
Abstract
Background
Pyrethroid resistance has been slower to emerge in Anopheles arabiensis than in An. gambiae s.s and An. funestus and, consequently, studies are only just beginning to unravel the genes involved. Permethrin resistance in An. arabiensis in Lower Moshi, Tanzania has been linked to elevated levels of both P450 monooxygenases and β-esterases. We have conducted a gene expression study to identify specific genes linked with metabolic resistance in the Lower Moshi An. arabiensis population.
Methods
Microarray experiments employing an An. gambiae whole genome expression chip were performed on An. arabiensis, using interwoven loop designs. Permethrin-exposed survivors were compared to three separate unexposed mosquitoes from the same or a nearby population. A subsection of detoxification genes were chosen for subsequent quantitative real-time PCR (qRT-PCR).
Results
Microarray analysis revealed significant over expression of 87 probes and under expression of 85 probes (in pairwise comparisons between permethrin survivors and unexposed sympatric and allopatric samples from Dar es Salaam (controls). For qRT-PCR we targeted over expressed ABC transporter genes (ABC ‘2060’), a glutathione-S-transferase, P450s and esterases. Design of efficient, specific primers was successful for ABC ‘2060’and two P450s (CYP6P3, CYP6M2). For the CYP4G16 gene, we used the primers that were previously used in a microarray study of An. arabiensis from Zanzibar islands. Over expression of CYP4G16 and ABC ‘2060’ was detected though with contrasting patterns in pairwise comparisons between survivors and controls. CYP4G16 was only up regulated in survivors, whereas ABC ‘2060’ was similar in survivors and controls but over expressed in Lower Moshi samples compared to the Dar es Salaam samples. Increased transcription of CYP4G16 and ABC ‘2060’ are linked directly and indirectly respectively, with permethrin resistance in Lower Moshi An. arabiensis.
Conclusions
Increased transcription of a P450 (CYP4G16) and an ABC transporter (ABC 2060) are linked directly and indirectly respectively, with permethrin resistance in Lower Moshi An. arabiensis. Our study provides replication of CYP4G16 as a candidate gene for pyrethroid resistance in An. arabiensis, although its role may not be in detoxification, and requires further investigation.
Similar content being viewed by others

Background
Vector control programmes employing ITNs and IRS are a mainstay of malaria control [1, 2]. Continuous usage of insecticides under such malaria control programmes and/or agricultural application of insecticides has resulted in the development of resistance in major malaria vector species [3–7]. Anopheline mosquito populations in many parts of Africa have developed resistance to pyrethroids [8], which are widely used for indoor residual spraying (IRS) and the only insecticide class available for insecticide treated nets (ITNs). Pyrethroids have been shown to pose very low health risks to humans and other mammals, but are toxic to insects and knock them down (kill them), even at very low doses [9].
Insecticide resistance is considered a major threat to the continued success of IRS and ITNs, with reduction in efficacy of pyrethroids being a particular concern [8, 10]. The various mechanisms by which mosquitoes are thought to develop resistance to insecticides include metabolic resistance, target-site resistance, reduced penetration and behavioural avoidance. However, metabolic resistance and target-site resistance are major mechanisms assumed to be responsible for insecticide resistance [11] and are known to contribute to pyrethroid resistance in malaria vectors. While target-site resistance occurs when the site of action of insecticide is altered such that the insecticide no longer binds effectively, metabolic resistance involves over-expression of enzymes capable of detoxifying insecticides or amino acid substitutions within these enzymes, which alter the affinity of the enzyme for the insecticide [12]. The most common target site resistance mechanism constitutes two point mutations at amino acid position 1014 of the voltage-gated sodium channel gene, resulting in either a leucine-phenylalanine (L1014F) [13], or a leucine-serine (L1014S) substitution [14].
Detoxification enzymes include glutathione-S-transferases (GSTs), esterases and P450 monooxygenases. GST-based DDT resistance by breaking down DDT to non-toxic products is common in a number of anopheline species including An. gambiae[15–17]. It has been suggested that GSTs may play a role in pyrethroid resistance by detoxifying lipid peroxidation products induced by pyrethroids and/or by protecting from insecticide exposure induced oxidative stress [18]. GSTs might also confer a secondary role in pyrethroid resistance by sequestering the insecticide, hence reducing the total in vivo concentration of insecticide [19]. More recently, the study of Riveron et al., 2014 [20] revealed a more direct role played by GSTs in conferring insecticide resistance in malaria vectors.
Elevated levels or activity of esterase enzymes which hydrolyze ester bonds or sequester insecticides is one of the most common metabolic resistance mechanisms in organophosphate (OPs) resistant mosquito species. Malathion (OP) resistance in Anopheles culicifacies and Anopheles stephensi has been associated with an altered form of esterase that specifically metabolizes the molecule at a much faster rate than that in susceptible strains [21, 22].
Cytochrome P450-dependent monooxygenases are an important and diverse family of enzymes involved in the metabolism of numerous endogenous and exogenous compounds. Cytochrome P450 belongs to six families and increased transcription of genes belonging to the CYP4, CYP6, and CYP9 has been observed in various insecticide-resistant species [23]. Microarray-based approaches have identified three candidate P450 genes, CYP6M2, CYP6P3 and CYP6Z2 that were found to be repeatedly over-produced in pyrethroid resistant populations of An. gambiae [24–26]. All of these genes encode for enzymes that are able to bind to type I and type II pyrethroids but only CYP6P3 and CYP6M2 were shown to metabolize the insecticides [27, 28]. More recently, some studies demonstrated that CYP6M2 is also capable of metabolizing the organochlorine insecticide DDT in An. gambiae, hence demonstrating the first evidence for a metabolic cross-resistance in malaria vectors [29].
