
Abstract
Mesotoga strain PhosAc3 was the first mesophilic cultivated member of the order Thermotogales. This genus currently contain two described species, M. prima and M. infera. Strain PhosAc3, isolated from a Tunisian digestor treating phosphogypsum, is phylogenetically closely related to M. prima strain MesG1.Ag.4.2T. Strain PhosAc3 has a genome of 3.1 Mb with a G+C content of 45.2%. It contains 3,051 protein-coding genes of which 74.6% have their best reciprocal BLAST hit in the genome of the type species, strain MesG1.Ag.4.2T. For this reason we propose to assign strain PhosAc3 as a novel ecotype of the Mesotoga prima species. However, in contrast with the M. prima type strain, (i) it does not ferment sugars but uses them only in the presence of elemental sulfur as terminal electron acceptor, (ii) it produces only acetate and CO2 from sugars, whereas strain MesG1.Ag.4.2T produces acetate, butyrate, isobutyrate, isovalerate, 2-methyl-butyrate and (iii) sulfides are also end products of the elemental sulfur reduction in theses growth conditions.
Similar content being viewed by others


Introduction
Members of the order Thermotogales typically possess a sheath-like structure called a “toga” and are mostly known as thermophilic or hyperthermophilic bacteria. Most species within this order have been isolated from heated sub-seafloors, marine hydrothermal vents, terrestrial hot springs and oil field reservoirs. Interestingly, SSU rRNA genes of Thermotogales were also detected in samples from polluted environments such as sediments of harbors and sludge from waste water treatment plants [1]. Accordingly they were also found in mesothermic enrichment cultures, notably those capable of (i) reductively dechlorinating 2, 3, 4, 5-tetrachlorobiphenyl, (ii) oxidizing hydrocarbons [2]. We reported in 2011 the first cultivation and a preliminary description of a mesophilic bacterium pertaining to the Thermotogales (strain PhosAc3) which was tentatively named “Mesotoga sulfurireducens” [3]. This mesophilic isolate was shown to belong to a large group of uncultivated bacteria that is distantly related to the thermophilic genus Kosmotoga. Soon after, M. prima strain MesG1.Ag.4.2T isolated from sediments from Baltimore Harbor [4] and M. infera strain VNs100T isolated from a water sample collected in the area of an underground gas storage [5] were fully characterized and described as new species. Strain PhosAc3 was isolated from a digestor treating phosphogypsum inoculated with a mixture of marine sediments and sludge originating from a dump and a wastewater treatment plant in Tunisia. It grows at temperatures between 30°C and 50°C (optimum 40°C) and uses fructose and lactate as energy sources. Phylogenetic analyses based on 16S rRNA gene sequences revealed that strain PhosAc3 is closely related to M. prima strain MesG1.Ag.4.2T[3].
Here we report on further taxonomic and physiological studies on strain PhosAc3 and describe the draft genome sequence and its annotation. We show that while they belong to the same species, PhosAc3 and MesG1.Ag.4.2T strains exhibit significant phenotypic and metabolic differences and that their genomes differ by about 25% in gene content.
Organism information
Classification and features
Genomic sequences of strain PhosAc3 showed that it possesses two copies of the 16S rRNA gene. As for M. infera, the two 16S rRNA coding genes found in PhosAc3 are 100% identical (this was further confirmed by re-sequencing of PCR products obtained using two primers pairs specifically designed to target the two 16S rRNA gene loci respectively). This situation contrasts with that of MesG1.Ag.4.2T, which was reported to harbor two distinct 16S rRNA genes that are 99.1% identical (Theba_0197 and Theba_1521). The two 16S rRNA genes of strain PhosAc3 share 99.2% identity with the sequence of MesG1Ag4.2. 16S rRNA gene A. Experiments conducted by the DSMZ (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH) Identification Service on PhosAc3 DNA revealed 78.7% of DNA-DNA re-association with M. prima MesG1.Ag.4.2T, which is a sufficient criterion to classify both strain in the same species. The phylogenetic position of strain PhosAc3 is shown in Figure 1.

