
Abstract
Skeletal muscle is the largest metabolic organ of the human body. Maintaining the best quality control and functional integrity of mitochondria is essential for the health of skeletal muscle. However, mitochondrial dysfunction characterized by mitochondrial dynamic imbalance and mitophagy disruption can lead to varying degrees of muscle atrophy, but the underlying mechanism of action is still unclear. Although mitochondrial dynamics and mitophagy are two different mitochondrial quality control mechanisms, a large amount of evidence has indicated that they are interrelated and mutually regulated. The former maintains the balance of the mitochondrial network, eliminates damaged or aged mitochondria, and enables cells to survive normally. The latter degrades damaged or aged mitochondria through the lysosomal pathway, ensuring cellular functional health and metabolic homeostasis. Skeletal muscle atrophy is considered an urgent global health issue. Understanding and gaining knowledge about muscle atrophy caused by mitochondrial dysfunction, particularly focusing on mitochondrial dynamics and mitochondrial autophagy, can greatly contribute to the prevention and treatment of muscle atrophy. In this review, we critically summarize the recent research progress on mitochondrial dynamics and mitophagy in skeletal muscle atrophy, and expound on the intrinsic molecular mechanism of skeletal muscle atrophy caused by mitochondrial dynamics and mitophagy. Importantly, we emphasize the potential of targeting mitochondrial dynamics and mitophagy as therapeutic strategies for the prevention and treatment of muscle atrophy, including pharmacological treatment and exercise therapy, and summarize effective methods for the treatment of skeletal muscle atrophy.
Similar content being viewed by others

Introduction
Skeletal muscle, which accounts for approximately 40% of total body weight, is critical for movement control, postural support, heat production, and metabolic balance [1]. However, the delicate balance between protein synthesis and degradation in skeletal muscle is disrupted when individuals experience various adverse pathological and physiological stressors, resulting in a decrease in muscle mass and strength [2]. This process is commonly referred to as skeletal muscle atrophy and is characterized by a reduced cross-sectional area of muscle fibers, loss of muscle mass and strength, and loss of balance between protein synthesis and degradation. Skeletal muscle atrophy has significant health consequences, and it was recognized as a distinct disease according to its own International Classification of Diseases, 10th Revision, Clinical Modification (ICD-10-CM) code (M62.84) in 2016 [3]. Skeletal muscle atrophy not only is prevalent among older individuals but also may occur secondary to peripheral nerve injury, cancer, diabetes, heart failure, and other diseases. Consequently, skeletal muscle atrophy can be categorized into primary and secondary skeletal muscle atrophy based on the underlying predisposing factors [4]. Muscle fiber type conversion occurs in both types of skeletal muscle atrophy, involving transitions from slow muscle fibers to fast muscle fibers, as well as from fast muscle fibers to slow muscle fibers [5, 6]. The impact of muscle atrophy is not limited to a decrease in quality of life; it is also associated with increased morbidity and mortality [7]. On the other hand, maintaining skeletal muscle health is closely related to reducing morbidity and mortality, which highlights the importance of maintaining the best muscle mass in the human body [8]. Currently, skeletal muscle atrophy has evolved into a critical global clinical issue, necessitating the development of innovative treatment approaches.
Mitochondria are important sites for energy production in the body, and skeletal muscle contraction requires a large amount of energy supply. To sustain normal muscle function and movement, the energy needed for skeletal muscle contraction must align with the ATP molecules produced by mitochondria. Research has demonstrated that skeletal muscle fibers, also known as mitochondrial pools, have two unique mitochondrial populations, subsarcolemmal (SS) mitochondria and intermyofibrillar (IMF) mitochondria [9]. These separate mitochondrial pools work together and generate ample ATP through oxidative phosphorylation (OXPHOS) to support the basic and contractile energy needs of the skeletal muscle network. To elucidate the pathogenesis of skeletal muscle atrophy, various biological mechanisms, including the ubiquitin‒proteasome system, autophagy–lysosomal system, cysteine aspartic acid system, and calpain system, have been proposed [10]. Certainly, there is compelling evidence supporting the idea that the accumulation of mitochondrial dysfunction contributes to the energy deficit and alterations in the balance between protein synthesis and degradation observed in skeletal muscle [11,12,13]. This dysfunction can promote the development of skeletal muscle atrophy by activating various pathways, including accumulation of reactive oxygen species (ROS), decreased mitochondrial biogenesis, imbalance in mitochondrial dynamics, and impaired mitophagy [14, 15]. These evidences indicate that mitochondria play a particularly prominent role in maintaining skeletal muscle health, emphasizing its possibility and importance in preventing skeletal muscle atrophy.
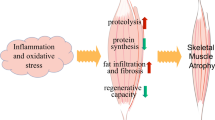
Mitochondria, as a dynamic organelle network, can be properly adjusted according to external pressure to meet the energy needs of skeletal muscle survival and contraction. The primary function of mitochondria relies on OXPHOS within the electron transport chain (ETC) to generate adenosine triphosphate (ATP), which serves as the primary “fuel” for cellular metabolic activities. In addition, mitochondria also play a role in programmed cell death, apoptosis, calcium regulation, cell metabolism regulation, and other biological processes [16]. The health of skeletal muscle is closely related to the integrity and optimal function of the mitochondrial network [17]. The normal operation of this network depends on several key processes, including mitochondrial dynamics (mediated by mitochondrial fusion and fission), mitophagy (involving mitochondrial self-clearance and protection), mitochondrial biogenesis, mitochondrial-derived vesicles, and others [18, 19]. Mitochondrial dynamics maintains normal morphology and quantity through continuous fusion and fission, while mitophagy degrades damaged mitochondria through the autophagy pathway, ensuring the health of the mitochondrial network [20, 21]. However, when individuals are exposed to a variety of endogenous stresses (denervation, DMD gene mutation, chronic inflammation, aging, etc.) and exogenous stresses (such as hypoxia, starvation, minimal exercise, cancer cachexia, etc.), reactive oxygen species (ROS) and the mitochondrial membrane potential in skeletal muscle rapidly increase, resulting in mitochondrial functional network disorders, including imbalances in mitochondrial dynamics and mitochondrial autophagy damage. Consequently, skeletal muscle atrophy is often characterized by an accumulation of mitochondrial ROS and a decline in mitochondrial function. Excessive ROS can induce the oxidation of myofibrillar proteins, promoting their degradation. Additionally, elevated ROS levels can impede the initial stage of mRNA translation, thereby inhibiting the protein synthesis pathway [22]. Moreover, there is an increase in the expression of two skeletal muscle-specific E3 ubiquitin ligases: muscle-specific ring finger protein 1 (MuRF1) and muscle atrophy F-box protein (Atrogin-1). During this period, several atrophy pathways are triggered in skeletal muscle: (1) mitochondrial disorders can lead to decreased mitochondrial membrane potential and excessive production of ROS, which in turn activate oxidative stress, inflammation, protein synthesis, and degradation-related signaling pathways, triggering muscle atrophy programs [23, 24]. (2) Various non-coding RNAs accelerate skeletal muscle atrophy by targeting mitochondrial dynamics and mitophagy-related proteins, resulting in reduced mitochondrial numbers and dysfunction [25,26,27,28] (Fig. 1).

The molecular mechanisms of skeletal muscle atrophy associated with mitochondrial dynamics and mitophagy. When individuals encounter diverse endogenous stressors, such as denervation, aging, chronic inflammation, and exogenous factors, such as hypoxia, starvation, or minimal exercise, the muscle cell mitochondrial membrane potential diminishes while reactive oxygen species accumulate. This scenario prompts the initiation of multiple signaling pathways and activation of various non-coding RNAs, fostering an imbalance in mitochondrial dynamics and disrupting mitophagy in muscle cells. This, in turn, accelerates the process of muscle atrophy and degradation
Therefore, maintaining the integrity of skeletal muscle mitochondria is crucial for skeletal muscle health and could be a potential therapeutic target for treating skeletal muscle atrophy. Restoring healthy mitochondrial function has been demonstrated to reduce muscle atrophy and improve health, especially by increasing mitochondrial dynamics and mitophagy. In this review, we concentrate on the current research status of mitochondrial dynamics and mitophagy in the context of skeletal muscle atrophy development and underscore the underlying mechanisms of action, suggesting a potential strategy for preventing and treating skeletal muscle atrophy by leveraging mitochondrial functions.
Intermodulation of mitochondrial dynamics and mitophagy during skeletal muscle atrophy
Skeletal muscle mitochondria, as highly adaptable organelles, require continuous monitoring to maintain structural and functional integrity. Mitochondrial dynamics and mitophagy are key mechanisms for maintaining mitochondrial quality. Recent studies have demonstrated a close relationship between mitophagy and mitochondrial dynamics. Mitochondrial dynamics help to isolate damaged mitochondria from a healthy mitochondrial network. Moreover, mitophagy may be the result of mitochondrial dynamics, which helps to degrade isolated damaged or abnormal mitochondria. These two proteins are not completely independent of each other but have a cross-talk relationship and jointly maintain a healthy mitochondrial network. Under normal physiological circumstances, mitochondrial dynamics intricately maintain the delicate equilibrium of the mitochondrial network by orchestrating seamless interplay between fusion and fission events. This dynamic process ensures the fulfillment of the body’s energy synthesis demands [29]. However, during the process of skeletal muscle atrophy, there is an imbalance in mitochondrial network homeostasis. Activation of FoxO3 family proteins occurs via the AMPK pathway, which recruits two muscle-specific E3 ubiquitin ligases: MuRF1 and Atrogin-1 [23]. Consequently, this activation leads to the overexpression of the mitochondrial fission-related proteins dynamin 1 Like (Drp1) and fission mitochondrial 1 (Fis1), resulting in excessive mitochondrial fission [30]. Excessive mitochondrial fission resulting from imbalanced mitochondrial network homeostasis leads to a fragmented state of the mitochondrial network. In response, an adaptive mitophagy pathway is triggered, which selectively eliminates damaged mitochondria from the network [31]. Taken together, these studies indicate that Drp1-mediated mitochondrial fission is a prerequisite for triggering mitophagy. Conversely, the inhibition of mitochondrial fission also initiates the initiation of mitophagy. The protein Fis1, which is responsible for mitochondrial fission, plays a crucial role in the pivotal intersections that trigger mitophagy within the primary pathways of the stress response. The absence of Fis1 leads to mitochondrial swelling and excessive fusion, thereby triggering mitophagy in skeletal muscle. Mechanistically, Fis1 downregulation leads to the accumulation of polyubiquitinated proteins in muscle fibers, which bind to the LC3 protein through the LC3-interacting region (LIR), thereby inducing abnormal mitophagy and leading to muscle atrophy and degeneration [32, 33]. Moreover, Fis1 depletion can also induce mitophagy through the PINK1/Parkin-independent pathway. Fis1 deficiency causes abnormal accumulation of STX17 in mitochondria, exposing the N-terminus and promoting self-oligomerization, thereby triggering mitophagy [34].
