
Abstract
The centromere drive model explaining rapid evolution of eukaryotic centromeres predicts higher frequency of positive selection acting on centromeric histone H3 (CenH3) in clades with asymmetric meiosis compared to the clades with only symmetric meiosis. However, despite the impression one might get from the literature, this key prediction of the centromere drive model has not only never been confirmed, but it has never been tested, because all the previous studies dealt only with the presence or absence instead of the frequency of positive selection. To provide evidence for or against different frequencies of positively selected CenH3 in asymmetrics and symmetrics, we have inferred the selective pressures acting on CenH3 in seventeen eukaryotic clades, including plants, animals, fungi, ciliates and apicomplexa, using codon-substitution models, and compared the inferred frequencies between asymmetrics and symmetrics in a quantitative manner. We have found that CenH3 has been evolving adaptively much more frequently in clades with asymmetric meiosis compared with clades displaying only symmetric meiosis which confirms the prediction of centromere drive model. Our findings indicate that the evolution of asymmetric meiosis required CenH3 to evolve adaptively more often to counterbalance the negative consequences of centromere drive.
Similar content being viewed by others

Introduction
Centromeric histone H3 (CenH3) is the cornerstone of the kinetochore, which ensures the proper segregation of chromosomes during cell division by mediating chromosomal attachment to spindle microtubules. This conserved function implies that the kinetochore should be evolving under strong purifying selection (i.e., it should be maintained as is) over evolutionary time, but multiple reports have suggested that CenH3 has been evolving adaptively (i.e., changing under positive selection) in various lineages of plants and animals1,2,3,4,5,6,7,8. This surprising paradox has been attributed to the recurrent evolutionary conflict between CenH3 and centromeric repeats over centromere control9,10. This conflict is central to the centromere drive theory, which explains the rapid evolution of eukaryotic centromeres and karyotypes and the establishment of reproductive barriers9,10,11,12,13. Under the model of centromere drive, centromeric repeats selfishly exploit the asymmetry of female meiosis (in which only one meiotic product survives, whereas male meiosis is usually symmetric with four surviving products) to secure preferential transmission to the egg at the expense of their homologous counterpart9,10. Expansions (or contractions) of centromeric repeats may lead to increased (or decreased) recruitment of CenH3 and thus to a larger (or smaller) kinetochore, which attracts more (or fewer) microtubules, resulting in the capture of such a kinetochore by the meiotic egg pole when it emanates more (or fewer) microtubules9,10. This driving centromere then rapidly spreads through a population, along with potential negative effects and hitchhiking mutations9,10. Therefore, CenH3 mutations that balance the binding capacity between homologous centromeres are positively selected because they suppress centromere drive along with the associated negative effects9,10.
The prediction that follows is that adaptively evolving CenH3 should be more frequent in clades with asymmetric meiosis than in clades with only symmetric meiosis, where there is no opportunity for conflict because all four meiotic products survive9,14. Although the prediction of differential CenH3 evolution between asymmetric and symmetric clades is a central pillar of the centromere drive theory, it has actually never been tested. Previous studies had suggested that CenH3 is indeed evolving adaptively only in lineages with asymmetric meiosis and under purifying selection in lineages with symmetric meiosis. However, data on CenH3 evolution in lineages with symmetric meiosis are scarce; so far only two clades of yeasts, two species of Plasmodium and two species of Ostreococcus have been analyzed15,16,17. Moreover, since there are also studies that detected a lack of positive selection in asymmetric clades15,18, the lack of detection of positive selection so far reported from species with symmetric meiosis is not sufficient to draw a solid conclusion on the differential CenH3 evolution in symmetrics and asymmetrics.
Taken together, the centromere drive model predicts higher frequency of positive selection episodes in the evolution of CenH3 in eukaryotic clades with asymmetric meiosis (from now on referred to as asymmetrics) than in clades with only symmetric meiosis (from now on referred to as symmetrics). Since not just centromere drive, but other factors may affect the incidence of positive selection events and thus mask the effects of centromere drive, it is necessary to analyze a large enough and representative dataset to detect statistically significant difference in positive selection frequency between the asymmetrics and symmetrics. To achieve such representativeness, we analyzed 191 CenH3 sequences (75% of which is analyzed here for the first time) sequences from a total of seventeen eukaryotic clades of plants, animals, fungi, ciliates and apicomplexa differing in meiotic symmetry (8 asymmetrics and 9 symmetrics; Table 1) by a unified methodical approach.
Methods
We obtained all 191 CenH3 sequences from GenBank and Joint Genome Institute databases19,20,21 using BLAST searches. The sources and accession numbers for all sequences are supplied in Supplementary File S1.
