
Abstract
Differentiation of neural lineages from mesenchymal stem cells has raised the hope of generating functional cells as seed cells for nerve tissue engineering. As important gene regulators, microRNAs (miRNAs) have been speculated to play a vital role in accelerating stem cell differentiation and repairing neuron damage. However, miRNA roles in directing differentiation of stem cells in current protocols are underexplored and the mechanisms of miRNAs as regulators of neuronal differentiation remain ambiguous. In this study, we have determined that miR-218 serves as crucial constituent regulator in neuronal differentiation of adipose stem cells (ASCs) through Wnt signaling pathway based on comprehensive annotation of miRNA sequencing data. Moreover, we have also discovered that miR-218 and Fibroblast Growth Factor-2 (FGF2) modulate neuronal differentiation in a sequential manner. These findings provide additional understanding of the mechanisms regulating stem cell neuronal differentiation as well as a new method for neural lineage differentiation of ASCs.
Similar content being viewed by others

Introduction
Mesenchymal stem cells are the ideal candidates for regenerative medicine and tissue engineering1. Generating neuronal cells from stem cells is an attractive approach given the limited intrinsic capacity of neurons in repairing neural tissue. In vitro studies have shown that the mesenchymal stem cells could differentiate into mature neurons expressing neuronal specific markers after exposure to various chemical agents2,3,4. However, these chemical induction methods are usually of low efficiency and considerable cytotoxicity. Recently, gene therapy has developed to meet this challenge. The approach involves the use of multipotential cells such as bone marrow-derived mesenchymal stem cells (BMSCs), muscle-derived stem cells (MDSCs) and adipose-derived stem cells (ASCs), which are engineered to overexpress factors that are of crucial roles of neurogenesis for promoting neuronal differentiation5,6,7.
Gene expression and related function of stem cell are controlled by a newly discovered class of short 22 nucleotides Micro-RNAs (miRNAs). MiRNAs interact with complex signal transduction pathways, including those involved in neuronal formation and development, by regulating the protein translation of specific cellular mRNAs and mRNAs degradation8,9,10,11. In the last decade, there has been an increase in our understanding of the role of miRNAs in neuronal development and stem cell neuronal differentiation, where miRNAs have shown to be involved in important genes that control cell pluripotency. Meanwhile, Researchers endeavor to manipulate the expression of particular miRNAs in order to promote stem cells differentiation into neural progenitor cells or authentic neural cells12,13,14,15. For example, miR-146a has been shown to be a key regulator of stem cell survival when the cells were incubated with induced factor (Diazoxide) by targeting Fas in the NF-κB signaling16. Let-7i was shown to be a novel and potent inhibitor of neuronal differentiation that targeted Mash1 and Ngn1 by participating in Sox2–Lin28 pathway on neurogenic process17. MiR-124 facilitates the maintaince of the neuronal state by targeting the Specificity protein (Sp1) which suppressed in differentiated neurons18. It has also been reported that the overexpression of miR-9 promotes neuronal differentiation by targeting the Tlx1 transcription factor and DNA binding-2 inhibitor19. Wang et al. have suggested that JMJD1C represses stem cell neuronal differentiation at least partially by epigenetically sustaining miR-302 expression and thus JMJD1C knockdown is sufficient to trigger neuronal differentiation upon withdrawal of exogenous FGF20.
In this study, we have successfully induced neuronal differentiation of ASCs by Retinoic Acid (RA) treatment. To systemically monitor the expression of miRNAs during ASCs differentiation, we determined the miRNA expression profile of ASCs, incubated with or without RA by high-throughput deep sequencing using Applied Biosystems SOLiD System. After comprehensive analysis of miRNAs sequencing data (miRNA profiling), we found that miR-218 is specifically expressed in neuronal differentiation. Furthermore, our study highlighted an intricate gene regulatory network and pathway (Wnt signaling) which is in turn highly related with miR-218 expression. Consequently, our results showed that the interaction of miR-218 and Wnt signaling had a crucial role which efficiently facilitate the differentiation of ASCs. After confirming that the expression of miR-218 alone was not enough to differentiate ASCs into neuronal cells, we demonstrated that miR-218 and Fibroblast Growth Factor-2 (FGF2) together regulate the generation of neuronal cells from ASCs in a temporally sequential manner. Our study, for the first time has provided a new insight into the time-sequential regulation mechanisms of neuronal differentiation.
Results
MiRNA expression profile analysis revealed that the Wnt signaling pathway and miR-218 were crucial for neuronal differentiation of ASCs
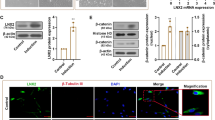
After 15 days of incubation with RA, the neurite outgrowth has been observed when the ASCs is differentiated into the neuronal lineage. The differentiation of ASCs into the neuronal lineage is confirmed by the expression of the neural terminal differentiation marker, βIII-Tubulin, using immunofluorescence (Fig. 1a). The protein expression of differentiation markers such as OCT4, SOX2, βIII-TUBULIN and MAP2 are also monitored at different time points (Day 0, Day 2, Day 5, Day 10 and Day 15) of RA treatment. (Figure 1b,c). The reduction of stemness markers (OCT4 and SOX2) is accompanied by enhancing the neural cell markers (βIII-TUBULIN and MAP2). The percentage of cells quantifies for this transformation (see Supplementary Figs S1 and S2). To elucidate the expression pattern of miRNAs during neuronal differentiation, high-throughput deep sequencing is performed using an Applied Biosystems SOLiD System. From the miRNA profiling results, about dysregulated 654 miRNAs are summarized. With the fold-change and Z-test analysis in the sequencing results, we have found that the expression levels of miRNAs are widely affected while the ASCs are differentiated into neuronal lineage and some miRNAs expression levels are more tempestuously regulated, including miR-146a, miR-196b, miR-31, miR-218, miR-214, miR-203, miR-124, miR-26a, miR-222, miR-375, miR-9, and let-7 family (Fig. 1d). The expression levels of some miRNAs are implicated in the development of neurons, such as miR-9, miR-214 and the let-7 family (the expression levels of miR-9, miR-146a and miR-214 are detected at 3 time points, see supplementary Fig. S3). Meanwhile, we evaluate the target genes of this miRNA pool by bioinformatics and subject to DAVID database. The functions of the target genes predicted by obviously altering miRNAs are annotated with KEGG signaling pathway analysis. From the P-Value analysis in the terms of the biological process, the Wnt signaling pathway (P - Value = 6.3) is likely to be critical for ASCs neuronal differentiation (Fig. 1e). The key gene expressions in Wnt signaling pathway (Wnt3a, Tcl4, Lef1, β-Catenin and Axin2 in Wnt/β-Catenin pathway) predicted with bioinformatics are validated by qRT-PCR (Fig. 1f). After addition of ICG-001 protein and subsequently adding RA for 15 days (anti-Wnt/RA group), Wnt signaling pathway is effectively inhibited by decreasing the expression levels of phosphorylation FZD (p-FZD) and β-CATENIN (Fig. 1g,h). As expected, OCT4 and SOX2 protein levels are unchanged and βIII-TUBULIN is undetectable in anti-Wnt/RA group (Fig. 1i,j).

