
Abstract
Cooperative behavior, a natural, pervasive and yet puzzling phenomenon, can be significantly enhanced by networks. Many studies have shown how global network characteristics affect cooperation; however, it is difficult to understand how this occurs based on global factors alone, low-level network building blocks, or motifs are necessary. In this work, we systematically alter the structure of scale-free and clique networks and show, through a stochastic evolutionary game theory model, that cooperation on cliques increases linearly with community motif count. We further show that, for reactive stochastic strategies, network modularity improves cooperation in the anti-coordination Snowdrift game and the Prisoner’s Dilemma game but not in the Stag Hunt coordination game. We also confirm the negative effect of the scale-free graph on cooperation when effective payoffs are used. On the flip side, clique graphs are highly cooperative across social environments. Adding cycles to the acyclic scale-free graph increases cooperation when multiple games are considered; however, cycles have the opposite effect on how forgiving agents are when playing the Prisoner’s Dilemma game.
Similar content being viewed by others


Introduction
The puzzling existence of cooperation in the face of the seemingly selfish process of natural selection has occupied researchers across many disciplines for over 30 years1,2. This interest extends beyond the most common biological and social contexts2,3 to a wide array of literature including, the theory of the firm in business where coordination of functions and competition with external entities are key considerations4, coordination during preparation for natural disasters in emergency management literature5, how cooperation influences morality emergence6, in the psychology literature and the physical mechanisms that give rise to human cooperation provided by neuroscience7,8, to name a few. A common way to model cooperation is by evolving an interacting population of social dilemma game players; a canonical game is the Prisoner’s Dilemma9,10.
This line of research has revealed a host of factors that contribute to the survival of cooperation11 including, remarkably, the interaction structure itself (see12 for a review). Evolving organisms often reproduce locally which in turn causes feedback between locally distributed traits and genetic relatedness between traits, this leads to interaction assortment and population viscosity13,14 that sustains cooperative behavior; however, what properties of structure drive what characteristics of cooperation and how? One way to tackle this problem that has proven useful is to consider evolving players of social dilemma games within a network, where the games provide a framework for understanding the simplest local interactions and the network frames community-level and global interaction12. This general approach has shown that relatively sparse connectivity15 or less frequent interaction16 helps encourage cooperation. More modular structures also encourages cooperation relative to networks with less separable communities17. It has also been commonly shown that heterogeneous networks, such as scale-free graphs with a power-law degree distribution dependence18, encourage cooperation18,19,20. Even so, it turns out that the advantage of cooperators disappears in scale-free networks when payoffs are averaged over neighbors rather than summed, because hubs are easily invaded by defectors even if cooperators favor hub locations21,22. Cycle length within a network is also a key characteristic and the strongest evolutionary dynamics tend to occur within cycles23. Short and long cycle effects compete with one another; short cycles pull cooperation lower while long cycles drive cooperation higher in the Prisoner’s Dilemma game24 and Ref. 25 shows that short cycles similarly limit cooperation.
In the present investigation into how the properties of structure drive cooperation we consider cooperation over four two-person, symmetric, stylized games common in the literature (i.e., Stag Hunt (SH), Snowdrift (SD), Harmony Game (HG) and Prisoner’s Dilemma (PD)), rather than compare the effects of one or two games, which is most common. In a social context any individual game may prove relevant for a given situation at a certain point in time and so the representative game is subject to change over time26,27; however, altering network structure to adapt to changing circumstances is costly28. In fact, network structure has been shown to actually inhibit cooperation when costs are high relative to benefits in the Snowdrift game29 and the common assumption of heterogeneous degree distribution encouraging cooperation is negated by participation costs30 and payoff averaging21,22. Therefore, in order to design enduring structures that influence behaviors either towards or away from cooperation we need to consider a wide range of social situations that the population may encounter.
How networks affect cooperation is usually considered as relative to average global structural properties, but this approach cannot give an accurate picture of local network characteristics nor their influence on evolving processes or functions, this is the purview of network motifs31. Motifs are common, repeating, statistically significant, subgraph building blocks, that lie below both the global and community level32,33,34 and just above the lowest node and link level of a network. The notion of network motifs applies to biological, social and information network contexts; where, for example, motifs found in the world wide web are distinct from genetic networks35. Motifs, which can be efficiently detected in real-world networks36,37, are functionally separable modules which interact dynamically within an organism and between an organism and its environment to both provide core functionality and adapt to the environment38. Since the environment is not static, but changes over time, evolutionary goals also vary, therefore, modular structure and network motifs must emerge to improve survival in this dynamic environment39. Certain motifs may be indicators of social status in networks40 and influence network dynamics as well41. Finally, if a range, or spectrum of motifs is considered a network hierarchy may be determined, when the network is directed42.
In the present work we use network motifs to help us understand the consequences of subtle changes in network structure which have very little effect on average global network characteristics but influence cooperation emergence.