Because the quality of the alignment is absolutely crucial for selection analyses, we used BAli-Phy software, which accounts for alignment uncertainties and avoids problems with biasing alignments towards guide trees because alignments and phylogenetic trees are estimated simultaneously22,23. The codon alignments and phylogenetic trees were jointly inferred in BAli-Phy v2.3.5 using M0 substitution model and RS07 model for indels. Because ciliate nuclear code is not implemented in BAli-Phy v2.3.5, Tetrahymena sequences were aligned at amino acid level using LG substitution model and RS07 model for indels and then backtranslated to nucleotide sequences. For each of the seventeen clades, we ran ten independent chains until they converged and then pooled the results. We masked each codon with a reliability score below 80% as “NNN” prior to selection analyses, and we used maximum a posteriori tree for all selection analyses, except primates, where a known species tree was used. All BAli-Phy alignments of CenH3 before and after masking the unreliable residues are supplied in Supplementary File S2. When only a partial sequence of CenH3 was available, we treated the missing part of the gene as missing data, and the gaps were replaced with “?”. The alignments and phylogenetic trees that we used for selection analyses are supplied in Supplementary File S3.
Once we had alignments and phylogenetic trees of the CenH3 sequences for each of the seventeen clades (Supplementary Files S2 and S3), we employed codon substitution models to infer the selective pressures acting on a protein from the non-synonymous/synonymous substitution rate ratio (dN/dS = ω). Non-synonymous substitutions in a codon lead to amino acid changes, while synonymous substitutions do not. If there is no selective pressure (neutral evolution), non-synonymous and synonymous substitutions are expected to occur at the same rate, with ω = 1. Purifying selection, which keeps the protein as it is, is indicated by ω < 1, and positive selection favoring substitutions that change the amino acids in a protein is indicated by ω > 1. If purifying selection is relaxed, ω tends to be elevated towards 1. Likewise, if positive selection is relaxed, ω tends to decrease towards 1.
We inferred three positive selection measures for each of the 16 analyzed clades: (i) the proportion of positively selected branches in the tree, (ii) the proportion of positively selected codon in the alignments and (iii) the overall ω ratio. To determine the frequency of positively selected branches, we ran the data through the branch-site random effects likelihood (BS-REL) model of codon substitution24. BS-REL allows ω to vary across both codons and branches and infers selective regimes independently for each branch of a given phylogeny, pooling information across all codons24,25. To assess the frequency of positively selected codons, we analyzed CenH3 from each of the seventeen clades using the mixed effects model of evolution (MEME) model of codon substitution that is capable of identifying instances of positive selection at the level of individual codons25. Both BS-REL and MEME analyses were performed using BS-REL and MEME modules as implemented on the DataMonkey web server26. Finally, we examined the overall ω ratio to evaluate CenH3 evolution across all codons and branches for each clade using a one-ratio (M0) model with codon frequencies option set to F3 × 4 in the codeml module of PAML4.7 27.
To assess whether asymmetrics and symmetrics significantly differ in the frequency of positively selected branches and codons and the overall ω ratio, we employed Mann-Whitney U test. However, it is possible that the phylogenetic relationships between analyzed clades may violate statistical independence of analyzed values required for Mann-Whitney U test. Therefore, we have also assessed the differences between asymmetrics and symmetrics using phylogenetically corrected statistical analyses. For that purpose, we have constructed a dated phylogenetic tree of all seventeen clades using divergence times from TIMETREE (Fig. 1;28,29). Statistical significances of phylogenetically corrected differences between asymmetrics and symmetrics were inferred using phylogenetic generalized linear models (pgls) as implemented in caper R-package (pgls function30). Pgls analyses were performed in R 3.2.3 31. The R-script used for pgls analyses is supplied in Supplementary File S4.

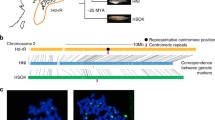
Phylogenetic tree of analyzed clades.
A dated phylogenetic tree of all seventeen analyzed clades. The dating is based on divergence times obtained from TIMETREE. Scale axes shows time in millions of years. Clades having asymmetric meiosis are labeled with (A), while clades with symmetric meiosis are labeled with (S).