(a) Immunofluorescence photomicrograph image of βIII-TUBULIN (red) and HOECHST (HOE, blue) staining in ASCs of non-treated cells (-RA group) and RA-treated cells (+RA group) for 15 days (D 15). (b) Western blotting of OCT4, SOX2, βIII-TUBULIN and MAP2 in the –RA and +RA groups at different time points and GAPDH is used as control. (c) Proteins expression levels, quantified by determining the gray value. (d) The expression ratios and evaluation, based on the mathematical model (using Fold-change and Z-test method), for all-known miRNAs that are detected between -RA and +RA groups. (e) Top KEGG pathways are summed up from the DAVID database based on P-value. (f) Expressions of key gene (Wnt3a, Tcf4, Lef1, β-Catenin and Axin2) in Wnt signaling pathway are detected by qRT-PCR between -RA and +RA groups (** p < 0.01, n = 4). (g) Western blotting analysis of Wnt signaling pathway marker proteins (FZD and phosphorylated FZD in the cytoplasm and the β-CATENIN in the nuclear) in control group (non-treated), RA group (RA-treated for 15 days) and anti-Wnt/RA group (pre-treated by the inhibitor of Wnt signaling pathway, ICG-001 and the group is incubated with RA for 15 days), GAPDH is used as control. (h) The proteins expression levels, quantified by determining the gray values (n = 2). (i) Western blotting analysis of the stem cell stemness marker (OCT4 and SOX2) and neural cell marker (βIII-TUBULIN) expression in control group, RA group and anti-Wnt/RA group and GAPDH is used as control. (j) Proteins expression levels, quantified by determining the gray value (n = 2).
We further investigate the Wnt signaling pathway genes. From KEGG analysis results, the red pentacles reveal the key genes closely related to the ASCs neuronal differentiation process (Fig. 2a). The genes involving in Wnt signaling (red pentacles) or regulation of Wnt signaling (blue pentacles) are targeted by miRNAs (blue circles). From our sequencing and predicted data, the top level among the differentially expressed miRNAs is highlighted and reveals that miR-218 has significantly up-regulation after RA treatment (Z-test = 42.3, Fig. 2b), which is accord with the previous reports about the key regulator in Wnt Signaling21,22,23. Therefore, miR-218 is considered to be crucial for ASCs neuronal differentiation. Indeed, we find RA supplementation in culture medium increases the endogenous miR-218 expression by almost 8.5-fold (Fig. 2c) and downregulates the expression of the OCT4 and SOX2 simultaneously (Fig. 2d,e). However, after anti-miR-218 transfection, subsequent RA treatment does not enhance βIII-TUBULIN expression (in anti-miR-218/RA group, Fig. 2d,e).

(a) In the +RA group (RA-treated group), based on the database prediction (TargetScan version 6.2 database) and the KEGG analysis, the genes involved in the Wnt signaling pathway (frame fill with green) and several key genes (red pentacles) in the Wnt signaling pathway. (b) Diagram showing the link between the key miRNAs and its target genes which are involved in or targeted on the regulation of Wnt signaling pathway. The genes included in Wnt signaling pathway (red pentacles) and regulated Wnt signaling (blue pentacles) are targeted by miRNAs (blue circles) and the key miRNAs (miR-218, miR-29a and miR-200 et al.) are summarized according the Z-test ranking results. MiR-218 (Z-test = 42.3) is found to be in the front rank. (c) Relative miR-218 expression for the control group (RA-non-treated cells), RA group (RA-treated for 15 days) and anti-miR-218/RA group (ASCs are transfected by anti-miR-218 plasmids for 48 h and RA treated for 15 days) are determined by qRT-PCR (**p < 0.01, n = 8). (d) Western blotting analysis of OCT4, SOX2 and βIII-TUBULIN expression in the control group, RA group and anti-miR-218/RA group and GAPDH is used as control. (e) Protein expression levels, quantified by determining the gray value (n = 3).
Taken together, these data demonstrated that the Wnt signaling pathway and miR-218 both participate and positively promote ASCs neuronal differentiation.
MiR-218 regulates Wnt signaling pathways but is insufficient to induce ASCs differentiation into neural cells
MiR-218 is specifically active in developing motor neurons. The robust upregulation of miR-218 in ASCs, differentiate ASCs into the neural lineage inspires us to investigate whether over expressing miR-218 may induce the neuronal differentiation of ASCs through Wnt signaling pathway. The miR-218 transfection markedly increases the endogenous miR-218 levels by almost 100-fold (Fig. 3a). While the transfection of anti-miR-218 significantly decrease the miR-218 expression. The expression of miR-218 target genes like Robo1, Robo2 and Lamb3 and the Wnt signaling pathway antagonist genes such as Sfrp2 and Dkk2 are validated by qRT-PCR with the cells transfected with miR-218 and anti-miR-218 (Fig. 3b). In comparison to the transfection of anti-miR-218 and the controls (miR-NC), transfection of miR-218 dramatically enhances p-FZD levels and elevates nuclear accumulation of β-CATENIN (Fig. 3c,d). These results indicate that miR-218 transfection can activate Wnt signaling pathway. Meanwhile, it seems that the anti-Wnt does not have any effect on the expression of miR-218 (see Supplementary Fig. S4).