Both metabolic resistance and target-site resistance have been documented in An. arabiensis, one of the dominant vector species of malaria in sub-Saharan Africa [30, 31] and in some areas such as the Great Rift Valley in East Africa, An. arabiensis is the predominant malaria vector species [32]. To date, the kdr mutations have been reported in several countries including Uganda [33], Sudan [34, 35], Cameroon [36], and Tanzania [37–39]. In addition to target site resistance, metabolic resistance has also been documented in different countries [40–42].
The study of dynamics of insecticide resistance in Lower Moshi An. arabiensis from 2009 to 2013 [39] has shown that the population has developed resistance to all pyrethroids tested (permethrin, deltamethrin and lambdacyhalothrin) with the presence of L1014F mutation at very low frequency. Tanzania has been scaling-up the use of pyrethroid-based LLINs which reached its universal coverage under Universal Coverage Campaign [43]. Therefore, higher pyrethroid resistance is expected following higher selection pressure on this Lower Moshi An. arabiensis population and it is likely that pyrethroid resistance will spread to other areas in the country. The previous study by Matowo et al., 2010 [44] had revealed significantly higher levels of oxidase and β-esterase enzymes in wild Lower Moshi An. arabiensis than in a laboratory reference strain. However, the genes responsible for elevated levels of these detoxification enzymes remained unknown, although over-expression of a few detoxification genes has been documented in An. arabiensis populations in Hai district, adjacent to Lower Moshi. [45]. Therefore, the aim of this study was to explore and identify specific genes involved in metabolic resistance to permethrin in Lower Moshi populations of An. arabiensis from Lower Moshi.
Methods
Mosquito sample collection and identification
Collections were conducted in the rainy season between May and June 2011 in Mabogini village (37°21′E, 3°24′S), Mabogini is an agricultural area of about 15 km south of Moshi town, in which rice is cultivated in two growing seasons, the main season in which rice is sown is mid-June with irrigation from June to October, and a second season of sporadic cultivation from September to February. The rice paddies thus provide breeding sites for Anopheles arabiensis, the predominant malaria vector in the area, throughout much of the year.
The larvae were collected from two specific localities, Mbugani (37°22' E, 3°25'S) and Harusini (37°21'E, 3°23'S) in Mabogini village which are 4 km. apart. Additional larval collections were made from Mbugani in August 2012.
Anopheline larvae were also collected from temporary breeding habitats in Ilala (Gerezani) (39°16' E, 6°49'S) and Kinondoni (Kawe) (39°13' E, 6°43'S) in Dar es salaam about 700 km from Moshi, in August and October 2011.
The field-collected anopheline larvae were transferred into an insectary and reared to adults. Emergent adults were split by sex before mating could occur and fed with 10% sugar solution.
Virgin females were preferred following the previous whole-body microarray experiments [46] that detected large transcriptional changes in mated females where the number of genes differentially expressed increased with time post mating.
At 3 days-old, adult females from Mbugani were exposed to 0.75% permethrin for one hour using a standard WHO tube bioassay procedure [47] with mortality scored 24 hours after the exposure period. Adult mosquitoes from Harusini, Moshi and Dar es Salaam were not exposed to insecticide. At their fourth day post emergence, all unexposed (control) females and the survivors (resistant mosquitoes) were killed in 75% ethanol. A hind leg was removed from each mosquito and stored over silica gel in an individually-labelled tube before placing the remainder of the specimens in RNA later in an Eppendorf tube. DNA was extracted from the legs that were removed from each specimen for species identification.
Species identification focused on the most prevalent local vectors of An. gambiae sensu lato; An. arabiensis and An. gambiae s.s. Mosquitoes were identified to species level using standard PCR method [48]. Only those identified as An. arabiensis were further analysed in this study.
Whole genome microarrays
Experimental design
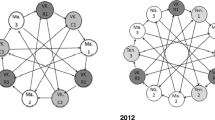
The study was designed to comprise of two microarray experiments. The first involved three groups of samples collected in 2011 from Mbugani (insecticide exposed, selecting for the 24% and 54% of the most resistant females [39] and unexposed) and Harusini (unexposed). Although initially suspected that Harusini might display a lower resistance level than Mbugani, its close proximity made predictions ambiguous and only results from hybridizations of Mbugani permethrin-selected against Mbugani unexposed and Mbugani permethrin-selected against Harusini unexposed were used. In the second experiment, to investigate temporal repeatability of results we repeated the experiment from Mbugani (permethrin-selected against. unexposed) using a collection from 2012. In this second experiment a pyrethroid-susceptible sample from Kinondoni and Ilala in Dar-es-Salaam [40] was also included, primarily to permit an allopatric comparison with the population from Mbugani (insecticide exposed that selected for the 54% of the most resistant females [39]. However, we found that comparisons involving the Dar-es-Salaam sample yielded very little additional resolution for the analysis, which coupled with suspected technical errors in some of the hybridizations led to their exclusion.
2011: Mbugani permethrin selected vs Mbugani unexposed
2011: Mbugani permethrin-selected vs. Harusini unexposed
2012: Mbugani permethrin-selected vs. Mbugani unexposedComparisons in experiment 1 consisted of three independent biological replicates while those of experiment 2 consisted of four independent biological replicates (Figure 1); and two technical repeats of dye swaps to control for dye bias. Hybridization and analysis followed an interwoven loop design.

Interwoven microarray experimental loop design for experiment 1 and 2.