Rooted maximum likelihood phylogeny of SSU rRNA sequences from Thermotogales (45 sequences, 1265 nucleotide positions). Numbers at nodes represent the bootstrap values estimated by the non-parametric bootstrap procedure implemented in Treefinder (100 replicates of the original alignments) and the posterior probabilities computed by Mr Bayes (only values greater than 50% and 0.5 respectively, are shown). The scale bar represents the average number of substitution per site.
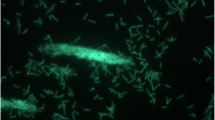
Strain PhosAc3 is a Gram-negative, pleomorphic bacterium. Cells appeared mostly as chains with a rod to coccoid shape of 2–4 μm long and 1–2 μm in diameter (Figure 2). They were non-motile. Strain PhosAc3 is a strict anaerobe. It is a mesophilic bacterium with an optimal growth temperature at 40°C (range 30-50°C). Additional analyses were performed to complete the characterisation of strain PhosAc3 using the same experimental procedures as detailed previously [3, 4]. The optimal growth NaCl concentration was found at 2 g. L−1 (range 0–30 g. L−1). The optimum pH range for growth was 6.9 (range 6.7-7.9). Elemental sulfur (10 g. L−1) was used as terminal electron acceptor, but not thiosulfate, sulfate or sulfite. Strain PhosAc3 used poorly yeast extract but requires it at low concentration (at 0.5 g. L−1) to grow on sugars, peptides and organic acids (arabinose, fructose, glucose, maltose, mannose, raffinose, saccharose, xylose, cellobiose, peptone, lactate and pyruvate) probably as vitamins and other growth factor sources. In contrast, the following substrates were not utilized: galactose, lactose, ribose, gelatin, casein, xylan, cellulose, acetate, butyrate, fumarate, succinate, ethanol, methanol, 1-propanol, and propionate. No growth by fermentation was observed with any combination of yeast extract and peptides or sugars in the absence of elemental sulfur, contrasting with what was reported for M. prima strain MesG1Ag4.2 (Additional file 1: Table S1). Surprisingly, acetate was also required at low concentration (2 mM) to initiate growth most likely to serve as carbon source for anabolism and thus was latter systematically added to the culture medium. End products of sugar metabolism were acetate and CO2. Sulfide production resulted from reduction of elemental sulfur. In any conditions of cultures, hydrogen was detected only as traces with concentrations around 1 μM measured in the gas phase. Finally, no growth was detected with H2/CO2 gas (200 kPa) in the headspace, with or without acetate added to the culture medium.

Phase contrast micrographs of strain PhosAc3. Scale bar: 10 μm.
All these informations on strain PhosAc3 are summarized in Table 1.

Circular representation of the Mesotoga strain PhosAc3 chromosome. Circles display (from the outside): (1) GC percent deviation (GC window - mean GC) in a 1000-bp window, (2) Predicted CDSs transcribed in the clockwise direction, (3) Predicted CDSs transcribed in the counterclockwise direction. Genes displayed in (2) and (3) are color-coded according different categories. Red and blue: MaGe validated annotations, orange: MicroScope automatic annotation with a reference genome, purple: Primary/Automatic annotations. (4) GC skew (G+C/G-C) in a 1000-bp window. (5) rRNA (blue), tRNA (green), misc_RNA (orange), Transposable elements (pink) and pseudogenes (grey).
Chemotaxonomic data
The fatty acid analysis was performed by the DSMZ on a PhosAc3 culture stopped at the end of exponential phase. Fatty acids were extracted using the method of Miller [12], analyzed by gas chromatography (gas chromatograph, model 6890 N, Agilent Technologies) and the resulting profile was determined using the Microbial Identification System (MIDI, Sherlock Version 6.1; database, TSBA40). The fatty acid pattern of strain Phos Ac3 was similar to that of M. infera (Additional file 1: Table S2). In contrast to these bacteria, C14 was not detected in M. prima type species (MesG1.Ag.4.2T) thus suggesting that strain PhosAc3 should be considered as novel ecotype of M. prima species.