(a) The relative miR-218 expression levels for ASCs transfected with control plasmid (miR-NC group), miR-218 plasmid (miR-218 group) and anti-miR-218 plasmid (anti-miR-218 group) after 48 h are detected by qRT-PCR (**p < 0.01, n = 4). (b) The qRT-PCR results of some miR-218 target (Robo1, Robo2, Lamb3, Sfrp2 and Dkk2) mRNAs expression, detected in miR-NC group, miR-218 group and anti-miR-218 group. (c) Western blotting results of Wnt signaling pathway markers (FZD and p-FZD in the cytoplasm and β-CATENIN in the nuclear) in the miR-NC group, miR-218 group and anti-miR-218 group and GAPDH is used as control. (d) Proteins expression levels, quantified by determining the gray value (n = 2). (e) The qRT-PCR results of Oct4, Sox2, βIII-Tubulin, Map2 and Nestin mRNA expression levels in the RA-treated group (RA group) and three plasmid treated groups (miR-NC group, miR-218 group and anti-miR-218 group). (f) Western blotting results of βIII-TUBULIN, MAP2 and NESTIN in the four groups (RA group, miR-NC group, miR-218 group and anti-miR-218 group) and GAPDH is used as control. (g) Protein expression level, quantified by determining the gray value (n = 2).
However, both the mRNAs levels of Oct4, Sox2, βIII-Tubulin, Map2 and Nestin (Fig. 3e) and the protein expression studies (Fig. 3f,g) prove that, overexpression of miR-218 alone cannot induce βIII-Tubulin, Map2 and Nestin expression in the absence of RA. These results confirm that, although miR-218 positively regulates Wnt signaling pathway, which alone is insufficient to induce ASCs differentiation into neural cells.
FGF2 and miR-218 co-operate sequentially in ASCs neural differentiation
Previous studies demonstrate that the FGF signaling pathway participates in neurogenesis and central nervous system formation24,25,26. In our earlier studies, we identified that addition of FGF2 (10 ng/mL) may work as a pre-induction factor and affect ASCs neuronal differentiation27. Herein, we speculate that FGF2 may interact with miR-218 to induce neuronal differentiation. To confirm this, we pre-treat ASCs with FGF2 (10 ng/mL) for 10 days followed by transfecting with miR-218. Subsequently, elevation of miR-218 levels in ASCs (+FGF2/miR-218 group) increases the expression of Wnt signaling pathway markers (p-FZD and β-CATENIN) in the +FGF2/miR-218 group compared to the -FGF2/-miR-218 and +FGF2/anti-miR-218 groups (Fig. 4a,b). This indicates, FGF2 and miR-218 work synergistically for enhancing Wnt signaling pathway. The morphological transformation to neural-like cells and the expression of βIII-Tubulin are confirmed by immunofluorescent imaging and photomicrograph (Fig. 4c). At the same time, the two-color Flow Cytometry (flow cytometric dot plots) shows, there is an increase (0.1% to 41.4%) of double-positive (βIII-Tubulin + cells) cells in +FGF2/miR-218 group compared to +FGF2/anti-miR-218 group (Fig. 4d). These results indicate, the temporal relationship between FGF2 and miR-218 on the neuronal differentiation. The FGF2 pretreatment cooperatively interacts with miR-218 to induce ASCs into neural lineage.

(a) Western blotting analysis of three markers of Wnt signaling pathway in –FGF2/-miR-218 group (without any FGF2 or plasmid treated), +FGF2/anti-miR-218 group (ASCs were incubated with 10 ng/mL FGF2 for 10 days and transfected with anti-miR-218 plasmid for 48 h), +FGF2/miR-218 group (ASCs were incubated with 10 ng/mL FGF2 for 10 days and transfected with miR-218 plasmid for 48 h) and GAPDH is used as control. (b) Proteins expression levels, quantified by determining the gray value (n = 3). (c) Immunofluorescence image of βIII-TUBULIN (red) and HOE (blue) staining in two groups (+FGF2/anti-miR-218 group and +FGF2/miR-218 group). Photomicrograph image of ASCs in +FGF2/miR-218 group. Scale bars, 20 μm. (d) The ASCs neuronal differentiation (βIII-TUBULIN positive expression in the +FGF2/anti-miR-218 group and +FGF2/miR-218 group) are performed by two-color flow cytometric analysis. The dot blot and data table (the red frame in the quadrants and red section in the column diagram) shows the proportion of the βIII-TUBULIN positive expression in +FGF2/anti-miR-218 group and +FGF2/miR-218 group. (e) Schematic illustration and timelines of ASCs cultured with FGF2 and miR-218 shows the sequential regulation method. ASCs are transfected with miR-218 plasmid. After 48 h (D 2), cells are maintained in medium supplemented with 10 ng/mL FGF2 for 10 days (D 12) (miR-218/+FGF2 group). (f) Western blotting analysis for OCT4, SOX2 and βIII-TUBULIN protein in miR-218/+FGF2, anti-miR-218/+FGF2 and GAPDH is used as control. (g) Proteins expression levels, quantified by determining the gray value (n = 2).
To evaluate in sequential manner, we treat cells with FGF2 followed by transfecting with miR-218 (miR-218/+FGF2 group) or anti-miR-218 (anti-miR-218/+FGF2 group) (Fig. 4e). Interestingly, neither morphological changes nor βIII-TUBULIN expression can be detected (Fig. 4f,g) in the above treated cells. These results indicate that pre-induction with FGF2 is necessary to facilitate the neuronal induction effect of miR-218 in ASCs. In addition, we find that miR-218 and FGF2 does not form the negative feedback loop (see Supplementary Fig. S5).
Taken together, our data indicate, elevation of FGF2 and miR-218 cooperatively induces stem cells to differentiate in a temporally sequential manner via the Wnt signaling pathway.
