
Abstract
Background
The host specificity and host range of the dry bean beetle, Acanthoscelides obtectus (Coleoptera: Chrysomelidae, Bruchinae), a seed predator of beans, is poorly known. In addition, the female oviposition preference and larval performance relationship is complicated by the respective importance of seed coat and cotyledon, because, paradoxically, females lay eggs on the basis of stimuli of the seed coat alone, without directly being able to assess the quality of the cotyledon’s suitability for larval development. Conversely, the thickness of seed coat may prevent first instar larvae from entering the seeds, even if cotyledons are suitable for development.
Methods
The seeds of 62 leguminous species and 75 cultivars and accessions occurring in Hungary were evaluated for preference-performance relations. The preference of female bean beetles for seeds was measured in no-choice egg-laying tests. The ability of first instar larvae to overcome the seed coat as a physical barrier was tested with intact seed coat, while pre-drilled seed coats allowed the larvae to assess the suitability of cotyledon for development. The number of emerging adults was recorded. The thickness of seed coats and the weight of seeds were measured. Nonparametric tests and logistic regression were used for the statistical analyses of data and effect sizes were also calculated.
Results
Seeds of 18 leguminous species (35% of them Lathyrus) supported larval development to adults if the seed coat was pre-drilled; however, only nine leguminous species supported development to adults if the seed coat was intact. Seed coat thickness beyond a critical threshold of 0.1 mm strongly influenced survival of first instar larvae. There was no overall positive correlation between oviposition preference and larval performance, except for 16 so-called acceptable non-hosts (Kendall’s τ = 0.3088). A. obtectus females also showed an ovipositional hierarchy of legume species even in no-choice tests.
Conclusions
The results suggest that whereas the use of some acceptable non-host species by the A. obtectus is possible in seed stores, the same is unlikely under outdoor conditions, where the recognition of a diverse set of seed pod-related compounds would be necessary to induce egg-laying.
Similar content being viewed by others

Background
In a seminal paper, Janzen [1] discussed the importance of seed coats and cotyledons of tropical legume species in their interaction with a bruchid beetle, Callosobruchus maculatus (F.). Based on no-choice oviposition and larval development investigations in the laboratory, he found extreme host selectivity of the species, which he attributed to the diversity of chemical compounds present in the hosts, and to physical traits, such as thickness, hardness and smoothness of the seed coat. However, more than forty years after Janzen’s paper, the host specificity of bruchids is still insufficiently known.
Although only implicitly, Janzen [1] also became one of the pioneers of female preference-larval performance studies, asking whether insect mothers optimally select substrates for larval development, and whether oviposition on novel or non-hosts are attempts of host range expansions [2, 3]. The latter hypothesis is supported by the fact that the range of plant species promoting survival of larvae is often wider than the egg-laying preference of the mother [4, 5]. The preference-performance theory postulates that females select egg-laying substrates in order to maximize their progeny’s fitness [3, 6, and references therein]. The female’s selection behaviour in a choice situation, where several items are presented simultaneously, and the female’s encounters with substrates of different quality is a sequential process, usually fits to the hierarchy of the genetically-based template of hosts [7]. Therefore, a positive correlation between the mother’s preference and offspring performance (usually the yield of adult offspring from a particular substrate) is probable. A meta-analysis of such studies [8], however, concluded that the results were contradictory: positive and negative correlations were equally abundant. Moreover, among the 23 studies of oviposition preference, there were only four no-choice tests. Females often make oviposition “mistakes”, e.g., laying eggs on toxic or otherwise unsuitable substrates; however, it is possible that these merely reflect the neural limitations of insects [9]. The strength of the relationship seems dichotomous according to the degree of host specialization of herbivorous insects. Specialists show strong positive preference-performance relationships, whereas in generalists, it may not exist at all [8, 10]. Several studies did not find genetic correlations between adult preference and larval performance either [11, 12]. The negative relationships refer not only to the lack of adaptation due to metabolic trade-offs, but to the importance of hitherto unconsidered life history components and environmental factors [8, 13, 14].
The preference-performance relationship arises in a different version for seed predator bruchid species. The complicating factor is the seed coat (testa, seed integument) that may or may not provide information on the quality of cotyledon for the egg-laying females or first instar larvae, but at the same time can impose formidable physical and chemical barriers for the larvae to enter the seed. The information on which the female bases her decision to oviposit may not adequately refer to the suitability of cotyledon, and the larva, following an exhausting period of drilling through a tough testa, may still die soon after contacting it. Despite the enormous advances in chemical analysis and the wealth of information of seed chemical composition in Leguminosae [15], relatively little is known of how seed coat chemistry affects host selection by bruchids.
Preference-performance studies evaluating a wide range of hosts and non-hosts are lacking due to the usually narrow host specialization of bruchids [1, 16]. Nevertheless, the bruchid species, Acanthoscelides obtectus (Say), Callosobruchus spp. and Zabrotes subfasciatus (Boheman) (Coleoptera, Chrysomelidae, Bruchinae), that are capable of propagating in storage on leguminous seeds, provide an opportunity to study host specificity. On a limited number of hosts, negative genetic correlations were found between preference and performance components in C. maculatus [17]. Furthermore, A. obtectus larvae reared on chickpea (Cicer arietinum L.) for ≥ 50 generations had lower survival rates in comparison to those reared on the natural host, Phaseolus vulgaris L. [18, 19]. It was concluded that there was a fitness cost for using an unusual host, and that whereas no genetic correlation was detected between preference and performance on Phaseolus, it existed on C. arietinum.
This paper investigates the potential host range and tests the preference-performance relations of A. obtectus. This species propagates on two species of cultivated beans (P. vulgaris and P. coccineus L.) in the temperate zone. In Europe, it can continuously reproduce on stored dry beans and therefore can achieve a pest status on this commodity. It also seasonally infests beans grown outdoors, laying eggs on nearly mature seeds within pods [20]. At harvest, infested seeds are collected, and the cycle is repeated in bean stores. Females do not stick eggs to the seed surface but place them among the seeds. First instar larvae have legs and can move among seeds to select sites on seeds to enter, and they can exercise independent selection behaviour, if a choice of seeds is available. While drilling through the seed coat, they avoid ingesting most of it [21], producing fine powder that indicates the site of boring. This behaviour is also observed with other bruchid species [1, 22, 23] and suggests the presence of potential chemical factors in the seed coat. Larvae tunnel into the cotyledon, develop through four instars and pupate within the seed.
The tribe Phaseoleae (Leguminosae) comprises ca. 89 genera, including Glycine, Lablab, Vigna and Phaseolus [24]. A. obtectus is an oligophagous species primarily attacking beans within the Phaseolus genus. However, spontaneous occurrence in some cultivars of the above three genera are also known, but it is a rare event and poorly documented, along with occasional observations concerning infestations in other legume genera such as Lupinus (tribe Genisteae), Cicer (Cicereae), Vicia, Lens and Lathyrus (Fabeae) [5, 25,26,27,28]. In seed storage facilities, only dry seeds are available, so selection of seeds for oviposition is dependent on the number of species being stored. Therefore, while selection choices for females can be limited, her oviposition decisions may be important for offspring survival.