To be judged significant a probe was required to (a) exhibit a false discovery rate-corrected probability of q < 0.05 in all three comparisons, and (b) exhibit a consistent directionality in expression, i.e. either Mbugani permethrin-selected groups > unexposed groups or Mbugani permethrin-selected groups < unexposed groups in all three comparisons. Owing to the use of within population (i) selected vs. (ii) unexposed comparisons, the magnitude of differential expression is expected to be relatively low, because (i) is a subset of (ii) [20] and therefore no fold-change threshold was applied as a significance criterion.
RNA extraction and labelling
Both four day old unfed unmated female mosquitoes that survived one hour exposure to 0.75% permethrin, held for 24 hours post–exposure and unexposed mosquitoes were killed using 70% absolute ethanol and stored in RNAlater® at -20°C. Total RNA was extracted from batches of ten mosquitoes using the RNAqueousR-4PCR Kit (Ambion) according to manufacturer’s instructions and treated with DNase I (Qiagen). The quality and quantity of the RNA was checked using a using a Nanodrop spectrophotometer (Nanodrop Technologies, Wilmington, DE, USA). and Bioanalyser (Agilent Technologies, Stratagene, USA) respectively. Extracted mRNA was dye-labelled using the Low Input Quick Amp Labelling Kit (Agilent Technologies, Stratagene, USA) according to manufacturer’s instructions. Labelling was repeated for samples that had a low cDNA yield (<15 ng/μl) and poor dye incorporation (<6.0 pmol/μl of each dye).
The labelled samples were hybridized using a Gene Expression Hybridization Kit (Agilent Technologies). Washing, scanning and feature extraction were performed according to the manufacturer’s recommendations.
Microarray analysis
Analysis of the microarray data was performed on corrected spot intensities using the LIMMA 2.4.1 software package [49] for R 2.3.0. Normalization of signal intensities was followed by ANOVA F-test using the MAANOVA package in R [50]. Microarray data are available in the ArrayExpress database (http://www.ebi.ac.uk/arrayexpress) under accession number E-MTAB-2513 (SURV vs UNEXPOSED loop design) and E-MTAB-2514 (SURV vs DAR loop design).
Quantitative real time PCR (qPCR)
Technical repeatability of microarray results for a subset of candidate genes was assessed using Quantitative PCR using the cDNA synthesised from the same RNA samples used in the microarray experiments. The samples were quantified using a Nanodrop spectrophotometer (Nanodrop Technologies, Wilmington, DE, USA). cDNA was synthesised from ~0.5-1 μg of RNA using oligo(dT)20 (50 μM) and SuperScript III (200U) (Invitrogen) and purified through a DNA-binding column (Qiagen). The quality and quantity of cDNA was measured using the Nanodrop spectrophotometer (Nanodrop Technologies, Wilmington, DE, USA) and the cDNA samples were stored at -70°C until further use.
Primers for qPCR were designed using NCBI primer BLAST (http://www.ncbi.nlm.nih.gov/tools/primer-blast/) by using Xm codes from Vector Base. Serial dilutions of cDNA were used for standard curve production to determine PCR efficiency and specificity of the primer pairs. The qPCR reactions were performed using the Agilent MXPro Real-Time PCR detection system (Agilent Technologies, Stratagene, USA).
A total volume of 20 μl contained 10 μl Brilliant III SYBR Green, 300 nM of primers, 2.5 μl of cDNA (1:10 dilution) and the total volume made up with sterile-distilled water. The cycling conditions for all primer sets consisted of 95°C for 15 minutes followed by 40 cycles of 60°C for 10s, 95°C for 30s, 95°C for 1 minute, 55°C for 30s and 95°C for 30s. Three biological replicates were run for each sample on a plate. Each gene was measured in at least six real-time PCR runs (replicate reactions/biological repeats). The real-time PCR (qPCR) reactions were run along with two normalising ‘control’ genes, ribosomal S7 and elongation factor EF1. Primer pairs exhibiting a linear relationship between Ct values and template concentration in standard curves, and high PCR amplification efficiency were chosen for further analysis.
Data analysis of gene expression
Outlier data points were identified and excluded from analysis based on obvious deviations in both the normal shape of amplification curves and the Ct values of other repeated observations (biological triplicates). The mean of the threshold cycles (Ct) for each gene were normalised against the average values for ribosomal S7 and elongation factor EF1. To compare gene expression between treatments, we calculated the ΔΔCt [51]. The fold-change in gene expression for each target gene, normalized to the ribosomal S7 and elongation factor EF1 relative to a susceptible strain of An. arabiensis from Dar es Salaam, was calculated according to the 2-ΔΔCt method incorporating PCR efficiency [51]. Basic data analysis (regression and t-tests were performed in Excel. with p < 0.05 used to assess significant difference between treatments for the t-tests.
Ethical considerations
The study received ethical clearance from the KCMC Research Ethics Committee.
Results
Microarray experiments involving Moshi samples indicated that 87 probes were significantly up regulated in all three comparisons and 85 down-regulated coming from a total of 133 genes (Figure 2). There was a high correlation among fold changes between the different experimental comparisons, suggesting consistency of results (Figure 3). Surprisingly, the genes that were down regulated include multiple detoxification genes, including several that have come up as top upregulated hits in previous An. gambiae s.s. arrays, including P450s repeatedly resistance-associated in previous work on An. gambiae s.s. (CYP6M2 and CYP6P3). Among the up-regulated genes were 3 independent ABC transporters, one of which ABCB4 (represented by 3 alternate transcripts) exhibited almost all significant probes. Also 3 probes for GSTe7 were up-regulated. Full microarray results from both experiments have been submitted as Additional file 1. However, few genes were selected for qPCR. Of about thirty primers that were designed, only six passed the criteria for qPCR. These primers are shown in Table 1 below including the primers for CYP4G16, the gene that was documented in other studies reporting on insecticide resistance in An. arabiensis and two normalising ‘control’ genes, ribosomal S7 and elongation factor EF1 (Table 1). Only two genes, ABC 2060 and CYP4G16 were the most significantly over-transcribed genes in qPCR (Figure 4).