Similarly, abnormalities in mitochondrial fusion also influence mitophagy. Several studies have shown that skeletal muscle atrophy in aged mice is partially caused by the downregulation of mitofusin2 (MFN2) expression [35, 36]. Loss of MFN2 inhibits the process of mitochondrial fusion, leading to an extensive amount of broken or swollen mitochondria in skeletal muscle, accompanied by a decrease in mitophagy flux. Interestingly, MFN2 deficiency can also induce ROS-dependent adaptive signaling pathways through the induction of hypoxia inducible factor 1 subunit alpha (HIF-1α) transcription factor and BCL2 interacting protein 3 (BNIP3), which can further compensate for the loss of mitophagy [35]. This compensatory mechanism aims to counterbalance the loss of mitophagy and minimize mitochondrial damage. Similarly, in the skeletal muscle of young mice, knockout of MFN2 results in impaired mitophagy and the manifestation of muscle aging-related phenotypes [36]. These findings indicate that changes in proteins related to mitochondrial dynamics can affect mitophagy through various mechanisms to maintain a healthy mitochondrial network.
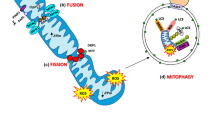
Impaired mitophagy also affects the occurrence of mitochondrial dynamics. In addition to their involvement in mitochondrial dynamics, proteins associated with mitochondrial dynamics, such as Drp1, MFN1, and MFN2, also serve as substrates for the PINK1–parkin pathway of mitochondrial autophagy. For instance, PTEN induced kinase 1 (PINK1) can directly phosphorylate Drp1 at Ser616 [37]. Indeed, Drp1 is a prerequisite for initiating mitochondrial fission [38]. In addition, MFN2 rather than MFN1 has been shown to be phosphorylated by PINK1 and to become a receptor for the ubiquitin ligase Parkin, which activates mitophagy through ubiquitination mediated degradation [39]. Similarly, mitophagy through the PINK1–parkin-independent pathway can also regulate mitochondrial dynamics. Under stress conditions, the mitophagy receptor FUNDC1 can dephosphorylate and dissociate OPA1 and subsequently bind to DNM1L, impacting mitochondrial fusion or fission [40]. Obviously, a distinct intermodulation exists between mitochondrial dynamics and mitophagy during skeletal muscle atrophy (Fig. 2).

Mutual regulation of mitophagy and mitochondrial dynamics. Initially, PINK1, a pivotal protein in mitophagy, directly phosphorylates DRP1 and MFN2, facilitating the activation of both mitochondrial fission and fusion. Moreover, phosphorylated MFN2 functions as a receptor for Parkin, subsequently initiating mitophagy. Furthermore, any mitochondrial breakage or swelling arising from irregularities in mitochondrial fusion or fission prompts the activation of mitophagy, ensuring the sustenance of a healthy mitochondrial network
The physiological and pathological significance of mitochondrial dynamics in skeletal muscle atrophy
Mitochondrial dynamics involve the continuous fusion and fission of mitochondria and are now recognized as fundamental processes in mitochondrial metabolism and turnover [41]. It can be regulated by various cellular signaling pathways to ensure cell survival under different stressful conditions. Mitochondrial dynamics have provided numerous advantages to mitochondria, such as efficient transport, enhanced homogenization of mitochondrial populations, and effective oxidative phosphorylation. When this process is disrupted, it can trigger a series of diseases, including skeletal muscle atrophy [42]. Mitochondrial fusion typically leads to elongation of the mitochondria, resulting in an extended mitochondrial network and improved network connectivity. This elongation may contribute to an increased level of oxidative phosphorylation within the cell [29]. Furthermore, mitochondrial fusion helps to dilute the concentration of damaged mitochondria, preventing the accumulation of dysfunctional mitochondria. This process is crucial for maintaining the overall health and functionality of the organism [43]. On the other hand, mitochondrial fission plays a role in separating damaged mitochondria from the healthy mitochondrial network, allowing for their rejuvenation and increasing energy output efficiency [44]. Therefore, reasonable fusion and fission are crucial for the health of the mitochondrial network.
The imbalance between mitochondrial fission and fusion leads to mitochondrial fragmentation, which is a prerequisite for mitophagy. Excessive mitochondrial fusion hinders proper mitophagy, while excessive mitochondrial fission results in an excess of independent mitochondria that are inefficient at energy production [45]. The regulation of these processes involves specific proteins, particularly members of the dynamin superfamily [46]. Key proteins involved in mitochondrial dynamics include the mitochondrial fission proteins Drp1, Fis1, MFF Mid49, and Mid51 and the mitochondrial fusion proteins MFN1/2 and optic atrophy 1 (OPA1). These proteins utilize GTP hydrolysis to drive associated proteins in the mitochondrial membrane and cytoplasm, facilitating mitochondrial fusion and fission events. Disruptions in the expression of proteins involved in mitochondrial dynamics within skeletal muscle can contribute to muscle loss under various physiological and pathological conditions, such as age-related muscle atrophy, muscle wasting associated with cancer cachexia, and sarcopenia resulting from physical inactivity [47,48,49]. The specific process of skeletal muscle atrophy caused by an imbalance in mitochondrial dynamics is described in the following sections (Fig. 3).

The importance of mitochondrial dynamics in maintaining skeletal muscle health. An imbalance in mitochondrial fission and fusion precipitates mitochondrial dysfunction, a primary cause of muscle atrophy and degeneration. Conversely, restoring mitochondrial dynamics through treatment in the physiological state of atrophic muscles can impede the advancement of muscle atrophy
Mitochondrial fusion dysregulation and skeletal muscle atrophy under different physiological/pathological conditions
Mitochondrial fusion is the process of fusing two adjacent mitochondrial outer membranes and inner membranes to form new mitochondria, ensuring the efficient exchange of substances within mitochondria (mitochondrial DNA, proteins, metabolites, etc.) [50]. Mitochondrial fusion consists of two crucial processes: outer membrane fusion mediated by MFN and inner membrane fusion mediated by OPA1. These processes rely on the coordination of proteins located on the outer mitochondrial membrane (OMM) and inner mitochondrial membrane (IMM). This fusion enables a greater frequency of metabolic substance exchange, the transfer of membrane potential from the inner mitochondrial membrane’s matrix side to the outer membrane, enhanced OXPHOS reactions, and reduced production of reactive oxygen species. These effects are particularly prominent in metabolically active cells, such as skeletal muscle cells, which possess a tightly interconnected mitochondrial network [51]. In addition, the excitation–contraction coupling process of skeletal muscle cells is also supported by mitochondrial fusion, and the mitochondrial fusion rate is positively correlated with the degree of skeletal muscle fiber contraction [52]. Surprisingly, the efficiency of mitochondrial fusion varies between different types of muscle fibers, with higher fusion rates observed in oxidative fibers than in glycolytic fibers [53]. The reason for this difference is that the content of mitochondria in glycolytic fibers is low. Compared with oxidative fibers, glycolytic fibers are less dependent on oxidative phosphorylation and more prone to fatigue. Consequently, the mitochondrial network is adaptively organized according to the muscle fiber type to match the metabolism and contraction characteristics of each muscle fiber type. Specifically, mitochondria in oxidative muscle fibers exhibit a lattice-like pattern, while glycolytic fibers exhibit a more fragmented mitochondrial network [53]. Interestingly, studies have shown that the types of muscle fibers involved in the process of skeletal muscle atrophy caused by different causes can often change according to different causes, including the transition from slow muscle to fast muscle and from fast muscle to slow muscle [6, 54].
In recent decades, extensive research has demonstrated the essential roles of fusion proteins, including MFN1, MFN2, and OPA1, in various cellular functions [52, 55]. The absence or reduction of these proteins has also been associated with the development of skeletal muscle atrophy under different physiological and pathological conditions [25, 56, 57]. First and foremost, abundant evidence suggests that mitochondrial fusion plays a vital biological role during early development. Notably, complete deletion of MFN1, MFN2, or OPA1 has been shown to result in embryonic lethality [58]. Second, during individual development, specific MFN2 deletion can lead to damage of neuromuscular synapses and cause skeletal muscle atrophy [59]. Conversely, it should be noted that in the study mentioned, overexpression of MFN2 specifically in neurons was found to rescue the loss of neuromuscular synapses and skeletal muscle atrophy in transgenic SOD1G93A mice (a mouse model of amyotrophic lateral sclerosis) and aged mice (a mouse model of skeletal muscle natural atrophy) [59]. Similarly, a recent study demonstrated that overexpression of MFN2 in skeletal muscle of aged mice led to slight muscle hypertrophy, indicating that MFN2 overexpression may be a viable approach to mitigate age-related muscle atrophy [60]. Additionally, in humans, the loss of MFN2 and subsequent failure of mitochondrial fusion are associated with the development of Charcot–Marie–Tooth type 2A (CMT2A, a neurodegenerative disease characterized by clinical manifestations such as muscle atrophy) [61]. Although the structures and functions of MFN1 and MFN2 are similar, some studies have shown that there are some differences between the two in terms of functional regulation and disease correlation. For instance, a study conducted by Gao Song’s team revealed that MFN2 exhibits greater membrane fusion efficiency than does MFN1 in primates [61]. This disparity is attributed to the persistent dimer formation of MFN2 even after hydrolysis at the GTPase structural domain interface. Therefore, the ratio of MFN1 to MFN2 plays a significant role in modulating the mitochondrial fusion process. On the other hand, MFN2, but not MFN1, is capable of connecting the endoplasmic reticulum (ER) to mitochondria, establishing a physical foundation for important processes associated with ER–mitochondrial interactions, including calcium ion (Ca2+) signaling [62]. Notably, specific knockout of OPA1 in skeletal muscle can cause more severe muscle atrophy-related phenotypes than a deletion of MFN1 or MFN2 can, indicating that OPA1 can have an important impact on skeletal muscle atrophy in other biological processes other than mitochondrial fusion [56, 63]. In terms of molecular mechanisms, the loss of OPA1 in skeletal muscle can induce the accumulation of intracellular ROS and the release of mitochondrial DNA (mtDNA), which can trigger the nuclear translocation of different transcription factors, including FoxO3, NFkB, ATF4, and Nrf2. Subsequently, these transcription factors enter the nucleus and interact with skeletal muscle atrophy-related genes, FGF21 and inflammatory cytokines (e.g., IL6, IL10, TNF), resulting in skeletal muscle atrophy [56].
In summary, disruption of mitochondrial fusion in skeletal muscle leads to an imbalance between protein synthesis and degradation, resulting in accelerated skeletal muscle atrophy. These findings underscore the importance of fusion proteins in maintaining skeletal muscle health and suggest that they could be potential therapeutic targets for preventing or treating muscle atrophy.