Genome sequencing information
Genome project history
This organism was selected for sequencing on the basis of its environmental and biotechnological relevance to issues in global carbon cycling, bioremediation of polluted soils and its significance in studying the evolutionary mechanisms of adaptation to moderate temperatures [13]. The genome project and an improved-high-quality-draft genome sequence have been deposited in the GOLD and IMG databases respectively. A summary of the project information is shown in Table 2.
Growth conditions and DNA isolation
Genomic DNA was isolated from an exponentially growing culture of strain PhosAc3 using the protocol of Marteinsson et al. [14].
Genome sequencing and assembly
De novo whole-genome shotgun sequencing was performed by combining a single and a long paired end (8 kbp) non-cloned libraries sequencing using the Roche Titanium pyrosequencing GS FLX+ technology (MWG Eurofins). This produced 350,813 reads with an average length of 439 bp for a total number of sequenced bases of 154,143,916 representing a sequencing depth of 49×. Using Celera Assembler software (v.6.1) both data sets could be assembled into four scaffolds including 14 large contigs (>1,000 bp) and 127 small contigs.
Genome annotation
Gene predictions annotation and comparative genomic analyses were performed using the MicroScope annotation platform [15]. The predicted CDSs were translated and used to search the National Center for Biotechnology Information non-redundant database, UniProt, TIGRFam, Pfam, PRIAM, KEGG, COGs, and InterPro. These data sources were combined to assert a product description for each predicted protein. Non-coding genes and miscellaneous features were predicted using tRNAscan-SE, RNAMMer, Rfam, TMHMM, and signalP. Additional gene prediction analysis and functional annotation were performed within the Integrated Microbial Genomes Expert Review platform [16]. CRISPR were searched using CRISPRFinder [17]. Table 3 presents the project information and its association with MIGS version 2.0 compliance [18].
Genome properties
The overall genome size estimated for M. prima strain PhosAc3 is 3,113,612 bp, significantly larger than that of the M. prima type strain MesG1.Ag.4.2 (2,974,229 bp) [19] and is composed of a unique circular chromosome (no plasmid was found in contrast to MesG1.Ag.4.2T). The average genome G+C content of strain PhosAc3 of 45.19% is close to that of MesG1.Ag.4.2T (45.45%). It contains two ribosomal operons, 47 tRNAs and 3,051 predicted protein-coding genes (Table 3; Figure 3).
Insights from the genome sequence
Like Mesotoga prima (strain MesG1.Ag.4.2T), Mesotoga strain PhosAc3 possesses a significantly larger genome (3.11 and 2.97 Mb respectively) than their thermophilic counterparts within the Thermotogales whose genome size ranged from 1.86 to 2.30 Mb. Of the 3051 protein encoding genes (CDS) of strain PhosAc3, 2392 (78.4%) have their best homologs (satisfying the bi-directional best hit criterion) in the complete genome of M. prima and are clustered in 273 syntons (cluster of at least two contiguous genes) in the two strains (SM Figure 1). For comparison sake, the two Mesotoga strains, MesG1.Ag.4.2T and PhosAc3, share respectively 1468 and 1542 CDS with the closely related species Kosmotoga olearia strain TBF 19.5.1 (SM Figure 1). It seems that the supplementary genes found in Mesotoga strain PhosAc3 (not present in K. olearia) have been acquired by LGT mostly from mesophilic members of the Firmicutes (peculiarly within the Clostridiales order) to the Mesotoga (data not shown) with whom they share the same microbial habitat [19]. As previously observed for Mesotogaprima strain MesG1.Ag.4.2T, the largest fractions of the genes presumably acquired by LGT are involved in amino acids transport and metabolism (COG category E), secondary metabolite biosynthesis (COG category Q) and signal transduction mechanisms (COG category T) (Table 4; data not shown). Of the 659 genes of Mesotoga strain PhosAc3 with no detectable homologs in M. prima strain MesG1.Ag.4.2T, the majority (65%) are annotated as unknown function proteins, 58 (8,8%) correspond to transposons and the rest to poorly characterized functions (COG categories S and R).