With respect to A. obtectus there are several areas of research that have yet to be addressed, namely, to confirm host range, oviposition decisions under a no-choice environment and subsequent larval performance and survival. Females rank seeds by size [29], but are they able to judge their suitability for larval development based on information provided solely by the seed coat? The following hypotheses were also tested: (1) seed coat thickness determines penetration by first instar larvae, (2) ingestion of seed coat is toxic to the larvae, and (3) in the absence of the seed coat, the suitability of the cotyledon determines survival of larvae.
Materials and methods
The experimental insect
The test beetles originated from a continuous laboratory rearing maintained on commercial P. vulgaris beans for at least five years at 26–28 °C and 18/6-light/dark regime as described by Szentesi [30]. Following emergence, several hundred adults were placed into 10 × 15 cm size jars on corrugated paper, fed honey-water and allowed to mate. Adults are sexually mature at emergence from beans, and females can lay eggs by the third day. One-to-three-day old males and females were separated based on the shape and coloration of the pygidium and used for the experiments. To obtain eggs for the experiments, several hundred beetles were placed into an 18 cm high and 20 cm diameter glass cylinder, with the bottom opening capped with 1 × 1 mm mesh metal screen. The weevils laid eggs on a single layer of dry beans placed above the screen, and the eggs fell into a lower collecting dish when the cylinder was shaken (see more details in Szentesi [30]). As new beetles were put into the device daily, a continuous supply of eggs was provided. Daily collection of eggs allowed precise timing of emergence of the “black-headed” stage of first instar larvae. At this stage of development, the eggshell becomes transparent, and the head of the larva turns black in the egg, indicating the commencement of hatching of larvae within several hours, i.e., within 24 h at 26–28 °C.
Plant species used in the tests
In this study, 62 species from the Leguminosae family belonging to the tribes Cercideae, Caesalpinieae, Genisteae, Amorpheae, Phaseoleae, Robinieae, Galegeae, Cicereae, Trifolieae and Fabeae, and 75 plant selections of six legume and four non-legume species (the latter as an outgroup) were tested for egg-laying and larval development (see Additional file 1: Tables S1, S2 and S3 for names of species, cultivars and accessions). These species either occur naturally in Hungary, are naturalized, or are cultivated as food or fodder. Only species with seeds which had masses large enough to support development of an A. obtectus larva were selected [31]. Seed mass was determined at room temperature with the help of an electronic balance (Sartorius A210P, Germany) operating with 0.1 mg accuracy.
As to developmental suitability of the plant species de Boer and Hanson’s [32] classification was used. Hosts (H) are those plant species that fully support development through generations and regularly harbour natural infestations. There are only two such species, the common bean (P. vulgaris) and the scarlet runner bean (P. coccineus) that fulfil these conditions in Hungary. Acceptable non-hosts (ANH) are not recognised and used as suitable substrates for larval development in nature. They grow usually asynchronously with bruchid phenology in space and time, and thus bruchid females are only occasionally constrained to use them, e.g., in seed stores. Although some adults may emerge, developmental time is typically long and larval mortality high. Chickpea is an example that belongs to this group. The third group, non-hosts (NH), comprises of plants that are nutritionally inadequate or toxic, and never supports development, although occasional egg-laying cannot be excluded in seed stores. The seed coat, the primary barrier to the cotyledon, was defined as penetrable if a first instar larva of A. obtectus was able to bore through it in case of an intact seed, and the cotyledon was considered suitable for development if an adult emerged from a seed regardless of whether the seed coat was intact or pre-drilled.
The seed samples originated from field or cultivated collections, and some were obtained from plant breeding companies. The samples were stored at a dry, cool place until use. Plant identification was carried out by the author.
Egg-laying tests
Because no-choice egg-laying tests were carried out, “preference” is not used in the sense of choice tests, where selection by an insect is based on free movement among items offered, but instead refers to a position in the hierarchy of host-range. The measure of such preference for a plant species was the number of eggs laid/female on seeds. For the no-choice tests, the seeds of plant species were kept at 26–28 °C and 70–80% RH to allow them to take up moisture a week before the bioassay commenced. Thereafter, three seeds of the same plant line were placed into a 2 cm × 5 cm glass vial, and three 1–2-day old female and male adults introduced into each vial. A piece of white linen was fixed by a rubber band onto the vial to prevent adults from escaping. The vials were placed in a controlled environment chamber in darkness (L:D/0:24 h), and maintained until all weevils had died, and then number of eggs laid over the entire lifetime was counted. There were seven replicates for each plant species and selections.
Larval performance test
Success of larval development on the different hosts was assessed by the yield of adult offspring. Seeds of all legume species and plant selections were handled as described in the previous section.
Two parallel treatments, with 45 replicates each, were set up. In one treatment the seeds were left intact, in the other they were pre-drilled with a high-speed electric drill (Triplex Miniplex, France) bit 0.14 mm in diameter, corresponding to the diameter of the first instar A. obtectus larva. One or two holes per seed were made at the ends and in the middle, respectively (depending on the size of seed) under a binocular microscope. Efforts were taken to drill through the seed testa only. Seeds were placed individually into glass vials (1 × 6 or 2 × 8 cm diam. × length) corresponding to seed size, and a single “black-headed” A. obtectus egg was placed onto the inner wall of the vial, ca. 1–2 cm above the seed, with the help of a fine wet brush, then the vial was capped with a cotton stopper. The vials were placed at 26–28 °C and 60–80% RH in dark conditions. To calculate the duration of time needed for development, the time of egg hatching and the L1s’ boring into the seeds were recorded by daily inspection from the beginning of the experiment. The observation of adult emergence was started after ca. 30 days. At least four months were allowed for development, then the seeds were opened under a dissecting microscope and the dead life stages recorded.
After establishing egg mortality, the remaining number of larval instars, such as L1 dead outside beans, plus dead L1, L2, etc. and pupae inside seeds, were taken as 100%; this value was then divided up among instars recorded. The instar of each larva was verified by head capsule measurement. All developmental malformations were also recorded. During dissection of the seeds, seed coat thickness was measured on ≥ 10 randomly selected seeds, using a micrometre under a microscope.
Effect of bean seed coat on larval development
To test the hypothesis that the bean seed coat is not only a physical barrier against entering the cotyledon but also contains chemicals detrimental to the L1s, artificial beans (balls) were prepared. The ca. 150 mg size balls were prepared from finely pulverized cotyledons (seed coat removed) of P. vulgaris cv. Valja, and seed coat powder of the same cultivar was added in 2.5, 5 and 10% w/w concentrations. The seed mass of this bean variety is 240.7 ± 5.4 mg (mean ± SE, N = 35), and the seed coat is < 10% of the mass. The cotyledon was milled by a water-cooled grinder (Tekmar A-10, IKA, Germany) to avoid chemical deterioration from heat. The balls consisted of a standard amount of bean cotyledon powder (80%) mixed with 20% water-soluble potato starch. These balls served as controls. Adequate portions of the potato starch were substituted with seed coat to create the above concentrations, e.g., in case of balls containing 10% seed coat powder, there were 80% cotyledon and 10% starch powders. Addition of distilled water created a pastry from which 6 mm diam. balls were made by hand, and they were dried at 40 °C for a day. The acceptability of the artificial beans for egg-laying was tested with three males and three females and three balls for each treatment. To assess the balls’ suitability for larval development, one egg with a “black-headed” larva was placed on individual balls in vials in 13 respective replicates/treatment. The number of eggs laid, the percentage dead larval stages without or after boring into the balls, as well as that of emerging adults were recorded.