The function and regulation of mitochondrial fission in skeletal muscle atrophy
Mitochondrial division is a complex regulatory process that mainly depends on DRP1. This process is vital for various cellular functions, including cell fission, programmed cell death, and maintenance of healthy mitochondria [64]. The sites of mitochondrial fission occur at mitochondrial–endoplasmic reticulum (ER) contact points marked by mtDNA, and the identification of these sites relies primarily on the association between mitochondria and the ER [65]. Following identification, the DRP1 protein is recruited to the identified cleavage site, where it forms a circular protein segmentation apparatus around the mitochondria, facilitating the contraction and division of the mitochondrial membrane [66]. Mitochondrial fission is not solely driven by the DRP1 protein. In mammals, DRP1 typically collaborates with classical dynamin 2 (DYN2) to facilitate a smooth process of mitochondrial fission [67]. Although the molecular mechanism of mitochondrial fission integrity has largely unknown, studies in recent decades have shown that activation of the DRP1 protein is carried out in a variety of ways, including through interactions with receptor proteins (Fis1, MFF, MID49, and MID51) [68], posttranslational modification [69], and contact activation with various organelles [70]. These findings indicate the complexity and versatility of Drp1 regulation in orchestrating mitochondrial fission. Furthermore, a recent study employing superresolution microscopy conducted a comprehensive analysis of mitochondrial fission, resulting in the redefinition of two spatially distinct types of fission: midzone fission and peripheral fission. These terms describe whether fission occurs at central or other locations in mitochondria [64]. The study revealed that peripheral division allows damaged material to be released into smaller mitochondria for mitophagy, whereas division in the intermediate region leads to mitochondrial proliferation. This study has provided a new direction for understanding the molecular components and regulation of mitochondrial fission mechanisms.
Currently, numerous studies are investigating the crucial role of mitochondrial fission in maintaining the overall health of skeletal muscle. Both excessive and insufficient mitochondrial fission have been associated with the development of various severe human diseases, including skeletal muscle atrophy. These studies underscore the significance of maintaining a balanced mitochondrial fission process for the proper functioning of skeletal muscle and highlight the potential implications for understanding and treating related disorders. In the following section, we emphasize the progress and mechanism of mitochondrial fission in skeletal muscle atrophy through the key proteins of Drp1.
Function and regulation of Drp1 in skeletal muscle atrophy
Drp1 is a critical protein involved in mitochondrial fission. The targeted deletion of DRP1 in muscle has been shown to induce severe skeletal muscle atrophy by impairing protein synthesis and disrupting Ca2+ homeostasis between the cytoplasm and mitochondria [47, 71]. As a therapeutic target, restoring the expression of DRP1 in aging Drosophila can alleviate age-related sarcopenia by enhancing mitochondrial morphology and function in muscle tissue and promoting effective mitophagy [72]. Paradoxically, in middle-aged mice, both knockdown and overexpression of Drp1 have negative effects on mitochondrial mass, leading to several key features of muscle atrophy. These findings indicate that targeting Drp1 expression is unlikely to be a feasible approach for combating muscle atrophy [73]. Similarly, in clinical settings, extensive evidence has emerged highlighting a significant link between mitochondrial dynamics and secondary skeletal muscle atrophy in patients with amyotrophic lateral sclerosis (ALS). Dysfunction of mitochondria mediated by Drp1 overactivation is considered to be one of the key factors leading to ALS pathology [74, 75]. The contrasting findings observed in studies examining mitochondrial fission in both mammals and Drosophila add an intriguing layer of complexity to our understanding. Ultimately, these conflicting results underscore the importance of maintaining mitochondrial division within the physiological context to ensure the integrity of skeletal muscle mass and function.
Unveiling the role and control of Fis1 in skeletal muscle atrophy
Within yeast, Fis1 acts as the exclusive receptor for Drp1 located on the outer mitochondrial membrane, facilitating the process of mitochondrial division by increasing the cytoplasmic protein Dnm1 (Dynamin1) [76]. In mammals, Fis1 is the most studied but not the only participant in mitochondrial fission. Rather, receptors such as MFF, Mid49, and Mid51 have emerged, suggesting the responsibility of ushering Drp1 to the outer mitochondrial membrane [77, 78]. Individual aging and long-term sedentary behavior can induce a mitochondrial fission phenotype characterized by increased Fis1 expression, accompanied by muscle mass loss [79, 80]. Several human studies have demonstrated that aerobic exercise training can remodel skeletal muscle mitochondrial fission in aging and sedentary populations, promoting the development of mitochondria into a more fused tubular network structure [79, 81]. This remodeling helps maintain higher mitochondrial oxidation capacity and substrate utilization. However, it is worth noting that other forms of exercise, such as resistance exercise, may not induce similar remodeling of mitochondrial networks [82]. In the context of fasting-induced muscle atrophy, temporarily inhibiting Fis1 represents a promising strategy to impede disease progression [30]. On the other hand, acute deletion of Fis1 in skeletal muscle has detrimental effects. This action typically triggers excessive mitochondrial fusion, resulting in ultrastructural anomalies, heightened ROS accumulation within mitochondria, reduced mitochondrial respiration, and an aggregation of polyubiquitinated proteins in muscle fibers. Ultimately, this cascade leads to muscle atrophy, degeneration, diminished mobility, and a shortened lifespan [32, 33]. Therefore, the precise regulation of Fis1 is crucial for maintaining skeletal muscle health and preventing pathological conditions associated with muscle atrophy.
Balancing mitochondrial dynamics: a feasible strategy against skeletal muscle atrophy
Abnormal mitochondrial fusion and fission are common features of both primary and secondary skeletal muscle atrophy. This not only highlights the importance of maintaining normal mitochondrial dynamics for skeletal muscle health but also suggests that the restoration of balanced mitochondrial dynamics may reverse the onset of skeletal muscle atrophy. For example, patients with ALS often exhibit excessive activation of mitochondrial fission by Drp1, resulting in massive accumulation of fragmented mitochondria, muscle atrophy and paralysis. However, studies using an exogenous inhibitor called P110, which inhibits the interaction between Drp1 and Fis1, have shown promising results. Treatment of mice with P110 significantly improved locomotion and improved the amount of healthier muscle tissue [83]. Similarly, acute deletion of OPA1 in adults leads to impaired mitochondrial fusion and increased fission, accompanied by increased FGF21 secretion in the muscle, resulting in severe skeletal muscle atrophy [84]. Mechanistically, OPA1 deficiency leads to ER stress and mitochondrial DNA release, which triggers the upregulation of the transcription factor FGF21. Moreover, ER stress triggers skeletal muscle degradation through the unfolded protein response (UPR) and the FoxO signaling pathway [56]. Studies in mice have shown that blocking excessive mitochondrial fission by downregulating the expression of Drp1 can effectively reduce the accumulation of the muscle factor FGF21 and largely alleviate skeletal muscle atrophy caused by OPA1 deficiency [85]. Obviously, it is feasible to treat skeletal muscle atrophy by targeting mitochondrial dynamics.
In addition, the expression level of the muscle factor FGF21 is usually positively correlated with unhealthy aging and muscle atrophy [86]. Transient inhibition or overexpression of FGF21 has been found to exert a health-promoting effect on skeletal muscle and enhance its adaptation to stress responses induced by perturbations in mitochondrial dynamics [87]. For example, in mouse and C2C12 cell I/RI (ischemia‒reperfusion injury) models, overexpression of FGF21 has been shown to inhibit the activation of Drp1. This inhibition of Drp1 activation by FGF21 protects mitochondria from excessive fission and contributes to the mitigation of histological damage in muscle, improvements in serum indices, and a reduction in apoptotic cell numbers in mice [87]. Furthermore, deletion of the muscle factor FGF21 has been shown to protect against starvation‐induced muscle atrophy and weakness [88]. These results indicate that in response to the detrimental effects of imbalanced mitochondrial fusion and fission on skeletal muscle, the body activates compensatory mechanisms through coordinated pathways. This compensatory response aims to restore dysregulated mitochondrial dynamics and maintain muscle health. Therefore, it is crucial to restore mitochondrial dynamics within appropriate limits in atrophic skeletal muscle to promote muscle health and counteract the progression of muscle atrophy.
The role of mitophagy in skeletal muscle atrophy
Overview of the mitophagy pathway in skeletal muscle
Mitophagy, a particular autophagic process mediated by the PINK1–parkin pathway that selectively eliminates extra or aberrant mitochondria, is a quality control mechanism for the removal of damaged mitochondria. PTEN-induced kinase 1 (PINK1) is a Ser/Thr kinase located in the outer mitochondrial membrane whose expression is induced by phosphatase and tens of homologs (PTEN). Parkin is a protein that is encoded by Park2 and has E3 ubiquitin protein ligase activity. Multiple studies have demonstrated that Parkin plays a protective role in maintaining skeletal muscle contraction and healthy mitochondrial function [89]. Skeletal muscle atrophy, including atrophy induced by fasting and aging, is exacerbated in the absence of Parkin or when its mitochondrial localization is reduced [90, 91]. Under normal physiological conditions, PINK1 is sequentially targeted by the mitochondrial outer membrane protein outer membrane translocase (TOM) from the mitochondrial outer membrane to the mitochondrial inner membrane and is degraded by the mitochondrial matrix processing peptidase MPP. It is then cleaved by the protease PARL in the mitochondrial inner membrane, resulting in the dissociation of PINK1 from TOM [92, 93]. Cleaved PINK1 is transferred to the cytoplasm, preventing the recruitment and activation of Parkin on the mitochondrial outer membrane (OMM), thereby inhibiting the onset of mitosis. Conversely, when mitochondria in skeletal muscle sustain damage, such as a loss of membrane potential or an excessive accumulation of ROS, the normal entry of PINK1 into the IMM is hindered. Under TOM action, PINK1 accumulates on the OMM as a dimer, where it recruits and activates Parkin and ultimately triggers mitophagy [94]. The underlying molecular mechanism involves the accumulation and dimerization of PINK1 on the OMM, which triggers autophosphorylation of Ser228 and Ser402, which is essential for the recruitment of Parkin by PINK1 [95, 96]. Once parkin is recruited to the OMM, PINK1 phosphorylates parkin at Ser65, resulting in its partial activation. Subsequently, parkin is fully activated by the addition of ubiquitin, which in turn is phosphorylated by PINK1 [97]. Mitochondrial outer membrane proteins, including VDAC1, MFN1/2, FIS1, etc., are subsequently ubiquitinated by activated parkin (parkin-interacting proteins are available in the IntAct database at https://www.ebi.ac.uk/intact). The ubiquitinated protein products, arising from parkin-mediated ubiquitination, are subsequently recruited to autophagosomes through interactions with members of the ATG8 protein family, such as LC3-II or GABARAP. These autophagosomes contain ubiquitinated cargo and then undergo fusion with lysosomes to form autophagic lysosomes. Within the autophagic lysosomes, the contents, including mitochondria exhibiting impaired ubiquitination or abnormal function, are efficiently cleared by various lysosomal enzymes [98, 99] (Fig. 4).

