant Biol. 2020;20(1):72.
Shan B, Bao G, Shi T, Zhai L, Bian S, Li X. Genome-wide identification of BBX gene family and their expression patterns under salt stress in soybean. BMC Genomics. 2022;23(1):820.
Song Y, Ma B, Guo Q, Zhou L, Zhou X, Ming Z, et al. MYB pathways that regulate UV-B-induced anthocyanin biosynthesis in blueberry (Vaccinium corymbosum). Front Plant Sci. 2023;14:1125382.
Bian Y, Ballington J, Raja A, Brouwer C, Reid R, Burke M, et al. Patterns of simple sequence repeats in cultivated blueberries (Vaccinium section Cyanococcus spp.) and their use in revealing genetic diversity and population structure. Mol breeding. 2014;34(2):675–89.
Gupta V, Estrada AD, Blakley I, Reid R, Patel K, Meyer MD, et al. RNA-Seq analysis and annotation of a draft blueberry genome assembly identifies candidate genes involved in fruit ripening, biosynthesis of bioactive compounds, and stage-specific alternative splicing. GigaScience. 2015;4:5.
Colle M, Leisner CP, Wai CM, Ou S, Bird KA, Wang J, et al. Haplotype-phased genome and evolution of phytonutrient pathways of tetraploid blueberry. GigaScience. 2019;8(3):1–15.
Song Y, Ma B, Guo Q, Zhou L, Lv C, Liu X, et al. UV-B induces the expression of flavonoid biosynthetic pathways in blueberry (Vaccinium corymbosum) calli. Front Plant Sci. 2022;13:1079087.
Fang K, Zhang A, Xi W. Genome-wide identification and expression analysis of the B-box gene family in peach fruit during postharvest cold storage and subsequent shelf life. Postharvest Biol Tec. 2021;172:111387.
Shalmani A, Fan S, Jia P, Li GF, Muhammad I, Li YM, et al. Genome identification of B-BOX gene family members in seven Rosaceae species and their expression analysis in response to flower induction in Malus domestica. Molecules. 2018;23:1763.
Cao Y, Han Y, Meng D, Li D, Jiao C, Jin Q, et al. B-BOX genes: genome-wide identification, evolution and their contribution to pollen growth in pear (Pyrus bretschneideri Rehd.). BMC Plant Biol. 2017;17(1):156.
Jin HQ, Xing MG, Cai CM, Li S. B-box proteins in Arachis duranensis: genome-wide characterization and expression profiles analysis. Agronmy-Basel. 2020;10:23.
Wang Y, Zhai Z, Sun Y, Feng C, Peng X, Zhang X, et al. Genome-wide identification of the B-BOX genes that respond to multiple ripening related signals in sweet cherry fruit. Int J Mol Sci. 2021;22:1622.
Song Z, Bian Y, Liu J, Sun Y, Xu D. B-box proteins: Pivotal players in light-mediated development in plants. J Integr Plant Biol. 2020;62:1293–309.
Bai S, Tao R, Tang Y, Yin L, Ma Y, Ni J, et al. BBX16, a B-box protein, positively regulates light-induced anthocyanin accumulation by activating MYB10 in red pear. Plant Biotechnol J. 2019;17:1985–97.
Bai S, Tao R, Yin L, Ni J, Yang Q, Yan X, et al. Two B-box proteins, PpBBX18 and PpBBX21, antagonistically regulate anthocyanin biosynthesis via competitive association with Pyrus pyrifolia ELONGATED HYPOCOTYL 5 in the peel of pear fruit. Plant J. 2019;100:1208–23.
Ulm R, Nagy F. Signalling and gene regulation in response to ultraviolet light. Curr Opin Plant Biol. 2005;8(5):477–82.
Yang G, Zhang C, Dong H, Liu X, Guo H, Tong B, et al. Activation and negative feedback regulation of SlHY5 transcription by the SlBBX20/21-SlHY5 transcription factor module in UV-B signaling. Plant Cell. 2022;34(5):2038–55.