Results and Discussion
To determine the frequency of positively selected branches, we have run the BS-REL analysis for each of the seventeen clades. After correction for multiple testing, BS-REL detected positively selected branches in five clades (clades with pcorr ≤ 0.05), all of them from the asymmetrics, while no positively selected branches were detected in the symmetrics (Table 1). When compared, the frequency of positively selected branches in the symmetrics was significantly lower than in the asymmetrics (pMW = 0.027, ppgls = 0.012), which is in accordance with the prediction of the centromere drive model. However, such a result does not rule out positive selection from symmetrics because the corrections for multiple testing may be too conservative and may lead to false-negative results. Therefore, we have also used uncorrected p-values and compared the proportions of positively selected branches between asymmetrics and symmetrics (Table 1). Again, symmetrics had a significantly lower proportion of branches under positive selection than asymmetrics (pMW = 0.001, ppgls = 0.0005; Fig. 2A).

Distinct CenH3 evolution patterns in the asymmetrics and symmetrics.
Box plots show differences between asymmetrics and symmetrics in (A) the proportion of positively selected branches after the correction for multiple testing; (B) the proportion of positively selected codons and (C) the average ω. N – the number of clades analyzed in each group (see Table 1 for details), pMW – significance from Mann-Whitney U Test, ppgls – significance from pgls analysis.
To assess the frequency of positively selected codons (from all codons in the alignment) in asymmetrics and symmetrics, we analyzed CenH3 from each of the seventeen clades using MEME (see Methods for details). The symmetrics had a significantly lower proportion of positively selected codons than asymmetrics (pMW = 0.002, ppgls = 0.002; Fig. 2B) as predicted by the centromere drive model. Moreover, the asymmetrics and symmetrics differed also in the distribution of positively selected codons across functional domains of the CenH3 protein (N-terminus and histone fold domain - HFD).
Since the HFD is required for centromere targeting and directly interacts with centromeric DNA28, it is thought to play a prominent role in the centromere drive3,13 and asymmetrics might thus be expected to experience a higher frequency of positive selection events in the HFD than symmetrics. In agreement with this expectation, in symmetrics, only two (Bryophyta and Trichoderma) of nine clades (22%) showed positively selected codon in the HFD, while in asymmetrics, five of eight (63%) displayed positively selected codons in this domain (Table 1).
Finally, to evaluate whether and how the asymmetrics and symmetrics differ in CenH3 evolution on a global scale, i.e., across all codons and branches, we examined the overall ω ratio for each clade. In each clade, CenH3 showed an average ω < 1 (Table 1), suggesting that most codons have been evolving under purifying selection for most of their evolutionary history. This observation may be expected because positive selection usually acts on only a few codons for a limited amount of time30. However, we detected a striking difference in the average ω between asymmetrics and symmetrics (Fig. 2C). The symmetrics showed significantly lower ω than the asymmetrics (pMW = 0.006; ppgls = 0.0007; Fig. 2C). Such a low ω implies that CenH3 has been evolving under much stronger purifying selection over evolutionary time in the symmetrics than in the asymmetrics and/or that the asymmetrics experienced more episodes of positive selection which is in accordance with the BS-REL and MEME analyses (see above).
Using three different models of codon substitution, we have shown that CenH3 has been evolving under different selective pressures in clades with opportunities for centromere drive (asymmetrics) compared with clades without such opportunities (symmetrics). In accordance with the predictions of the centromere drive model and previous reports based on smaller datasets15,16,17, episodes of positive selection acting on CenH3 appear to be much less frequent in symmetrics (Fig. 2A–C).
Interestingly, we have detected adaptively evolving CenH3 in the ciliate genus Tetrahymena from asymmetrics (Table 1) where the previous study reported the absence of positive selection18. In contrast to Elde et al. who suggested unsuppressed centromere drive in Tetrahymena18, our results indicate that centromere drive might have been suppressed in these ciliates. On the other hand, in CenH3 of Drosophila and Primates, we have detected positive selection only in N-terminus (Table 1), while previous studies reported positively selected codons in both N-terminus and HFD1,5,7. This discrepancies may be attributed to the fact that, for the detection of positively selected codons, we have used branch-site models of codon substitution instead of site models employed in the previous studies. Although the branch-site models and site models may perform differently because they have different assumptions32, it is important that we have used the same branch-site models consistently in all our analyses, an approach allowing us to compare CenH3 evolution in symmetrics and asymmetrics.