The mechanism of mitophagy under normal and abnormal physiological conditions. Under normal conditions, PINK1-mediated mitochondrial targeting and degradation inhibit parkin recruitment, thus preventing mitophagy initiation. In contrast, within damaged muscle mitochondria, PINK1 accumulation on the outer membrane triggers parkin recruitment and activation, initiating mitophagy. This cascade involves PINK1 accumulation, dimerization, and phosphorylation, which are crucial for parkin recruitment. Upon recruitment, PINK1 phosphorylates parkin at Ser65, activating it and promoting subsequent ubiquitin release
As mentioned above, while the PINK1–parkin pathway plays a significant role, it is not the exclusive mechanism responsible for mediating mitophagy. Other pathways are also involved in the process of mitophagy. Mitophagy can also be initiated through receptor-dependent pathways, which are characterized by the absence of parkin-mediated ubiquitin signaling [100]. For instance, various PINK1–parkin-independent receptors, including BNIP3, BNIP3L/NIX, FUNDC1, BCL2L13, PHB2, ATAD3B, and SPATA33, have been identified. These receptors are activated in response to different external stress conditions and bind to other E3 ubiquitin ligases. Then, LC3 or GABARAP proteins are recruited to connect mitochondria and autophagosomes, thereby triggering mitochondrial autophagy (these receptor proteins are listed in Table 1). This process is similar to that of the Atg32 transcription factor in yeast [101]. These proteins often possess a highly conserved LIR domain that facilitates their interaction with LC3 or GABARAP proteins [102]. Consequently, these proteins facilitate the recruitment of LC3 or GABARAP proteins, thereby establishing connections between mitochondria and autophagosomes. For example, recent studies in skeletal muscle have shown that AMBRA1 (autophagy and beclin 1 regulator 1) relies on the E3 ubiquitin ligase HUWE1 (HECT, UBA, and WWE domain containing E3 ubiquitin protein ligase 1) to induce mitophagy, and specific knockout of AMBRA1 in mouse skeletal muscle can seriously impair skeletal muscle mass and mitophagy [103, 104]. Consequently, it is imperative to expand our understanding of mitophagy beyond the PINK1–parkin pathway and consider the involvement of receptor-dependent pathway-related proteins.
Mitophagy: a potential therapeutic target for muscle atrophy
In the context of muscle physiology, mitophagy plays a crucial role in the degradation and recycling of damaged or aged mitochondria. Mitochondrial dysfunction caused by blocked mitophagy is usually considered to be one of the main factors causing skeletal muscle atrophy [120]. Given this perspective, an increasing number of studies have focused on the mechanism of mitophagy in the process of muscle atrophy and suggested that preventing and treating muscle atrophy may be useful (Table 2). These studies aimed to enhance the understanding of the intricate processes involved and provide insights into potential therapeutic strategies for the prevention and treatment of muscular dystrophy.
Pharmacological modulation of mitophagy in preventing and treating skeletal muscle atrophy
Pharmacological modulation of mitophagy has emerged as a promising approach for enhancing muscle function and preventing muscle atrophy. Notably, in the context of age-related skeletal muscle atrophy, the significant accumulation of ROS in mitochondria regulates the activity of mitophagy-related proteins through oxidative modification, accompanied by oxidative damage and decreased mitochondrial membrane potential, resulting in excessive activation of mitophagy and damage to muscle tissue [133]. To address this issue, researchers have investigated pharmacological interventions aimed at rebalancing mitophagy and alleviating oxidative stress to preserve muscle integrity and function. For instance, apigenin, a natural flavonoid with potent antioxidant properties, has shown promise in alleviating age-related skeletal muscle atrophy. By reducing oxidative stress and inhibiting excessive mitophagy and apoptosis, apigenin contributes to the preservation of muscle mass and function in aging mice [122]. It is commonly observed that aging is linked to excessive mitophagy, which may result in muscle dysfunction. Theoretically, age-related skeletal muscle atrophy can be mitigated by reducing oxidative stress and inhibiting mitophagy and apoptosis [80]. However, contrasting findings have emerged from several studies, indicating that skeletal muscle mitophagy is impaired during aging. Impaired mitophagy can predictably lead to the accumulation of damaged mitochondria in vivo, disruption of the mitochondrial network, and initiation of the apoptotic program, which together lead to skeletal muscle atrophy [134, 135]. Under this physiological condition, several studies have demonstrated that urolithin A [123, 136] and tomatidine [124] can enhance muscle function by inducing mitophagy in age-related skeletal muscle atrophy. More notably, a recent study definitively highlighted that there was no discernible difference in the level of mitophagy observed in the muscles of young and aged mice [137]. These conflicting conclusions regarding mitophagy in skeletal muscle underscore the complexity of this process (Table 3). Thus, we summarize several factors contributing to the conflicting conclusions regarding mitophagy in aging skeletal muscle. First, different studies employ different subjects, experimental conditions, and technical methods, which can lead to variations in the results. Second, skeletal muscle atrophy is a prolonged process, and mitophagy may exert varying effects at distinct stages of the atrophy process. Finally, sex differences have been shown to influence mitophagy regulation [138].
Furthermore, in cases of exogenous skeletal muscle atrophy, neurological skeletal muscle atrophy caused by nervous system injury or disease is common. Oxidative stress and inflammatory responses are typically considered early events triggering disease onset. In the treatment of neurogenic skeletal muscle atrophy, drug interventions such as antioxidants or antiinflammatories can significantly mitigate abnormal mitophagy and decelerate the progression of skeletal muscle atrophy. For instance, a range of antioxidant and antiinflammatory medications, including isoquercitrin [139], celecoxib [127], and salidroside [126], have been demonstrated to be able to impede the inflammatory response and oxidative stress in denervated skeletal muscle. Mechanistically, these medications can hinder mitophagy and proteolysis, enhance blood circulation in the targeted muscles, and ultimately ameliorate muscle atrophy resulting from denervation.
Stabilizing mitophagy through targeted exercise: a nondrug approach to treating skeletal muscle atrophy
In recent years, exercise therapy has also become an efficient nondrug therapy for the treatment of skeletal muscle atrophy. Numerous studies have demonstrated that targeted exercise and training can enhance mitophagy, the process of removing damaged mitochondria, thereby countering muscle atrophy caused by muscle damage or metabolic disorders. This effect is achieved through the activation of various nuclear proteins in the body, including those involved in the PINK1–parkin pathway and other related pathways. Muscle use and exercise are known to stimulate myogenesis and muscle growth. Myogenesis, driven by myoblast myogenic differentiation, is a pivotal process in skeletal muscle development and the maintenance of homeostasis. Mitophagy plays an indispensable role in facilitating this process [174]. Conversely, prolonged lack of exercise results in a higher rate of muscle protein breakdown than synthesis, leading to reduced skeletal muscle load and consequently accelerating muscle atrophy [175]. Interestingly, diverse types of exercise modulate mitophagy through distinct signaling or regulatory pathways to deter skeletal muscle atrophy. For instance, resistance exercise can prevent and mitigate skeletal muscle atrophy by triggering AMPK-mediated mitophagy and preserving the health of the mitochondrial network in skeletal muscle. Mechanistically, it downregulates the expression of E3 ubiquitin ligases, such as atrogin-1 and MuRF1, in skeletal muscle through the AMPK/FoxO3 signaling pathway and reduces skeletal muscle atrophy caused by the inhibition of protein synthesis and imbalance of ubiquitination/degradation [128, 176]. Furthermore, it has been demonstrated that aerobic/endurance exercise can mitigate skeletal muscle atrophy by promoting the mitochondrial localization of mitophagy biomarkers (including PINK1 and parkin) and the conversion of key autophagy proteins (from LC3-I to LC3-II) [128, 177]. Further evidence suggests that the enhancement of mitophagy flux in skeletal muscle induced by endurance exercise relies on parkin [178]. Different forms of exercise target the restoration of mitochondrial content and function to physiological norms by modulating mitophagy, thus mitigating the buildup of dysfunctional mitochondria and halting the advancement of skeletal muscle atrophy [176, 179].
Indeed, despite multiple studies examining the therapeutic effects of mitophagy on skeletal muscle atrophy induced by various causes, inconsistencies in the data persist. These conflicting results highlight the complexity of mitophagy regulation and its role in skeletal muscle atrophy, which may vary depending on the specific conditions, models, and factors under investigation. Further research is necessary to unravel the specific underlying mechanisms involved and clarify the impact of mitophagy in the context of skeletal muscle atrophy (Fig. 5).

Schematic representation of potential therapeutic targets for skeletal muscle atrophy: mitophagy. Pharmacological modulation and exercise therapy are proposed as interventions to prevent and delay skeletal muscle atrophy. These drugs aim to restore physiological mitophagy levels and reduce the accumulation of damaged mitochondria in the body
Conclusions
Currently, skeletal muscle atrophy is a considerable global health concern, as evidenced by notable increases in both the prevalence and mortality rates of affected individuals. Skeletal muscle is the body’s most extensive metabolic organ. The significance of mitochondrial homeostasis in supporting muscle activity and preserving muscle health is increasingly acknowledged. Key mechanisms of mitochondrial quality control, such as mitochondrial dynamics and mitophagy, play a critical role in maintaining metabolic balance and maintaining mitochondrial health in skeletal muscle. The interaction between mitochondrial dynamics and mitophagy has not been fully explored, and a comprehensive understanding of the complete mechanism governing skeletal muscle atrophy has yet to be obtained. In this review, we focus on the effects of changes in mitochondrial dynamics and mitophagy at the basic physiological level on skeletal muscle health and summarize the reasons for these conflicting data. Crucially, we discussed the potential of targeted regulation of and mitophagy as a viable strategy for treating skeletal muscle atrophy. In the future, leveraging mitochondrial dynamics and mitophagy to alleviate skeletal muscle atrophy will undoubtedly become a feasible and promising strategy.
Availability of data and materials
No data was used for the research described in the article.
Abbreviations
- OXPHOS:
-
Oxidative phosphorylation
- ROS:
-
Reactive oxygen species
- FoxO3:
-
Forkhead box O3
- MuRF1:
-
Muscle-specific ring finger protein 1
- Atrogin-1:
-
Muscle atrophy F-box protein
- DRP1:
-
Dynamin 1 like
- Fis1:
-
Fission, mitochondrial 1
- LIR:
-
LC3 interaction region
- MFN1:
-
Mitofusin 1
- MFN2:
-
Mitofusin 2
- HIF-1α:
-
Hypoxia inducible factor 1 subunit alpha
- BNIP3:
-
BCL2 interacting protein 3
- PINK1:
-
PTEN induced kinase 1
- OPA1:
-
Optic atrophy 1
- OMM:
-
Outer mitochondrial membrane
- IMM:
-
Inner mitochondrial membrane
- CMT2A:
-
Charcot–Marie–Tooth type 2A
- ER:
-
Endoplasmic reticulum
- Ca2+:
-
Calcium ions
- mtDNA:
-
Mitochondrial DNA
- FGF21:
-
Fibroblast growth factor 21
- MID49:
-
Mitochondrial elongation factor 2
- MID51:
-
Mitochondrial elongation factor 1
- ALS:
-
Amyotrophic lateral sclerosis
References
Baskin KK, Winders BR, Olson EN. Muscle as a “mediator” of systemic metabolism. Cell Metab. 2015;21(2):237–48.