Here forward, we introduce the evolutionary model we have developed for this work, highlight the effect of varying structures on cooperation across multiple games and delve into two example structures which show the lowest average cooperation across games (a scale-free graph) and highest average cooperation across games (a clique-type graph), relative to the fully-connected case. Our results show that networks which lack either sufficient cyclic structures or contain densely connected cliques suffer significantly reduced cooperation, particularly in coordination situations, adding cycles helps here but when the social environment becomes more competitive this can be counterproductive. On the flip side, connecting highly cooperative clique communities together improves cooperation linearly with the frequency of local community motif structures, which extend interfaces to neighboring communities and give a cohesive structure for coordination and cooperation locally. Cycles can either help (coordination) or hinder (competition) depending on the social environment that is encouraged by a particular network which spread prevailing community behavioral norms to other communities.
Results
Model
The model we have developed for this work operates on four levels: pair-wise games, strategy space, the learning model and the interaction structure or network. Game play occurs between agents who play two-player repeated games with each of their nearest neighbors and receive payoffs according to the moves of each player, commensurate with the payoff structure of the game, equation (4). We consider four games, including the Prisoner’s Dilemma, or PD (the strongest social dilemma), Stag Hunt (SH), Snow Drift (SD) and the Harmony Game, or HG (the weakest dilemma). During game play, agents, with memory of their opponent’s last move within a repeated game, determine their next move according to their stochastic reactive strategy, which is based on the three-component, Markovian, mixed-strategy framework from43,44, where y is the probability to trust on the first move, p is the probability to reciprocate a cooperative move and q is the probability to forgive defection. Since y acts in a single round within a repeated game and the other strategies repeat throughout y is plagued by noise, indeed it can be problematic to collect enough samples to show the meaningful evolution of y (trust)17. Therefore, we treat y as a discrete rather than a continuous variable in this work; this treatment decreases y noise considerably and improves run convergence. Learning, or strategy updating, is done asynchronously via the Fermi rule, equation (1), from relatively weak to strong selection,  . We also employ a social interpretation (which we refer to as SOC, see equation (2)) of the Fermi rule where β is inversely proportional to the payoff noise across each agent’s nearest neighbors; we describe the model in greater detail in the Methods section.
. We also employ a social interpretation (which we refer to as SOC, see equation (2)) of the Fermi rule where β is inversely proportional to the payoff noise across each agent’s nearest neighbors; we describe the model in greater detail in the Methods section.
In our quest to increase the model’s sensitivity and performance we abandoned the synchronous updating rule, replicator equation and stochastic y methods from prior work17 in favor of asynchronous updating, the Fermi rule and a discrete y variable. These changes also allowed us to verify cooperation improvement (shown in Fig. 1) with modularity, studied by17, under different updating and evolutionary rules. In addition, we tested more modular structures up to a maximum modularity Q32 of nearly 0.9. A graph generator was required to test the modularity range from 0.6 to 0.9 in nearly equal increments; we developed the generator based on45.

Cooperation change with structure.
The figure shows difference in average cooperation  across a set of the four most common two player symmetric games (i.e., Harmony Game, Stag Hunt, Snow Drift and Prisoner’s Dilemma) between a set of networks in modularity order. The point symbol is the corresponding graph index from the networks figure in the Methods section. The network modularity (Q in32) is shown along the horizontal axis; sample graphs are shown above the plot. Graph node color represents community membership, which we determined using the random-walk method described by60. The most significant outliers are the Barabási-Albert (BA) graph47,48 (graph index 27) which experiences the least improvement (negative) in cooperation and the clique graph (graph index 37) which evolves much higher cooperation (both compared to the full graph). Both of these cases are noted in the figure with a blue arrow.
across a set of the four most common two player symmetric games (i.e., Harmony Game, Stag Hunt, Snow Drift and Prisoner’s Dilemma) between a set of networks in modularity order. The point symbol is the corresponding graph index from the networks figure in the Methods section. The network modularity (Q in32) is shown along the horizontal axis; sample graphs are shown above the plot. Graph node color represents community membership, which we determined using the random-walk method described by60. The most significant outliers are the Barabási-Albert (BA) graph47,48 (graph index 27) which experiences the least improvement (negative) in cooperation and the clique graph (graph index 37) which evolves much higher cooperation (both compared to the full graph). Both of these cases are noted in the figure with a blue arrow.
For payoffs, we use the average performance of a node over its neighborhood (see equation (3)) rather than a the neighborhood sum used by46. We chose this approach both to associate results of this study with prior modularity work17 and to increase the generality of our results since the averaging rule produces dynamics invariant to payoff matrix translations22. Averaging payoffs may negate the benefit of the scale-free network21 but sparse cliques still enjoy broad cooperation improvement, which we shall discuss in the next section.