Statistical procedures
Variance homogeneity of variables (larval mortality outside and inside seeds, and adult emergence from intact or pre-drilled seeds, etc.), notwithstanding various transformations, did not meet conditions of parametric tests, therefore nonparametric tests were used. Between-group comparisons were performed for three variables (e.g., plant groups) with Kruskal–Wallis ANOVA, and for two variables (e.g., intact and pre-drilled seed coat) with Wald-Wolfowitz runs test. The results are presented as medians and quartiles if data were obtained in percentages.
As a nonparametric version of discriminant function analysis, logistic regressions of the generalized linear models [33] were applied to reject or accept two major null hypotheses: (a) seed coat thickness does not influence L1 mortality, and (b) the suitability of seed cotyledon does not affect larval development. The continuous dependent variables indicated dichotomous distributions, therefore they were transformed to binary categorical variables at biologically reasonable cutpoints, and the coding of variables was performed accordingly: independent variables (xi), such as seed coat thickness, was assigned 1 if < 0.1 mm, and 0 if > 0.1 mm; cotyledon supporting larval development and adult emergence was given 1 if ‘suitable’, and 0 if ‘unsuitable’. Dependent variables (yi), such as L1 mortality outside seeds (N = 140), successful penetration of the seed coat (N = 140), larval development inside seeds (N = 212), and adult emergence (N = 212) were coded 1 if L1 mortality was < 50%, if L1 entered the seeds, if larval mortality inside seeds was < 30%, and if adult emergence from seeds was > 10%. Zero codes were assigned to the opposite outcomes (Additional file 1: Tables S6 and S7). Such zero–one “reference cell” coding [34] produced χ2-tables, where the left upper cells (A) were 11, and the right lower ones (D) were 00.
For logistic regression, STATISTICA ver. 6’s [35] GLZ module with binomial distribution, logit link function and sigma-restricted parameter estimation was used. The respective analyses used only a single predictor variable and one response variable. The program provided the estimate of the categorical predictor that maximized the probability of the dependent variable. For goodness of fit, Wald- and χ2-statistics of log-likelihood were calculated.
With the dichotomous data, effect sizes (risk difference, risk ratio, odds ratio and confidence intervals, CI) for L1 mortality and adult emergence were estimated by the methods described in [36, 37]. The calculations were based on 2 × 2 χ2-tables (Additional file 1: Tables S6 and S7). The odds ratio (OR) of the outcome was computed by the probabilities of cells: [p(1)/(1-p(1)]/[p(0)/(1-p(0)], where in p(1) the binary independent predictor is 1, and in p(0) it is 0. In order to be able to calculate effect size for adult emergence, 0.5 was added to all values in cells, because one element (adult emergence from NH plants) of the frequency table was zero. Interactions between dichotomous variables (e.g., ‘penetrable’/’impenetrable’ and intact/pre-drilled seed coat) were computed by multiplying two odds ratios (OR10 × OR01), assuming H0 (that there is no interaction) if the odds ratio cell OR11 equalled the result [38]. One-way ANOVA was used to evaluate the effects of concentrations of seed coat powder on larval mortality. All statistical procedures were performed with STATISTICA 6 [35].
Results
Female oviposition preference
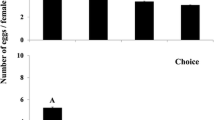
Bruchid females deposited significantly decreasing number of eggs/female on seeds in the order of host (H), acceptable non-host (ANH) and non-host (NH) species groups. Females laid a mean (± SE) of 38.6 ± 0.8 eggs on 22 H species and plant selections, 18.6 ± 0.7 eggs on 55 ANH species and plant selections, and 13.5 ± 0.5 eggs on 63 NH species and plant selections, and significantly more eggs on larger seeds (Table 1). Mean mass of H seeds was 340.8 mg, that of ANH 235.6 mg, and that of NH 70.3 mg (Table 1). All 66 plant species (62 leguminous and 4 outgroup species) received eggs, but females laid less than 15 eggs on 55%, less than 30 eggs on 35%, and between 30–45 eggs on 10% of plant species. Figure 1 shows the distribution of eggs among the leguminous tribes. Not surprisingly, A. obtectus females laid the highest number of eggs on members of the tribe Phaseoleae, where the main hosts are also found. Within Phaseoleae, soybeans (Glycine) were the least preferred (Additional file 1: Table S1). Comparable responses were noted to some species within the Caesalpinieae, Genisteae, Robinieae, Cicereae and Fabeae (Additional file 1: Table S1). Relatively high numbers of eggs were laid on some non-host species: Gleditsia delavayi Franch. (Caesalpinieae), Laburnum alpinum (Mill.) Bercht. & J. Presl, L. anagyroides Medik. (Genisteae), and Robinia viscosa Vent. (Robinieae). C. arietinum (Cicereae) and Vicia faba L. (Fabeae) are known as occasional hosts. The number of eggs laid/female on them fell into the medium and high categories, respectively. A mean number of less than 10 eggs/female were laid on Vicia tenuifolia Roth, Robinia pseudoacacia L. and Amorpha fruticosa L., and on some other members of the Fabeae tribe (Additional file 1: Table S1).

Distribution of the number of eggs laid/female A. obtectus across leguminous tribes. Data are means (± SE), points represent outlier values. The order of tribes corresponds to the phylogenetic relations presented by [24]
Females accepted the artificial seeds incorporated with seed coat as an oviposition substrate and laid similar number of eggs as on the control (Table 2).
Larval performance
The major criteria of larval performance were: (a) L1 mortality outside seeds due to the thickness of seed coat, and (b) within-seed mortality of various developmental stages in the cotyledon. Both variables were modulated by the intact or pre-drilled status of seeds. The seed coat of host (H) and acceptable non-host (ANH) species was significantly thinner (Table 1). L1 mortality outside intact seeds was significantly lower for the H (24.75%) and ANH groups (93.3%), in comparison with the NH group (100%, all are medians, Table 1). For the two H and 16 ANH species, Table 3 provides data of seed coat thickness, whereas Additional file 1: Tables S2 and S3 give similar information for NH, and plant selections of H and ANH groups. Remarkably, seed coat constitutes a barrier even on the primary host, beans (P. vulgaris). Contrary to the very low larval mortality inside seeds, there was substantial mortality outside intact seeds of bean cultivars (27.3%, 13.6–53.3, median, lower and upper quartiles, N = 21), but only 4.4% (0–11.4) for pre-drilled seeds. Similar values can be given for other legume species from which adults emerged, with the difference that the upper level of mortality usually reached 100% with intact seeds, with some exceptions such as Vigna unguiculata (L.) Walp. and V. angularis (Willd.) Ohwi & H. Ohashi, where the seed coat was extremely thin (Tables 3 and 4). The best survival was seen in runner beans (P. coccineus), with 0% (intact seed) and 2.2% (pre-drilled seed) larval mortality outside the seeds, respectively.