Discussion
Micro-RNAs inhibit translation and mediate mRNA decay through sequence-specific base pairings with specific region of target genes. Researchers endeavor to manipulate the expression of particular miRNAs in order to promote stem cells differentiation into neural progenitor cells or authentic neural cells28,29,30. In this study, we analyzed miRNA expression profiling that significantly changed when ASCs were treated with RA for neuronal differentiation. We identified that certain miRNAs exhibited tremendous changes in their expression during differentiation (including let-7 family, miR-146a, miR-196b, miR-218, miR-214, miR-203, miR-124, miR-26a, miR-222, miR-375, miR-9), which have been shown to facilitate neurogenesis15,31,32,33,34,35,36.
Similarly, we predicted from bioinformatics analysis37,38,39, and then found, that Wnt signaling pathway was closely involved and played a pivotal role in the neural differentiation process of ASCs. In the Wnt signaling pathway (involving Fzd/β-Catenin pathway), the combination of Wnt proteins and the receptors led to an increase in activity of glycogen synthase kinase 3β (Gsk3β) and Axin2. Then, the β-Catenin undergoes a nuclear translocation where it accumulated and formed complexes with transcription factors, activating a number of intracellular signaling pathways40,41,42. Previous evidence revealed that Wnt signaling pathway promoted stem cell self-renewal and participated in neurogenesis43. Wiggan et al. hypothesized that, during early placode development, Pax3 and Sox2 activated Wnt signaling pathway through the enhancer N-1c44. Their research revealed that, the direct involvement of Wnt signaling in the initiation of neural plate development. Studies by Elizalde et al. indicated that Wnt-4 and Wnt-11 (Wnt signaling pathway regulator) promoted early neuronal differentiation by diminishing the RA-induced downregulation of Oct4 and Nanog and the upregulation of Pax6, Ascl1, Hoxc5, and Neurod145.
Furthermore, certain miRNAs have shown to be involved with important genes that controlled the cell pluripotency and mediated the induction of pluripotent stem cells by targeting the Wnt signaling pathway. For example, considerable evidence suggested that the Wnt signaling pathway has been regulated by miR-499, miR-355, miR-375, miR-27, miR-29, miR-17, miR-142 and miR-21846,47,48,49,50,51,52,53,54. The Dkk2, Sfrp2 and Sost are reported as the Wnt signaling pathway inhibitors and miR-218 targets these inhibitors as a positive feedback loop with Wnt signaling pathway23,55. Based on the sequencing data, we selected miR-218 for further investigation, not only because of its extreme differential expression in comparison to other miRNAs (Fig. 1d) but mainly of its involvement in the Wnt signaling pathway (Fig. 2b–d).
To further support the hypothesis that Wnt signaling pathway and miR-218 were closely involved in the process of ASCs neural differentiation, we used anti-miR-218 and the inhibitor ICG-001 to block ASCs neuronal differentiation and supplemented the ASCs culture medium with RA. Our experimental results verified the predictive identification of miR-218 and Wnt signaling in neural differentiation and using our modified bioinformatics analysis method further demonstrated their function as key cellular triggers of neuronal differentiation. Interestingly, miR-218 did not appear to play an instructive role neither in mesenchymal stem cell fate determination nor in motor neuron fate determination on its own. Given that the overexpression of miR-218 alone was not sufficient to induce the formation of motor neurons in chick neural tube or mouse embryonic stem cells (ESCs) and to induce the neuronal differentiation of ASCs as well56,57. Therefore, we speculated that there might exist a method of combinative regulation in the ASCs neuronal differentiation.
FGF2 belongs to the family of heparin-binding growth factors and has been described as a mitotic activator in the stem cells differentiations27,58. The FGF2 treatment of cultured stem cells provided mitogenic support and predominated in the induction. In the present study, we included FGF2 in the culture medium as a pre-induction factor for neural differentiation. With this pre-induction and the overexpression of miR-218, Wnt signaling pathway was stimulated to a greater extent compare to other conditions such as +FGF2/anti-miR-218 or miR-218. Additionally, we demonstrated that, synergistically, the supplementation of FGF2 and overexpression of miR-218 prompted the differentiation of ASCs into neural cells (Fig. 4c,d).
A number of studies have investigated the effects of various inductive factors at different stages of differentiation in in-vitro. For instance, Tang et al. revealed that appropriate timing of Wnt-1 expression was necessary for the appropriate RA-induced expression of the neural phenotype in P19 cells59. Handorf et al. delineated two stages of chondrogenesis and identified that developmental days 9–12 represented an important regulatory point in the chondrogenic program of hMSCs by examining the sequential administration of TGFB1, BMP7, and IGF1 within specific temporal windows60. Similarly, the major principle of stem cells neuronal differentiation may have a multi-step process, and that cells exhibit spatial and temporal responses to signaling pathways and their regulators (i.e., miRNAs). Therefore, we classified mesenchymal stem cells neural differentiation could be divided into two sequential stages: “induction” and “differentiation”. The delivery of FGF2 was used for the induction of differentiation in the initial step, during which ASCs displayed a stage of increased activity triggered by FGF2 and were “conditioned” for the subsequent elevation of miR-218. Finally, we validated the sequential link between bFGF (FGF2) and miR-218 during ASC neural differentiation. Further investigations of the molecular mechanisms underpinned this link and the synergistic effects of FGF2 and miR-218 on the Wnt signaling pathway during neuronal differentiation were warranted.
In this study, we identified that Wnt signaling pathway and miR-218 were closely related to ASCs neuronal differentiation. We also demonstrated that miR-218 overexpression alone failed to induce ASCs neuronal differentiation and FGF2 pretreatment cooperatively interacted with miR-218 to generate neural cells. Furthermore, FGF2 and miR-218 were shown to operate in a temporally sequential manner to promote the differentiation of ASCs into the neural lineage. Our results augmented, the current understanding of the developmental processes of neural differentiation and provided important insights into how miRNAs contributed to this process, which could assist the development of novel inductive approaches for neural tissue regeneration.
Materials and Methods
ASCs isolation and differentiation
ASCs were obtained from four-week old female Sprague-Dawley rats (weight 100–130 g), as previously described61. The use of all animal samples were approved by and carried out in accordance to the medical ethics committee of Southeast University, China. ASCs were digested and seeded. The cells were cultured in basal medium composed of Dulbecco’s modified Eagle’s medium (DMEM; Thermo Fisher Scientific, USA), 5% fetal bovine serum (FBS; Gibco Lab., USA), 1% penicillin/streptomycin (Thermo Fisher Scientific) with or without FGF2 (Peprotech, USA). The medium was replaced every 3 days for a period of 10 days. ASCs were then seeded into 12-well plates and neuronal differentiation (RA-treated, +RA group) was performed over a period of 15 days using basal medium supplemented with 100 ng/mL Retinoic Acid (RA; Sigma Aldrich, USA). The non-treated group (control, −RA group) was cultured contemporaneously.