Cooperation and modularity
Broadly considering the results in Fig. 1, the network with the lowest overall change in cooperation relative to the full graph, across all games, is the Barabási-Albert (BA) graph47,48 in Fig. 1 (graph index 27). Cooperation change relative to each game is shown in Fig. 2 where significant drops in cooperation occur along the spectrum from low to high modularity. The most prominent dropouts in Fig. 2 fall into three categories, those with highly connected cliques (index 12, 13, 16, 19, 34, 35 in Fig. 2) whose modules approximate a full graph and so are less cooperative, the BA graph (index 27 in Fig. 2) which has reduced cooperation due to frequent invasion of hubs by defectors21,22 and approximately acyclic graphs (index 20, 26 in Fig. 2). The dropouts are the largest in the BA graph, especially in the Snowdrift and Stag Hunt games, loss of cooperation in the later is significant enough to reduce average cooperation across all games below the full graph (Fig. 1).

Effect of modularity on cooperation.
The figure shows Cooperation p (reciprocity) as relative to Modularity (Q) conditioned by game, one curve for each game. Significant cooperation changes (e.g., dropouts) are indicated by vertical lines with the corresponding network attached, along with the corresponding graph index from the network figure in the Methods section. Index 12, 13, 16, 19, 27, 34 and 35 correspond to local cooperation depressions (dropouts), network 37 from Fig. 1 is also shown. Approximately acyclic graphs are highlighted on the graph (20, 26) for each game; networks are shown for each on the plot.
From the ST-plane results in Fig. 3, forgiveness is affected the least by changes in network structure. The simple 5 × 5 lattice structure (see SI1 for lattice details) improves cooperation significantly across the plane but the Snowdrift game shows the most improvement, but cooperation dominates the majority of the ST plane for the clique graph (rightmost column in Fig. 3). Since payoffs are averaged based on a node’s performance against each of its neighbors (confirming21,22 for reactive stochastic strategies), cooperation in the BA graph (27 in Fig. 3) is reduced across all games, even compared with the fully connected graph (leftmost column in Fig. 3).

ST Plane versus structure.
The figure shows cooperation across the ST-plane for y (top row), p and q (bottom row) for the full graph (left column), the BA graph (index 27), the 5 × 5 square lattice (index 17) and the clique graph (right column, index 37). Coordinates within each ST-plane represent the payoffs of the Harmony Game (HG), Snowdrift (SD), Stag Hunt (SH) and Prisoner’s Dilemma (PD) games; divisions between each game are shown as white dashed lines and the game region is noted in the bottom of the figure. See SI1 for detailed plots comparing each network with the the full graph by game.
Beyond the payoff structure effect reported by21,22, we hypothesize that some of the cooperation erosion in the BA graph is due to its acyclic nature rather than its preferential attachment property or heterogeneous degree distribution per se. The evolutionary dynamics of games on cycles can favor cooperation, especially for more competitive games like the Prisoner’s Dilemma23 and analytical work has shown that a combination of relatively large girth and fewer short cycles should improve cooperation24, so an undefined girth (acyclic) may limit cooperation likewise. To test this, we developed an experiment in the next section that attempts to increase scale-free graph competition through adding cycles from single-edge changes to the network.
Scale-free graph edge experiment
To provide evidence for the effect of cycles on cooperation we constructed a simple illustrative example by systematically adding cycles of different lengths to the BA graph, thus gradually increasing its girth. From Fig. 4(A), when girth rises average cooperation across all games increases proportionally. However, girth fails to tell the complete story. If higher degree hubs are included in the cycle (Fig. 4(A) point d), cooperation is higher than the same length cycle that connects through a lower degree node (Fig. 4(A) point e). Cooperation does not continually improve as cycle length increases, a limit is reached where new cycles must be added to achieve further improvement. If we add another cycle (Fig. 4(A) point h) cooperation improves but if a new cycle is added by splitting a large cycle (Fig. 4(A) point i), thus decreasing girth, cooperation decreases relative to the two-cycle case (point h). These effects are similar in nature from strong to weak selection, including the SOC β rule, equation (2).

Effect of cycles on cooperation.
Panel (A) shows the change in reciprocity p across all games by single-edge additions to the BA graph (Fig. 1 index 27). Each point is represented by a square with length (width) proportional to the girth of the graph, or the length of the shortest cycle. The girth value is shown within each data point square for the  . The graph corresponding to each data point is shown above; the shortest cycle is shown in orange. The vertical axis represents the average reciprocal cooperation (p) level across games (PD, SD, HG, SH) relative to the full graph (Fig. 1 index 1). The four selection strength β curves are shown (0.001, 1, 10 and the SOC adaptive method, equation (2)). Panel (B) conditions the cooperation results by game relative to the full graph, for
. The graph corresponding to each data point is shown above; the shortest cycle is shown in orange. The vertical axis represents the average reciprocal cooperation (p) level across games (PD, SD, HG, SH) relative to the full graph (Fig. 1 index 1). The four selection strength β curves are shown (0.001, 1, 10 and the SOC adaptive method, equation (2)). Panel (B) conditions the cooperation results by game relative to the full graph, for  . The full graph is indicated by the horizontal red line. Point symbols are for each respective point index in panel (A). A summary of the effects in panel (B) is shown in Table 1.