A recent study suggested that the evolution of centromeric DNA repeats and CenH3 might be driven by selection of centromere-linked genes rather than by centromere drive in inbred lines of maize33. However, selection on centromere-linked genes and centromere drive are not mutually exclusive evolutionary mechanisms. If the selection on centromere-linked genes was the only mechanism responsible for the evolution of centromeres and kinetochore proteins, there should be no competition for surviving meiotic products and symmetrics and asymmetrics should not differ in the frequency of positive selection acting on CenH3. The observation of different frequency of positively selected CenH3 between symmetrics and asymmetrics (Table 1; Fig. 1) suggests that centromere drive has indeed been operating in asymmetrics. Furthermore, in Saccharomyces (symmetrics), centromeric DNA has been rapidly evolving due to very high mutation rate34,35, yet their CenH3 does not show almost any signs of positive selection16,17 (Table 1). Since there is no opportunity for centromere drive in Saccharomyces due their symmetric meiosis, these findings suggest that meiotic asymmetry may indeed be the main factor responsible for the adaptive evolution of CenH3.
In the light of the centromere drive model and the results of our and previous studies, it is likely that before asymmetric meiosis appeared on the evolutionary scene, CenH3 had been evolving under strong purifying selection with occasional episodes of positive selection. After meiotic asymmetry appeared, positive selection occurred at a higher frequency because CenH3 was forced to counteract the negative consequences of centromere drive. Interestingly, some asymmetrics have evolved holokinetic chromosomes36 that have been hypothesized to suppress centromere drive10,17. The absence of positively selected CenH3 recently found in holokinetic plant genus Luzula37 and the fact that holokinetic chromosomes have so for been reported only in asymmetrics support this hypothesis.
Additional Information
How to cite this article: Zedek, F. and Bureš, P. CenH3 evolution reflects meiotic symmetry as predicted by the centromere drive model. Sci. Rep. 6, 33308; doi: 10.1038/srep33308 (2016).
References
Malik, H. S. & Henikoff, S. Adaptive evolution of Cid, a centromere-specific histone in Drosophila. Genetics 157, 1293–1298 (2001).
Talbert, P. B., Masuelli, R., Tyagi, A. P., Comai, L. & Henikoff, S. Centromeric localization and adaptive evolution of an Arabidopsis histone H3 variant. Plant Cell 14, 1053–1066 (2002).
Cooper, J. L. & Henikoff, S. Adaptive evolution of the histone fold domain in centromeric histones. Mol. Biol. Evol. 21, 1712–1718 (2004).
Hirsch, C. D., Wu, Y., Yan, H. & Jiang, J. Lineage-Specific Adaptive Evolution of the Centromeric Protein CENH3 in Diploid and Allotetraploid Oryza Species. Mol. Biol. Evol. 26, 2877–2885 (2009).
Schueler, M. G., Swanson, W., Thomas, P. J., Green, E. D. & Progra, N. C. S. Adaptive Evolution of Foundation Kinetochore Proteins in Primates. Mol. Biol. Evol. 27, 1585–1597 (2010).
Zedek, F. & Bureš, P. Evidence for Centromere Drive in the Holocentric Chromosomes of Caenorhabditis. Plos One 7, e30496 (2012).
Beck, E. A. & Llopart, A. Widespread Positive Selection Drives Differentiation of Centromeric Proteins in the Drosophila melanogaster subgroup. Sci Rep 5, 17197 (2015).
Neumann, P. et al. Centromeres Off the Hook: Massive Changes in Centromere Size and Structure Following Duplication of CenH3 Gene in Fabeae Species. Mol Biol Evol, doi: 10.1093/molbev/msv070 (2015).
Henikoff, S., Ahmad, K. & Malik, H. S. The centromere paradox: Stable inheritance with rapidly evolving DNA. Science 293, 1098–1102 (2001).
Malik, H. S. & Henikoff, S. Major Evolutionary Transitions in Centromere Complexity. Cell 138, 1067–1082 (2009).
Pardo-Manuel de Villena, F. & Sapienza, C. Female meiosis drives karyotypic evolution in mammals. Genetics 159, 1179–1189 (2001).
Yoshida, K. & Kitano, J. The contribution of female meiotic drive to the evolution of neo-sex chromosomes. Evolution 66, 3198–3208 (2012).
Molina, W. F., Martinez, P. A., Bertollo, L. A. & Bidau, C. J. Evidence for meiotic drive as an explanation for karyotype changes in fishes. Mar. Genomics 15, 29–34 (2014).
Roach, K. C., Ross, B. D. & Malik, H. S. Adaptive Evolution of Centromeric Proteins. ELS, doi: 10.1002/9780470015902.a0022868.
Talbert, P. B., Bryson, T. D. & Henikoff, S. Adaptive evolution of centromere proteins in plants and animals. J. Biol. 3, 18 (2004).
Baker, R. E. & Rogers, K. Phylogenetic Analysis of Fungal Centromere H3 Proteins. Genetics 174, 1481–1492 (2006).