MiRNA analysis
Total RNA were extracted from RA-treated and non-treated cells and altered miRNAs expression were detected by sequencing of the Applied Biosystems SOLiD System, as previously described37,62,63. Altered expression of miRNAs and prediction of effects on their target genes were analyzed following the method (based on the Z-test calculation method, a mathematical model to evaluate the comprehensive repression rate of specific mRNAs using total miRNA expression profiling) reported previously37,64,65. Briefly, the target genes levels between RA-treated (+RA group) and non-treated (−RA group) cells were predicted by the tools of TargetScan version 6.2 database. The identified lists of genes were subjected to functional annotation, clustering and analysis using the KEGG analysis based on the Database for Annotation, Visualization and Integrated Discovery (DAVID) Bioinformatics Database (https://david.ncifcrf.gov/).
Quantitative real-time PCR
The miRNAs and mRNAs expression were measured by quantitative Real-Time PCR (qRT-PCR) using ABI 7500 System (Life Technologies, USA) and standard protocols (All the Primers are listed in Supplementary Table S1 online). Relative amounts were determined using the 2−ΔΔCt method.
ICG-001 treatment
ICG-001 protein inhibits the Wnt signaling pathway by binding to the element-binding protein (CBP)66. ASCs were seeded into 12-well plates and maintained at CO2 incubator. After 24 h of seeding, the final concentrations of 5 μM ICG-001 (Selleckchem, USA) was added into the ASCs medium for 2 days, followed by 100 ng/mL of RA was added and further cultured for 15 days (anti-Wnt/RA group).
Western blotting
Total cellular proteins and nuclear proteins were separately extracted using the Total Protein Isolation Kit (Sangon Company, China) and Nuclear Protein Isolation Kit (Sangon Company). Proteins were quantified by Bicinchoninic acid assay kit (Sangon Company). The protein samples were loaded on SDS–PAGE gels and electrophoresed under standard conditions. Western blotting was performed using nitrocellulose membranes. After blocking, membranes were incubated with primary antibodies (1:200–1:500) at 4 °C overnight. After rinsing, incubation was conducted with secondary horseradish peroxidase-conjugated goat anti-rabbit or mouse IgG antibody (Bioss Biotechnology, China) and exposed to film. Primary antibodies included anti-Frizzled (FZD, Immunoway, Catalog: YT1783, USA), anti-phospho-Frizzled (p-FZD, Immunoway, Catalog: YP0173), anti-β-Catenin (Boster Biotechnology, Catalog: BM1575, China), anti-Oct4 (Santa Cruz Biotechnology, Catalog: sc-5279, USA), anti-Sox2 (Santa Cruz Biotechnology, Catalog: sc-17320), anti-βIII-Tubulin (Santa Cruz Biotechnology, Catalog: sc-58888), anti-Map2 (Santa Cruz Biotechnology, Catalog: sc-20172), anti-Nestin (Santa Cruz Biotechnology, Catalog: sc-20978), and anti-Gapdh (Cell Signaling Technology, Catalog: #2118, USA).
MiRNA transfection
ASCs were seeded into 12-well plates, after 24h of seeding, 5 μL Superfectin Transfection Reagent (Qiagen, Canada) was added per well, according to the manufacturer’s instructions. Transfections were performed using 20 nM of miR-NC, anti-miR-218 or miR-218, respectively. The miRNA plasmids were designed and enhanced green fluorescence protein (eGFP) was used as a reporter gene. After 48 h, transfected cells and transfection efficiency was measured and assayed by detection of eGFP. In the anti-miR-218/RA group, the anti-miR-218 plasmids were transfected for 2 days and the ASCs medium was supplemented with 100 ng/mL RA for 13 days. The βIII-tubulin positive expressions were analyzed by Flow Cytometry using C-Flow software.
FGF2 pretreatment
ASCs medium was supplemented with 10 ng/mL FGF2 for 10 days, followed by transfected with miR-NC, anti-miR-218 or miR-218 plasmids for 2 days. In order to evaluate the temporal relationship, the chronological sequence was exchanged.
Flow cytometry
Cells were trypsinized with 0.25% trypsin solution (Sigma Aldrich) and fixed with 4% formaldehyde (Sangon Company) in PBS at 37 °C for 0.5 h. After rinsing, the cells were incubated with rabbit anti-rat βIII-Tubulin primary antibody (1:200 in 1% BSA; Santa Cruz Biotechnology) at 37 °C for 4 h then incubated with the appropriate amount of the secondary antibody, goat anti-rabbit Alexa-Fluor 647 (1:500 in 1% BSA; Invitrogen, USA) for 1 h at 37 °C. During the whole experiment the cells were protected from light. Finally, the cell samples were subjected to Flow Cytometry and corresponding data were analyzed by C-Flow software.
Immunofluorescent analysis
After induction of RA and/or miR-218 for a period of time, ASCs were subjected to immunofluorescent staining by rabbit anti-rat βIII-Tubulin primary antibody to detect differentiated neural cells. After washing with PBS, the samples were incubated with the appropriate secondary antibody, mouse anti-rabbit Alexa-Fluor 647 (1:200 in 1% BSA; Cell Signaling Technology) in the dark. Then samples were washed twice with PBS, the nuclei stained with 10 μg/mL Hoechst 33342 (HOE, Sigma Aldrich,) for 0.5 h, and images were obtained using a Revolution XD confocal laser scanning microscope (Andor, Belfast, Northern Ireland).
Statistical analysis
All data were expressed as mean ± SD. Differences were compared using the Student’s t-test; p-values < 0.05 were considered statistically significant (*p < 0.05, **p < 0.01).