. The full graph is indicated by the horizontal red line. Point symbols are for each respective point index in panel (A). A summary of the effects in panel (B) is shown in Table 1.
Since our model separates behaviors into three factors, trust (y), reciprocity (p) and forgiveness (q), we can see some of the inner workings of the effect cycles have on cooperation, shown in Fig. 4(B). First we see that in all respects BA-graph cooperation in the PD game is higher than the full graph; however, the coordination games (SH and HG) lose ground and are less cooperative and the anti-coordination game, SD, decreases in cooperation likewise but is able to remain slightly more cooperative than the full graph due to its modularity. The dearth of cycles does also tend to make cooperation more difficult in coordination situations, though the effect is modest. Moreover, there is a marked difference between games; the positive effect of girth on y and p strengthens in the SH and SD games but then reverses in the PD game where y and q reduce with girth, see Table 1 for a summary of these effects. Long cycles help trust and reciprocity evolve in the anti-coordination game (Snowdrift) but when the game shifts to a more competitive situation (PD) longer cycles actually reduce trust and forgiveness; reciprocity is unaffected. The difference lies in the evolutionary dynamics where long cycles provide more opportunities to dampen defection through forgiveness than short cycles. Further, when a cycle is lengthened a higher proportion of nodes in the network are within the cycle, which are known to favor cooperators over defectors due to the supporting effect of cooperators just behind the boundary between communities; defectors fail to support in kind23. In the more competitive PD game the ability of forgivers in the cycle to support boundary nodes that experience defection is compromised, this causes the strategy boundary to move in favor of defectors as cycles lengthen.
Cycles do seem to play a key role in cooperation emergence; however, acheiving highly cooperative structures through adding cycles is difficult, in-fact, doing so by splitting cycles is an ineffective way to increase cooperation. Even though cooperation increases by adding cycles to the BA graph we were not able to meet the cooperation levels of the full graph in our eight edge addition experiment though the social (SOC) rule came close to achieving parity (Fig. 4 point h). To create the most cooperative structures, we must start with more modular structures that are initially much more cooperative than the full graph, demonstrated by17 for the Prisoner’s Dilemma game.
Clique graph edge experiment
To synthesize a more cooperative environment we start with a network that has the highest average payoff across games (a clique graph in Fig. 1, index 37) and add edges that join communities such that short cycles (e.g., triangular motifs) are not affected. From Fig. 5, average cooperation increases as edges are added. We continue to add edges to create another complete cycle between cliques and cooperation increases likewise. Because the motif changes are controlled only one cyclic motif is affected by these edge additions, namely the community motif in Fig. 6, all other changing motifs are acyclic. Since we expect acyclic structures to be less cooperative (Fig. 2) and adding edges between communities decreases modularity (9% total decrease for index 11 versus index 1 in Fig. 5(A)) it follows that the community motif is largely responsible for the cooperation improvement in this clique experiment in spite of the opposing effect from lower modularity; in this example cooperation improved by 1% relative to the full graph for each edge added. We found this effect to be robust to selection strength as well as the social (SOC) selection rule and confirmed the magnitude of the effect with the replicator equation utilized in17.

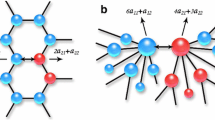
Motif changes between cliques.
The figure shows each of the graph motifs35 of size 3 and 4 that are found within the graphs. In our clique edge-addition experiment we added edges in a way that did not increase short cycles and this is demonstrated by unchanging quantities of triangle, square and dual triangle motifs. The largest changing motif with a cycle is the community motif. Long interface motifs increase linearly throughout the experiment (interface B motif) and at the highest rate that indicates an increase in long cycles.
The motif results are separated by game and y, p, q component in Fig.5(B). There is a considerable difference in the SD (strong T) versus SH (weak T) games; forgiveness does not evolve in the former (strong) case versus the later (weak) and HG remains unaffected by the community motif increase. In contrast with the girth study (Fig. 4), p increases with the community motif in all games except HG and no reversal occurs in any game, compared to the PD reversal (Fig. 4) when cycles are added to the BA graph. Since the clique graph provides a highly cooperative environment defection is largely contained, demonstrated by the unchanging forgiveness (q) as interfaces with other communities are added.
Discussion
To summarize, there are four main structural factors that affect cooperation in these settings, some positive and others negative, each act at either the local or global level of the network (see Table 1). These structural factors include connection degree15, modularity17 and as we have shown in this work, cycles23,24,25 and community motifs.
There are differences, however, in how networks affect cooperation between pure strategy evolution and stochastic strategy evolution. For pure strategies in the PD game25, showed that triangles limit cooperation under high noise conditions, but we show that even a single triangle can increase average cooperation across games when stochastic strategies are used across noise levels. This results in a different ideally-cooperative structure for stochastic reactive strategies across games, many long cycles are desired. This contrasts with pure strategies in the PD, where acyclic networks are desired under high noise conditions and randomly connected triangular structures are desired under low noise conditions25.