Mortality of various developmental stages inside intact seeds were substantially different from that within pre-drilled ones; however, the critical event for development in the seeds was invariably the survival of first instar larvae. Tables 1 and 4 provide results for H and ANH species, and further data are available in Additional file 1: Tables S2 and S3 for bean, pea and soybean varieties, as well as for NH species, showing differences among host types. Because larvae were unable to penetrate the intact seed coat of NH species, there was no larval establishment inside NH seeds. In comparison, larval establishment was 75% (44–93%, median and quartiles) within pre-drilled seeds. It was observed with all plant groups that many larvae entered seeds via the pre-drilled hole but then exited and died. Seed testa frequently had several shallow pits, where L1 attempted to bore in (e.g., all Gleditsia japonica Miq. seeds, both intact and pre-drilled, had such traces). Although it is well documented that additional A. obtectus larvae may enter through the hole made by a pioneer larva [39], the ratio of L1-made and artificial holes was 1.5:1 on the most preferred bean seeds (P. coccineus), i.e., many L1 larvae did not use the pre-prepared holes on this host. In some instances (e.g., Caragana or Onobrychis genera), L1s entered seeds through the hilum. First instar larvae entering Gleditsia seeds through an artificial hole made at the embryo area invariably died within the embryo. In cases where the cotyledons were soft (several Glycine cultivars/accessions and Caragana), larvae made longer tunnels before dying.
The experiment that aimed to elucidate the effects of seed coat of beans on larval development proved that, besides being a physical barrier, the seed coat also inhibited larval development at the lowest concentration incorporated into artificial seeds (Table 2). Conversely, controls (without any seed coat content) were fully suitable for development.
Significantly more adults emerged from H species and plant selections than either from ANH species and plant selections or NH species, and whether intact or pre-drilled (Table 1). Of the 62 legume species, A. obtectus larvae developed into adults in 18 (29%) species in four tribes, if the seed coat was pre-drilled, compared to only nine species if they had intact seed coat (Table 4). However, the picture varied considerably concerning plant selections (Additional file 1: Table S3). Although adult emergence in P. vulgaris was generally high, at the cultivar level it ranged between 51 and 100%. In P. coccineus, all larvae developed into adults. Interestingly, even intact cowpea (Vigna) seeds supported larval development to adults, whereas in white lupin (Lupinus albus), adults emerged from pre-drilled seeds only. Pre-drilled soybean (Glycine) plant selections and hyacinth bean (Lablab) seeds assisted higher survival to adults than intact seeds. This is also paralleled by the length of developmental time needed until adult emergence (Table 4); for instance, it was two to three times longer in G. max (L.) Merr., V. angularis and P. sativum L. in comparison with beans. Of the 27 plant selections of garden peas, adults emerged from 24 (88.9%), however, only from 13 (48.1%) of these if the testa was intact. Similar values occurred for 17 G. max plant selections: adult emerged from six (35.3%) of them, but only from four of these with an intact seed coat. A surprising feature is the asymmetric distribution of adult emergence between the Vicia and Lathyrus genera.
Of the 18 leguminous species that had pre-drilled seed testa and were suitable for survival through to adults, in five (28%) species some adults that emerged showed malformation. Furthermore, in 10 leguminous species, larvae developed through to adults but were unable to emerge. Typical malformation was a substantial decrease in elytra width and length: the elytra became shorter and triangular. There were 0.1% malformed adults in beans, 33.0% in peas, 31.6% in L. tuberosus L., 6.7% in L. sativus, and 0.7% in V. faba.
Female preference versus larval performance
Additional file 1: Table S5 provides the most important nonparametric correlation coefficients referring to the overall relationship between plant traits and insect responses. Accordingly, there was a significant positive relationship between preference and performance only in the ANH group, i.e., between the number of eggs laid/female and adult emergence (Kendall’s τ = 0.3088, intact seeds, N = 55). The correlations between seed mass and the number of eggs laid were extremely low in all three plant groups but were significant in ANH and NH species. The thicker the seed coat, the higher was the first instar larval mortality outside on ANH and NH seeds, and seed coat thickness also affected L1 larval mortality on H seeds too.
The logistic regression provided evidence that L1 mortality outside seeds was due to different seed coat thicknesses. With the condition of less than 50% L1 mortality outside seeds, 15.6% (7.1–32.6, median and quartiles, N = 19) mortality occurred if the seed coat was thin (0.08 ± 0.001 mm, mean ± SE, N = 78), in comparison with 100% (97.8–100, median and quartiles, N = 61), if the seed coat was thick (0.15 ± 0.005 mm, N = 62) and with mortality higher than 50%. (Wald test for seed coat thickness was 8.2, df = 1, p = 0.0043; log-likelihood: -48.4; goodness of fit χ2: 17.9, df = 1, p < 0.001.) Approaching the same hypothesis from another angle, i.e., if the seed coat was assigned as’penetrable’ or ‘impenetrable’, significantly higher number of L1 entered seeds with ‘penetrable’ seed coat (Wald test: 27.3, df = 1, p < 0.001; log-likelihood: -81.4; χ2 = 31.1, df = 1, p < 0.001). Here, only those cases were considered where L1 larvae entered a seed then died immediately after it. This result is interesting, because ‘penetrable’ seed coat thickness was 0.0998 ± 0.004 mm (N = 64), whereas ‘impenetrable’ was 0.119 ± 0.005 mm (N = 76), a mere 0.02 mm difference.
Larval performance and adult emergence did depend on the ‘quality’ or ‘suitability’ of seed cotyledons. With the criterion of less than 30% larval mortality inside seeds, mortality was 2.27% (0–7.33, median and quartiles, N = 87) if the cotyledon was ‘suitable’ for larval development, in comparison with ‘unsuitable’ cotyledons (80.0%, 60–95.5, median and quartiles, N = 60), and with mortality higher than 30%. (Wald test for cotyledon suitability was 27.6, df = 1, p < 0.001; log-likelihood = -131.5; χ2 = 29.8, df = 1, p < 0.001.) Adult emergence from ‘suitable’ seeds was 37.8% (10–84.1, median and quartiles, N = 123) vs. from’unsuitable’ cotyledons: 0% (N = 89), thus logistic regression for adult emergence could not be performed. Nevertheless, effect sizes could be computed from the χ2 table.