Volcano plot of all probes significant in all of three experimental comparisons of An. arabiensis permethrin-exposed survivors vs. unexposed samples (average of the three experiments). Red triangles show probes of candidate genes chosen for subsequent qRT-PCR, with probes for the same gene within dashed ovals; the tight cluster of overexpressed gene probes are grouped together.

Scatterplot illustrating the consistency of expression (measured as log 2 fold change) for significant probes among the three experiments. R2 value indicates all data; value in parenthesis the R2 value for genes selected for subsequent qRT-PCR.

Relative expression levels of four candidate genes between sample groups of An. arabiensis differing in predicted susceptibility. Data are delta-delta CT values calculated from 5–9 biological repeats. **P < 0.01; *P < 0.05 (t-tests).
Discussion
Transcriptional analysis of pyrethroid resistance in the four day old Lower Moshi An. arabiensis females was investigated using the An. gambiae whole genome microarray [25] and quantitative real-time qPCR. Since a fully sequenced genome for An. arabiensis was not available; we used heterologous hybridization of the An. arabiensis samples to an An. gambiae microarray. The microarray analysis of this study clearly indicates that the differences among arrays could not be appropriately determined. In the previous studies, the use of An. gambiae ‘detox chip’ could successfully determine the gene expression of pyrethroid resistant populations of other species including An. funestus [53], An. stephensi [54] and An. arabiensis [28]. Gene expression of pyrethroid resistant An. arabiensis populations from Pemba Island was also successfully determined by using An. gambiae whole genome microarray [40]. The microarray analysis of this study clearly indicates that there were some very interesting hits among the up regulated genes most notably three independent ABC transporters and GSTe7.
Transcription of four genes was evaluated in this study using quantitative real-time (qPCR). It included one ABC transporter gene ABC 2060 that was up regulated, two P450s (CYP6M2, CYP6P3) that were down regulated in microarray experiments but previously associated with pyrethroid resistance and one P450 gene CYP4G16 that was identified through published literature and implicated in permethrin resistance in An. arabiensis [40]. Only two genes, CYP4G16 and ABC 2060, showed high expression levels (p < 0.05) in the pyrethroid resistant Lower Moshi An. arabiensis and it is possible that the two genes may be playing a role in the observed resistance to pyrethroids in the Lower Moshi An. arabiensis population. The low levels of expression of the two P450s, CYP6M2 and CYP6P3 in the pyrethroid resistant Lower Moshi An. arabiensis implies that the observed pyrethroid resistance results from other mechanisms rather than insecticide metabolism.
CYP4G16 has previously been recorded as an over-transcribed gene in An. arabiensis strain from Cameroon [28], Pemba Island Zanzibar [40] and in a laboratory strain of An. arabiensis from Sudan that was resistant to DDT, permethrin and deltamethrin [41]. In Pemba Island up-regulation of CYP4G16 in resistant An. arabiensis ranged from 2.0 to 4.5 fold compared to susceptible strains of An. arabiensis from Dar es Salaam and Unguja and it has been suggested that it plays a role in cuticular-based resistance. Cytochrome P450s CYP4G17 and CYP4G16 of the Anopheles gambiae are closely related to the Drosophila melanogaster CYP4G1, the specific insect P450 with decarbonylase function that is involved in hydrocarbon biosynthesis. CYP4G16 and CYP4G17 proteins are found in the mosquito head and the carcass of the abdomen, MozAtlas [55] but not the midgut or malpighiam tubules, where P450 detoxification genes have been localized previously.
In this study, metabolic resistance to pyrethroids in Lower Moshi An. arabiensis has been shown to be associated with increased transcription of P450 gene, CYP4G16 and ABC transporter. Following qPCR analysis, CYP4G16 was over-expressed in Lower Moshi pyrethroid resistant An. arabiensis compared to susceptible strains of An. arabiensis from Dar es Salaam. (FC = 3.92, p = 0.007). The gene ABC 2060 was over-expressed with the FC of 5.07, (p = 0.009). ABC transporters have been linked to insecticide resistance in several species such as Helicoverpa armigera [56], Aedes spp. [57, 58], Culex pipiens complex [59] and Anopheles gambiae [60]. ABC transporters have also been correlated with insect resistance to Bacillus thuringiensis [61]. Members of the ABC-transporter family (ABC-transporters) pump foreign molecules out of insect cells using an ATP-dependent mechanism [62–65]. They are also involved in lipid transport to the cuticle [62]. Interestingly, AGAP002060 is a member of the ABCH family, which have been implicated in transportation of lipids to the cuticle [62]. The previous study on An.arabiensis populations a few kilometers from Mabogini village [45] (the current study site) revealed over-transcription of esterase AGAP006227, the UDPGT AGAP006775 and the cytochrome P450s CYP9J4 and CYP6P1. However, its involvement in insecticide resistance is not clear. In addition, the gene AGAP000987 encoding the cuticle protein CPAP3-A1b was also strongly over-transcribed in An. arabiensis in the area. Cuticle proteins play an important role in pyrethroid resistance through altered insecticide penetration and over-transcription of genes encoding them have been reported in pyrethroid resistant species including Anopheles species [45, 66].
Conclusions
The study compared the gene expression profiles of permethrin-resistant and susceptible populations of An. arabiensis using quantitative real-time PCR (qPCR). CYP6P3 is one of the leading candidate genes for metabolic resistance in West African An. gambiae and its down regulation suggests that quite different mechanisms may operate in the different species.