Additional Information
How to cite this article: Hu, F. et al. MiR-218 Induces Neuronal Differentiation of ASCs in a Temporally Sequential Manner with Fibroblast Growth Factor by Regulation of the Wnt Signaling Pathway. Sci. Rep. 7, 39427; doi: 10.1038/srep39427 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Mizuno, H., Tobita, M. & Uysal, A. C. Concise Review: Adipose-Derived Stem Cells as a Novel Tool for Future Regenerative Medicine. Stem Cells 30, 804–810 (2012).
Woodbury, D., Schwarz, E. J., Prockop, D. J. & Black, I. B. Adult Rat and Human Bone Marrow Stromal Cells Differentiate Into Neurons. J Neurosci Res (2000).
Safford, K. M., Safford, S. D., Gimble, J. M., Shetty, A. K. & Rice, H. E. Characterization of neuronal/glial differentiation of murine adipose-derived adult stromal cells. Exp Neurol 187, 319–328, doi: 10.1016/j.expneurol.2004.01.027 (2004).
Kim, S. et al. Neural differentiation potential of peripheral blood- and bone-marrow-derived precursor cells. Brain Res 1123, 27–33, doi: 10.1016/j.brainres.2006.09.044 (2006).
Jung, J. S. et al. MBD6 is a direct target of Oct4 and controls the stemness and differentiation of adipose tissue-derived stem cells. Cell Mol Life Sci 70, 711–728, doi: 10.1007/s00018-012-1157-4 (2013).
Billon, N. et al. Cooperation of Spl and p300 in the induction of the CDK inhibitor p21(WAF1/CIP1) during NGF-mediated neuronal differentiation. Oncogene 18, 2872–2882, doi: 10.1038/sj.onc.1202712 (1999).
Cheng, F. et al. Neurogenin 2 Converts Mesenchymal Stem Cells into a Neural Precursor Fate and Improves Functional Recovery after Experimental Stroke. Cellular Physiology and Biochemistry 33, 847–858, doi: 10.1159/000358657 (2014).
He, L. & Hannon, G. J. Micrornas: Small RNAs with a big role in gene regulation. Nature Reviews Genetics 5, 522–531, doi: 10.1038/nrg1379 (2004).
Stappert, L., Roese-Koerner, B. & Bruestle, O. The role of microRNAs in human neural stem cells, neuronal differentiation and subtype specification. Cell and Tissue Research 359, 47–64, doi: 10.1007/s00441-014-1981-y (2015).
Collino, F., Bruno, S., Deregibus, M. C., Tetta, C. & Camussi, G. In Vitamins and Hormones: Stem Cell Regulators Vol. 87 Vitamins and Hormones (ed G. Litwack ) 291–320 (2011).
Gangaraju, V. K. & Lin, H. MicroRNAs: key regulators of stem cells. Nature Reviews Molecular Cell Biology 10, 116–125 (2009).
Cui, Y. et al. MiR-125b orchestrates cell proliferation, differentiation and migration in neural stem/progenitor cells by targeting Nestin. Bmc Neuroscience 13, doi: 10.1186/1471-2202-13-116 (2012).
Tay, Y., Zhang, J. Q., Thomson, A. M., Lim, B. & Rigoutsos, I. MicroRNAs to Nanog, Oct4 and Sox2 coding regions modulate embryonic stem cell differentiation. Nature 455, 1124–U1112, doi: 10.1038/Nature07299 (2008).
Card, D. A. G. et al. Oct4/Sox2-regulated miR-302 targets cyclin D1 in human embryonic stem cells. Molecular and Cellular Biology 28, 6426–6438, doi: 10.1128/Mcb.00359-08 (2008).
Kosik, K. S. & Krichevsky, A. M. The elegance of the microRNAs: A neuronal perspective. Neuron 47, 779–782, doi: 10.1016/j.neuron.2005.08.019 (2005).
Suzuki, Y., Kim, H. W., Ashraf, M. & Haider, H. K. Diazoxide potentiates mesenchymal stem cell survival via NF-κB-dependent miR-146a expression by targeting Fas. American Journal of Physiology-Heart and Circulatory Physiology 299, H1077–H1082 (2010).
Cimadamore, F., Amador-Arjona, A., Chen, C., Huang, C. T. & Terskikh, A. V. SOX2-LIN28/let-7 pathway regulates proliferation and neurogenesis in neural precursors. P Natl Acad Sci USA 110, E3017–E3026, doi: 10.1073/pnas.1220176110 (2013).
Mondanizadeh, M. et al. MicroRNA-124 Regulates Neuronal Differentiation of Mesenchymal Stem Cells by Targeting Sp1 mRNA. Journal of Cellular Biochemistry 116, 943–953, doi: 10.1002/jcb.25045 (2015).
Zhao, C., Sun, G., Ye, P., Li, S. & Shi, Y. MicroRNA let-7d regulates the TLX/microRNA-9 cascade to control neural cell fate and neurogenesis. Scientific reports 3, 1329, doi: 10.1038/srep01329 (2013).
Wang, J. L., Park, J. W., Drissi, H., Wang, X. F. & Xu, R. H. Epigenetic Regulation of miR-302 by JMJD1C Inhibits Neural Differentiation of Human Embryonic Stem Cells. Journal of Biological Chemistry 289, 2384–2395, doi: 10.1074/jbc.M113.535799 (2014).
Tu, Y. Y. et al. MicroRNA-218 Inhibits Glioma Invasion, Migration, Proliferation, and Cancer Stem-like Cell Self-Renewal by Targeting the Polycomb Group Gene Bmi1. Cancer Research 73, 6046–6055, doi: 10.1158/0008-5472.can-13-0358 (2013).
Zhang, W. B., Zhong, W. J. & Wang, L. A signal-amplification circuit between miR-218 and Wnt/beta-catenin signal promotes human adipose tissue-derived stem cells osteogenic differentiation. Bone 58, 59–66, doi: 10.1016/j.bone.2013.09.015 (2014).
Liu, Y. et al. MiR-218 reverses high invasiveness of glioblastoma cells by targeting the oncogenic transcription factor LEF1. Oncol Rep 28, 1013–1021, doi: 10.3892/or.2012.1902 (2012).
Xu, W., Wang, X., Xu, G. & Guo, J. Basic fibroblast growth factor expression is implicated in mesenchymal stem cells response to light-induced retinal injury. Cell Mol Neurobiol 33, 1171–1179, doi: 10.1007/s10571-013-9983-y (2013).