Effect sizes and risk analyses showed that L1 larvae had 19 times larger chance to have < 50% mortality outside seeds by odds ratio (OR). Risk difference (RD) indicated 45% higher survival for larvae if they happened to bore in a seed with seed coat thickness of < 0.1 mm, in comparison with seeds having thicker testa (Additional file 1: Table S6). However, the regression coefficient (φ2) explained only 10% of variance of seed coat effect, referring to other important factors affecting larval entry. On the other hand, when facing penetrable/impenetrable seed coat (results are not shown in table), L1 larvae had three times higher risk to die facing ‘impenetrable’ seed coat (RR = 3.01 ± 1.3, mean ± SE, CI95 = 1.9 & 4.8). However, chances for larvae to enter a seed with penetrable testa was 7.5 times higher (OR = 7.5 ± 1.5, CI95 = 3.5 & 16.1), even if they died after the first bites from the cotyledon. Here, the regression coefficient explained a relatively high level (21%) of the variance.
The risk of > 30% larval mortality inside seeds increased ca. 2 times (RR = 1.95 ± 1.13, CI95 = 1.52 & 2.49) in cotyledons unsuitable for reaching later developmental stages. The odds for such an outcome were high (OR = 4.85 ± 1.28, CI95 = 2.96 & 7.94, results are not shown in table). The chance for reaching adulthood in seeds from which > 10% adults emerged was ca. 4 times higher (RR in Additional file 1: Table S7), than in cotyledons allowing only 1–2 weevils to successfully complete their development. The odds for adulthood in suitable seeds were extremely high (OR = 523.42 ± 1.23, CI95 = 348.74 & 785.60) due to the asymmetry caused by the NH seeds. The regression coefficient (φ2) explained a high portion (54%) of the variance (Additional file 1: Table S7).
As expected, there was an interaction between penetrable/impenetrable and intact/pre-drilled states of seed coats: the joint effect (OR11 = 0.6718) was larger than the multiplied value (0.4820) of their respective effects (OR10 = 0.7179 and OR01 = 0.6714). The joint effect of both variables is 1.4 times higher than the combined effect of each variable acting separately.
Discussion
This study could not prove an overall positive correspondence between host preference and host suitability. However, 16 leguminous plant species (Table 4) did show positive significant relationship between oviposition preference and larval performance (Additional file 1: Table S5), and these are called acceptable non-hosts (ANH) [32]. Nine of these legume species supported development into adult stage even if the seed coats were intact. Besides the two Phaseolus species, rearing in laboratories is possible on chickpea through generations [18, 40], and occasional infestations occur in stores on garden peas or faba beans (Szentesi unpubl. results). How females assess host suitability of ANH species through the seed coat remains an intriguing aspect of the positive relationship between female choice and larval success. The distribution of larval mortality and adult emergence (Tables 1 and 4, Additional file 1: Tables S2 and S3) among plant groups, and the results of logistic regression (Additional file 1: Table S6), overwhelmingly emphasize the importance of the seed coat in allowing or preventing the use of a seed. For instance, regression coefficients (φ2) explained 10% and 21% of the variance, respectively, of the effect of seed coat thickness and ‘penetrability’, supporting the hypothesis that several yet unknown physical and chemical factors are also important. In spite of the fact that a preference for larger H seeds has been proven in bruchids in choice-tests [29, 41], the seed mass of plant groups had negligible effects on the number of eggs laid/female (Additional file 1: Table S5), and the significant positive preference relationship in ANH and NH species might be attributable to physical traits, e.g., curvature of the seed surface.
In the present study only a limited number of possible variables that may affect preference-performance relationships (seed mass, number of eggs laid/female and seed coat thickness) was taken into consideration. Other important factors (e.g., chemicals, further physical, environmental, life history traits, mobility, predators, host genotype) that influence host selection [8, 10, 42] were not investigated. In addition, experience may promote acceptance of ANH species [43, Szentesi unpubl. results] and may even lead to host-race formation [44], but neurological constraints can impose limitations to perception and integration of stimuli [9]. In the light of many factors influencing preference-performance relationship, and specifically egg-laying, it is doubtful whether the oviposition response of A. obtectus females on ANH and NH species was “erroneous” or “defective”, as there are possible alternative explanations for the behaviour [45, 46].
Is there a hierarchy in host selection in no-choice tests?
The widely known hierarchy-threshold model [7] for individual host selection in choice situations explains that specificity is bound to genetic and physiological status, and that the rank-order of hosts is invariable. Oviposition occurs when acceptability of a substrate exceeds the motivational threshold, and the different acceptability levels create a rank-order. An alternative hypothesis [47] states that it is the actual set of eggs available for laying that decides acceptance: higher number of eggs motivate acceptance of low-quality hosts. In this study, however, no-choice tests were applied. A. obtectus females did produce a rank-order of ANH plants; however, except for the primary host beans, this was different from those obtainable in choice tests, and even more different concerning larval development (Szentesi unpubl. results, Additional file 1: Table S8). In choice tests, A. obtectus visits items in a sequential way and lays most eggs on beans. However, it never totally ignores other substrates (even glass beads), because variation in egg-laying is also modulated by factors such as egg-load and/or, in most cases, unknown physical and chemical stimuli. In this study females laid ca. 1/3 of the number of eggs on NH species compared to those deposited on H plants. This means that elements of acceptability of different NH plants could induce the motivational state to accept a lowered egg-laying threshold as time passed.
The host-range of A. obtectus
Pest status, width of geographic distribution and host range can be interconnected [48]. A. obtectus is narrowly specialized on some wild and cultivated members of the genus Phaseolus [49], but it is a widespread and important pest, whose host affiliation may also include some species of pantropical leguminous tribes [50]. In the present study, P. vulgaris and P. coccineus represented the hosts, the second providing better preference/performance ratio as also confirmed by [25]. Some species and cultivars of the following genera belong to the ANH group: Lupinus, Glycine, Lablab, Vigna, Cicer, Vicia, Lens, Lathyrus, and Pisum (Table 4), largely agreeing with findings of [5, 16, 26, 27]. Differences depend on whether the authors applied a sufficiently wide spectrum of plant selections, because differences in acceptance among these can be as large as, or larger than, between species (Additional file 1: Tables S2 and S3).
Pre-drilled seeds contributed to the understanding of host-range by demonstrating how important chemical constituents of the cotyledons were when seed coats did not function as barriers. The surprisingly high ratio of Lathyrus species supporting development to adulthood (Table 4) could be possible only in case of pre-drilled seed coats. It is noteworthy that most of those plant species that allowed some larval development were members of the tribe Fabeae, and specifically of the genus Lathyrus and much less of Vicia (Additional file 1: Table S2).
The importance of seed coat
The legume seed coat is a significant barrier. In Janzen’s [1] work, no bruchid larva was able to penetrate through > 0.4 mm thick seed coat. From another perspective, Thiery [51, 52] demonstrated that seed coat hardness coupled with low (ca. 6%) moisture content caused higher than 90% L1 mortality with A. obtectus on P. vulgaris. The high L1 mortality as a result of intact seed testa in case of plants grown in nature in this study (Additional file 1: Table S2) calls attention to further factors. These complementary findings indicate that both physical (thickness, smoothness and hardness) and chemical traits of the seed coat contribute to mortality, and it seems that no study has attempted to separate the respective effects.