Based on the quantitative real-time PCR q(PCR), one P450 monooxygenase gene CYP4G16 and ABC transporter ABC 2060 were over-transcribed in permethrin resistance in Lower Moshi An. arabiensis. However, further investigation is needed to clarify its role in observed resistance. There is no real evidence that ABC 2060 transporter gene is linked to observed resistance, although significant lower expression of this gene was observed in a susceptible strain from Dar es Salaam than the Moshi population (i.e. either survivors or Moshi control). However, the two genes identified are most likely not the only genes involved in pyrethroid resistant An. arabiensis from Lower Moshi. Further investigation is needed especially in microarray analysis to identify more genes that may be involved in insecticide resistance in the country and its effects on the current malaria control interventions.
Abbreviations
- ITNs:
-
Insecticide-treated nets
- LLINs:
-
Long-lasting insecticide treated nets
- IRS:
-
Indoor residual spraying
- DDT:
-
Dichlorodiphenyltrichloroethane
- OPs:
-
Organophosphates
- GSTs:
-
Glutathione S-transferases
- P450:
-
Cytochrome P450 monooxygenase enzymes
- RNA:
-
Ribonucleic acid
- qPCR:
-
Quantitative Polymerase Chain Reaction
- RT-qPCR:
-
Reverse transcription and quantitative polymerase chain reaction
- cDNA:
-
Complementary Deoxyribonucleic acid
- CYP:
-
Genes encoding cytochrome
- NCBI:
-
National Center for Biotechnology Information
- ANOVA:
-
Analysis of variance
- MAANOVA:
-
Multivariate Analysis of Variance
- MCDC:
-
Malaria Capacity Development Consortium
- KCMC:
-
Kilimanjaro Christian Medical Centre
- WHO:
-
World Health Organization.
References
Zaim M, Jambulingam P: Global insecticide use for vector-borne disease control 3rd edition. 2007, Geneva: World Health Organization
WHO: World malaria report 2013. 2013, Geneva: World Health Organization
Abuelmaali SA, Elaagip AH, Basheer MA, Frah EA, Ahmed FTA, Elhaj HFA, Seidahmed OME, Hamid MMA WD: Impacts of agricultural practices on insecticide resistance in the malaria vector Anopheles arabiensis in Khartoum state, Sudan. PLoS One. 2013, 8 (12): doi:10.1371/annotation/d40e811c-993d-40ec-8294-420402282448
Czeher C, Labbo R, Arzika I, Duchemin JB: Evidence of increasing Leu-Phe knock down resistance- mutation in Anopheles gambiae from Niger following a nationwide long-lasting insecticide treated nets implementation. Malar J. 2008, 7: 189-10.1186/1475-2875-7-189.
Protopopoff N, Verhaegen K, Van Bortel W, Roelants P, Marcotty T, Baza D, D’Alessandro U, Coosemans M: A significant increase in kdr in Anopheles gambiae associated with an intensive vector control intervention in Burundi highlands. Trop Med Int Health. 2008, 13: 1479-1487. 10.1111/j.1365-3156.2008.02164.x.
Yadouleton A, Martin T, Padonou G, Chandre F, Asidi A, Djogbenou L, Dabiré R, Aïkpon R, Boko M, Glitho I, Akogbeto M: Cotton pest management practices and the selection of pyrethroid resistance in Anopheles gambiae population in Northern Benin. Parasit Vectors. 2011, 4: 60-10.1186/1756-3305-4-60.
Diabate A, Baldet T, Chandre F, Akogbéto M, Guiguemde RT, Darriet F, Brengues C, Guillet P, Hemingway J, Graham JS, Hougard JM: The role of agricultural use of insecticides in resistance to pyrethroids in Anopheles gambiae s.l in Burkina Faso. Am J Trop Med Hyg. 2002, 67: 617-622.
Ranson H, N'Guessan R, Lines J, Moiroux N, Nkuni Z, Corbel V: Pyrethroid resistance in African anopheline mosquitoes: what are the implications for malaria control?. Trends Parasitol. 2011, 27 (2): 91-98. 10.1016/j.pt.2010.08.004.
Zaim M, Aitio A, Nakashima N: Safety of pyrethroid-treated mosquito nets. Med Vet Entomol. 2000, 14: 1-5. 10.1046/j.1365-2915.2000.00211.x.
Global Malaria Programme WHO: Global plan on insecticide resistance management in malaria vectors (GPIRM). 2012, Geneva: World Health Organization
Coleman M, Hemingway J: Insecticide resistance monitoring and evaluation in disease transmitting mosquitoes. J Pest Sci. 2007, 32 (2): 69-76. 10.1584/jpestics.R07-01.
Hemingway J, Hawkes NJ, McCarroll L, Ranson H: The molecular basis of insecticideresistance in mosquitoes. Insect Biochem Mol Biol. 2004, 34 (7): 653-665. 10.1016/j.ibmb.2004.03.018.
Martinez Torres D, Chandre F, Williamson MS, Darriet F, Berge JB, Devonshire AL, Guillet P, Pasteur N, Pauron D: Molecular characterization of pyrethroid knockdown resistance (kdr) in the major malaria vector Anopheles gambiae s.s. Insect Mol Biol. 1998, 7 (2): 179-184. 10.1046/j.1365-2583.1998.72062.x.
Ranson H, Jensen B, Vulule JM, Wang X, Hemingway J, Collins FH: Identification of a point mutation in the voltage-gated sodium channel gene of Kenyan Anopheles gambiae associated with resistance to DDT and pyrethroids. Insect Mol Biol. 2000, 9 (5): 491-497. 10.1046/j.1365-2583.2000.00209.x.