Wang, Y. et al. Efficient derivation of human embryonic stem cell lines from discarded embryos through increases in the concentration of basic fibroblast growth factor. Human Cell 25, 16–23, doi: 10.1007/s13577-011-0039-7 (2012).
Suzuki, Y., Yanagisawa, M., Yagi, H., Nakatani, Y. & Yu, R. K. Involvement of beta 1-Integrin Up-regulation in Basic Fibroblast Growth Factor- and Epidermal Growth Factor-induced Proliferation of Mouse Neuroepithelial Cells. Journal of Biological Chemistry 285, 18443–18451, doi: 10.1074/jbc.M110.114645 (2010).
Hu, F. H. et al. Effects of Epidermal Growth Factor and Basic Fibroblast Growth Factor on the Proliferation and Osteogenic and Neural Differentiation of Adipose-Derived Stem Cells. Cell Reprogram 15, 224–232, doi: 10.1089/cell.2012.0077 (2013).
Annibali, D. et al. A New Module in Neural Differentiation Control: Two MicroRNAs Upregulated by Retinoic Acid, miR-9 and-103, Target the Differentiation Inhibitor ID2. Plos One 7, doi: 10.1371/journal.pone.0040269 (2012).
Neo, W. H. et al. MicroRNA miR-124 Controls the Choice between Neuronal and Astrocyte Differentiation by Fine-tuning Ezh2 Expression. Journal of Biological Chemistry 289, 20788–20801, doi: 10.1074/jbc.M113.525493 (2014).
Zhang, L. et al. Isoflurane Inhibits Embryonic Stem Cell Self-Renewal and Neural Differentiation Through miR-9/E-cadherin Signaling. Stem cells and development 24, 1912–1922, doi: 10.1089/scd.2014.0397 (2015).
Chen, H. et al. miR-7 and miR-214 are specifically expressed during neuroblastoma differentiation, cortical development and embryonic stem cells differentiation, and control neurite outgrowth in vitro . Biochem Bioph Res Co 394, 921–927, doi: 10.1016/j.bbrc.2010.03.076 (2010).
Asli, N. S. & Kessel, M. Spatiotemporally restricted regulation of generic motor neuron programs by miR-196-mediated repression of Hoxb8. Developmental Biology 344, 857–868, doi: 10.1016/j.ydbio.2010.06.003 (2010).
Rajaram, K., Harding, R. L., Hyde, D. R. & Patton, J. G. miR-203 regulates progenitor cell proliferation during adult zebrafish retina regeneration. Developmental Biology 392, 393–403, doi: 10.1016/j.ydbio.2014.05.005 (2014).
Visvanathan, J., Lee, S., Lee, B., Lee, J. W. & Lee, S.-K. The microRNA miR-124 antagonizes the anti-neural REST/SCP1 pathway during embryonic CNS development. Gene Dev 21, 744–749 (2007).
Forrest, A. R. et al. Induction of microRNAs, mir-155, mir-222, mir-424 and mir-503, promotes monocytic differentiation through combinatorial regulation. Leukemia 24, 460–466 (2009).
Kosik, K. S. The neuronal microRNA system. Nature Reviews Neuroscience 7, 911–920, doi: 10.1038/nrn2037 (2006).
Xu, P., Zhu, Y., Sun, B. & Xiao, Z. Colorectal cancer characterization and therapeutic target prediction based on microRNA expression profile. Scientific reports 6, doi: 10.1038/srep20616 (2016).
Sun, B., Liu, R., Ye, N. & Xiao, Z. D. Comprehensive evaluation of microRNA expression profiling reveals the neural signaling specific cytotoxicity of superparamagnetic iron oxide nanoparticles (SPIONs) through N-methyl-D-aspartate receptor. PLoS One 10, e0121671, doi: 10.1371/journal.pone.0121671 (2015).
Sun, B., Yang, F., Hu, F. H., Huang, N. P. & Xiao, Z. D. Comprehensive annotation of microRNA expression profiles. Bmc Genet 14, 120 (2013).
Aman, A. & Piotrowski, T. Wnt/β-catenin and Fgf signaling control collective cell migration by restricting chemokine receptor expression. Developmental cell 15, 749–761 (2008).
Baker, J. C., Beddington, R. S. P. & Harland, R. M. Wnt signaling in Xenopus embryos inhibits Bmp4 expression and activates neural development. Gene Dev 13, 3149–3159, doi: 10.1101/gad.13.23.3149 (1999).
Rharass, T. et al. Ca2+-mediated Mitochondrial Reactive Oxygen Species Metabolism Augments Wnt/beta-Catenin Pathway Activation to Facilitate Cell Differentiation. Journal of Biological Chemistry 289, 27937–27951, doi: 10.1074/jbc.M114.573519 (2014).
Inestrosa, N. C. & Varela-Nallar, L. Wnt signalling in neuronal differentiation and development. Cell and Tissue Research 359, 215–223, doi: 10.1007/s00441-014-1996-4 (2015).
Wiggan, O. & Hamel, P. A. Pax3 regulates morphogenetic cell behavior in vitro coincident with activation of a PCP/non-canonical Wnt-signaling cascade. Journal of Cell Science 115, 531–541 (2002).
Elizalde, C. et al. Distinct Roles for Wnt-4 and Wnt-11 During Retinoic Acid-Induced Neuronal Differentiation. Stem Cells 29, 141–153, doi: 10.1002/stem.562 (2011).
Zhang, L. L. et al. MiR-499 induces cardiac differentiation of rat mesenchymal stem cells through wnt/beta-catenin signaling pathway. Biochem Bioph Res Co 420, 875–881, doi: 10.1016/j.bbrc.2012.03.092 (2012).
Zhang, J. et al. Effects of miR-335-5p in Modulating Osteogenic Differentiation by Specifically Downregulating Wnt Antagonist DKK1. Journal of Bone and Mineral Research 26, 1953–1963, doi: 10.1002/jbmr.377 (2011).
Wang, Y. et al. miR-375 regulates rat alveolar epithelial cell trans-differentiation by inhibiting Wnt/beta-catenin pathway. Nucleic Acids Research 41, 3833–3844, doi: 10.1093/nar/gks1460 (2013).