Chemical factors in the seed coat
One obvious factor that should direct the attention to phytochemicals in seed coats is the observation with several bruchid species [1, 22, 23], A. obtectus included [21, Szentesi pers. obs.], that L1 larvae consume little or none of the seed coat. It was also demonstrated by Stamopoulos and Huignard [53], and in this study (Table 2), that the consumption of bean seed testa was toxic to A. obtectus larvae. A multitude of chemical factors have been identified in legume seeds coats [15]. For example, A. obtectus L1s did not even attempt to penetrate the intact seed coat in the Genisteae tribe, due probably to quinolizidine alkaloids that are present in the testa [54].
Chemical factors in the cotyledon
The cotyledon ultimately determines whether larval development can reach the adult stage. A wide spectrum of secondary plant substances occurs in leguminous tribes, but one of the most characteristic groups is non-protein (also called toxic) amino acids [55]. Not only are their remarkable amounts inside seeds (up to 8% of dry weight [56]) important, but also is their taxonomic distribution in Fabeae. The cotyledon of most Lathyrus species contains a diverse array of toxic amino acids, most frequently homoarginine and lathyrine, whereas Vicia species can be characterized by the dominance of canavanine [57, 58]. In this study, within the Fabeae tribe, V. faba was the only species that allowed development of A. obtectus to adulthood (Table 4). Canavanine can be a significant factor influencing survival of A. obtectus in NH Vicia seeds, in spite of the fact that A. obtectus larvae show a remarkable tolerance to this compound: at 2.0% w/w concentration some adults (4% of the control) still emerged from artificial seeds (Szentesi unpubl. results). However, toxic amino acids occur in many other legume species and they act in combination with several other chemical groups to form the chemical resistance profile of a seed [59]. Indicators of the effects of this complex milieu are malformations and slower development, presumably consequences of the higher metabolic costs of handling substances in the cotyledon.
Evolutionary considerations
The seed coat not only excludes potential exploiters; it also effectively divides the “perception space” of egg-laying insects. Whereas insects can directly perceive plant quality by probing in cases of leaves and fruits, A. obtectus females seem to be unable to judge suitability of cotyledons by the seed coat. The possibility exists that the seed coat can mediate information on the inner quality of the seed; however, the stimulus complex leading to egg-laying on an unsuitable seed is not known. The outcome of preference-performance conditions is further complicated in the field. Although A. obtectus was able to lay eggs into V. unguiculata pods in the laboratory [28], this would not necessarily happen in nature. These findings also direct attention to the need of more tests with pods of different ANH plant species, as this must be the first step to host-range expansion.
An intriguing result of this study was the surprisingly high frequency of development of A. obtectus to adulthood reared on meadow peas (Lathyrus species), when seed coats were pre-drilled. There are no reasons for assuming any evolutionary connection in the relationship, because (1) larvae cannot enter Lathyrus seeds with intact testa, (2) some Lathyrus species do not contain toxic substances to A. obtectus larvae by chance or domestication, and (3) A. obtectus females most likely do not recognise the suitability of Lathyrus seeds for larvae, as they assess only pods for egg-laying in nature. (During the 30 years of collecting legume seeds from Lathyrus species from the field, A. obtectus has never been reared, Szentesi and T. Jermy unpubl. obs.)
As in several other instances [60], the egg-laying by A. obtectus females onto ANH seeds also raises the question as to whether host range expansion has occurred. The first steps in host shifts are behavioural events [61,62,63], viz. the ultimate conditions are recognition and acceptance of the new plant species as host. Although oviposition preference and larval performance likely are governed by different gene complexes [4, 64, 65], there must be genetic covariance between preference and performance to adapt to a new plant species [66, 67]. Egg-laying should be accompanied by physiological adaptations of larvae to handle compounds metabolically once they are inside seeds [68, 69]. Unless preadaptation [70] played a role, as was recently suggested with a related species (A. macrophthalmus) [71, 72], host shift with A. obtectus is less likely. The behaviour of not ingesting the seed coat while boring in, observed with L1 larvae of several bruchid species, can be adaptive. It ensures that, in case of a host shift, larval acceptance or rejection behaviour would be related to the chemicals of cotyledon only.
The following traits maintain A. obtectus within its current status of host specialization: (a) first instar larvae are not constrained to enter an acceptable non-host seed in a no-choice environment, because they have legs, and as long as their energy reserves allow it, they can actively seek suitable hosts; (b) first instar larvae rarely enter acceptable non-host seeds with intact seed testa; and (c) the life cycle of A. obtectus takes place both in stores and in the field, and this spatial segregation regularly interrupts possible breeding and selection on potential additional hosts. Even if generations of beetles were produced on acceptable non-host seeds, the recognition of these plants as suitable oviposition sites must happen based on traits of pods (not seed coats) in the field that, as a first step, would require substantial genetic changes.
Conclusions
This study provides evidence that the seed coat of leguminous plant species hampers both A. obtectus females from judging the suitability of the cotyledon for successful larval development and neonate larvae to enter seeds. Although females are prone to oviposit on dry seeds of many leguminous plant species, the L1 population suffers high mortality while drilling through the seed testa. Making the seed coat penetrable by artificial holes exposed the larvae to possible chemicals factors in the cotyledon that resulted in additional mortality with an exception of 16 leguminous species designated as’acceptable non-hosts’. Apart from this plant group, the female preference and larval performance relation was negative. Further investigations should clarify the role of chemicals of seed testa and cotyledon of leguminous plants in their acceptance and rejection by females and neonate larvae.
Availability of data and materials
The datasets used and/or analysed during the current study are available from the author on reasonable request. Additional information is available in Additional file 1: Tables.
Abbreviations
- H:
-
Host
- ANH:
-
Acceptable non-host
- NH:
-
Non-host
- KW:
-
Kruskal-Wallis
- OR:
-
Odds ratio
- RD:
-
Risk difference
- RR:
-
Risk ratio
- CI:
-
Confidence interval
- GLZ:
-
Generalized linear model
References
Janzen DH. How southern cowpea weevil larvae (Bruchidae: Callosobruchus maculatus) die on nonhost seeds. Ecology. 1977;58:921–7.
Jaenike J. On optimal oviposition behavior in phytophagous insects. Theor Pop Biol. 1978;14:350–6.
Mayhew PJ. Adaptive patterns of host-plant selection by phytophagous insects. Oikos. 1997;79:417–28.
Wiklund C. The evolutionary relationship between adult oviposition preferences and larval host plant range in Papilio machaon. Oecologia. 1975;18:185–97.
Jermy T, Szentesi Á. The role of inhibitory stimuli in the choice of oviposition site by phytophagous insects. Entomol Exp Appl. 1978;24:458–71.
Thompson JN. Evolutionary ecology of the relationship between oviposition preference and performance of offspring in phytophagous insects. Entomol Exp Appl. 1988;47:3–14.