Baehr LM, Hughes DC, Waddell DS, Bodine SC. SnapShot: skeletal muscle atrophy. Cell. 2022;185(9):1618-1618.e1.
Anker SD, Morley JE, von Haehling S. Welcome to the ICD-10 code for sarcopenia. J Cachexia Sarcopenia Muscle. 2016;7(5):512–4.
Riuzzi F, Sorci G, Arcuri C, Giambanco I, Bellezza I, Minelli A, et al. Cellular and molecular mechanisms of sarcopenia: the S100B perspective. J Cachexia Sarcopenia Muscle. 2018;9(7):1255–68.
Yang X, Xue P, Liu Z, Li W, Li C, Chen Z. SESN2 prevents the slow-to-fast myofiber shift in denervated atrophy via AMPK/PGC-1α pathway. Cell Mol Biol Lett. 2022;27(1):66.
Ciciliot S, Rossi AC, Dyar KA, Blaauw B, Schiaffino S. Muscle type and fiber type specificity in muscle wasting. Int J Biochem Cell Biol. 2013;45(10):2191–9.
Argilés JM, Campos N, Lopez-Pedrosa JM, Rueda R, Rodriguez-Mañas L. Skeletal muscle regulates metabolism via Interorgan crosstalk: roles in health and disease. J Am Med Dir Assoc. 2016;17(9):789–96.
Landi F, Calvani R, Martone AM, Salini S, Zazzara MB, Candeloro M, et al. Normative values of muscle strength across ages in a “real world” population: results from the longevity check-up 7+ project. J Cachexia Sarcopenia Muscle. 2020;11(6):1562–9.
Vincent AE, White K, Davey T, Philips J, Ogden RT, Lawless C, et al. Quantitative 3D mapping of the human skeletal muscle mitochondrial network. Cell Rep. 2019;26(4):996-1009.e4.
Yin L, Li N, Jia W, Wang N, Liang M, Yang X, et al. Skeletal muscle atrophy: from mechanisms to treatments. Pharmacol Res. 2021;172: 105807.
Sieber CC. Malnutrition and sarcopenia. Aging Clin Exp Res. 2019;31(6):793–8.
Cochet C, Belloni G, Buondonno I, Chiara F, D’Amelio P. The role of nutrition in the treatment of sarcopenia in old patients: from restoration of mitochondrial activity to improvement of muscle performance, a systematic review. Nutrients. 2023;15(17):3703.
Kubat GB, Bouhamida E, Ulger O, Turkel I, Pedriali G, Ramaccini D, et al. Mitochondrial dysfunction and skeletal muscle atrophy: causes, mechanisms, and treatment strategies. Mitochondrion. 2023;72:33–58.
Nunnari J, Suomalainen A. Mitochondria: in sickness and in health. Cell. 2012;148(6):1145–59.
Kang C, Yeo D, Ji LL. Muscle immobilization activates mitophagy and disrupts mitochondrial dynamics in mice. Acta Physiol (Oxf). 2016;218(3):188–97.
Abate M, Festa A, Falco M, Lombardi A, Luce A, Grimaldi A, et al. Mitochondria as playmakers of apoptosis, autophagy and senescence. Semin Cell Dev Biol. 2020;98:139–53.
Hood DA, Memme JM, Oliveira AN, Triolo M. Maintenance of skeletal muscle mitochondria in health, exercise, and aging. Annu Rev Physiol. 2019;81:19–41.
Hong X, Isern J, Campanario S, Perdiguero E, Ramírez-Pardo I, Segalés J, et al. Mitochondrial dynamics maintain muscle stem cell regenerative competence throughout adult life by regulating metabolism and mitophagy. Cell Stem Cell. 2022;29(9):1298-1314.e10.
Pfanner N, Warscheid B, Wiedemann N. Mitochondrial proteins: from biogenesis to functional networks. Nat Rev Mol Cell Biol. 2019;20(5):267–84.
Song J, Herrmann JM, Becker T. Quality control of the mitochondrial proteome. Nat Rev Mol Cell Biol. 2021;22(1):54–70.
Roca-Portoles A, Tait SWG. Mitochondrial quality control: from molecule to organelle. Cell Mol Life Sci. 2021;78(8):3853–66.
Lian D, Chen MM, Wu H, Deng S, Hu X. The role of oxidative stress in skeletal muscle myogenesis and muscle disease. Antioxidants. 2022;11(4):755.
Sandri M, Sandri C, Gilbert A, Skurk C, Calabria E, Picard A, et al. Foxo transcription factors induce the atrophy-related ubiquitin ligase atrogin-1 and cause skeletal muscle atrophy. Cell. 2004;117(3):399–412.
Zhang H, Qi G, Wang K, Yang J, Shen Y, Yang X, et al. Oxidative stress: roles in skeletal muscle atrophy. Biochem Pharmacol. 2023;214: 115664.
Yang X, Xue P, Chen H, Yuan M, Kang Y, Duscher D, et al. Denervation drives skeletal muscle atrophy and induces mitochondrial dysfunction, mitophagy and apoptosis via miR-142a-5p/MFN1 axis. Theranostics. 2020;10(3):1415–32.
Hu Z, Klein JD, Mitch WE, Zhang L, Martinez I, Wang XH. MicroRNA-29 induces cellular senescence in aging muscle through multiple signaling pathways. Aging. 2014;6(3):160–75.
Goljanek-Whysall K, Soriano-Arroquia A, McCormick R, Chinda C, McDonagh B. miR-181a regulates p62/SQSTM1, parkin, and protein DJ-1 promoting mitochondrial dynamics in skeletal muscle aging. Aging Cell. 2020;19(4): e13140.
Alessio E, Buson L, Chemello F, Peggion C, Grespi F, Martini P, et al. Single cell analysis reveals the involvement of the long non-coding RNA Pvt1 in the modulation of muscle atrophy and mitochondrial network. Nucleic Acids Res. 2019;47(4):1653–70.
Wai T, Langer T. Mitochondrial dynamics and metabolic regulation. Trends Endocrinol Metab. 2016;27(2):105–17.
Romanello V, Guadagnin E, Gomes L, Roder I, Sandri C, Petersen Y, et al. Mitochondrial fission and remodelling contributes to muscle atrophy. EMBO J. 2010;29(10):1774–85.
Green DR, Van Houten B. SnapShot: mitochondrial quality control. Cell. 2011;147(4):950.
Lee TT, Chen PL, Su MP, Li JC, Chang YW, Liu RW, et al. Loss of Fis1 impairs proteostasis during skeletal muscle aging in Drosophila. Aging Cell. 2021;20(6): e13379.
Zhang Z, Sliter DA, Bleck CKE, Ding S. Fis1 deficiencies differentially affect mitochondrial quality in skeletal muscle. Mitochondrion. 2019;49:217–26.
Xian H, Yang Q, Xiao L, Shen HM, Liou YC. STX17 dynamically regulated by Fis1 induces mitophagy via hierarchical macroautophagic mechanism. Nat Commun. 2019;10(1):2059.
Sebastián D, Sorianello E, Segalés J, Irazoki A, Ruiz-Bonilla V, Sala D, et al. Mfn2 deficiency links age-related sarcopenia and impaired autophagy to activation of an adaptive mitophagy pathway. EMBO J. 2016;35(15):1677–93.
Liang J, Zhang H, Zeng Z, Wu L, Zhang Y, Guo Y, et al. Lifelong aerobic exercise alleviates sarcopenia by activating autophagy and inhibiting protein degradation via the AMPK/PGC-1α signaling pathway. Metabolites. 2021;11(5):323.
Han H, Tan J, Wang R, Wan H, He Y, Yan X, et al. PINK1 phosphorylates Drp1S616 to regulate mitophagy-independent mitochondrial dynamics. EMBO Rep. 2020;21(8): e48686.
Liu H, Ho PWL, Leung CT, Pang SYY, Chang EES, Choi ZYK, et al. Aberrant mitochondrial morphology and function associated with impaired mitophagy and DNM1L-MAPK/ERK signaling are found in aged mutant Parkinsonian LRRK2R1441G mice. Autophagy. 2021;17(10):3196–220.
Chen Y, Dorn GW. PINK1-phosphorylated mitofusin 2 is a Parkin receptor for culling damaged mitochondria. Science. 2013;340(6131):471–5.
Chen M, Chen Z, Wang Y, Tan Z, Zhu C, Li Y, et al. Mitophagy receptor FUNDC1 regulates mitochondrial dynamics and mitophagy. Autophagy. 2016;12(4):689–702.
Giacomello M, Pyakurel A, Glytsou C, Scorrano L. The cell biology of mitochondrial membrane dynamics. Nat Rev Mol Cell Biol. 2020;21(4):204–24.
Chan DC. Mitochondrial dynamics and its involvement in disease. Annu Rev Pathol. 2020;15:235–59.
Chan DC. Fusion and fission: interlinked processes critical for mitochondrial health. Annu Rev Genet. 2012;46:265–87.
Toyama EQ, Herzig S, Courchet J, Lewis TL, Losón OC, Hellberg K, et al. Metabolism. AMP-activated protein kinase mediates mitochondrial fission in response to energy stress. Science. 2016;351(6270):275–81.
Westermann B. Bioenergetic role of mitochondrial fusion and fission. Biochim Biophys Acta. 2012;1817(10):1833–8.
Jimah JR, Hinshaw JE. Structural insights into the mechanism of dynamin superfamily proteins. Trends Cell Biol. 2019;29(3):257–73.
Favaro G, Romanello V, Varanita T, Andrea Desbats M, Morbidoni V, Tezze C, et al. DRP1-mediated mitochondrial shape controls calcium homeostasis and muscle mass. Nat Commun. 2019;10(1):2576.
Huang DD, Fan SD, Chen XY, Yan XL, Zhang XZ, Ma BW, et al. Nrf2 deficiency exacerbates frailty and sarcopenia by impairing skeletal muscle mitochondrial biogenesis and dynamics in an age-dependent manner. Exp Gerontol. 2019;119:61–73.
Mao X, Gu Y, Sui X, Shen L, Han J, Wang H, et al. Phosphorylation of dynamin-related protein 1 (DRP1) regulates mitochondrial dynamics and skeletal muscle wasting in cancer cachexia. Front Cell Dev Biol. 2021;9: 673618.
Gao S, Hu J. Mitochondrial fusion: the machineries in and out. Trends Cell Biol. 2021;31(1):62–74.
Romanello V, Sandri M. The connection between the dynamic remodeling of the mitochondrial network and the regulation of muscle mass. Cell Mol Life Sci. 2021;78(4):1305–28.