Ranson H, Rossiter L, Ortelli F, Jensen B, Wang XL, Roth CW, Collins FH, Hemingway J: Identification of a novel class of insect glutathione S-transferases involvedin resistance to DDT in the malaria vector Anopheles gambiae. Biochem J. 2001, 359: 295-304. 10.1042/0264-6021:3590295.
Gunasekaran K, Muthukumaravel S, Sahu SS, Vijayakumar T, Jambulingam P: Glutathione S-transferase activity in Indian vectors of malaria: A defense mechanism against DDT. J Med Entomol. 2011, 48 (3): 561-569. 10.1603/ME10194.
Prapanthadara LA, Ketterman AJ: Qualitative and quantitative changes in glutathioneS-transferases in the mosquito Anopheles gambiae confer DDT-resistance. Biochem Soc Trans. 1993, 21 (3): 304S-
Vontas JG, Small GJ, Hemingway J: Glutathione S-transferases as antioxidant defence agents confer pyrethroid resistance in Nilaparvata lugens. Biochem J. 2001, 357: 65-72. 10.1042/0264-6021:3570065.
Kostaropoulos I, Papadopoulos AI, Metaxakis A, Boukouvala E, Papadopoulou-Mourkidou E: Glutathione S-transferase in the defence against pyrethroids in insects. Insect Biochem Mol Biol. 2001, 31 (4–5): 313-319.
Riveron JM, Yunta C, Ibrahim SS, Djouaka R, Irving H, Menze BD, Ismail HM, Hemingway J, Ranson H, Albert A, Wondji CS: A single mutation in the GSTe2 gene allows tracking of metabolically based insecticide resistance in a major malaria vector. Gen Biol. 2014, 15: R27-10.1186/gb-2014-15-2-r27.
Herath PR, Miles SJ, Davidson G: Fenitrothion (OMS 43) resistance in the taxon Anopheles culicifacies Giles. J Trop Med Hyg. 1981, 84 (2): 87-88.
Hemingway J: The genetics of malathion resistance in Anopheles stephensi from Pakistan. Trans R Soc Trop Med Hyg. 1983, 77 (1): 106-108. 10.1016/0035-9203(83)90030-5.
Feyereisen R, Lawrence IG, Kostas I, Sarjeet SG: Insect Cytochrome P450. Comprehensive Molecular Insect Science. 2005, Amsterdam: Elsevier, 1-77.
Muller P, Warr E, Stevenson BJ, Pignatelli PM, Morgan JC, Stephen A, Yawson E, Mitchell SN, Ranson H, Hemingway J, Paine MJ, Donnelly MJ: Field-caught permethrin-resistant Anopheles gambiae overexpress CYP6P3, a P450 that metabolises pyrethroids. PLoS Genet. 2008, 4: el000286-
Djouaka RF, Bakare AA, Coulibaly ON, Akogbeto MC, Ranson H, Hemingway J, Strode C: Expression of the cytochrome P450s,CYF6P3 and CYF6M2 are significantly elevated in multiple pyrethroid resistant populations of Anopheles gambiaes.s. from Southern Benin and Nigeria. BMC Genomics. 2008, 9: 538-10.1186/1471-2164-9-538.
Muller P, Donnelly MJ, Ranson H: Transcription profiling of a recently colonised pyrethroid resistant Anopheles gambiae strain from Ghana. BMC Genomics. 2007, 8: 3626-
Stevenson BJ, Bibby J, Pignatelli P, Muangnoicharoen S, O'Neill PM, Lian LY, Muller P, Nikou D, Steven A, Hemingway J, Sutcliff MJ, Paine MJ: Cytochrome P450 6 M2 from the malaria vector Anopheles gambiae metabolizes pyrethroids: Sequential metabolism of deltamethrin revealed. Insect Biochem Mol Biol. 2011, 41 (7): 492-502. 10.1016/j.ibmb.2011.02.003.
Muller P, Chouaibou M, Pignatelli P, Etang J, Walker ED, Donnelly MJ, Simard F, Ranson H: Pyrethroid tolerance is associated with elevated expression of antioxidants and agricultural practice in Anopheles arabiensis sampled from an area of cotton fields in Northern Cameroon. Mol Ecol. 2008, 17 (4): 1145-1155.
Mitchell SN, Stevenson BJ, Muller P, Wilding CS, Egyir-Yawson A, Field SG, Hemingway J, Paine MJ, Ranson H, Donnelly MJ: Identification and validation of a gene causing cross-resistance between insecticide classes in Anopheles gambiae from Ghana. Proc Natl Acad Sci U S A. 2012, 109 (16): 6147-6152. 10.1073/pnas.1203452109.
Gillies MT, Coetzee M: A supplement to the Anophelinae of Africa south of the Sahara (Afrotropical region). Johannesburg S Afr Inst Med Res. 1987, 55: 1-143.
Coetzee M, Crag M, Le Sueur D: Distribution of African malaria mosquitoes belonging to the Anopheles gambiae complex. Parasitol Today. 2000, 16: 74-77. 10.1016/S0169-4758(99)01563-X.
Minakawa N, Sonye G, Mogi M, Githeko A, Yan G: The effects of climatic factors on the distribution and abundance of malaria vectors in Kenya. J Med Entomol. 2002, 39: 833-841. 10.1603/0022-2585-39.6.833.
Verhaeghen K, Bortel WV, Roelants P, Backeljau T, Coosemans M: Detection of the East and West African kdr mutation in Anopheles gambiae and Anopheles arabiensis from Uganda using a new assay based on FRET/Melt Curve analysis. Malar J. 2006, 5: 16-10.1186/1475-2875-5-16.