Wang, T. & Xu, Z. Y. miR-27 promotes osteoblast differentiation by modulating Wnt signaling. Biochem Bioph Res Co 402, 186–189, doi: 10.1016/j.bbrc.2010.08.031 (2010).
Liu, W. et al. TCF3, a novel positive regulator of osteogenesis, plays a crucial role in miR-17 modulating the diverse effect of canonical Wnt signaling in different microenvironments. Cell Death & Disease 4, doi: 10.1038/cddis.2013.65 (2013).
Kapinas, K., Kessler, C. B. & Delany, A. M. miR-29 Suppression of Osteonectin in Osteoblasts: Regulation During Differentiation and by Canonical Wnt Signaling. Journal of Cellular Biochemistry 108, 216–224, doi: 10.1002/jcb.22243 (2009).
Hu, W. H. et al. miR-142-3p promotes osteoblast differentiation by modulating Wnt signaling. Molecular Medicine Reports 7, 689–693, doi: 10.3892/mmr.2012.1207 (2013).
Zhang, W.-B., Zhong, W.-J. & Wang, L. A signal-amplification circuit between miR-218 and Wnt/β-catenin signal promotes human adipose tissue-derived stem cells osteogenic differentiation. Bone 58, 59–66 (2014).
Egea, V. et al. Tissue inhibitor of metalloproteinase-1 (TIMP-1) regulates mesenchymal stem cells through let-7f microRNA and Wnt/beta-catenin signaling. P Natl Acad Sci USA 109, E309–E316, doi: 10.1073/pnas.1115083109 (2012).
Hassan, M. Q. et al. miR-218 Directs a Wnt Signaling Circuit to Promote Differentiation of Osteoblasts and Osteomimicry of Metastatic Cancer Cells. Journal of Biological Chemistry 287, 42084–42092, doi: 10.1074/jbc.M112.377515 (2012).
Thiebes, K. P. et al. miR-218 is essential to establish motor neuron fate as a downstream effector of Isl1-Lhx3. Nature Communications 6, 7718, doi: 10.1038/ncomms8718 (2015).
Amin, N. D. et al. Loss of motoneuron-specific microRNA-218 causes systemic neuromuscular failure. Science 350, 1525–1529, doi: 10.1126/science.aad2509 (2015).
Chang, J. et al. Fibroblast Growth Factor Signaling Is Essential for Self-renewal of Dental Epithelial Stem Cells. Journal of Biological Chemistry 288, 28952–28961, doi: 10.1074/jbc.M113.506873 (2013).
Tang, K. et al. Wnt-1 promotes neuronal differentiation and inhibits gliogenesis in P19 cells. Biochem Bioph Res Co 293, 167–173 (2002).
Handorf, A. M. & Li, W. J. Induction of mesenchymal stem cell chondrogenesis through sequential administration of growth factors within specific temporal windows. Journal of cellular physiology 229, 162–171, doi: 10.1002/jcp.24428 (2014).
Zuk, P. A. et al. Multilineage cells from human adipose tissue: Implications for cell-based therapies. Tissue Eng 7, 211–228 (2001).
Li, S. et al. Assessment of nanomaterial cytotoxicity with SOLiD sequencing-based microRNA expression profiling. Biomaterials 32, 9021–9030, doi: 10.1016/j.biomaterials.2011.08.033 (2011).
Li, S. et al. MicroRNAs as participants in cytotoxicity of CdTe quantum dots in NIH/3T3 cells. Biomaterials 32, 3807–3814, doi: 10.1016/j.biomaterials.2011.01.074 (2011).
Sun, B., Yang, F., Hu, F. H., Huang, N. P. & Xiao, Z. D. Comprehensive annotation of microRNA expression profiles. Bmc Genet 14, 120 (2012).
Sun, B., Liu, R., Ye, N. & Xiao, Z. D. Comprehensive Evaluation of microRNA Expression Profiling Reveals the Neural Signaling Specific Cytotoxicity of Superparamagnetic Iron Oxide Nanoparticles (SPIONs) through N-Methyl-D-Aspartate Receptor. Plos One 10, 11, doi: 10.1371/journal.pone.0121671 (2015).
Emami, K. H. et al. A small molecule inhibitor of beta-catenin/CREB-binding protein transcription [corrected]. Proc Natl Acad Sci USA 101, 12682–12687, doi: 10.1073/pnas.0404875101 (2004).
Acknowledgements
This work was supported by the National Basic Research Program of China (No. 2013CB932902), the National Natural Science Foundation of China (No. 61071047 and 31371003), the Natural Science Foundation of Jiangsu Province (No. BK2012122), the Postdoctoral Science Foundation of Jiangsu Province (1501017C), the Fundamental Research Funds for the Central Universities (2242016R20026) and the Specialized Research Fund for the Doctoral Program of Higher Education of China (No. 20130092110030).
Author information
Authors and Affiliations
Contributions
Fei-Hu Hu and Bo Sun conceived and coordinated the study and wrote the paper. Xian-Hui Meng and Peng Xu analyzed the sequencing experiments shown in Fig. 2. Yan-Liang Zhu analyzed the experiments shown in Fig. 4. Gao-Jun Teng and Zhong-Dang Xiao provided technical assistance and contributed to the preparation of this study. All authors reviewed the results and approved the final version of the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Hu, F., Sun, B., Xu, P. et al. MiR-218 Induces Neuronal Differentiation of ASCs in a Temporally Sequential Manner with Fibroblast Growth Factor by Regulation of the Wnt Signaling Pathway. Sci Rep 7, 39427 (2017). https://doi.org/10.1038/srep39427
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep39427
- Springer Nature Limited
This article is cited by
-
Comprehensive analysis of differentially expressed profiles of mRNA, lncRNA, and miRNA of Yili geese ovary at different egg-laying stages
BMC Genomics (2022)
-
Adult stem cell response to doped bioactive borate glass
Journal of Materials Science: Materials in Medicine (2020)
-
Conversion of Neural Stem Cells into Functional Neuron-Like Cells by MicroRNA-218: Differential Expression of Functionality Genes
Neurotoxicity Research (2020)