Eisner V, Lenaers G, Hajnóczky G. Mitochondrial fusion is frequent in skeletal muscle and supports excitation-contraction coupling. J Cell Biol. 2014;205(2):179–95.
Mishra P, Varuzhanyan G, Pham AH, Chan DC. Mitochondrial dynamics is a distinguishing feature of skeletal muscle fiber types and regulates organellar compartmentalization. Cell Metab. 2015;22(6):1033–44.
Lang F, Khaghani S, Türk C, Wiederstein JL, Hölper S, Piller T, et al. Single muscle fiber proteomics reveals distinct protein changes in slow and fast fibers during muscle atrophy. J Proteome Res. 2018;17(10):3333–47.
Li M, Wang L, Wang Y, Zhang S, Zhou G, Lieshout R, et al. Mitochondrial fusion via OPA1 and MFN1 supports liver tumor cell metabolism and growth. Cells. 2020;9(1):E121.
Tezze C, Romanello V, Desbats MA, Fadini GP, Albiero M, Favaro G, et al. Age-associated loss of OPA1 in muscle impacts muscle mass, metabolic homeostasis, systemic inflammation, and epithelial senescence. Cell Metab. 2017;25(6):1374-1389.e6.
Xi QL, Zhang B, Jiang Y, Zhang HS, Meng QY, Chen Y, et al. Mitofusin-2 prevents skeletal muscle wasting in cancer cachexia. Oncol Lett. 2016;12(5):4013–20.
Chen H, Detmer SA, Ewald AJ, Griffin EE, Fraser SE, Chan DC. Mitofusins Mfn1 and Mfn2 coordinately regulate mitochondrial fusion and are essential for embryonic development. J Cell Biol. 2003;160(2):189–200.
Wang L, Gao J, Liu J, Siedlak SL, Torres S, Fujioka H, et al. Mitofusin 2 regulates axonal transport of calpastatin to prevent neuromuscular synaptic elimination in skeletal muscles. Cell Metab. 2018;28(3):400.
Cefis M, Dargegen M, Marcangeli V, Taherkhani S, Dulac M, Leduc-Gaudet JP, et al. MFN2 overexpression in skeletal muscles of young and old mice causes a mild hypertrophy without altering mitochondrial respiration and H2 O2 emission. Acta Physiol. 2024. https://doi.org/10.1111/apha.14119.
Li YJ, Cao YL, Feng JX, Qi Y, Meng S, Yang JF, et al. Structural insights of human mitofusin-2 into mitochondrial fusion and CMT2A onset. Nat Commun. 2019;10(1):4914.
de Brito OM, Scorrano L. Mitofusin 2 tethers endoplasmic reticulum to mitochondria. Nature. 2008;456(7222):605–10.
Varanita T, Soriano ME, Romanello V, Zaglia T, Quintana-Cabrera R, Semenzato M, et al. The OPA1-dependent mitochondrial cristae remodeling pathway controls atrophic, apoptotic, and ischemic tissue damage. Cell Metab. 2015;21(6):834–44.
Kleele T, Rey T, Winter J, Zaganelli S, Mahecic D, Perreten Lambert H, et al. Distinct fission signatures predict mitochondrial degradation or biogenesis. Nature. 2021;593(7859):435–9.
Lewis SC, Uchiyama LF, Nunnari J. ER-mitochondria contacts couple mtDNA synthesis with mitochondrial division in human cells. Science. 2016;353(6296): aaf5549.
Tilokani L, Nagashima S, Paupe V, Prudent J. Mitochondrial dynamics: overview of molecular mechanisms. Essays Biochem. 2018;62(3):341–60.
Lee JE, Westrate LM, Wu H, Page C, Voeltz GK. Multiple dynamin family members collaborate to drive mitochondrial division. Nature. 2016;540(7631):139–43.
Kalia R, Wang RYR, Yusuf A, Thomas PV, Agard DA, Shaw JM, et al. Structural basis of mitochondrial receptor binding and constriction by DRP1. Nature. 2018;558(7710):401–5.
Ma R, Ma L, Weng W, Wang Y, Liu H, Guo R, et al. DUSP6 SUMOylation protects cells from oxidative damage via direct regulation of Drp1 dephosphorylation. Sci Adv. 2020;6(13): eaaz0361.
Nagashima S, Tábara LC, Tilokani L, Paupe V, Anand H, Pogson JH, et al. Golgi-derived PI(4)P-containing vesicles drive late steps of mitochondrial division. Science. 2020;367(6484):1366–71.
Dulac M, Leduc-Gaudet JP, Reynaud O, Ayoub MB, Guérin A, Finkelchtein M, et al. Drp1 knockdown induces severe muscle atrophy and remodelling, mitochondrial dysfunction, autophagy impairment and denervation. J Physiol. 2020;598(17):3691–710.
Rana A, Oliveira MP, Khamoui AV, Aparicio R, Rera M, Rossiter HB, et al. Promoting Drp1-mediated mitochondrial fission in midlife prolongs healthy lifespan of Drosophila melanogaster. Nat Commun. 2017;8(1):448.
Dulac M, Leduc-Gaudet JP, Cefis M, Ayoub MB, Reynaud O, Shams A, et al. Regulation of muscle and mitochondrial health by the mitochondrial fission protein Drp1 in aged mice. J Physiol. 2021;599(17):4045–63.
Qi Z, Huang Z, Xie F, Chen L. Dynamin-related protein 1: a critical protein in the pathogenesis of neural system dysfunctions and neurodegenerative diseases. J Cell Physiol. 2019;234(7):10032–46.
Petrozziello T, Bordt EA, Mills AN, Kim SE, Sapp E, Devlin BA, et al. Targeting tau mitigates mitochondrial fragmentation and oxidative stress in amyotrophic lateral sclerosis. Mol Neurobiol. 2022;59(1):683–702.
Mozdy AD, McCaffery JM, Shaw JM. Dnm1p GTPase-mediated mitochondrial fission is a multi-step process requiring the novel integral membrane component Fis1p. J Cell Biol. 2000;151(2):367–80.
Otera H, Wang C, Cleland MM, Setoguchi K, Yokota S, Youle RJ, et al. Mff is an essential factor for mitochondrial recruitment of Drp1 during mitochondrial fission in mammalian cells. J Cell Biol. 2010;191(6):1141–58.
Yu R, Liu T, Jin SB, Ning C, Lendahl U, Nistér M, et al. MIEF1/2 function as adaptors to recruit Drp1 to mitochondria and regulate the association of Drp1 with Mff. Sci Rep. 2017;7(1):880.
Axelrod CL, Fealy CE, Mulya A, Kirwan JP. Exercise training remodels human skeletal muscle mitochondrial fission and fusion machinery towards a pro-elongation phenotype. Acta Physiol (Oxf). 2019;225(4): e13216.
Yeo D, Kang C, Gomez-Cabrera MC, Vina J, Ji LL. Intensified mitophagy in skeletal muscle with aging is downregulated by PGC-1alpha overexpression in vivo. Free Radic Biol Med. 2019;130:361–8.
Ruegsegger GN, Pataky MW, Simha S, Robinson MM, Klaus KA, Nair KS. High-intensity aerobic, but not resistance or combined, exercise training improves both cardiometabolic health and skeletal muscle mitochondrial dynamics. J Appl Physiol. 2023;135(4):763–74.
Marshall RN, McKendry J, Smeuninx B, Seabright AP, Morgan PT, Greig C, et al. Acute resistance exercise training does not augment mitochondrial remodelling in master athletes or untrained older adults. Front Physiol. 2022;13:1097988.
Joshi AU, Saw NL, Vogel H, Cunnigham AD, Shamloo M, Mochly-Rosen D. Inhibition of Drp1/Fis1 interaction slows progression of amyotrophic lateral sclerosis. EMBO Mol Med. 2018;10(3): e8166.
Pereira RO, Tadinada SM, Zasadny FM, Oliveira KJ, Pires KMP, Olvera A, et al. OPA1 deficiency promotes secretion of FGF21 from muscle that prevents obesity and insulin resistance. EMBO J. 2017;36(14):2126–45.
Romanello V, Scalabrin M, Albiero M, Blaauw B, Scorrano L, Sandri M. Inhibition of the fission machinery mitigates OPA1 impairment in adult skeletal muscles. Cells. 2019;8(6):597.
Roh E, Hwang SY, Yoo HJ, Baik SH, Cho B, Park YS, et al. Association of plasma FGF21 levels with muscle mass and muscle strength in a national multicentre cohort study: Korean Frailty and Aging Cohort Study. Age Ageing. 2021;50(6):1971–8.
Li B, Liu L. Fibroblast growth factor 21, a stress regulator, inhibits Drp1 activation to alleviate skeletal muscle ischemia/reperfusion injury. Lab Invest. 2022;102(9):979–88.
Oost LJ, Kustermann M, Armani A, Blaauw B, Romanello V. Fibroblast growth factor 21 controls mitophagy and muscle mass. J Cachexia Sarcopenia Muscle. 2019;10(3):630–42.
Gouspillou G, Godin R, Piquereau J, Picard M, Mofarrahi M, Mathew J, et al. Protective role of Parkin in skeletal muscle contractile and mitochondrial function. J Physiol. 2018;596(13):2565–79.
Peker N, Sharma M, Kambadur R. Parkin deficiency exacerbates fasting-induced skeletal muscle wasting in mice. NPJ Parkinsons Dis. 2022;8(1):159.
Chen CCW, Erlich AT, Crilly MJ, Hood DA. Parkin is required for exercise-induced mitophagy in muscle: impact of aging. Am J Physiol Endocrinol Metab. 2018;315(3):E404–15.
Meissner C, Lorenz H, Weihofen A, Selkoe DJ, Lemberg MK. The mitochondrial intramembrane protease PARL cleaves human Pink1 to regulate Pink1 trafficking. J Neurochem. 2011;117(5):856–67.
Deas E, Plun-Favreau H, Gandhi S, Desmond H, Kjaer S, Loh SHY, et al. PINK1 cleavage at position A103 by the mitochondrial protease PARL. Hum Mol Genet. 2011;20(5):867–79.
Nguyen TN, Padman BS, Lazarou M. Deciphering the molecular signals of PINK1/Parkin mitophagy. Trends Cell Biol. 2016;26(10):733–44.
Aerts L, Craessaerts K, De Strooper B, Morais VA. PINK1 kinase catalytic activity is regulated by phosphorylation on serines 228 and 402. J Biol Chem. 2015;290(5):2798–811.
Okatsu K, Uno M, Koyano F, Go E, Kimura M, Oka T, et al. A dimeric PINK1-containing complex on depolarized mitochondria stimulates Parkin recruitment. J Biol Chem. 2013;288(51):36372–84.
Gladkova C, Maslen SL, Skehel JM, Komander D. Mechanism of Parkin activation by PINK1. Nature. 2018;559(7714):410–4.
Lazarou M, Sliter DA, Kane LA, Sarraf SA, Wang C, Burman JL, et al. The ubiquitin kinase PINK1 recruits autophagy receptors to induce mitophagy. Nature. 2015;524(7565):309–14.