Further, we do confirm the result from25, that even a single triangle decreases cooperation in the PD game, but limit this effect to the forgiveness q part of cooperation (see Fig. 4(B), PD panel, point b).
Modularity may be essential for improving cooperation globally17 but the modules themselves must be connected sparsely (see Fig. 2), as15 suggests with the b/c > k rule, to sustain cooperation. Indeed some of the most modular structures we tested, including graph index 42 in Fig. 1, were significantly less cooperative than the structures in our clique experiment (Fig. 5) due to dense connections within their modules and cooperation dropouts (i.e., low cooperation though the network is modular) occurred commonly in networks with highly connected modules (see graphs 12, 13, 16, 19, 34, 35 in Fig. 2). Further improvement in already cooperative clique networks requires community-to-community connections but in a controlled way without increasing short cycles that reduce support for cooperators at the boundary of communities. This community motif both joins together agents in cohesive groups and extends an interface to other groups as well.
The social environment is far different between the clique network model and the Barabási-Albert47,48 (BA) scale-free model. BA-graph cooperation is high relative to the fully connected graph in competitive situations, modeled by the Prisoner’s Dilemma; however, in the Stag Hunt and Snowdrift games cooperation is comparatively low but can hang on and even improve modestly as communities are connected better but this same connectivity becomes a liability under competition (Fig. 4) by allowing defection to invade through the same pathways. Taken as a whole, though, cooperation improvement is lower in scale-free graphs than well mixed or fully connected graphs, but we do find that cooperation is higher on lattices than scale free, contrary to human cooperation experiments50 that show both lattice and scale free networks behave the same, but aligns with the theoretical literature that shows improvements in the lattice over well mixed populations17,48,51.
In the clique case (Fig. 5), cooperation is robust therefore adding links between communities gives additional opportunities for reciprocal action. These same paths allow defectors to invade scale-free network hubs and this cycle of invasion erodes forgiveness and trust. Acyclic structures are an asset in competitive situations, represented by the Prisoner’s Dilemma game, because they limit the spread of defection to otherwise cooperative communities; however, when cycles are absent and coordination is needed, where agents attempt to predict52 future moves of their partners, acyclic communities lack sufficient support at external boundaries that cycles can provide23 and so are unable to efficiently coordinate which reduces fitness of the global population.
We have also developed a socially adaptive rule SOC in which the noise is proportional to the local environment noise in payoffs and show that these results are similar to that of strong selection (β > 10) and this constrains the usefulness of this rule to relatively strong selection found in microbial cooperation53, rather than human cooperation in which the moves of players have a more significant impact than payoffs54.
In this work, we have studied cooperation emergence in both the BA and clique structures through subtle single-edge network changes. We are unaware of any comparable study that delves into single-edge effects, nor the effect of network motifs on cooperation emergence, which we show to be linear in our clique example (Fig. 5). We also confirm, for reactive strategies, that scale-free network cooperation is significantly reduced, as reported by21,22 and confirm the robustness of these results from weak to strong selection, including a socially adaptive rule (SOC).
Research that extends the present work includes investigating the size and connectivity of graph clique modules and extending the strategy space to a four-part framework that includes the Pavlov strategy, for example extending55 and the present work.
Methods
We evolve our population of social dilemma game players in a network environment (see Fig. 7) where nodes represent players and links the interactions where games are played. A prior modularity study17 used an updating rule from18; however, this rule gives up the ability to adjust selection strength. In this work we apply the more conventional Fermi rule, equation (1), in order to take advantage selection adjustment. Additionally,17 performed synchronous updating whereas in the present work we employ asynchronous updating. Finally, we modified the y, p, q stochastic strategy framework, from43,44, slightly by making y a discrete rather than continuous variable. These changes were largely motivated by the need to decrease simulation convergence time and we found that these rules, especially asynchronous updating, decreased convergence time considerably. These changes also allowed us to confirm the modularity effect on cooperation17, with asynchronous updating and the Fermi rule, from weak to strong selection.

Networks.
All graphs, except for the edge addition BA and clique experiments in are shown in this figure. The modularity value (Q32) is shown above each panel along with the graph index in parentheses. Individual communities or modules are indicated by color where nodes with the same color belong to the same community or module. Community detection is performed by a random walk method described by60. The number of nodes is shown on each graph in the figure.

In the Fermi equation (1),  is the probability that node o will adopt the strategy of neighbor n,
is the probability that node o will adopt the strategy of neighbor n,  and
and  are the payoffs of o and neighbor n respectively, β is the evolutionary selection strength and Δ is the difference between the maximum and minimum payoff achievable in a given game. The later Δ parameter allows us to vary payoffs across a wide range of games and yet evolve the population in a common way54. As
are the payoffs of o and neighbor n respectively, β is the evolutionary selection strength and Δ is the difference between the maximum and minimum payoff achievable in a given game. The later Δ parameter allows us to vary payoffs across a wide range of games and yet evolve the population in a common way54. As  a small payoff advantage results in a high probability of switching strategies conversely when
a small payoff advantage results in a high probability of switching strategies conversely when  the probability of switching strategies becomes 1/2 irrespective of payoff.
the probability of switching strategies becomes 1/2 irrespective of payoff.