Courtney SP, Chen GK, Gardner A. A general model for individual host selection. Oikos. 1989;55:55–65.
Gripenberg S, Mayhew PJ, Parnell M, Roslin T. A meta-analysis of preference-performance relationships in phytophagous insects. Ecol Lett. 2010;13:383–93.
Bernays EA. Neural limitations in phytophagous insects: implications for diet breadth and evolution of host affiliation. Annu R Entomol. 2001;46:703–28.
de la Charlery Masselière M, Facon B, Hafsi A, Duyck P-F. Diet breadth modulates preference-performance relationships in a phytophagous insect community. Sci Rep. 2017;7:16934.
Gu H, Cao A, Walter GH. Host selection and utilisation of Sonchus oleraceus (Asteraceae) by Helicoverpa armigera (Lepidoptera: Noctuidae): a genetic analysis. Ann Appl Biol. 2001;138:293–9.
Cotter SC, Edwards OR. Quantitative genetics of preference and performance on chickpeas in the noctuid moth Helicoverpa armigera. Heredity. 2006;96:396–402.
Scheirs J, Jordaens K, De Bruyn L. Have genetic trade-offs in host use been overlooked in arthropods? Evol Ecol. 2005;19:551–61.
Rios RS, Cardenas M, Gonzalez K, et al. Effects of host plant and maternal feeding experience on population vital rates of a specialized leaf beetle. Arthr-Plant Inter. 2013;7:109–18.
Bisby FA, Buckingham J, Harbome JB. Phytochemical dictionary of the Leguminosae. Dordrecht: Chapman & Hall/Springer-Science+Business Media, B.V; 1994.
Johnson CD. Adaptive radiation of Acanthoscelides in seeds: examples of legume-bruchid interactions. In: Stirton CH, Zarucchi JL, editors. Advances in legume biology. Monographs in Systematic Botany from the Missouri Botanical Garden. 1989;29:755–80.
Fox CW. A quantitative genetic analysis of oviposition preference and larval performance on two hosts in the bruchid beetle Callosobruchus maculatus. Evolution. 1993;47:166–75.
Tucić N, Mikuljanac S, Stojković O. Genetic variation and covariation among life-history traits in population of Acanthoscelides obtectus maintained on different hosts. Entomol Exp Appl. 1997;85:247–56.
Milanović D, Gliksman I. Selection responses and quantitative-genetic analysis of preadult performance on two host plants in the bean weevil, Acanthoscelides obtectus. Entomol Exp Appl. 2004;113:125–33.
Jarry M. Histoire naturelle de la bruche du haricot dans un agrosysteme du Sud-Ouest de la France. Contribution a l’étude de la structure et de la dynamique des populations d’Acanthoscelides obtectus Say dans les stocks et les cultures de Phaseolus vulgaris. Dissertation, University of Pau; 1984.
Thiery D. Etude du comportement des larves neonates d’ Acanthoscelides obtectus Say (Coleoptere, Bruchidae) en presence des graines de leur plante-hôte Phaseolus vulgaris L. Dissertation, Francois Rabelais University of Tours; 1981.
Coe M, Coe C. Large herbivores, Acacia trees and bruchid beetles. SA J Sci. 1987;83:624–35.
Southgate BJ. Observations on the larval emergence in Callosobruchus chinensis (Coleoptera: Bruchidae). Entomol Gen. 1984;9:177–80.
Lewis G, Schrire B, MacKinder B, Lock M. Legumes of the world. Kew: Royal Botanic Garden; 2005.
Leroi B, Jarry M. Relations d’ Acanthoscelides obtectus avec différentes espèces de Phaseolus: Influence sur la fécondité et possibilités de développement larvaire. Entomol Exp Appl. 1981;30:73–82.
Vukasović P, Glumac S, Martinović A. Contribution a l’étude de la polyphagie de la bruche du haricot (Acanthoscelides obtectus Say). Lept Nauć Radova Poljopriv Fakult Novom Sadu. 1961;5:1–35 ((In Serb.)).
Hamraoui A, Regnault-Roger C. Oviposition and larval growth of Acanthoscelides obtectus Say (Col., Bruchidae) in regard to host and non-host plants from Leguminosae family. J Appl Entomol. 1995;119:195–9.
Jarry M, Bonet A. La bruche du haricot, Acanthoscelides obtectus Say (Coleoptera, Bruchidae), est-elle un danger pour le cowpea, Vigna unguiculata (L.) Walp.? Agronomie. 1982;2:963–8.
Szentesi Á. Resource assessment and clutch size in the bean weevil Acanthoscelides obtectus. Pest Manag Sci. 2003;59:431–6.
Szentesi Á. Studies on the mass-rearing of Acanthoscelides obtectus Say (Col., Bruchidae). Acta Phytopathol Acad Sci Hung. 1972;7:453–63.
Szentesi Á, Jermy T. Predispersal seed predation in leguminous species: seed morphology and bruchid distribution. Oikos. 1995;73:23–32.
De Boer G, Hanson FE. Food plant selection and induction of feeding preference among host and non-host plants in larvae of the tobacco hornworm Manduca sexta. Entomol Exp Appl. 1984;35:177–93.
Tabachnick BG, Fidell LS. Using multivariate statistics. Boston: Pearson Education, Inc.; 2013.
Hosmer DW, Lemeshow S. Applied logistic regression. New York: John Wiley and Sons, Inc.; 2000.
StatSoft, Inc. STATISTICA (data analysis software system), version 6. www.statsoft.com. 2003.
Nakagawa S, Cuthill IC. Effect size, confidence interval and statistical significance: a practical guide for biologists. Biol Rev. 2007;82:591–605.
Kline RB. Beyond significance testing. Statistics reform in the behavioral sciences. Washington: American Psychological Association; 2013.
Kleinbaum DG, Klein M. Logistic regression. A self-learning text. New York: Springer-Verlag; 2002.
Labeyrie V. Etude de la repartition des larves neonates d'Acanthoscelides obtectus Say. Proc XIIIth Int Congr Entomol, London; 1965. p. 310-311
Vuts J, Woodcock CM, König L, Powers SJ, Pickett JA, et al. Host shift induces changes in mate choice of the seed predator Acanthoscelides obtectus via altered chemical signalling. PLoS ONE. 2018;13:e0206144.
Mitchell R. The evolution of oviposition tactics in the bean weevil, Callosobruchus maculatus (F.). Ecology. 1975;56:696–702.
Cronin JT, Abrahamson WG. Goldenrod stem galler preference and performance: effects of multiple herbivores and plant genotypes. Oecologia. 2001;127:87–96.
Zhang P-J, Liu SS. Experience induces a phytophagous insect to lay eggs on a nonhost plant. J Chem Ecol. 2006;32:747–53.
Ikonen A, Sipura M, Miettinen S, Tahvanainen J. Evidence for host race formation in the leaf beetle Galerucella lineola. Entomol Exp Appl. 2003;108:179–85.
Larsson S, Ekbom B. Oviposition mistakes in herbivorous insects: confusion or a step towards a new host plants. Oikos. 1995;72:155–60.