Talbert, P. B., Bayes, J. J. & Henikoff, S. Evolution of centromeres and kinetochores: A two-part fugue in The Kinetochore (eds De Wulf, P. & Earnshaw, W. C. ) 193–230 (Springer: Berlin,, 2008).
Elde, N. C., Roach, K. C., Yao, M. C. & Malik, H. S. Absence of positive selection on centromeric histones in Tetrahymena suggests unsuppressed centromere: drive in lineages lacking male meiosis. J. Mol. Evol. 72, 510–520 (2011).
Benson, D. A., Karsch-Mizrachi, I., Lipman, D. J., Ostell, J. & Wheeler, D. L. GenBank. Nucleic Acids Research 33, (Database issue), D34–D38 (2005).
Grigoriev, I. V. et al. The Genome Portal of the Department of Energy Joint Genome Institute. Nucleic Acids Res. 40, (Database issue), D26–D32 (2012).
Nordberg, H. et al. The genome portal of the Department of Energy Joint Genome Institute: 2014 updates. Nucleic Acids Res. 42, D26–D31 (2014).
Redelings, B. D. & Suchard, M. A. Joint Bayesian Estimation of Alignment and Phylogeny. Syst. Biol. 54, 401–418 (2005).
Redelings, B. D. Erasing Errors Due to Alignment Ambiguity When Estimating Positive Selection. Mol. Biol. Evol. 31, 1979–1993 (2014).
Kosakovsky Pond, S. L. et al. A random effects branch-site model for detecting episodic diversifying selection. Mol. Biol. Evol. 28, 3033–3043 (2011).
Murrell, B. et al. Detecting individual sites subject to episodic diversifying selection. PLoS Genetics 8, e1002764 (2012).
Delport, W., Poon, A. F., Frost, S. D. & Kosakovsky Pond, S. L. Datamonkey 2010: a suite of phylogenetic analysis tools for evolutionary biology. Bioinformatics 26, 2455–2457 (2010).
Yang, Z. PAML 4: phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 24, 1586–1591 (2007).
Hedges, S. B., Dudley, J. & Kumar, S. TimeTree: a public knowledge-base of divergence times among organisms. Bioinformatics 22, 2971–2972 (2006).
Hedges, S. B., Marin, J., Suleski, M., Paymer, M. & Kumar, S. Tree of Life Reveals Clock-Like Speciation and Diversification. Mol Biol Evol 32, 835–845 (2015).
Orme, D. et al. Caper: Comparative Analyses of Phylogenetics and Evolution in R. URL: http://cran.r-project.org/web/packages/caper/caper.pdf (2012) (Date of access: 01/03/2016).
R Core Team. R: a language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. URL: http://www.R-project.org/ (2013).
Yang, Z. & dos Reis, M. Statistical properties of the branch-site test of positive selection. Mol Biol Evol 28, 1217–1228 (2011).
Schneider, K. L., Xie, Z., Wolfgruber, T. K. & Presting, G. G. Inbreeding drives maize centromere evolution. PNAS 113, E987–E996 (2016).
Bensasson, D., Zarowiecki, M., Burt, A. & Koufopanou, V. Rapid evolution of yeast centromeres in the absence of drive. Genetics 178, 2161–2167 (2008).
Bensasson, D. Evidence for a high mutation rate at rapidly evolving yeast centromeres. BMC Evol Biol 11, 211 (2011).
Bureš, P. & Zedek, F. Holokinetic drive: Centromere drive in chromosomes without centromeres. Evolution 68, 2412–2420 (2014)
Zedek, F. & Bureš, P. Absence of positive selection on CenH3 in Luzula suggests that holokinetic chromosomes may suppress centromere drive. Annals of Botany, doi: 10.1093/aob/mcw186 (in press).
Acknowledgements
We greatly appreciate the computational resources that were provided by MetaCentrum (the program LM2010005) and CERIT-SC (the program Centre CERIT Scientific Cloud, part of the Operational Program Research and Development for Innovations, Reg. no. CZ.1.05/3.2.00/08.0144) because our analyses requiring almost 1,500 CPU days would otherwise be impossible to perform. This work was funded by the Czech Science Foundation, grant no. 13–29362S.
Author information
Authors and Affiliations
Contributions
F.Z. and P.B. conceived the study, F.Z. analyzed the data, F.Z. and P.B. wrote the manuscript. Both authors reviewed the manuscript prior to the submission.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Zedek, F., Bureš, P. CenH3 evolution reflects meiotic symmetry as predicted by the centromere drive model. Sci Rep 6, 33308 (2016). https://doi.org/10.1038/srep33308
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep33308
- Springer Nature Limited