Conclusion
Based on taxonomic and genomic criteria, Mesotoga strain PhosAc3 should be considered a novel strain of the M. prima species. Besides numerous similarities, both strains exhibit clear differences in their phenotypic features and even to their gene content. They may therefore represent distinct ecotypes as previously defined [20, 21]. Strain PhosAc3, like M. infera, is capable of significant growth on simple substrates (sugars and organic acids) only in the presence of elemental sulfur as terminal electron acceptor suggesting that these substrates are oxidized rather than fermented. This sharply contrasts with the reported fermentative metabolism of sugars of M. prima type species (strain MesG1.Ag.4.2T). Other differences between strain PhosAc3 and M. prima strain MesG1.Ag.4.2T include the end products of sugar metabolism, the optimum NaCl concentration for growth and the range of electron acceptors used (Table 1). The availability of the genome sequences of two Mesotoga strains offers a good opportunity to look in further details the genomic determinants that may be responsible of the metabolic differences observed between the two strains. Moreover, the comparison with other Thermotogales genomes should bring relevant information regarding the bacterial adaptation to novel ecological niches (from hot to mesothermic biotopes) and the importance of lateral gene transfer in such evolutionary processes [13].
References
Nesbo CL, Dlutek M, Zhaxybayeva O, Doolittle WF: Evidence for existence of “mesotogas”, members of the order Thermotogales adapted to low-temperature environments. Appl Environ Microbiol 2006, 72: 5061–5068. 10.1128/AEM.00342-06
Chouari R, Le Paslier D, Daegelen P, Ginestet P, Weissenbach J, Sghir A: Novel predominant archaeal and bacterial groups revealed by molecular analysis of an anaerobic sludge digester. Environ Microbiol 2005, 7: 1104–1115. 10.1111/j.1462-2920.2005.00795.x
Ben Hania W, Ghodbane R, Postec A, Brochier-Armanet C, Hamdi M, Fardeau M-L, et al.: Cultivation of the first mesophilic representative (“ Mesotoga ”) within the order Thermotogales . Syst Appl Microbiol 2011, 34: 581–585. 10.1016/j.syapm.2011.04.001
Nesbo CL, Bradnan DM, Adebusuyi A, Dlutek M, Petrus AK, Foght J, et al.: Mesotoga prima gen. nov., sp nov., the first described mesophilic species of the Thermotogales . Extremophiles 2012, 16: 387–393. 10.1007/s00792-012-0437-0
Ben Hania W, Postec A, Aullo T, Ranchou-Peyruse A, Erauso G, Brochier-Armanet C, et al.: Mesotoga infera sp. nov., a mesophilic member of the order Thermotogales , isolated from an underground gas storage aquifer. Int J Syst Evol Microbiol 2013, 63: 3003–3008. 10.1099/ijs.0.047993-0
Woese CR, Kandler O, Wheelis ML: Towards a natural system of organisms: proposal for the domains Archaea, Bacteria , and Eucarya . Proc Natl Acad Sci U S A 1990, 87: 4576–4579. 10.1073/pnas.87.12.4576
Reysenbach A: Phylum BII. Thermotogae phy. nov. In Bergey’s Manual® of Systematic Bacteriology. Volume Volume One : The Archaea and the Deeply Branching and Phototrophic Bacteria. Edited by: Boone DR, Castenholz RW, Garrity GM. New York: Springer; 2001:369–387.
Reysenbach A: Class I. Thermotogae class nov. In Bergey’s Manual® of Systematic Bacteriology. Volume Volume One : The Archaea and the Deeply Branching and Phototrophic Bacteria. Edited by: Boone DR, Castenholz RW, Garrity GM. New York: Springer; 2001:369–370.
Reysenbach A: Order I. Thermotogales ord. nov. In Bergey’s Manual® of Systematic Bacteriology. Volume Volume One : The Archaea and the Deeply Branching and Phototrophic Bacteria. Edited by: Boone DR, Castenholz RW, Garrity GM. New York: Springer; 2001:369–370.
Reysenbach A: Thermotogaceae fam. nov. In Bergey’s Manual® of Systematic Bacteriology. Volume Volume One : The Archaea and the Deeply Branching and Phototrophic Bacteria. Edited by: Boone DR, Castenholz RW, Garrity GM. New York: Springer; 2001:369–370.
Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, et al.: Gene ontology: tool for the unification of biology. The gene ontology consortium. Nat Genet 2000, 25: 25–29. 10.1038/75556
Miller LT: Single derivatization method for routine analysis of bacterial whole-cell fatty acid methyl esters, including hydroxy acids. J Clin Microbiol 1982, 16: 584–586.
Zhaxybayeva O, Swithers KS, Lapierre P, Fournier GP, Bickhart DM, DeBoy RT, et al.: On the chimeric nature, thermophilic origin, and phylogenetic placement of the Thermotogales . Proc Natl Acad Sci U S A 2009, 106: 5865–5870. 10.1073/pnas.0901260106
Marteinsson V, Watrin L, Prieur D, Caprais J-C, Raguenès G, Erauso G: Sulfur-metabolizing hyperthermophilic anaerobic Archaea isolated from hydrothermal vents in the Southwestern Pacific Ocean. Int J Syst Bacteriol 1995, 45: 623–632. 10.1099/00207713-45-4-623
Vallenet D, Engelen S, Mornico D, Cruveiller S, Fleury L, Lajus A, et al.: MicroScope: a platform for microbial genome annotation and comparative genomics. Nucleic Acids Res Database (Oxford). 2009;2009:bap021. Epub 2009 Nov 25. Database (Oxford). 2009;2009:bap021. Epub 2009 Nov 25.
Markowitz VM, Chen I-MA, Palaniappan K, Chu K, Szeto E, Grechkin Y, et al.: IMG: the integrated microbial genomes database and comparative analysis system. Nucleic Acids Res 2012, 40: D115-D122. 10.1093/nar/gkr1044
Grissa I, Vergnaud G, Pourcel C: CRISPRFinder: a web tool to identify clustered regularly interspaced short palindromic repeats. Nucleic Acids Res 2007, 35: W52-W57. 10.1093/nar/gkm360
Field D, Garrity G, Gray T, Morrison N, Selengut J, Sterk P, et al.: The minimum information about a genome sequence (MIGS) specification. Nat Biotechnol 2008, 26: 541–547. 10.1038/nbt1360
Zhaxybayeva O, Swithers KS, Foght J, Green AG, Bruce D, Detter C, et al.: Genome sequence of the mesophilic Thermotogales Bacterium Mesotoga prima MesG1.Ag.4.2 reveals the largest Thermotogales genome to date. Genome Biol Evol 2012, 4: 700–708. 10.1093/gbe/evs059
Fenchel T: Microbiology. Biogeography for bacteria. Science 2003, 301: 925–926. 10.1126/science.1089242
Nesbo CL, Dlutek M, Doolittle WF: Recombination in Thermotoga : implications for species concepts and biogeography. Genetics 2006, 172: 759–769.
Acknowledgments
Sequencing cost was supported by Protéus SA. We thank Zoe Rouy from the MicroScope team for providing access to this powerful annotation platform. C.B.-A. is member of the Institut Universitaire de France. C. B-A is funded by the ANR-10-BINF-01-01 (Ancestrome) grants.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
WB, KF and MLF performed experiments to complete the characterization of strain PhosAc3. WB and KF prepared genomic DNA of strain PhosAc3 for sequencing at GATC. GE and JLM processed the raw data through annotation pipelines. GE and AD did expert annotation searches. CB and AP performed phylogenetic analyses. CP, representing PROTEUS Cie, funded the project. GE and BO drafted the paper. All authors read and approved the final manuscript.
Electronic supplementary material
40793_2014_945_MOESM1_ESM.docx
Additional file 1: Table S1.: Differential characteristics between Mesotoga prima strains and M. infera. Table S2. Fatty acids composition of Mesotoga prima strains and M. infera. Figure S1. Number of common (core) or strain-specific genes in Mesotoga strains and Kosmotoga olearia TBF 19.5.1. (DOCX 123 KB)
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Ben Hania, W., Fadhlaoui, K., Brochier-Armanet, C. et al. Draft genome sequence of Mesotoga strain PhosAC3, a mesophilic member of the bacterial order Thermotogales, isolated from a digestor treating phosphogypsum in Tunisia. Stand in Genomic Sci 10, 12 (2015). https://doi.org/10.1186/1944-3277-10-12
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1944-3277-10-12