Fox CW, Lalonde RG. Host confusion and the evolution of insect diet breadths. Oikos. 1993;67:577–81.
Hopkins RJ, Ekbom B. The pollen beetle, Meligethes aeneus, changes egg production rate to match host quality. Oecologia. 1999;120:274–8.
Tuda M, Chou LY, Niyomdham C, Buranapanichpan S, Tateishi Y. Ecological factors associated with pest status in Callosobruchus (Coleoptera: Bruchidae): high host specificity of non-pests to Cajaninae (Fabaceae). J Stored Prod Res. 2005;41:31–45.
Simmonds MSJ, Blaney WM, Birch ANE. Legume seeds: the defenses of wild and cultivated species of Phaseolus against attack by bruchid beetles. Ann Bot. 1989;63:177–84.
Johnson CD. Seed beetle host specificity and systematics of the Leguminosae. In: Polhill RM, Raven PH, editors. Advances in legume systematics. Kew: Royal Botanical Gardens; 1981. p. 995–1027.
Thiery D. Influence de la teneur en eau et de la dureté du tégument des graines de Phaseolus vulgaris sur la fréquence de pénétration des larves néonates d’Acanthoscelides obtectus. Entomol Exp Appl. 1982;32:141–5.
Thiery D. Hardness of some fabaceous seed coats in relation to larval penetration by Acanthoscelides obtectus (Say) (Coleoptera: Bruchidae). J Stored Prod Res. 1984;20:177–81.
Stamopoulos DC, Huignard J. L’influence des diverses parties de la graine de haricot (Phaseolus vulgaris) sur le développement des larves d’Acanthoscelides obtectus (Coléoptère Bruchidae). Entomol Exp Appl. 1980;28:38–46.
Szentesi Á, Wink M. Fate of quinolizidine alkaloids through three trophic levels: Laburnum anagyroides (Leguminosae) and associated organisms. J Chem Ecol. 1991;17:1557–73.
Fowden L. The non-protein amino acids of plants. In: Reinhold L, Liwschitz Y, editors. Progress in phytochemistry, vol. 2. London: Interscience Publication; 1970. p. 203–66.
Bell EA, Lackey JA, Polhill RM. Systematic significance of canavanine in the Papilionoideae. Biochem Syst Ecol. 1978;6:201–12.
Bell EA. Toxic amino acids in the Leguminosae. In: Harborne J, editor. Phytochemical ecology. London and New York: Academic; 1972. p. 163–77.
Bell EA, Tirimanna ASL. Association of amino acids and related compounds in the seeds of forty-seven species of Vicia: their taxonomic and nutritional significance. Biochem J. 1965;97:104–11.
Birch ANE, Simmonds MSJ, Blaney WM. (1989) Chemical interactions between bruchids and legumes. In: Stirton CH, Zarucchi JL, editors. Advances in legume biology. Monographs in Systematic Botany from the Missouri Botanical Garden. 1989;29:781–809.
Thompson JN, Pellmyr O. Evolution of oviposition behavior and host preference in Lepidoptera. Annu R Entomol. 1991;36:65–89.
Futuyma DJ. Selective factors in the evolution of host choice by phytophagous insects. In: Ahmad S, editor. Herbivorous insects. Host-seeking behavior and mechanisms. New York: Academic Press, Inc.; 1983. p. 227–44.
Jermy T. Evolution of insect/host plant relationships. Am Nat. 1984;124:609–30.
Courtney SP, Kibota TT. Mother doesn’t know best: selection of hosts by ovipositing insects. In: Bernays EA, editor. Insect-plant interactions, vol. II. Boca Raton: CRC Press; 1990. p. 161–88.
Berenbaum MR, Feeny PP. Chemical mediation of host-plant specialization the papilionid paradigm. In: Tilmon KJ, editor. Specialization speciation and radiation. The evolutionary biology of herbivorous insects. Berkeley: University of Californisa Press; 2008. p. 3–19.
Messina FJ, Gompert Z. Evolution of host acceptance and its reversibility in a seed beetle. Ecol Entomol. 2017;42:42–50.
Via S. Genetic covariance between oviposition preference and larval performance in an insect herbivore. Evolution. 1986;40:778–85.
Jaenike J. Genetic population structure of Drosophila tripunctata: patterns of variation and covariation of traits affecting resource use. Evolution. 1989;43:1467–82.
Huignard J, Baehr JC, Desroches P, Mandon N. Adaptation of a Callosobruchus maculatus strain to Vicia faba, as its new host plant. In: Städler E, Rowell-Rahier M, Baur R, editors. Insect-plant relationships. Dordrecht: Kluwer Academic Publishers; 1996. p. 156–9.
Jankovic-Tomanic M, Jovanovic DS, Savkovic U, et al. Host expansion modifies activity of phosphatases in a legume store pest Acanthoscelides obtectus (Say). J Stored Prod Res. 2015;62:32–5.
Janzen DH. On ecological fitting. Oikos. 1985;45:308–10.
Tuda M, Wu L-H, Tateishi Y, Niyomdham C, Buranapanichpan S, Morimoto K, et al. A novel host shift and invaded range of a seed predator, Acanthoscelides macrophthalmus (Coleoptera: Chrysomelidae: Bruchinae), of an invasive weed Leucaena leucocephala. Entomol Sci. 2009;12:1–8.
Tuda M, Wu LH, Yamada N, Wang CP, Wu WJ, Buranapanichpan S, Kagoshima K, et al. Host shift capability of a specialist seed predator of an invasive plant: roles of competition, population genetics and plant chemistry. Biol Invasions. 2014;16:303–13.
White R and ILDIS (International Legume Database & Information Service). 2018. https://ildis.org/index.shtml. Accessed Jan 2021.
Acknowledgements
I thank Tibor Jermy and Dénes Schmera for comments on an earlier draft, and Margit Mydló, Klára Ferencz, Teréz Tihanyi and Katalin Ujj for very valuable help in experimental work. András Horánszky and Gábor Fekete assisted in identification of plant species. I am indebted to Donald C. Weber (Beltsville) and József Vuts (Harpenden) for helpful advice and linguistic improvement of the text. Suggestions by anonymous referees greatly improved the manuscript. The provision from several seed samples by plant breeder companies is gratefully acknowledged. The study was supported by the Hungarian National Research Grant OTKA-1434.
Funding
OTKA-1434 of the Hungarian National Research Grant.
Author information
Authors and Affiliations
Contributions
Not applicable. The author read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This article does not contain any work with protected animal species, therefore, no ethical approval/permission was necessary from an appropriate ethics committee/review board local, regional or national board, to carry out sampling and experiments described in the Methods.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1.
Additional tables.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Szentesi, Á. How the seed coat affects the mother’s oviposition preference and larval performance in the bean beetle (Acanthoscelides obtectus, Coleoptera: Chrysomelidae, Bruchinae) in leguminous species. BMC Ecol Evo 21, 171 (2021). https://doi.org/10.1186/s12862-021-01892-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12862-021-01892-9