Besides the fixed notion of β we define a dynamic value, equation (2), that adapts to the local noise of a node’s neighborhood; 1/β is proportional to noise56,57,58. The rationale for this is based on a social context where decision-making confidence is not fixed but relative to the situation or local environment that a player experiences. If I see a variety of payoffs in my neighborhood I am less confident with simply imitating the most successful since the environmental uncertainty adds doubt to the observed payoff values that may change. Conversely, I am more likely to take a risk if the environment is certain. We refer to this social rule as SOC in this work. In equation (2),  is the neighborhood variance of node o and
is the neighborhood variance of node o and  is the mean payoff of the same neighborhood.
is the mean payoff of the same neighborhood.

The SOC rule has the added advantage of reducing early imitation errors in our simulations since initial strategies  are picked from a uniform distribution for each run, a static β results in volatile early generations where it is common for nodes to cycle through several strategies before becoming satisfied.
are picked from a uniform distribution for each run, a static β results in volatile early generations where it is common for nodes to cycle through several strategies before becoming satisfied.
The payoff  , equation (3) from17, for each node o is determined by averaging the sum of its repeated game payoff over all of its nearest neighbors. The number of neighbors (degree) of node o is ko, the repeated game discount factor w is drawn from a uniform distribution between 0.8 and 0.9 and the number of games
, equation (3) from17, for each node o is determined by averaging the sum of its repeated game payoff over all of its nearest neighbors. The number of neighbors (degree) of node o is ko, the repeated game discount factor w is drawn from a uniform distribution between 0.8 and 0.9 and the number of games  .
.

Our asynchronous updating scheme follows the coevolution method described by46 with the exception that we do not evolve structure but rather evolve the population to convergence before starting anew with a different network. In this scheme, one node is picked at random from the network and then one of its neighbors is chosen at random. Each of the chosen nodes (players) plays a repeated game versus their immediate neighbors and record their average payoff. Strategies of the chosen nodes update according to the Fermi function, equation (1). Updates continue to take place until convergence is achieved. We define convergence as  asynchronous updates with no strategy switching (
asynchronous updates with no strategy switching ( is the number of nodes in the network). If a run does not converge within
is the number of nodes in the network). If a run does not converge within  updates then the last 100 updates are averaged together; this occurs in approximately 10% of cases in the Snow Drift game but rarely in other games (the problem with Snow Drift convergence was also reported by46). We also averaged 1000 updates for non-converging cases and found no significant difference compared to an average of 100 updates.
updates then the last 100 updates are averaged together; this occurs in approximately 10% of cases in the Snow Drift game but rarely in other games (the problem with Snow Drift convergence was also reported by46). We also averaged 1000 updates for non-converging cases and found no significant difference compared to an average of 100 updates.
All runs were executed on a 256-core, CrayXE6 computer. In total, 1024 runs were collected for each network structure, game and β value in order to sample the ST payoff plane sufficiently. More samples were needed to improve the clarity of the ST-planes in Fig. 1, thus a total of 10240 samples were collected, per game, for the full, BA and clique graphs. For the payoff matrix, similar to17, we adopt a simplified symmetric two-player repeated game, shown in equation (4) from2,59, where the defecting player receives T (temptation to defect) if their opponent cooperates but receives P = 0 if their opponent defects, (the punishment for mutual defection). Here we extend the PD game  and
and  from17 to the Stag Hunt
from17 to the Stag Hunt  and
and  , Snow Drift
, Snow Drift  and
and  and Harmony games
and Harmony games  and
and  . For each run, new T and S values are drawn from a uniform distribution within each respective game region.
. For each run, new T and S values are drawn from a uniform distribution within each respective game region.

The modularity index was calculated based on community structure measurement Q developed by32, where Q = 1 is a maximum, or the strongest community structure. Community detection was done by a random-walk60 method.
Additional Information
How to cite this article: Gianetto, D. A. and Heydari, B. Sparse cliques trump scale-free networks in coordination and competition. Sci. Rep. 6, 21870; doi: 10.1038/srep21870 (2016).
References
Axelrod, R. Launching “the evolution of cooperation”. J. Theor. Biol. 299, 21–4 (2012).
Nowak, M. A. Evolving cooperation. J. Theor. Biol. 299, 1–8 (2012).
Perc, M. & Szolnoki, A. Coevolutionary games-a mini review. Biosystems 99, 109–125 (2010).
Kogut, B. & Zander, U. What firms do? coordination, identity and learning. Organ. Sci. 7, 502–518 (1996).
Guastello, S. J. & Bond Jr, R. W. Coordination learning in stag hunt games with application to emergency management. Nonlinear Dynamics Psychol Life Sci 8, 345–74 (2004).