Himeidan YE, Chen H, Chandre F, Donnelly MJ: Short report: permethrin and DDT resistance in the malaria vector Anopheles arabiensis from eastern Sudan. Am J Trop Med Hyg. 2007, 77: 1066-1068.
Abdalla H, Matambo TS, Koekemoer LL, Mnzava AP, Hunt RH, Coetzee M: Insecticide susceptibility and vector status of natural populations of Anopheles arabiensis from Sudan. Trans R Soc Trop Med Hyg. 2008, 102: 263-271. 10.1016/j.trstmh.2007.10.008.
Ndjemaï HNM, Patchoké S, Atangana J, Etang J, Simard F, Bilong Bilong CF, Reimer L, Cornel A, Lanzaro CG, Fondjo E: The distribution of insecticide resistance in Anopheles gambiae s.l populations from Cameroon: an update. Trans R Soc Trop Med Hyg. 2008, 103 (11): 1127-1138.
Kulkarni MA, Rowland M, Alifrangis M, Mosha FW, Matowo J, Malima R, Justin P, Kweka E, Lyimo I, Magesa S, Salanti A, Rau ME, Drakeley C: Occurrence of leucine to phenylalanine knockdown resistance (kdr) mutation in Anopheles arabiensis populations Tanzania detected by a high throughput SSOP-ELISA method. Malar J. 2006, 5: 56-10.1186/1475-2875-5-56.
Protopopoff N, Matowo J, Malima R, Kavishe R, Kaaya R, Wright A, West P, Kisinza W, Mosha FW, Kleinschmidt I, Rowland M: High level of resistance to pyrethroids and DDT and reduced susceptibility to bendiocarb in malaria vector Anopheles gambiae in North Western Tanzania. Malar J. 2013, 12: 149-10.1186/1475-2875-12-149.
Matowo J, Kitau J, Kabula B, Oxborough RM, Mosha FW, Rowland M: Dynamics of pyrethroid resistance and the frequency of kdr mutation in Anopheles arabiensis in rural villages of Lower Moshi, North-Eastern Tanzania. J Parasitol Vector Biol. 2014, 6 (3): 31-41.
Jones CM, Haji KA, Khatib BO, Bagi J, Mcha J, Devine GJ, Daley M, Kabula B, Ali AS, Majambere S, Ranson H: The dynamics of pyrethroid resistance in Anopheles arabiensis from Zanzibar and an assessment of the underlying genetic basis. Parasite Vectors. 2013, 6: 343-10.1186/1756-3305-6-343.
Nardini L, Christian RN, Coetzer N, Ranson H, Coetzee M, Koekemoer LL: Detoxification enzymes associated with insecticide resistance in laboratory strains of Anopheles arabiensis of different geographic origin. Parasite Vectors. 2012, 5: 113-10.1186/1756-3305-5-113.
Mouatcho JC, Munhenga G, Hargreaves K, Brooke BD, Coetzee M, Koekemoer LL: Pyrethroid resistance in a major African malaria vector Anopheles arabiensis from Mamfene, northern KwaZulu-Natal, South Africa. S Afr J Sci. 2009, 105: 127-131.
Universal Coverage Campaign - Final Impact. 2011,http://medatanzania.org/?%20p?=?ucc_final,
Matowo J, Kulkarni MA, Mosha FW, Oxborough R, Kitau J, Tenu F, Rowland M: Biochemical basis of permethrin resistance in Anopheles arabiensis from Lower Moshi, north-eastern Tanzania. Malar J. 2010, 9: 193-10.1186/1475-2875-9-193.
Nkya TE, Akhouayri I, Kisinza W, David JP: Impact of environment on mosquito response to pyrethroid insecticides: facts, evidences and prospects. Insect Biochem Mol Biol. 2013, 43: 407-416. 10.1016/j.ibmb.2012.10.006.
Rogers DW, Whitten MMA, Thailayil J, Soichot J, Levashina EA, Catteruccia F: Molecular and cellular components of the mating machinery in Anopheles gambiae females. Proc Natl Acad Sci U S A. 2008, 105 (49): 19390-19395. 10.1073/pnas.0809723105.
WHO: Test procedures for insecticide resistance monitoring in malaria vector mosquitoes. 2013, Geneva: World Health Organization
Scott JA, Brogdon WG, Collins FH: Identification of single specimens of the Anopheles gambiae complex by the polymerase chain reaction. Am J Trop Med Hyg. 1993, 49: 520-529.
Smyth GK L: Linear models for microarray data. Bioinformatics and Computational Biology Solutions using R and BIOCONDUCTOR. Edited by: Gentleman R, Carey V, Dudoit S, Irizarry R, Huber W. 2005, New York: Springer, 397-420.
Wu H, Kerr M, Cui X, Churchill G: MAANOVA: A Software Package for the Analysisof Spotted cDNA Microarray Experiments. The Analysis of Gene Expression Data. Edited by: Parmigiani G, Garrett E, Irizarry R, Zeger S. 2003, London: Springer, 313-341.
Schmittgen TD, Livak KJ: Analyzing real-time PCR data by the comparative C-T method. Nat Protoc. 2008, 3: 1101-1108. 10.1038/nprot.2008.73.
Witzig C, Parry M, Morgan JC, Irving H, Steven A, Cuamba N, Kerah-Hinzoumbé C, Ranson H, Wondji CS: Genetic mapping identifies a major locus spanning P450 clusters associated with pyrethroid resistance in kdr-free Anopheles arabiensis from Chad. Heredity. 2013, 110: 389-397. 10.1038/hdy.2012.112.