Bravo-San Pedro JM, Kroemer G, Galluzzi L. Autophagy and mitophagy in cardiovascular disease. Circ Res. 2017;120(11):1812–24.
Villa E, Marchetti S, Ricci JE. No Parkin zone: mitophagy without Parkin. Trends Cell Biol. 2018;28(11):882–95.
Kanki T, Wang K, Cao Y, Baba M, Klionsky DJ. Atg32 is a mitochondrial protein that confers selectivity during mitophagy. Dev Cell. 2009;17(1):98–109.
Stolz A, Ernst A, Dikic I. Cargo recognition and trafficking in selective autophagy. Nat Cell Biol. 2014;16(6):495–501.
Di Rita A, Peschiaroli A, D’Acunzo P, Strobbe D, Hu Z, Gruber J, et al. HUWE1 E3 ligase promotes PINK1/PARKIN-independent mitophagy by regulating AMBRA1 activation via IKKα. Nat Commun. 2018;9(1):3755.
Gambarotto L, Metti S, Chrisam M, Cerqua C, Sabatelli P, Armani A, et al. Ambra1 deficiency impairs mitophagy in skeletal muscle. J Cachexia Sarcopenia Muscle. 2022;13(4):2211–24.
Li E, Li X, Huang J, Xu C, Liang Q, Ren K, et al. BMAL1 regulates mitochondrial fission and mitophagy through mitochondrial protein BNIP3 and is critical in the development of dilated cardiomyopathy. Protein Cell. 2020;11(9):661–79.
Wu X, Zheng Y, Liu M, Li Y, Ma S, Tang W, et al. BNIP3L/NIX degradation leads to mitophagy deficiency in ischemic brains. Autophagy. 2021;17(8):1934–46.
Liu L, Feng D, Chen G, Chen M, Zheng Q, Song P, et al. Mitochondrial outer-membrane protein FUNDC1 mediates hypoxia-induced mitophagy in mammalian cells. Nat Cell Biol. 2012;14(2):177–85.
Chen Z, Siraj S, Liu L, Chen Q. MARCH5-FUNDC1 axis fine-tunes hypoxia-induced mitophagy. Autophagy. 2017;13(7):1244–5.
Li M, Jia J, Zhang X, Dai H. Selective binding of mitophagy receptor protein Bcl-rambo to LC3/GABARAP family proteins. Biochem Biophys Res Commun. 2020;530(1):292–300.
Murakawa T, Yamaguchi O, Hashimoto A, Hikoso S, Takeda T, Oka T, et al. Bcl-2-like protein 13 is a mammalian Atg32 homologue that mediates mitophagy and mitochondrial fragmentation. Nat Commun. 2015;6:7527.
Shu L, Hu C, Xu M, Yu J, He H, Lin J, et al. ATAD3B is a mitophagy receptor mediating clearance of oxidative stress-induced damaged mitochondrial DNA. EMBO J. 2021;40(8): e106283.
Xu X, Zhang Y, Cheng H, Zhou R. SPATA33 functions as a mitophagy receptor in mammalian germline. Autophagy. 2021;17(5):1284–6.
Yoo SM, Yamashita SI, Kim H, Na D, Lee H, Kim SJ, et al. FKBP8 LIRL-dependent mitochondrial fragmentation facilitates mitophagy under stress conditions. FASEB J. 2020;34(2):2944–57.
Shirane-Kitsuji M, Nakayama KI. Mitochondria: FKBP38 and mitochondrial degradation. Int J Biochem Cell Biol. 2014;51:19–22.
Strappazzon F, Nazio F, Corrado M, Cianfanelli V, Romagnoli A, Fimia GM, et al. AMBRA1 is able to induce mitophagy via LC3 binding, regardless of PARKIN and p62/SQSTM1. Cell Death Differ. 2015;22(3):517.
Wei Y, Chiang WC, Sumpter R, Mishra P, Levine B. Prohibitin 2 is an inner mitochondrial membrane mitophagy receptor. Cell. 2017;168(1–2):224-238.e10.
Yan C, Gong L, Chen L, Xu M, Abou-Hamdan H, Tang M, et al. PHB2 (prohibitin 2) promotes PINK1-PRKN/Parkin-dependent mitophagy by the PARL-PGAM5-PINK1 axis. Autophagy. 2020;16(3):419–34.
Schlattner U, Tokarska-Schlattner M, Epand RM, Boissan M, Lacombe ML, Kagan VE. NME4/nucleoside diphosphate kinase D in cardiolipin signaling and mitophagy. Lab Invest. 2018;98(2):228–32.
Cen X, Chen Y, Xu X, Wu R, He F, Zhao Q, et al. Pharmacological targeting of MCL-1 promotes mitophagy and improves disease pathologies in an Alzheimer’s disease mouse model. Nat Commun. 2020;11(1):5731.
Peker N, Donipadi V, Sharma M, McFarlane C, Kambadur R. Loss of Parkin impairs mitochondrial function and leads to muscle atrophy. Am J Physiol Cell Physiol. 2018;315(2):C164–85.
Ballarò R, Lopalco P, Audrito V, Beltrà M, Pin F, Angelini R, et al. Targeting mitochondria by SS-31 ameliorates the whole body energy status in cancer- and chemotherapy-induced cachexia. Cancers. 2021;13(4):850.
Wang D, Yang Y, Zou X, Zhang J, Zheng Z, Wang Z. Antioxidant apigenin relieves age-related muscle atrophy by inhibiting oxidative stress and hyperactive mitophagy and apoptosis in skeletal muscle of mice. J Gerontol A Biol Sci Med Sci. 2020;75(11):2081–8.
Singh A, D’Amico D, Andreux PA, Fouassier AM, Blanco-Bose W, Evans M, et al. Urolithin A improves muscle strength, exercise performance, and biomarkers of mitochondrial health in a randomized trial in middle-aged adults. Cell Rep Med. 2022;3(5): 100633.
Fang EF, Waltz TB, Kassahun H, Lu Q, Kerr JS, Morevati M, et al. Tomatidine enhances lifespan and healthspan in C. elegans through mitophagy induction via the SKN-1/Nrf2 pathway. Sci Rep. 2017;7:46208.
Mito T, Vincent AE, Faitg J, Taylor RW, Khan NA, McWilliams TG, et al. Mosaic dysfunction of mitophagy in mitochondrial muscle disease. Cell Metab. 2022;34(2):197-208.e5.
Huang Z, Fang Q, Ma W, Zhang Q, Qiu J, Gu X, et al. Isoquercitrin delays denervated soleus muscle atrophy by inhibiting oxidative stress and inflammation. Front Pharmacol. 2019;10:997.
Zhang L, Li M, Wang W, Yu W, Liu H, Wang K, et al. Celecoxib alleviates denervation-induced muscle atrophy by suppressing inflammation and oxidative stress and improving microcirculation. Biochem Pharmacol. 2022;203: 115186.
Zeng Z, Liang J, Wu L, Zhang H, Lv J, Chen N. Exercise-induced autophagy suppresses sarcopenia through Akt/mTOR and Akt/FoxO3a signal pathways and AMPK-mediated mitochondrial quality control. Front Physiol. 2020;11: 583478.
Ito A, Hashimoto M, Tanihata J, Matsubayashi S, Sasaki R, Fujimoto S, et al. Involvement of Parkin-mediated mitophagy in the pathogenesis of chronic obstructive pulmonary disease-related sarcopenia. J Cachexia Sarcopenia Muscle. 2022;13(3):1864–82.
Marcangeli V, Youssef L, Dulac M, Carvalho LP, Hajj-Boutros G, Reynaud O, et al. Impact of high-intensity interval training with or without l-citrulline on physical performance, skeletal muscle, and adipose tissue in obese older adults. J Cachexia Sarcopenia Muscle. 2022;13(3):1526–40.
Konopka AR, Suer MK, Wolff CA, Harber MP. Markers of human skeletal muscle mitochondrial biogenesis and quality control: effects of age and aerobic exercise training. J Gerontol A Biol Sci Med Sci. 2014;69(4):371–8.
Balan E, Schwalm C, Naslain D, Nielens H, Francaux M, Deldicque L. Regular endurance exercise promotes fission, mitophagy, and oxidative phosphorylation in human skeletal muscle independently of age. Front Physiol. 2019;10:1088.
Sakellariou GK, Pearson T, Lightfoot AP, Nye GA, Wells N, Giakoumaki II, et al. Mitochondrial ROS regulate oxidative damage and mitophagy but not age-related muscle fiber atrophy. Sci Rep. 2016;6:33944.
Leduc-Gaudet JP, Reynaud O, Hussain SN, Gouspillou G. Parkin overexpression protects from ageing-related loss of muscle mass and strength. J Physiol. 2019;597(7):1975–91.
Drummond MJ, Addison O, Brunker L, Hopkins PN, McClain DA, LaStayo PC, et al. Downregulation of E3 ubiquitin ligases and mitophagy-related genes in skeletal muscle of physically inactive, frail older women: a cross-sectional comparison. J Gerontol A Biol Sci Med Sci. 2014;69(8):1040–8.
Ryu D, Mouchiroud L, Andreux PA, Katsyuba E, Moullan N, Nicolet-Dit-Félix AA, et al. Urolithin A induces mitophagy and prolongs lifespan in C. elegans and increases muscle function in rodents. Nat Med. 2016;22(8):879–88.
Jiménez-Loygorri JI, Villarejo-Zori B, Viedma-Poyatos Á, Zapata-Muñoz J, Benítez-Fernández R, Frutos-Lisón MD, et al. Mitophagy curtails cytosolic mtDNA-dependent activation of cGAS/STING inflammation during aging. Nat Commun. 2024;15:830.
Rosa-Caldwell ME, Lim S, Haynie WS, Brown JL, Lee DE, Dunlap KR, et al. Mitochondrial aberrations during the progression of disuse atrophy differentially affect male and female mice. J Cachexia Sarcopenia Muscle. 2021;12(6):2056–68.
Shen Y, Zhang Q, Huang Z, Zhu J, Qiu J, Ma W, et al. Isoquercitrin delays denervated soleus muscle atrophy by inhibiting oxidative stress and inflammation. Front Physiol. 2020;11:988.
De Palma C, Morisi F, Cheli S, Pambianco S, Cappello V, Vezzoli M, et al. Autophagy as a new therapeutic target in Duchenne muscular dystrophy. Cell Death Dis. 2012;3(11): e418.
Mucha O, Kaziród K, Podkalicka P, Rusin K, Dulak J, Łoboda A. Dysregulated autophagy and mitophagy in a mouse model of duchenne muscular dystrophy remain unchanged following heme oxygenase-1 knockout. Int J Mol Sci. 2021;23(1):470.
Luan P, D’Amico D, Andreux PA, Laurila PP, Wohlwend M, Li H, et al. Urolithin A improves muscle function by inducing mitophagy in muscular dystrophy. Sci Transl Med. 2021;13(588): eabb0319.