Roro AG, Dukker SAF, Melby TI, Solhaug KA, Torre S, Olesn JE. UV-B-induced inhibition of stem elongation and leaf expansion in pea depends on modulation of gibberellin metabolism and intact gibberellin signalling. J Plant Growth Regul. 2017;36(3):680–90.
Liang T, Shi C, Peng Y, Tan H, Xin P, Yang Y, et al. Brassinosteroid-activated BRI1-MES-SUPPRESSOR 1 inhibits flavonoid biosynthesis and coordinates growth and UV-B stress responses in plants. Plant Cell. 2020;32(10):3224–39.
Hectors K, Oevelen SV, Guisez Y, Prinsen E, Jansen MAK. The phytohormone auxin is a component of the regulatory system that controls UV-mediated accumulation of flavonoids and UV-induced morphogenesis. Physiol Plantarum. 2012;145:594–603.
Lyu G, Li D, Li S. Bioinformatics analysis of BBX family genes and its response to UV-B in Arabidopsis thaliana. Plant Signal Behav. 2020;15(9):1782647.
Heng Y, Lin F, Jiang Y, Ding M, Yan T, Lan H, et al. B-box containing proteins BBX30 and BBX31, acting downstream of HY5, negatively regulate photomorphogenesis in Arabidopsis. Plant Physiol. 2019;180(1):497–508.
Yadav A, Lingwan M, Yadukrishnan P, Masakapalli SK, Datta S. BBX31 promotes hypocotyl growth, primary root elongation and UV-B tolerance in Arabidopsis. Plant Signal Behav. 2019;14(5):e1588672.
Vaishak KP, Yadukrishnan P, Bakshi S, Kushwaha AK, Ramachandran H, Job N, et al. The B-box bridge between light and hormones in plants. J Photochem Photobiol B. 2019;191:164–74.
Huang Y, Xiong H, Xie Y, Lyu S, Miao T, Li T, et al. BBX24 interacts with DELLA to regulate UV-B-induced photomorphogenesis in Arabidopsis thaliana. Int J Mol Sci. 2022;23(13):7386.
Soitamo AJ, Piippo M, Allahverdiyeva Y, Battchikova N, Aro EM. Light has a specific role in modulating Arabidopsis gene expression at low temperature. BMC Plant Biol. 2008;8(1):13.
Kumar S, Stecher G, Li M, Knyaz C, Tamura K. MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol Biol Evol. 2018;35(6):1547–9.
Chen C, Chen H, Zhang Y, Thomas HR, Frank MH, He Y, et al. TBtools: an integrative toolkit developed for interactive analyses of big biological data. Mol Plant. 2020;13(8):1194–202.
Acknowledgements
Not applicable.
Funding
The work was supported by the National Natural Science Foundation of China (Grant no. 31700260).
Author information
Authors and Affiliations
Contributions
C.Z. designed and carried out the experiments; X.L., W.S. and B.M. carried out all bioinformatics analysis and wrote the manuscript; Q.G., L.Z. and X.Z supervised the experiments; Y.S. and K.W. contributed to genes expression analysis. C.Z. directed and revised the manuscript. All authors read, reviewed and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1:
Table S1. Homologous Arabidopsis and blueberry genes of the BBX family. Table S2. Gene ID of VcBBXs in blueberry and AtBBXs in Arobidopsis. Table S3. The acronyms and corresponding full name for predicted VcBBX interactors. Table S4. Information of BBX genes under UV-B radiation. Table S5. Pearson's correlation coefficients (r) of expression levels between B-Box and plant hormone signlaling genes under UV-B radiation. Table S6. Primers used in this study.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Liu, X., Sun, W., Ma, B. et al. Genome-wide analysis of blueberry B-box family genes and identification of members activated by abiotic stress. BMC Genomics 24, 584 (2023). https://doi.org/10.1186/s12864-023-09704-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12864-023-09704-8