Tomasello, M. & Vaish, A. Origins of human cooperation and morality. Annu Rev Psychol 64, 231–55 (2013).
Sanfey, A. G. Social decision-making: insights from game theory and neuroscience. Science 318, 598–602 (2007).
Rilling, J. K. & Sanfey, A. G. The neuroscience of social decision-making. Annu Rev Psychol 62, 23–48 (2011).
Maynard Smith, J. Evolution and the theory of games (Cambridge University Press, Cambridge; New York, 1982).
Axelrod, R. & Hamilton, W. D. The evolution of cooperation. Science 211, 1390–1396 (1981).
Nowak, M. A. Five rules for the evolution of cooperation. Science 314, 1560–3 (2006).
Szabó, G. & Fáth, G. Evolutionary games on graphs. Phys. Rep. 446, 97–216 (2007).
Taylor, P. D. Altruism in viscous populations an inclusive fitness model. Evol. Ecol. 6, 352–356 (1992).
Hamilton, W. D. The genetical evolution of social behaviour. i. J. Theor. Biol. 7, 1–16 (1964).
Ohtsuki, H., Hauert, C., Lieberman, E. & Nowak, M. A. A simple rule for the evolution of cooperation on graphs and social networks. Nature 441, 502–505 (2006).
Maciejewski, W., Fu, F. & Hauert, C. Evolutionary game dynamics in populations with heterogenous structures. PLoS Comput. Biol. 10, e1003567 (2014).
Gianetto, D. A. & Heydari, B. Network modularity is essential for evolution of cooperation under uncertainty. Sci Rep 5, 9340 (2015).
Santos, F. C. & Pacheco, J. Scale-free networks provide a unifying framework for the emergence of cooperation. PRL 95, 098104 (2005).
Vukov, J., Santos, F. C. & Pacheco, J. M. Cognitive strategies take advantage of the cooperative potential of heterogeneous networks. New J. Phys. 14, 063031 (2012).
Santos, F. C., Pinheiro, F. L., Lenaerts, T. & Pacheco, J. M. The role of diversity in the evolution of cooperation. J. Theor. Biol. 299, 88–96 (2012).
Szolnoki, A., Perc, M. & Danku, Z. Towards effective payoffs in the prisoners dilemma game on scale-free networks. Physica A 387, 2075–2082 (2008).
Tomassini, M., Pestelacci, E. & Luthi, L. Social dilemmas and cooperation in complex networks. Int. J. Mod. Phys. C 18, 1173–1185 (2007).
Ohtsuki, H. & Nowak, M. A. Evolutionary games on cycles. Proc Biol Sci 273, 2249–56 (2006).
Manshadi, V. H. & Saberi, A. Dynamics of prisoner’s dilemma and the evolution of cooperation on networks. In Proceedings of the 3rd Innovations in Theoretical Computer Science Conference, 227–235 (ACM, 2012).
Vukov, J., Szabó, G. & Szolnoki, A. Cooperation in the noisy case: Prisoner’s dilemma game on two types of regular random graphs. Phys Rev E Stat Nonlin Soft Matter Phys 73, 067103 (2006).
Helbing, D. & Johansson, A. Cooperation, norms and revolutions: a unified game-theoretical approach. PLoS One 5, e12530 (2010).
Ostrom, E. Collective action and the evolution of social norms. J. Econ. Perspect. 14, 137–158 (2000).
Liu, Y., Chen, X., Zhang, L., Tao, F. & Wang, L. Does migration cost influence cooperation among success-driven individuals? Chaos Soliton Fract 45, 1301–1308 (2012).
Hauert, C. & Doebeli, M. Spatial structure often inhibits the evolution of cooperation in the snowdrift game. Nature 428, 643–646 (2004).
Masuda, N. Participation costs dismiss the advantage of heterogeneous networks in evolution of cooperation. Proc. Biol. Sci. 274, 1815–21 (2007).
Hales, D. & Arteconi, S. Motifs in evolving cooperative networks look like protein structure networks. Netw. Heterog. Media 3, 239–249 (2008).
Clauset, A., Newman, M. E. J. & Moore, C. Finding community structure in very large networks. Phys Rev E Stat Nonlin Soft Matter Phys 70 doi: 10.1103/PhysRevE.70.066111 (2004).
Newman, M. E. Modularity and community structure in networks. P Natl Acad Sci 103, 8577–8582 (2006).
Newman, M. E. J. & Girvan, M. Finding and evaluating community structure in networks. Phys Rev E Stat Nonlin Soft Matter Phys 69, 026113 (2004).
Milo, R. et al. Network motifs: simple building blocks of complex networks. Science 298, 824–827 (2002).
Wernicke, S. Efficient detection of network motifs. EEE/ACM Trans. Comput. Biol. Bioinf. 3, 347–359 (2006).
Wernicke, S. & Rasche, F. Fanmod: a tool for fast network motif detection. Bioinformatics 22, 1152–3 (2006).