Christian RN, Strode C, Ranson H, Coetzer N, Coetzee M, Koekemoer L: Microarray analysis of a pyrethroid resistant African malaria vector, Anopheles funestus, from Southern Africa. Pestic Biochem Physiol. 2011, 99: 140-147. 10.1016/j.pestbp.2010.11.010.
Vontas J, David J-P, Nikou D, Hemingway J, Christophides GK, Louis C, Ranson H: Transcriptional analysis of insecticide resistance in Anopheles stephensi using cross-species microarray hybridization. Insect Mol Biol. 2007, 16: 315-324. 10.1111/j.1365-2583.2007.00728.x.
Baker D, Nolan T, Fischer B, Pinder A, Crisanti A, Russell S: A comprehensive gene expression atlas of sex- and tissue-specificity in the malaria vector, Anopheles gambiae. BMC Genomics. 2011, 12 (1): 296-10.1186/1471-2164-12-296.
Aurade RM, Jayalakshmi SK, Sreeramulu K: P-glycoprotein ATPase from the resistantpest, Helicoverpa armigera: purification, characterization and effect of various insecticides on its transport function. Biochim Biophys Acta. 2010, 1798: 1135-1143. 10.1016/j.bbamem.2010.02.019.
Bariami V, Jones CM, Poupardin R, Vontas J, Ranson H: Gene Amplification, ABC Transporters and Cytochrome P450s: Unraveling the Molecular Basis of Pyrethroid Resistance in the Dengue Vector. Aedes aegypti PLoS Negl Trop Dis. 2012, 6 (6): e1692-10.1371/journal.pntd.0001692. doi:10.1371/journal.pntd.0001692
Porretta D, Gargani M, Bellini R, Medici A, Punelli F, Urbanelli S: Defence mechanisms against insecticides temephos and diflubenzuron in the mosquito Aedes caspius: the P-glycoprotein efflux pumps. Med Vet Entomol. 2008, 22: 48-54. 10.1111/j.1365-2915.2008.00712.x.
Buss DS, McCaffery AR, Callaghan A: Evidence for p-glycoprotein modification of insecticide toxicity in mosquitoes of the Culex pipiens complex. Med Vet Entomol. 2002, 16: 218-222. 10.1046/j.1365-2915.2002.00365.x.
Roth CW, Holm I, Graille M, Dehoux P, Rzhetsky A, Wincker P, Weissenbach J, Brey PT: Identification of the Anopheles gambiae ATP-binding cassette transporter superfamily genes. Mol Cells. 2003, 15: 150-158.
Gahan LJ, Pauchet Y, Vogel H, Heckel DG: An ABC transporter mutation is correlated with insect resistance to Bacillus thuringiensis Cry1Ac toxin. PLoS Genet. 2010, 6: e1001248-10.1371/journal.pgen.1001248.
Broehan G, Kroeger T, Lorenzen M, Merzendorfer H: Functional analysis of the ATP-binding cassette (ABC) transporter gene family of Tribolium castaneum. BMC Gen. 2013, 14: 6-
Higgins CF, Linton KJ: The ATP switch model for ABC transporters. Nat Struct Mol Biol. 2004, 11: 918-926. 10.1038/nsmb836.
Linton KJ, Higgins CF: Structure and function of ABC transporters: the ATP switch provides flexible control. Pflugers Arch. 2007, 453: 555-567. 10.1007/s00424-006-0126-x.
George AM, Jones PM: Perspectives on the structure-function of ABC transporters: the Switch and Constant Contact Models. Prog Biophys Mol Biol. 2012, 109: 95-107. 10.1016/j.pbiomolbio.2012.06.003.
Wood OR, Hanrahan S, Coetzee M, Koekemoer LL, Brooke DB: Cuticle thickening associated with pyrethroid resistance in the major malaria vector Anopheles funestus. Parasite Vectors. 2010, 3: 67-10.1186/1756-3305-3-67.
Acknowledgements
We are grateful to Hyacinthe Toe, a PhD student at Liverpool School of Tropical Medicine for orienting JM in the laboratory for microarray analysis. Our sincere gratitude to the Vector group of Liverpool School of Tropical Medicine for their support and encouragement during microarray analysis; Professor Martin Donnelly conceived the loop design for microarray analysis.
We sincerely express our special thanks to Mr. Augustine Mtui, Rashid Athumani and Issa Masenga for the field larval collections and rearing; Grace Chuwa and Dickson Matowo for mosquito processing and preservation. This work was supported by the Malaria Capacity Development Consortium (MCDC) which is funded by The Wellcome Trust (Grant number WT084289MA).
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
JM conceived, designed, and performed the experiments; participated in data analysis and interpretation; wrote the first draft of the manuscript. CMJ participated in RNA extractions, labelling, hybridizations and scanning of the microarrays; contributed to data analysis and interpretation and reviewed the manuscript. BK contributed to sampling, processing and preservation of samples and review of the manuscript. KS processed mosquitoes and performed reverse transcription and qPCR. FM and MR critically revised the manuscript. HR provided technical support and facilitated the implementation of the microarrays work. DW co-designed the study, provided technical support, contributed to data analysis and interpretation and critically reviewed the manuscript. All authors have read and approved the final manuscript.
Electronic supplementary material
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Matowo, J., Jones, C.M., Kabula, B. et al. Genetic basis of pyrethroid resistance in a population of Anopheles arabiensis, the primary malaria vector in Lower Moshi, north-eastern Tanzania. Parasites Vectors 7, 274 (2014). https://doi.org/10.1186/1756-3305-7-274
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1756-3305-7-274