Pal R, Palmieri M, Loehr JA, Li S, Abo-Zahrah R, Monroe TO, et al. Src-dependent impairment of autophagy by oxidative stress in a mouse model of Duchenne muscular dystrophy. Nat Commun. 2014;5:4425.
Hughes MC, Ramos SV, Turnbull PC, Rebalka IA, Cao A, Monaco CMF, et al. Early myopathy in Duchenne muscular dystrophy is associated with elevated mitochondrial H2 O2 emission during impaired oxidative phosphorylation. J Cachexia Sarcopenia Muscle. 2019;10(3):643–61.
Tamura Y, Kitaoka Y, Matsunaga Y, Hoshino D, Hatta H. Daily heat stress treatment rescues denervation-activated mitochondrial clearance and atrophy in skeletal muscle. J Physiol. 2015;593(12):2707–20.
Takeda K, Kitaoka Y, Watanabe K, Miyakawa S, Lindley MR, Takemasa T. Effects of eicosapentaenoic acid intake on denervation-induced mitochondrial adaptation in mouse skeletal muscle. J Phys Fitness Sports Med. 2018;7(5):261–7.
Uemichi K, Shirai T, Matsuno R, Iwata T, Tanimura R, Takemasa T. The role of the mechanistic target of rapamycin complex 1 in the regulation of mitochondrial adaptation during skeletal muscle atrophy under denervation or calorie restriction in mice. Appl Physiol Nutr Metab. 2023;48(3):241–55.
Vainshtein A, Desjardins EM, Armani A, Sandri M, Hood DA. PGC-1α modulates denervation-induced mitophagy in skeletal muscle. Skelet Muscle. 2015;5:9.
Borgia D, Malena A, Spinazzi M, Desbats MA, Salviati L, Russell AP, et al. Increased mitophagy in the skeletal muscle of spinal and bulbar muscular atrophy patients. Hum Mol Genet. 2017;26(6):1087–103.
Memme JM, Oliveira AN, Hood DA. p53 regulates skeletal muscle mitophagy and mitochondrial quality control following denervation-induced muscle disuse. J Biol Chem. 2022;298(2): 101540.
Triolo M, Slavin M, Moradi N, Hood DA. Time-dependent changes in autophagy, mitophagy and lysosomes in skeletal muscle during denervation-induced disuse. J Physiol. 2022;600(7):1683–701.
Oeary MFN, Vainshtein A, Carter HN, Zhang Y, Hood DA. Denervation-induced mitochondrial dysfunction and autophagy in skeletal muscle of apoptosis-deficient animals. Am J Physiol Cell Physiol. 2012;303(4):C447–54.
Capitanio D, Vasso M, De Palma S, Fania C, Torretta E, Cammarata FP, et al. Specific protein changes contribute to the differential muscle mass loss during ageing. Proteomics. 2016;16(4):645–56.
Joseph AM, Adhihetty PJ, Wawrzyniak NR, Wohlgemuth SE, Picca A, Kujoth GC, et al. Dysregulation of mitochondrial quality control processes contribute to sarcopenia in a mouse model of premature aging. PLoS ONE. 2013;8(7): e69327.
Iqbal S, Ostojic O, Singh K, Joseph AM, Hood DA. Expression of mitochondrial fission and fusion regulatory proteins in skeletal muscle during chronic use and disuse. Muscle Nerve. 2013;48(6):963–70.
O’Leary MF, Vainshtein A, Iqbal S, Ostojic O, Hood DA. Adaptive plasticity of autophagic proteins to denervation in aging skeletal muscle. Am J Physiol Cell Physiol. 2013;304(5):C422-430.
Crane JD, Devries MC, Safdar A, Hamadeh MJ, Tarnopolsky MA. The effect of aging on human skeletal muscle mitochondrial and intramyocellular lipid ultrastructure. J Gerontol A Biol Sci Med Sci. 2010;65(2):119–28.
Distefano G, Standley RA, Dubé JJ, Carnero EA, Ritov VB, Stefanovic-Racic M, et al. Chronological age does not influence ex-vivo mitochondrial respiration and quality control in skeletal muscle. J Gerontol A Biol Sci Med Sci. 2017;72(4):535–42.
García-Prat L, Martínez-Vicente M, Perdiguero E, Ortet L, Rodríguez-Ubreva J, Rebollo E, et al. Autophagy maintains stemness by preventing senescence. Nature. 2016;529(7584):37–42.
Fulle S, Protasi F, Di Tano G, Pietrangelo T, Beltramin A, Boncompagni S, et al. The contribution of reactive oxygen species to sarcopenia and muscle ageing. Exp Gerontol. 2004;39(1):17–24.
Gouspillou G, Sgarioto N, Kapchinsky S, Purves-Smith F, Norris B, Pion CH, et al. Increased sensitivity to mitochondrial permeability transition and myonuclear translocation of endonuclease G in atrophied muscle of physically active older humans. FASEB J. 2014;28(4):1621–33.
Bujak AL, Crane JD, Lally JS, Ford RJ, Kang SJ, Rebalka IA, et al. AMPK activation of muscle autophagy prevents fasting-induced hypoglycemia and myopathy during aging. Cell Metab. 2015;21(6):883–90.
Cui H, Hu D, Liu Y, Zhao J. Identifying Acss1, Mtfp1 and Oxct1 as key regulators and promising biomarkers of sarcopenia in various models. Gene. 2024;896: 148053.
Stouth DW, vanLieshout TL, Mikhail AI, Ng SY, Raziee R, Edgett BA, et al. CARM1 drives mitophagy and autophagy flux during fasting-induced skeletal muscle atrophy. Autophagy. 2023. https://doi.org/10.1080/15548627.2023.2288528.
Rosa-Caldwell ME, Brown JL, Perry RA, Shimkus KL, Shirazi-Fard Y, Brown LA, et al. Regulation of mitochondrial quality following repeated bouts of hindlimb unloading. Appl Physiol Nutr Metab. 2020;45(3):264–74.
Marshall RN, Smeuninx B, Seabright AP, Morgan PT, Atherton PJ, Philp A, et al. No effect of five days of bed rest or short-term resistance exercise prehabilitation on markers of skeletal muscle mitochondrial content and dynamics in older adults. Physiol Rep. 2022;10(13): e15345.
Standley RA, Distefano G, Pereira SL, Tian M, Kelly OJ, Coen PM, et al. Effects of β-hydroxy-β-methylbutyrate on skeletal muscle mitochondrial content and dynamics, and lipids after 10 days of bed rest in older adults. J Appl Physiol. 2017;123(5):1092–100.
Kang C, Ji LL. PGC-1α overexpression via local transfection attenuates mitophagy pathway in muscle disuse atrophy. Free Radic Biol Med. 2016;93:32–40.
Yamashita SI, Kyuuma M, Inoue K, Hata Y, Kawada R, Yamabi M, et al. Mitophagy reporter mouse analysis reveals increased mitophagy activity in disuse-induced muscle atrophy. J Cell Physiol. 2021;236(11):7612–24.
Zhang QB, Liu AY, Fang QZ, Wang F, Wang H, Zhou Y. Effect of electrical stimulation on disuse muscular atrophy induced by immobilization: correlation with upregulation of PERK signal and Parkin-mediated mitophagy. Am J Phys Med Rehabil. 2023;102(8):692–700.
Uda M, Yoshihara T, Ichinoseki-Sekine N, Baba T, Yoshioka T. Potential roles of neuronal nitric oxide synthase and the PTEN-induced kinase 1 (PINK1)/Parkin pathway for mitochondrial protein degradation in disuse-induced soleus muscle atrophy in adult rats. PLoS ONE. 2020;15(12): e0243660.
Pileggi CA, Hedges CP, D’Souza RF, Durainayagam BR, Markworth JF, Hickey AJR, et al. Exercise recovery increases skeletal muscle H2O2 emission and mitochondrial respiratory capacity following two-weeks of limb immobilization. Free Radic Biol Med. 2018;124:241–8.
Petrick HL, Handy RM, Vachon B, Frangos SM, Holwerda AM, Gijsen AP, et al. Dietary nitrate preserves mitochondrial bioenergetics and mitochondrial protein synthesis rates during short-term immobilization in mice. J Physiol. 2023. https://doi.org/10.1113/JP284701.
Sin J, Andres AM, Taylor DJR, Weston T, Hiraumi Y, Stotland A, et al. Mitophagy is required for mitochondrial biogenesis and myogenic differentiation of C2C12 myoblasts. Autophagy. 2016;12(2):369–80.
English KL, Paddon-Jones D. Protecting muscle mass and function in older adults during bed rest. Curr Opin Clin Nutr Metab Care. 2010;13(1):34–9.
Luo L, Lu AM, Wang Y, Hong A, Chen Y, Hu J, et al. Chronic resistance training activates autophagy and reduces apoptosis of muscle cells by modulating IGF-1 and its receptors, Akt/mTOR and Akt/FOXO3a signaling in aged rats. Exp Gerontol. 2013;48(4):427–36.
Guan Y, Drake JC, Yan Z. Exercise-induced mitophagy in skeletal muscle and heart. Exerc Sport Sci Rev. 2019;47(3):151–6.
Chen CCW, Erlich AT, Hood DA. Role of Parkin and endurance training on mitochondrial turnover in skeletal muscle. Skelet Muscle. 2018;8(1):10.
Tarawan VM, Gunadi JW, Setiawan, Lesmana R, Goenawan H, Meilina DE, et al. Alteration of autophagy gene expression by different intensity of exercise in gastrocnemius and soleus muscles of Wistar rats. J Sports Sci Med. 2019;18(1):146–54.
Acknowledgements
Not applicable.
Funding
This work was supported by National Key Research and Development Program of China (2021YFD1200801); Sichuan Science and Technology Program (2021YFYZ0007, 2021ZDZX0008, 2021YFYZ0030, and sccxtd-2023-08-09); and the China Agriculture Research System (CARS-35).
Author information
Authors and Affiliations
Contributions
Linyuan Shen* and Li Zhu* proposed the idea for the article. Yuhang Lei†, Yanhao Qiu, and Qiuyang Chen participated in the literature searches. Yuhang Lei†, Mailin Gan†, Xingyu Wang, and Tianci Liao analyzed and summarized the literature. Yuhang Lei† and Mailin Gan† drafted this work and contributed to the original writing. Mengying Zhao, Lei Chen, Shunhua Zhang, Ye Zhao, Lili Niu, and Yan Wang reviewed and proofread this paper. Linyuan Shen* and Li Zhu* critically revised the manuscript. All the authors have read and agreed to the published version of the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no potential conflicts of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lei, Y., Gan, M., Qiu, Y. et al. The role of mitochondrial dynamics and mitophagy in skeletal muscle atrophy: from molecular mechanisms to therapeutic insights. Cell Mol Biol Lett 29, 59 (2024). https://doi.org/10.1186/s11658-024-00572-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s11658-024-00572-y