Wolf, D. M. & Arkin, A. P. Motifs, modules and games in bacteria. Curr Opin Microbiol 6, 125–134 (2003).
Kashtan, N. & Alon, U. Spontaneous evolution of modularity and network motifs. P Natl Acad Sci USA 102, 13773–13778 (2005).
Xie, W.-J. J., Li, M.-X. X., Jiang, Z.-Q. Q. & Zhou, W.-X. X. Triadic motifs in the dependence networks of virtual societies. Sci Rep 4, 5244 (2014).
Ghoneim, A., Abbass, H. & Barlow, M. Characterizing game dynamics in two-player strategy games using network motifs. IEEE Trans Syst Man Cybern B Cybern 38, 682–90 (2008).
Paulau, P. V., Feenders, C. & Blasius, B. Motif analysis in directed ordered networks and applications to food webs. Sci Rep 5, 11926 (2015).
Nowak, M. A. & Sigmund, K. Tit for tat in heterogeneous populations. Nature 355, 250–253 (1992).
Nowak, M. & Sigmund, K. The evolution of stochastic strategies in the prisoner’s dilemma. Acta Appl. Math. 20, 247–265 (1990).
Trajanovski, S., Kuipers, F. A., Martín-Hernández, J. & Van Mieghem, P. Generating graphs that approach a prescribed modularity. Comput Commun 36, 363–372 (2013).
Santos, F. C., Pacheco, J. M. & Lenaerts, T. Cooperation prevails when individuals adjust their social ties. PLoS Comput. Biol. 2, e140 (2006).
Albert, R. & Barabási, A. L. Statistical mechanics of complex networks. Rev. Mod. Phys. 74, 47–97 (2002).
Barabási, A.-L. L. & Albert, R. Emergence of scaling in random networks. Science 286, 509–512 (1999).
Szabó, G. & Hoke, C. Evolutionary prisoners dilemma game on a square lattice. Phys Rev E Stat Nonlin Soft Matter Phys 58, 69 (1998).
Gracia-Lázaro, C. et al. Heterogeneous networks do not promote cooperation when humans play a prisoner’s dilemma. Proc Natl Acad Sci USA 109, 12922–6 (2012).
Grim, P. The greater generosity of the spatialized prisoner?’s dilemma. J. Theor. Biol. 173, 353–359 (1995).
Engemann, D. A., Bzdok, D., Eickhoff, S. B., Vogeley, K. & Schilbach, L. Games people play-toward an enactive view of cooperation in social neuroscience. Front Hum Neurosci 6, 148 (2012).
Damore, J. A. & Gore, J. Understanding microbial cooperation. J. Theor. Biol. 299, 31–41 (2012).
Grujić, J. et al. A comparative analysis of spatial prisoner’s dilemma experiments: conditional cooperation and payoff irrelevance. Sci. Rep. 4, 4615 (2014).
Brauchli, K., Killingback, T. & Doebeli, M. Evolution of cooperation in spatially structured populations. J. Theor. Biol. 200, 405–417 (1999).
Szabó, G., Vukov, J. & Szolnoki, A. Phase diagrams for an evolutionary prisoners dilemma game on two-dimensional lattices. Phys Rev E Stat Nonlin Soft Matter Phys 72 (2005). doi: 10.1103/PhysRevE.72.047107.
Szabó, G. & Hauert, C. Evolutionary prisoners dilemma games with voluntary participation. Phys Rev E Stat Nonlin Soft Matter Phys 66 (2002). doi: 10.1103/PhysRevE.66.062903.
Ren, J., Wang, W.-X. & Qi, F. Randomness enhances cooperation: A resonance-type phenomenon in evolutionary games. Phys Rev E Stat Nonlin Soft Matter Phys 75 (2007). doi: 10.1103/PhysRevE.75.045101.
Nowak, M. A. & May, R. M. Evolutionary games and spatial chaos. Nature 359, 826–829 (1992).
Pons, P. & Latapy, M. Computing communities in large networks using random walks. J. Graph Algorithms Appl. 10, 191–218 (2006).
Acknowledgements
This material is in part based upon work supported by the National Science Foundation under Grant No. 1548521.
Author information
Authors and Affiliations
Contributions
Author D.G. created all figures and wrote the main manuscript. Author D.G. created the model, ran simulations and analyzed results, the work was supervised by BH who also helped refine the research questions. All authors reviewed the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Gianetto, D., Heydari, B. Sparse cliques trump scale-free networks in coordination and competition. Sci Rep 6, 21870 (2016). https://doi.org/10.1038/srep21870
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep21870
- Springer Nature Limited
This article is cited by
-
Fair Topologies: Community Structures and Network Hubs Drive Emergence of Fairness Norms
Scientific Reports (2017)
-
Predicting metabolic adaptation from networks of mutational paths
Nature Communications (2017)
-
Detecting the Collapse of Cooperation in Evolving Networks
Scientific Reports (2016)